Recent Advances in Origins of Life Research by Biophysicists in Japan


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Synthesis, Assembly, and Regulation of Primitive Biopolymers
3. Assembly and Co-Assembly of Primitive Compartments
4. Primitive Biopolymer Evolution
5. The Future of OoL Research: Incorporating Novel Biophysics Technologies and Methodologies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, E.; Morowitz, H.J. The Origin and Nature of Life on Earth: The Emergence of the Fourth Geosphere; Cambridge University Press: Cambridge, UK, 2016; ISBN 9781107121881. [Google Scholar]
- Scharf, C.; Virgo, N.; Cleaves, H.J., 2nd; Aono, M.; Aubert-Kato, N.; Aydinoglu, A.; Barahona, A.; Barge, L.M.; Benner, S.A.; Biehl, M.; et al. A Strategy for Origins of Life Research. Astrobiology 2015, 15, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, N.; Hogeweg, P.; Kaneko, K. Conceptualizing the origin of life in terms of evolution. Philos. Trans. A Math. Phys. Eng. Sci. 2017, 375, 20160346. [Google Scholar]
- Kvenvolden, K.A.; Lawless, J.G.; Ponnamperuma, C. Nonprotein amino acids in the murchison meteorite. Proc. Natl. Acad. Sci. USA 1971, 68, 486–490. [Google Scholar] [PubMed]
- Kvenvolden, K.; Lawless, J.; Pering, K.; Peterson, E.; Flores, J.; Ponnamperuma, C.; Kaplan, I.R.; Moore, C. Evidence for Extraterrestrial Amino-acids and Hydrocarbons in the Murchison Meteorite. Nature 1970, 228, 923–926. [Google Scholar] [PubMed]
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef]
- Miller, S.L. Production of Some Organic Compounds under Possible Primitive Earth Conditions 1. J. Am. Chem. Soc. 1955, 77, 2351–2361. [Google Scholar]
- Miller, S.L.; Urey, H.C. Organic Compound Synthes on the Primitive Eart: Several questions about the origin of life have been answered, but much remains to be studied. Science 1959, 130, 245–251. [Google Scholar] [CrossRef]
- Martins, Z.; Botta, O.; Fogel, M.L.; Sephton, M.A.; Glavin, D.P.; Watson, J.S.; Dworkin, J.P.; Schwartz, A.W.; Ehrenfreund, P. Extraterrestrial nucleobases in the Murchison meteorite. Earth Planet. Sci. Lett. 2008, 270, 130–136. [Google Scholar] [CrossRef]
- Parker, E.T.; Cleaves, H.J.; Dworkin, J.P.; Glavin, D.P.; Callahan, M.; Aubrey, A.; Lazcano, A.; Bada, J.L. Primordial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment. Proc. Natl. Acad. Sci. USA 2011, 108, 5526–5531. [Google Scholar]
- Johnson, A.P.; Cleaves, H.J.; Dworkin, J.P.; Glavin, D.P.; Lazcano, A.; Bada, J.L. The Miller volcanic spark discharge experiment. Science 2008, 322, 404. [Google Scholar] [CrossRef] [PubMed]
- Orgel, L.E. Molecular replication. Nature 1992, 358, 203–209. [Google Scholar] [PubMed]
- O’Flaherty, D.K.; Kamat, N.P.; Mirza, F.N.; Li, L.; Prywes, N.; Szostak, J.W. Copying of Mixed-Sequence RNA Templates inside Model Protocells. J. Am. Chem. Soc. 2018, 140, 5171–5178. [Google Scholar] [CrossRef] [PubMed]
- Mansy, S.S.; Szostak, J.W. Reconstructing the emergence of cellular life through the synthesis of model protocells. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Chandru, K.; Guttenberg, N.; Giri, C.; Hongo, Y.; Butch, C.; Mamajanov, I.; James Cleaves, H. Simple prebiotic synthesis of high diversity dynamic combinatorial polyester libraries. Commun. Chem. 2018, 1, 30. [Google Scholar] [CrossRef]
- Yu, S.-S.; Krishnamurthy, R.; Fernández, F.M.; Hud, N.V.; Schork, F.J.; Grover, M.A. Kinetics of prebiotic depsipeptide formation from the ester-amide exchange reaction. Phys. Chem. Chem. Phys. 2016, 18, 28441–28450. [Google Scholar]
- Thelen, A.E.; Nixon, C.A.; Chanover, N.J.; Molter, E.M.; Cordiner, M.A.; Achterberg, R.K.; Serigano, J.; Irwin, P.G.J.; Teanby, N.; Charnley, S.B. Spatial variations in Titan’s atmospheric temperature: ALMA and Cassini comparisons from 2012 to 2015. Icarus 2018, 307, 380–390. [Google Scholar] [CrossRef]
- Shock, E.L.; Canovas, P.; Yang, Z.; Boyer, G.; Johnson, K.; Robinson, K.; Fecteau, K.; Windman, T.; Cox, A. Thermodynamics of Organic Transformations in Hydrothermal Fluids. Rev. Mineral. Geochem. 2013, 76, 311–350. [Google Scholar] [CrossRef]
- Adamala, K.; Szostak, J.W. Competition between model protocells driven by an encapsulated catalyst. Nat. Chem. 2013, 5, 495–501. [Google Scholar]
- Frederix, P.W.J.M.; Scott, G.G.; Abul-Haija, Y.M.; Kalafatovic, D.; Pappas, C.G.; Javid, N.; Hunt, N.T.; Ulijn, R.V.; Tuttle, T. Exploring the sequence space for (tri-)peptide self-assembly to design and discover new hydrogels. Nat. Chem. 2015, 7, 30–37. [Google Scholar] [CrossRef]
- Watanabe-Nakayama, T.; Ono, K.; Itami, M.; Takahashi, R.; Teplow, D.B.; Yamada, M. High-speed atomic force microscopy reveals structural dynamics of amyloid β1-42 aggregates. Proc. Natl. Acad. Sci. USA 2016, 113, 5835–5840. [Google Scholar]
- Dani, A.; Huang, B.; Bergan, J.; Dulac, C.; Zhuang, X. Superresolution imaging of chemical synapses in the brain. Neuron 2010, 68, 843–856. [Google Scholar] [CrossRef]
- Renaud, J.-P.; Chari, A.; Ciferri, C.; Liu, W.-T.; Rémigy, H.-W.; Stark, H.; Wiesmann, C. Cryo-EM in drug discovery: Achievements, limitations and prospects. Nat. Rev. Drug Discov. 2018, 17, 471–492. [Google Scholar] [CrossRef]
- Yamada, A.; Yu, P.; Lin, W.; Okugawa, Y.; Boland, C.R.; Goel, A. A RNA-Sequencing approach for the identification of novel long non-coding RNA biomarkers in colorectal cancer. Sci. Rep. 2018, 8, 575. [Google Scholar] [CrossRef]
- Chen, H.; Shiroguchi, K.; Ge, H.; Xie, X.S. Genome-wide study of mRNA degradation and transcript elongation in Escherichia coli. Mol. Syst. Biol. 2015, 11, 781. [Google Scholar]
- Mizuno, C.M.; Guyomar, C.; Roux, S.; Lavigne, R.; Rodriguez-Valera, F.; Sullivan, M.B.; Gillet, R.; Forterre, P.; Krupovic, M. Numerous cultivated and uncultivated viruses encode ribosomal proteins. Nat. Commun. 2019, 10, 752. [Google Scholar] [CrossRef]
- Forterre, P. To be or not to be alive: How recent discoveries challenge the traditional definitions of viruses and life. Stud. Hist. Philos. Biol. Biomed. Sci. 2016, 59, 100–108. [Google Scholar] [CrossRef]
- Ohta, A.; Murakami, H.; Higashimura, E.; Suga, H. Synthesis of polyester by means of genetic code reprogramming. Chem. Biol. 2007, 14, 1315–1322. [Google Scholar]
- Kawahara-Kobayashi, A.; Hitotsuyanagi, M.; Amikura, K.; Kiga, D. Experimental evolution of a green fluorescent protein composed of 19 unique amino acids without tryptophan. Orig. Life Evol. Biosph. 2014, 44, 75–86. [Google Scholar]
- Kawahara-Kobayashi, A.; Masuda, A.; Araiso, Y.; Sakai, Y.; Kohda, A.; Uchiyama, M.; Asami, S.; Matsuda, T.; Ishitani, R.; Dohmae, N.; et al. Simplification of the genetic code: Restricted diversity of genetically encoded amino acids. Nucleic Acids Res. 2012, 40, 10576–10584. [Google Scholar] [CrossRef]
- Amikura, K.; Kiga, D. The number of amino acids in a genetic code. RSC Adv. 2013, 3, 12512. [Google Scholar]
- Amikura, K.; Sakai, Y.; Asami, S.; Kiga, D. Multiple amino acid-excluded genetic codes for protein engineering using multiple sets of tRNA variants. ACS Synth. Biol. 2014, 3, 140–144. [Google Scholar]
- Mukai, T.; Wakiyama, M.; Sakamoto, K.; Yokoyama, S. Genetic encoding of non-natural amino acids in Drosophila melanogaster Schneider 2 cells. Protein Sci. 2010, 19, 440–448. [Google Scholar]
- Harada, K.; Aoyama, S.; Matsugami, A.; Kumar, P.K.R.; Katahira, M.; Kato, N.; Ohkanda, J. RNA-directed amino acid coupling as a model reaction for primitive coded translation. Chembiochem 2014, 15, 794–798. [Google Scholar] [CrossRef]
- Froese, T.; Campos, J.I.; Fujishima, K.; Kiga, D.; Virgo, N. Horizontal transfer of code fragments between protocells can explain the origins of the genetic code without vertical descent. Sci. Rep. 2018, 8, 3532. [Google Scholar] [PubMed]
- Soai, K.; Matsumoto, A. Asymmetric Autocatalysis and the Origin of Homochirality. In Stereochemistry and Global Connectivity: The Legacy of Ernest L. Eliel Volume 2; Cheng, H.N., Maryanoff, C.A., Miller, B.D., Schmidt, D.G., Eds.; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2017; Volume 1258, pp. 27–47. ISBN 9780841232402. [Google Scholar]
- Wang, Z.; Xu, W.; Liu, L.; Zhu, T.F. A synthetic molecular system capable of mirror-image genetic replication and transcription. Nat. Chem. 2016, 8, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Sczepanski, J.T.; Joyce, G.F. A cross-chiral RNA polymerase ribozyme. Nature 2014, 515, 440–442. [Google Scholar]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; Duffy, C.D.; Sutherland, J.D. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar] [CrossRef]
- Xu, J.; Ritson, D.J.; Ranjan, S.; Todd, Z.R.; Sasselov, D.D.; Sutherland, J.D. Photochemical reductive homologation of hydrogen cyanide using sulfite and ferrocyanide. Chem. Commun. 2018, 54, 5566–5569. [Google Scholar]
- Shimamura, K.; Shimojo, F.; Nakano, A.; Tanaka, S. Ab initio molecular dynamics study of prebiotic production processes of organic compounds at meteorite impacts on ocean. J. Comput. Chem. 2019, 40, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, K.; Shimojo, F.; Nakano, A.; Tanaka, S. Meteorite impacts on ancient oceans opened up multiple NH production pathways. Phys. Chem. Chem. Phys. 2017, 19, 11655–11667. [Google Scholar] [CrossRef]
- Bada, J.L.; Chalmers, J.H.; Cleaves, H.J. Is formamide a geochemically plausible prebiotic solvent? Phys. Chem. Chem. Phys. 2016, 18, 20085–20090. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Botta, G.; Pino, S.; Costanzo, G.; Di Mauro, E. Genetics first or metabolism first? The formamide clue. Chem. Soc. Rev. 2012, 41, 5526–5565. [Google Scholar]
- Saladino, R.; Crestini, C.; Pino, S.; Costanzo, G.; Di Mauro, E. Formamide and the origin of life. Phys. Life Rev. 2012, 9, 84–104. [Google Scholar]
- Gillams, R.J.; Jia, T.Z. Mineral Surface-Templated Self-Assembling Systems: Case Studies from Nanoscience and Surface Science towards Origins of Life Research. Life 2018, 8, 10. [Google Scholar] [CrossRef]
- Ferris, J.P. Montmorillonite-catalysed formation of RNA oligomers: The possible role of catalysis in the origins of life. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1777–1786, discussion 1786. [Google Scholar] [CrossRef]
- Orgel, L.E. Polymerization on the rocks: Theoretical introduction. Orig. Life Evol. Biosph. 1998, 28, 227–234. [Google Scholar] [CrossRef]
- Fornaro, T.; Steele, A.; Brucato, J.R. Catalytic/Protective Properties of Martian Minerals and Implications for Possible Origin of Life on Mars. Life 2018, 8, 56. [Google Scholar]
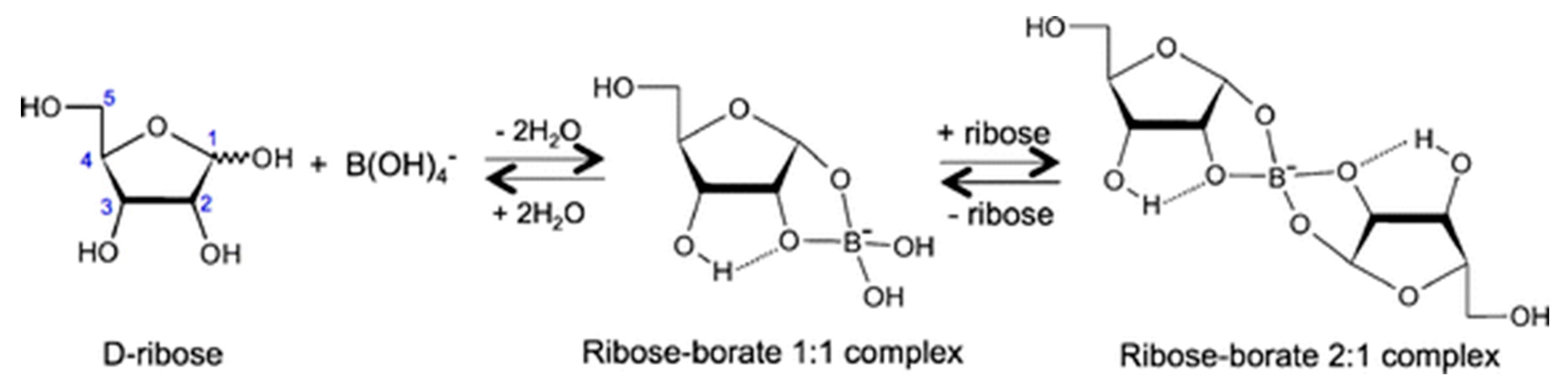
- Furukawa, Y.; Horiuchi, M.; Kakegawa, T. Selective stabilization of ribose by borate. Orig. Life Evol. Biosph. 2013, 43, 353–361. [Google Scholar]
- Furukawa, Y.; Kim, H.-J.; Hutter, D.; Benner, S.A. Abiotic regioselective phosphorylation of adenosine with borate in formamide. Astrobiology 2015, 15, 259–267. [Google Scholar] [CrossRef]
- Goedert, M.; Spillantini, M.G. A century of Alzheimer’s disease. Science 2006, 314, 777–781. [Google Scholar] [CrossRef]
- Watanabe-Nakayama, T.; Itami, M.; Kodera, N.; Ando, T.; Konno, H. High-speed atomic force microscopy reveals strongly polarized movement of clostridial collagenase along collagen fibrils. Sci. Rep. 2016, 6, 28975. [Google Scholar]
- Iida, Y.; Tamura, A. 3P076 Hydrolysis of lipid droplets and amyloid fibrils by the designed peptide (01D. Protein: Function, Poster). Biophysics 2013, 53, S224. [Google Scholar] [CrossRef]
- Iida, Y.; Tamura, A. 1P071 Hydrolysis of amyloid fibrils by artificially designed peptides (01D. Protein: Function, Poster, The 52nd Annual Meeting of the Biophysical Society of Japan(BSJ2014)). Biophysics 2014, 54, S152. [Google Scholar]
- Nishio, T.; Yoshikawa, Y.; Fukuda, W.; Umezawa, N.; Higuchi, T.; Fujiwara, S.; Imanaka, T.; Yoshikawa, K. Branched-Chain Polyamine Found in Hyperthermophiles Induces Unique Temperature-Dependent Structural Changes in Genome-Size DNA. Chemphyschem 2018, 19, 2299–2304. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Fujita, Y.; Ohno, H.; Suzuki, Y.; Hayashi, K.; Komatsu, K.R.; Kawasaki, S.; Hidaka, K.; Yonehara, S.; Sugiyama, H.; et al. Protein-driven RNA nanostructured devices that function in vitro and control mammalian cell fate. Nat. Commun. 2017, 8, 540. [Google Scholar] [PubMed]
- Ohno, H.; Saito, H. RNA/RNP Nanotechnology for Biological Applications. Biophysics 2016, 56, 023–026. [Google Scholar]
- Ohno, H.; Osada, E.; Saito, H. Design, assembly, and evaluation of RNA-protein nanostructures. Methods Mol. Biol. 2015, 1297, 197–211. [Google Scholar]
- Ohno, H.; Saito, H. RNA and RNP as Building Blocks for Nanotechnology and Synthetic Biology. Prog. Mol. Biol. Transl. Sci. 2016, 139, 165–185. [Google Scholar]
- Rahman, M.M.; Matsumura, S.; Ikawa, Y. Effects of molecular crowding on a bimolecular group I ribozyme and its derivative that self-assembles to form ribozyme oligomers. Biochem. Biophys. Res. Commun. 2018, 507, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Gulshan, M.A.; Matsumura, S.; Higuchi, T.; Umezawa, N.; Ikawa, Y. Comparative study of polyethylene polyamines as activator molecules for a structurally unstable group I ribozyme. Biosci. Biotechnol. Biochem. 2018, 82, 1404–1407. [Google Scholar]
- Rahman, M.M.; Matsumura, S.; Ikawa, Y. Artificial RNA Motifs Expand the Programmable Assembly between RNA Modules of a Bimolecular Ribozyme Leading to Application to RNA Nanostructure Design. Biology 2017, 6, 37. [Google Scholar]
- Berg, J.M.; Stryer, L.; Tymoczko, J.L.; Gatto, G.J. Biochemistry; Macmillan Learning: New York, NY, USA, 2015; ISBN 9781464126109. [Google Scholar]
- Monnard, P.-A.; Walde, P. Current Ideas about Prebiological Compartmentalization. Life 2015, 5, 1239–1263. [Google Scholar] [PubMed]
- Sunami, T.; Ichihashi, N.; Nishikawa, T.; Kazuta, Y.; Yomo, T. Effect of Liposome Size on Internal RNA Replication Coupled with Replicase Translation. Chembiochem 2016, 17, 1282–1289. [Google Scholar] [PubMed]
- Szostak, J.W.; Bartel, D.P.; Luisi, P.L. Synthesizing life. Nature 2001, 409, 387–390. [Google Scholar]
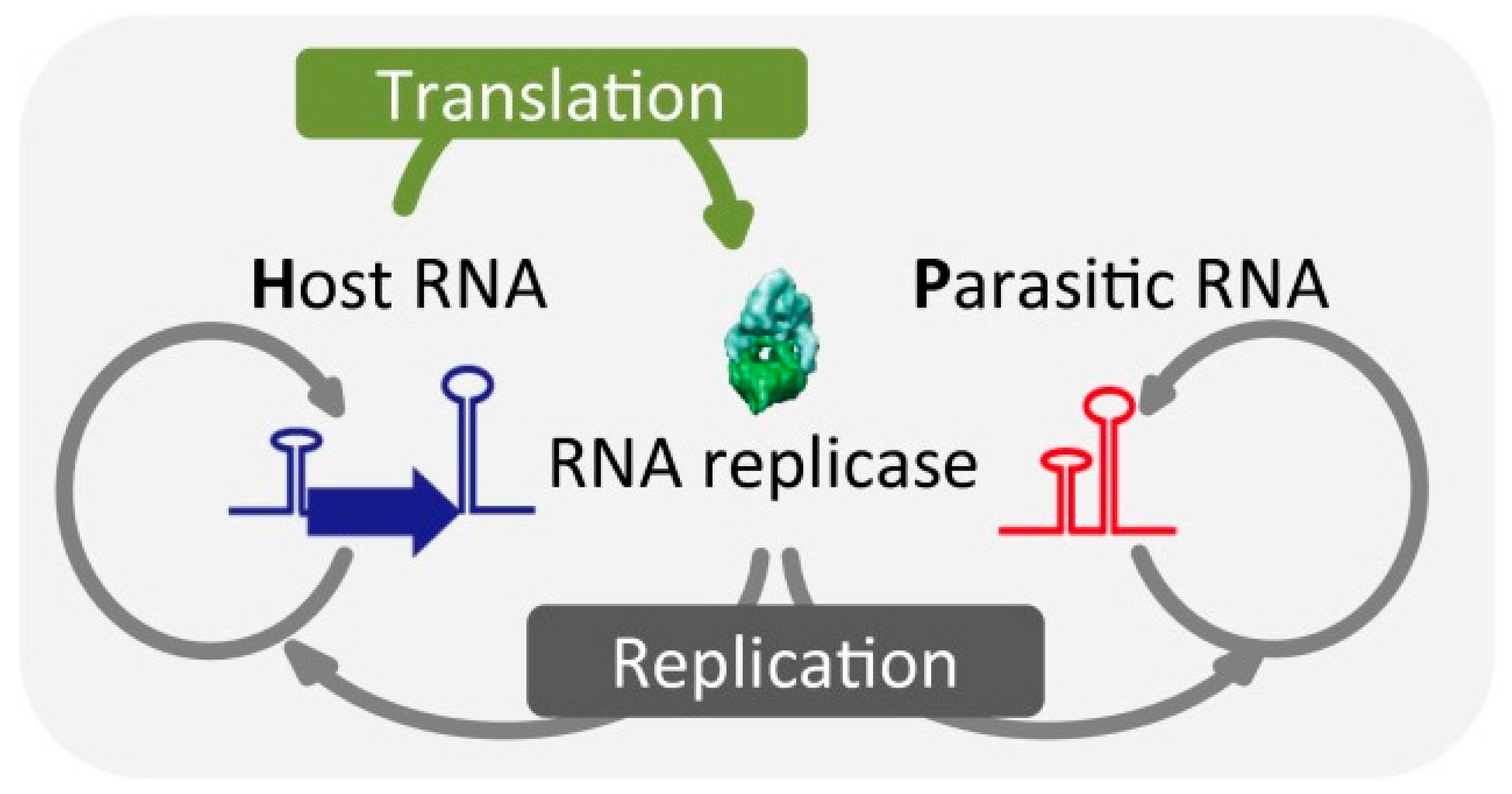
- Bansho, Y.; Furubayashi, T.; Ichihashi, N.; Yomo, T. Host-parasite oscillation dynamics and evolution in a compartmentalized RNA replication system. Proc. Natl. Acad. Sci. USA 2016, 113, 4045–4050. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, S.; Kun, Á.; Ryckelynck, M.; Coldren, F.; Szilágyi, A.; Jossinet, F.; Rick, C.; Nghe, P.; Szathmáry, E.; Griffiths, A.D. Transient compartmentalization of RNA replicators prevents extinction due to parasites. Science 2016, 354, 1293–1296. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.A.; Walde, P. From self-assembled vesicles to protocells. Cold Spring Harb. Perspect. Biol. 2010, 2, a002170. [Google Scholar]
- Deamer, D. The Role of Lipid Membranes in Life’s Origin. Life 2017, 7, 5. [Google Scholar] [CrossRef]
- Kamiya, K.; Kawano, R.; Osaki, T.; Akiyoshi, K.; Takeuchi, S. Cell-sized asymmetric lipid vesicles facilitate the investigation of asymmetric membranes. Nat. Chem. 2016, 8, 881–889. [Google Scholar]
- Watanabe, R.; Soga, N.; Yamanaka, T.; Noji, H. High-throughput formation of lipid bilayer membrane arrays with an asymmetric lipid composition. Sci. Rep. 2014, 4, 7076. [Google Scholar] [CrossRef]
- Tsuda, S.; Sakakura, T.; Fujii, S.; Suzuki, H.; Yomo, T. Shape Transformations of Lipid Vesicles by Insertion of Bulky-Head Lipids. PLoS ONE 2015, 10, e0132963. [Google Scholar]
- Shima, T.; Muraoka, T.; Hamada, T.; Morita, M.; Takagi, M.; Fukuoka, H.; Inoue, Y.; Sagawa, T.; Ishijima, A.; Omata, Y.; et al. Micrometer-size vesicle formation triggered by UV light. Langmuir 2014, 30, 7289–7295. [Google Scholar] [CrossRef]
- Ikeda, K.; Horiuchi, A.; Egawa, A.; Tamaki, H.; Fujiwara, T.; Nakano, M. Nanodisc-to-Nanofiber Transition of Noncovalent Peptide–Phospholipid Assemblies. ACS Omega 2017, 2, 2935–2944. [Google Scholar]
- Sheng, L.; Kurihara, K. Transformation of oil droplets into giant vesicles. Chem. Commun. 2016, 52, 7786–7789. [Google Scholar]
- Hao, F.; Tahara, K.; Kikuchi, J.-I. A Synthetic Cell Division System: Effect of Nonbilayer-forming Lipid on Division of Liposomal Membranes. Chem. Lett. 2014, 43, 811–813. [Google Scholar]
- Sakuma, Y.; Imai, M. From vesicles to protocells: The roles of amphiphilic molecules. Life 2015, 5, 651–675. [Google Scholar]
- Williams, G.H.; Stoeber, K. The cell cycle and cancer. J. Pathol. 2012, 226, 352–364. [Google Scholar]
- Becker, S.; Schneider, C.; Okamura, H.; Crisp, A.; Amatov, T.; Dejmek, M.; Carell, T. Wet-dry cycles enable the parallel origin of canonical and non-canonical nucleosides by continuous synthesis. Nat. Commun. 2018, 9, 163. [Google Scholar]
- Fox, S.; Pleyer, H.L.; Strasdeit, H. An automated apparatus for the simulation of prebiotic wet‒dry cycles under strictly anaerobic conditions. Int. J. Astrobiol. 2019, 18, 60–72. [Google Scholar] [CrossRef]
- Milshteyn, D.; Damer, B.; Havig, J.; Deamer, D. Amphiphilic Compounds Assemble into Membranous Vesicles in Hydrothermal Hot Spring Water but Not in Seawater. Life 2018, 8, 11. [Google Scholar]
- Hurwitz, S.; Harris, R.N.; Werner, C.A.; Murphy, F. Heat flow in vapor dominated areas of the Yellowstone Plateau Volcanic Field: Implications for the thermal budget of the Yellowstone Caldera: Heat flow in the yellowstone caldera. J. Geophys. Res. 2012, 117, 425. [Google Scholar] [CrossRef]
- Terasawa, H.; Nishimura, K.; Suzuki, H.; Matsuura, T.; Yomo, T. Coupling of the fusion and budding of giant phospholipid vesicles containing macromolecules. Proc. Natl. Acad. Sci. USA 2012, 109, 5942–5947. [Google Scholar] [CrossRef]
- Kurihara, K.; Okura, Y.; Matsuo, M.; Toyota, T.; Suzuki, K.; Sugawara, T. A recursive vesicle-based model protocell with a primitive model cell cycle. Nat. Commun. 2015, 6, 8352. [Google Scholar] [CrossRef]
- Kurihara, K.; Tamura, M.; Shohda, K.-I.; Toyota, T.; Suzuki, K.; Sugawara, T. Self-reproduction of supramolecular giant vesicles combined with the amplification of encapsulated DNA. Nat. Chem. 2011, 3, 775–781. [Google Scholar] [CrossRef]
- Rampioni, G.; D’Angelo, F.; Messina, M.; Zennaro, A.; Kuruma, Y.; Tofani, D.; Leoni, L.; Stano, P. Synthetic cells produce a quorum sensing chemical signal perceived by Pseudomonas aeruginosa. Chem. Commun. 2018, 54, 2090–2093. [Google Scholar] [CrossRef]
- Kuruma, Y. Creation of Simple Biochemical Systems to Study Early Cellular Life. Orig. Life Evol. Biosph. 2015, 45, 359–360. [Google Scholar] [CrossRef]
- Berhanu, S.; Ueda, T.; Kuruma, Y. Artificial photosynthetic cell producing energy for protein synthesis. Nat. Commun. 2019, 10, 1325. [Google Scholar] [CrossRef]
- Shimobayashi, S.F.; Ichikawa, M. Emergence of DNA-encapsulating liposomes from a DNA-lipid blend film. J. Phys. Chem. B 2014, 118, 10688–10694. [Google Scholar] [CrossRef]
- Kurokawa, C.; Fujiwara, K.; Morita, M.; Kawamata, I.; Kawagishi, Y.; Sakai, A.; Murayama, Y.; Nomura, S.-I.M.; Murata, S.; Takinoue, M.; et al. DNA cytoskeleton for stabilizing artificial cells. Proc. Natl. Acad. Sci. USA 2017, 114, 7228–7233. [Google Scholar] [CrossRef]
- Allolio, C.; Magarkar, A.; Jurkiewicz, P.; Baxová, K.; Javanainen, M.; Mason, P.E.; Šachl, R.; Cebecauer, M.; Hof, M.; Horinek, D.; et al. Arginine-rich cell-penetrating peptides induce membrane multilamellarity and subsequently enter via formation of a fusion pore. Proc. Natl. Acad. Sci. USA 2018, 115, 11923–11928. [Google Scholar] [CrossRef]
- Milletti, F. Cell-penetrating peptides: Classes, origin, and current landscape. Drug Discov. Today 2012, 17, 850–860. [Google Scholar] [CrossRef]
- Mahendran, K.R.; Niitsu, A.; Kong, L.; Thomson, A.R.; Sessions, R.B.; Woolfson, D.N.; Bayley, H. A monodisperse transmembrane α-helical peptide barrel. Nat. Chem. 2017, 9, 411–419. [Google Scholar] [CrossRef]
- Islam, M.Z.; Sharmin, S.; Moniruzzaman, M.; Yamazaki, M. Elementary processes for the entry of cell-penetrating peptides into lipid bilayer vesicles and bacterial cells. Appl. Microbiol. Biotechnol. 2018, 102, 3879–3892. [Google Scholar]
- Parvez, F.; Alam, J.M.; Dohra, H.; Yamazaki, M. Elementary processes of antimicrobial peptide PGLa-induced pore formation in lipid bilayers. Biochim. Biophys. Acta Biomembr. 2018, 1860, 2262–2271. [Google Scholar] [CrossRef]
- Islam, M.Z.; Sharmin, S.; Levadnyy, V.; Alam Shibly, S.U.; Yamazaki, M. Effects of Mechanical Properties of Lipid Bilayers on the Entry of Cell-Penetrating Peptides into Single Vesicles. Langmuir 2017, 33, 2433–2443. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Islam, M.Z.; Sharmin, S.; Dohra, H.; Yamazaki, M. Entry of a Six-Residue Antimicrobial Peptide Derived from Lactoferricin B into Single Vesicles and Escherichia coli Cells without Damaging their Membranes. Biochemistry 2017, 56, 4419–4431. [Google Scholar] [CrossRef]
- Nakao, H.; Hayashi, C.; Ikeda, K.; Saito, H.; Nagao, H.; Nakano, M. Effects of Hydrophilic Residues and Hydrophobic Length on Flip-Flop Promotion by Transmembrane Peptides. J. Phys. Chem. B 2018, 122, 4318–4324. [Google Scholar] [CrossRef]
- Suzuki, Y.; Endo, M.; Sugiyama, H. Lipid-bilayer-assisted two-dimensional self-assembly of DNA origami nanostructures. Nat. Commun. 2015, 6, 8052. [Google Scholar]
- Sato, Y.; Endo, M.; Morita, M.; Takinoue, M.; Sugiyama, H.; Murata, S.; Nomura, S.-I.M.; Suzuki, Y. Environment-Dependent Self-Assembly of DNA Origami Lattices on Phase-Separated Lipid Membranes. Adv. Mater. Interfaces 2018, 5, 1800437. [Google Scholar] [CrossRef]
- Sugiura, Y.; Ikeda, K.; Nakano, M. High Membrane Curvature Enhances Binding, Conformational Changes, and Fibrillation of Amyloid-β on Lipid Bilayer Surfaces. Langmuir 2015, 31, 11549–11557. [Google Scholar] [CrossRef]
- Vogele, K.; Frank, T.; Gasser, L.; Goetzfried, M.A.; Hackl, M.W.; Sieber, S.A.; Simmel, F.C.; Pirzer, T. Towards synthetic cells using peptide-based reaction compartments. Nat. Commun. 2018, 9, 3862. [Google Scholar] [CrossRef]
- Cooper, G.J.T.; Kitson, P.J.; Winter, R.; Zagnoni, M.; Long, D.-L.; Cronin, L. Modular redox-active inorganic chemical cells: iCHELLs. Angew. Chem. Int. Ed. Engl. 2011, 50, 10373–10376. [Google Scholar] [CrossRef]
- Yin, Y.; Niu, L.; Zhu, X.; Zhao, M.; Zhang, Z.; Mann, S.; Liang, D. Non-equilibrium behaviour in coacervate-based protocells under electric-field-induced excitation. Nat. Commun. 2016, 7, 10658. [Google Scholar] [CrossRef] [PubMed]
- Frankel, E.A.; Bevilacqua, P.C.; Keating, C.D. Polyamine/Nucleotide Coacervates Provide Strong Compartmentalization of Mg2+, Nucleotides, and RNA. Langmuir 2016, 32, 2041–2049. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.Z.; Hentrich, C.; Szostak, J.W. Rapid RNA exchange in aqueous two-phase system and coacervate droplets. Orig. Life Evol. Biosph. 2014, 44, 1–12. [Google Scholar] [CrossRef]
- Nakatani, N.; Sakuta, H.; Hayashi, M.; Tanaka, S.; Takiguchi, K.; Tsumoto, K.; Yoshikawa, K. Specific Spatial Localization of Actin and DNA in a Water/Water Microdroplet: Self-Emergence of a Cell-Like Structure. Chembiochem 2018, 19, 1370–1374. [Google Scholar] [CrossRef] [PubMed]
- Pir Cakmak, F.; Keating, C.D. Combining Catalytic Microparticles with Droplets Formed by Phase Coexistence: Adsorption and Activity of Natural Clays at the Aqueous/Aqueous Interface. Sci. Rep. 2017, 7, 3215. [Google Scholar] [CrossRef]
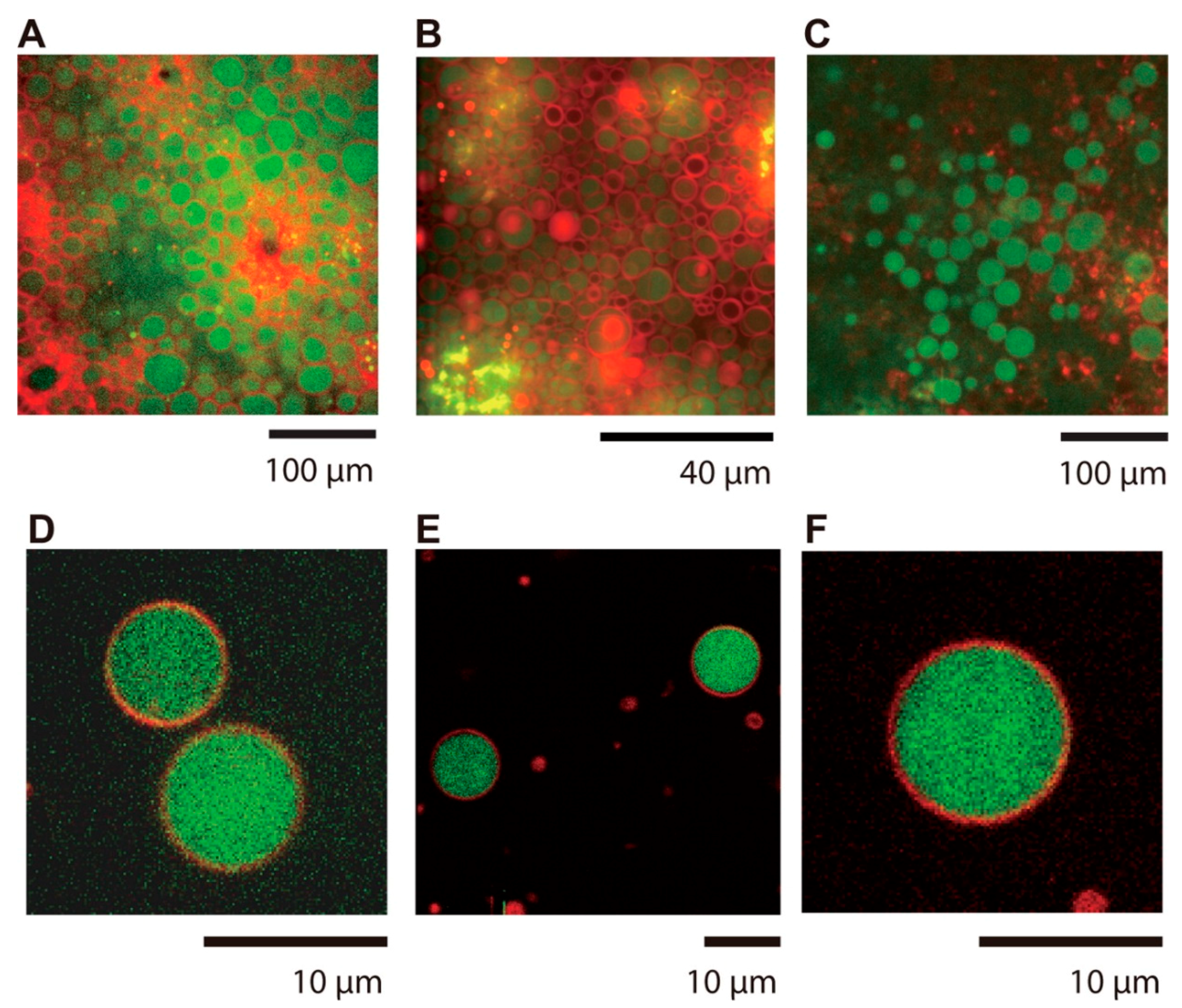
- Jia, T.Z.; Pappas, C.G.; Jain, A.; Kroiss, D.; Wijerathne, N.K.; Aramini, J.M.; Pal, A.; Szostak, J.W.; Ulijn, R.V.; Chandru, K.; et al. Biopolymer Self-Assembly and Combinatorial Evolution at the Origin of Life. In Proceedings of the 56th Annual Meeting of the Biophysical Society of Japan, Okayama, Japan, 15–17 September 2018. [Google Scholar]
- Robertson, M.P.; Joyce, G.F. The origins of the RNA world. Cold Spring Harb. Perspect. Biol. 2012, 4, a003608. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Nemoto, N.; Kumachi, S.; Arai, H. In Vitro Selection of Single-Domain Antibody (VHH) Using cDNA Display. Methods Mol. Biol. 2018, 1827, 269–285. [Google Scholar]
- Takahashi, K.; Sunohara, M.; Terai, T.; Kumachi, S.; Nemoto, N. Enhanced mRNA-protein fusion efficiency of a single-domain antibody by selection of mRNA display with additional random sequences in the terminal translated regions. Biophys. Physicobiol. 2017, 14, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Tada, S.; Zang, Q.; Wang, W.; Kawamoto, M.; Liu, M.; Iwashita, M.; Uzawa, T.; Kiga, D.; Yamamura, M.; Ito, Y. In vitro selection of a photoresponsive peptide aptamer to glutathione-immobilized microbeads. J. Biosci. Bioeng. 2015, 119, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, Y.; Kumachi, S.; Nishigaki, K.; Nemoto, N. Increasing the library size in cDNA display by optimizing purification procedures. Biol. Proced. Online 2013, 15, 7. [Google Scholar] [CrossRef]
- Mast, C.B.; Schink, S.; Gerland, U.; Braun, D. Escalation of polymerization in a thermal gradient. Proc. Natl. Acad. Sci. USA 2013, 110, 8030–8035. [Google Scholar] [CrossRef]
- Maeda, Y.T.; Tlusty, T.; Libchaber, A. Effects of long DNA folding and small RNA stem-loop in thermophoresis. Proc. Natl. Acad. Sci. USA 2012, 109, 17972–17977. [Google Scholar] [CrossRef]
- Svoboda, P.; Di Cara, A. Hairpin RNA: A secondary structure of primary importance. Cell. Mol. Life Sci. 2006, 63, 901–908. [Google Scholar] [CrossRef]
- Mizuuchi, R.; Blokhuis, A.; Vincent, L.; Nghe, P.; Lehman, N.; Baum, D. Mineral surfaces select for longer RNA molecules. Chem. Commun. 2019, 55, 2090–2093. [Google Scholar] [CrossRef]
- Mizuuchi, R.; Lehman, N. Limited Sequence Diversity Within a Population Supports Prebiotic RNA Reproduction. Life 2019, 9, 20. [Google Scholar] [CrossRef]
- Koonin, E.V.; Wolf, Y.I.; Katsnelson, M.I. Inevitability of the emergence and persistence of genetic parasites caused by evolutionary instability of parasite-free states. Biol. Direct 2017, 12, 31. [Google Scholar] [CrossRef]
- Ichihashi, N.; Usui, K.; Kazuta, Y.; Sunami, T.; Matsuura, T.; Yomo, T. Darwinian evolution in a translation-coupled RNA replication system within a cell-like compartment. Nat. Commun. 2013, 4, 2494. [Google Scholar] [CrossRef]
- Mizuuchi, R.; Ichihashi, N. Sustainable replication and coevolution of cooperative RNAs in an artificial cell-like system. Nat. Ecol. Evol. 2018, 2, 1654–1660. [Google Scholar] [CrossRef]
- Wilson, D.S.; Keefe, A.D.; Szostak, J.W. The use of mRNA display to select high-affinity protein-binding peptides. Proc. Natl. Acad. Sci. USA 2001, 98, 3750–3755. [Google Scholar] [CrossRef]
- Smith, G. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef]
- He, Y.; Wang, K.; Yan, N. The recombinant expression systems for structure determination of eukaryotic membrane proteins. Protein Cell 2014, 5, 658–672. [Google Scholar] [CrossRef]
- Fujii, S.; Matsuura, T.; Sunami, T.; Nishikawa, T.; Kazuta, Y.; Yomo, T. Liposome display for in vitro selection and evolution of membrane proteins. Nat. Protoc. 2014, 9, 1578–1591. [Google Scholar] [CrossRef]
- Fujii, S.; Matsuura, T.; Yomo, T. In vitro directed evolution of alpha-hemolysin by liposome display. Biophysics 2015, 11, 67–72. [Google Scholar] [CrossRef][Green Version]
- Uyeda, A.; Watanabe, T.; Kato, Y.; Watanabe, H.; Yomo, T.; Hohsaka, T.; Matsuura, T. Liposome-Based in Vitro Evolution of Aminoacyl-tRNA Synthetase for Enhanced Pyrrolysine Derivative Incorporation. Chembiochem 2015, 16, 1797–1802. [Google Scholar] [CrossRef]
- Suzuki, K.; Ikegami, T. Spatial-pattern-induced evolution of a self-replicating loop network. Artif. Life 2006, 12, 461–485. [Google Scholar] [CrossRef]
- Virgo, N.; Froese, T.; Ikegami, T. The positive role of parasites in the origins of life. In 2013 IEEE Symposium on Artificial Life (ALife); IEEE: Piscataway, NJ, USA, 2013. [Google Scholar]
- Matsubara, Y.J.; Kaneko, K. Optimal size for emergence of self-replicating polymer system. Phys. Rev. E 2016, 93, 032503. [Google Scholar] [CrossRef]
- Kamimura, A.; Kaneko, K. Reproduction of a protocell by replication of a minority molecule in a catalytic reaction network. Phys. Rev. Lett. 2010, 105, 268103. [Google Scholar] [CrossRef]
- Ueda, M.; Takeuchi, N.; Kaneko, K. Stronger selection can slow down evolution driven by recombination on a smooth fitness landscape. PLoS ONE 2017, 12, e0183120. [Google Scholar] [CrossRef]
- Kamino, K.; Kondo, Y.; Nakajima, A.; Honda-Kitahara, M.; Kaneko, K.; Sawai, S. Fold-change detection and scale invariance of cell-cell signaling in social amoeba. Proc. Natl. Acad. Sci. USA 2017, 114, E4149–E4157. [Google Scholar] [CrossRef]
- Saito, N.; Kaneko, K. Embedding dual function into molecular motors through collective motion. Sci. Rep. 2017, 7, 44288. [Google Scholar] [CrossRef]
- Witkowski, O.; Ikegami, T. Emergence of Swarming Behavior: Foraging Agents Evolve Collective Motion Based on Signaling. PLoS ONE 2016, 11, e0152756. [Google Scholar] [CrossRef]
- Maruyama, N.; Saito, D.; Hashimoto, Y.; Ikegami, T. Dynamic organization of flocking behaviors in a large-scale boids model. SIAM J. Sci. Comput. 2019, 375, 20160351. [Google Scholar] [CrossRef]
- Kojima, H.; Froese, T.; Oka, M.; Iizuka, H.; Ikegami, T. A Sensorimotor Signature of the Transition to Conscious Social Perception: Co-regulation of Active and Passive Touch. Front. Psychol. 2017, 8, 1778. [Google Scholar]
- Ikegami, T. Simulating Active Perception and Mental Imagery with Embodied Chaotic Itinerancy. J. Conscious. Stud. 2007, 14, 111–125. [Google Scholar]
- Sinapayen, L.; Masumori, A.; Ikegami, T. Learning by stimulation avoidance: A principle to control spiking neural networks dynamics. PLoS ONE 2017, 12, e0170388. [Google Scholar]
- Woodward, A.; Froese, T.; Ikegami, T. Neural coordination can be enhanced by occasional interruption of normal firing patterns: A self-optimizing spiking neural network model. Neural Netw. 2015, 62, 39–46. [Google Scholar] [CrossRef][Green Version]
- Oshima, T. Enigmas of biosyntheses of unusual polyamines in an extreme thermophile, Thermus thermophilus. Plant Physiol. Biochem. 2010, 48, 521–526. [Google Scholar] [CrossRef]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Shendure, J.; Balasubramanian, S.; Church, G.M.; Gilbert, W.; Rogers, J.; Schloss, J.A.; Waterston, R.H. DNA sequencing at 40: Past, present and future. Nature 2017, 550, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Ameta, S.; Winz, M.-L.; Previti, C.; Jäschke, A. Next-generation sequencing reveals how RNA catalysts evolve from random space. Nucleic Acids Res. 2014, 42, 1303–1310. [Google Scholar] [CrossRef]
- Pitt, J.N.; Ferré-D’Amaré, A.R. Rapid construction of empirical RNA fitness landscapes. Science 2010, 330, 376–379. [Google Scholar] [CrossRef]
- Ogawa, T.; Kryukov, K.; Imanishi, T.; Shiroguchi, K. The efficacy and further functional advantages of random-base molecular barcodes for absolute and digital quantification of nucleic acid molecules. Sci. Rep. 2017, 7, 13576. [Google Scholar] [CrossRef] [PubMed]
- Shiroguchi, K.; Jia, T.Z.; Sims, P.A.; Xie, X.S. Digital RNA sequencing minimizes sequence-dependent bias and amplification noise with optimized single-molecule barcodes. Proc. Natl. Acad. Sci. USA 2012, 109, 1347–1352. [Google Scholar] [CrossRef]
- Ohshiro, T.; Tsutsui, M.; Yokota, K.; Taniguchi, M. Quantitative analysis of DNA with single-molecule sequencing. Sci. Rep. 2018, 8, 8517. [Google Scholar] [CrossRef]
- Ohshiro, T.; Tsutsui, M.; Yokota, K.; Furuhashi, M.; Taniguchi, M.; Kawai, T. Detection of post-translational modifications in single peptides using electron tunnelling currents. Nat. Nanotechnol. 2014, 9, 835–840. [Google Scholar] [CrossRef]
- Ando, T.; Kodera, N.; Takai, E.; Maruyama, D.; Saito, K.; Toda, A. A high-speed atomic force microscope for studying biological macromolecules. Proc. Natl. Acad. Sci. USA 2001, 98, 12468–12472. [Google Scholar] [CrossRef]
- Ando, T.; Uchihashi, T.; Kodera, N.; Yamamoto, D.; Miyagi, A.; Taniguchi, M.; Yamashita, H. High-speed AFM and nano-visualization of biomolecular processes. Pflugers Arch. 2008, 456, 211–225. [Google Scholar]
- Hansma, H.G. Possible origin of life between mica sheets: Does life imitate mica? J. Biomol. Struct. Dyn. 2013, 31, 888–895. [Google Scholar] [CrossRef]
- Han, D.; Qi, X.; Myhrvold, C.; Wang, B.; Dai, M.; Jiang, S.; Bates, M.; Liu, Y.; An, B.; Zhang, F.; et al. Single-stranded DNA and RNA origami. Science 2017, 358, eaao2648. [Google Scholar]
- Kawamoto, A.; Namba, K. Structural Study of the Bacterial Flagellar Basal Body by Electron Cryomicroscopy and Image Analysis. Methods Mol. Biol. 2017, 1593, 119–131. [Google Scholar]
- Sato-Tomita, A.; Shibayama, N.; Happo, N.; Kimura, K.; Okabe, T.; Matsushita, T.; Park, S.-Y.; Sasaki, Y.C.; Hayashi, K. Development of an X-ray fluorescence holographic measurement system for protein crystals. Rev. Sci. Instrum. 2016, 87, 063707. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, T.; Azai, C.; Terauchi, K.; Yoshimura, M.; Ohta, T. Soft X-Ray Imaging of Cellular Carbon and Nitrogen Distributions in Heterocystous Cyanobacteria. Plant Physiol. 2018, 177, 52–61. [Google Scholar]
- Tosha, T.; Nomura, T.; Nishida, T.; Saeki, N.; Okubayashi, K.; Yamagiwa, R.; Sugahara, M.; Nakane, T.; Yamashita, K.; Hirata, K.; et al. Capturing an initial intermediate during the P450nor enzymatic reaction using time-resolved XFEL crystallography and caged-substrate. Nat. Commun. 2017, 8, 1585. [Google Scholar] [CrossRef]
- Iwakura, N.; Yokoyama, T.; Quaglia, F.; Mitsuoka, K.; Mio, K.; Shigematsu, H.; Shirouzu, M.; Kaji, A.; Kaji, H. Chemical and structural characterization of a model Post-Termination Complex (PoTC) for the ribosome recycling reaction: Evidence for the release of the mRNA by RRF and EF-G. PLoS ONE 2017, 12, e0177972. [Google Scholar] [CrossRef]
- Sugita, Y.; Matsunami, H.; Kawaoka, Y.; Noda, T.; Wolf, M. Cryo-EM structure of the Ebola virus nucleoprotein-RNA complex at 3.6 Å resolution. Nature 2018, 563, 137–140. [Google Scholar] [CrossRef]
- Sugita, Y.; Kawaoka, Y.; Noda, T.; Wolf, M. Structure of the Ebola Virus Nucleocapsid Core by Single Particle Cryo-Electron Microscopy. Microsc. Microanal. 2016, 22, 66–67. [Google Scholar] [CrossRef]
- Ozeki, Y.; Asai, T.; Shou, J.; Yoshimi, H. Multicolor Stimulated Raman Scattering Microscopy With Fast Wavelength-Tunable Yb Fiber Laser. IEEE J. Sel. Top. Quantum Electron. 2019, 25, 1–11. [Google Scholar] [CrossRef]
- Ito, T.; Obara, Y.; Misawa, K. Single-beam phase-modulated stimulated Raman scattering microscopy with spectrally focused detection. J. Opt. Soc. Am. B 2017, 34, 1004. [Google Scholar] [CrossRef]
- Otomo, K.; Hibi, T.; Kozawa, Y.; Nemoto, T. STED microscopy--super-resolution bio-imaging utilizing a stimulated emission depletion. Microscopy 2015, 64, 227–236. [Google Scholar]
- Otomo, K.; Hibi, T.; Fang, Y.-C.; Hung, J.-H.; Tsutsumi, M.; Kawakami, R.; Yokoyama, H.; Nemoto, T. Advanced easySTED microscopy based on two-photon excitation by electrical modulations of light pulse wavefronts. Biomed. Opt. Express 2018, 9, 2671–2680. [Google Scholar] [CrossRef]
- Shoji, K.; Kawano, R.; White, R.J. Spatially Resolved Chemical Detection with a Nanoneedle-Probe-Supported Biological Nanopore. ACS Nano 2019, 13, 2606–2614. [Google Scholar]
- Kawano, R. Nanopore Decoding of Oligonucleotides in DNA Computing. Biotechnol. J. 2018, 13, e1800091. [Google Scholar] [CrossRef] [PubMed]
- Mojarro, A.; Hachey, J.; Bailey, R.; Brown, M.; Doebler, R.; Ruvkun, G.; Zuber, M.T.; Carr, C.E. Nucleic acid extraction and sequencing from low-biomass synthetic Mars analog soils for in situ life detection. Mol. Biol. 2018. [Google Scholar] [CrossRef]
- Xin, Y.; Kim, J.; Ni, M.; Wei, Y.; Okamoto, H.; Lee, J.; Adler, C.; Cavino, K.; Murphy, A.J.; Yancopoulos, G.D.; et al. Use of the Fluidigm C1 platform for RNA sequencing of single mouse pancreatic islet cells. Proc. Natl. Acad. Sci. USA 2016, 113, 3293–3298. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Yamashita, H.; Hayakawa, M.; Onoe, H.; Takinoue, M. Capillary-based Centrifugal Microfluidic Device for Size-controllable Formation of Monodisperse Microdroplets. J. Vis. Exp. 2016. [Google Scholar] [CrossRef]
- Soga, N.; Watanabe, R.; Noji, H. Attolitre-sized lipid bilayer chamber array for rapid detection of single transporters. Sci. Rep. 2015, 5, 11025. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Soga, N.; Hara, M.; Noji, H. Arrayed water-in-oil droplet bilayers for membrane transport analysis. Lab Chip 2016, 16, 3043–3048. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef]
- Klein, A.M.; Mazutis, L.; Akartuna, I.; Tallapragada, N.; Veres, A.; Li, V.; Peshkin, L.; Weitz, D.A.; Kirschner, M.W. Droplet barcoding for single-cell transcriptomics applied to embryonic stem cells. Cell 2015, 161, 1187–1201. [Google Scholar] [CrossRef]
- World Premier International Research Center (WPI) Initiative. Japan Society for the Promotion of Science. Available online: https://www.jsps.go.jp/english/e-toplevel/ (accessed on 3 April 2019).
- LIMMS Website. Available online: http://limmshp.iis.u-tokyo.ac.jp/ (accessed on 3 April 2019).
- Invitational Fellowships for Research in Japan. Japan Society for the Promotion of Science. Available online: https://www.jsps.go.jp/english/e-inv/index.html (accessed on 3 April 2019).
- Bilateral Programs. Japan Society for the Promotion of Science. Available online: https://www.jsps.go.jp/english/e-bilat/index.html (accessed on 3 April 2019).
- Global Activities. Japan Science and Technology Agency (JST). Available online: http://www.jst.go.jp/inter/english/top.html (accessed on 3 April 2019).
- Kitazato, K.; Milliken, R.E.; Iwata, T.; Abe, M.; Ohtake, M.; Matsuura, S.; Arai, T.; Nakauchi, Y.; Nakamura, T.; Matsuoka, M.; et al. The surface composition of asteroid 162173 Ryugu from Hayabusa2 near-infrared spectroscopy. Science 2019. [Google Scholar] [CrossRef]
- Müller, T.G.; O’Rourke, L.; Barucci, A.M.; Pál, A.; Kiss, C.; Zeidler, P.; Altieri, B.; González-García, B.M.; Küppers, M. Physical properties of OSIRIS-REx target asteroid (101955) 1999 RQ 36: Derived from Herschel, VLT/ VISIR, and Spitzer observations. Astron. Astrophys. 2012, 548, A36. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, T.Z.; Kuruma, Y. Recent Advances in Origins of Life Research by Biophysicists in Japan. Challenges 2019, 10, 28. https://doi.org/10.3390/challe10010028
Jia TZ, Kuruma Y. Recent Advances in Origins of Life Research by Biophysicists in Japan. Challenges. 2019; 10(1):28. https://doi.org/10.3390/challe10010028
Chicago/Turabian StyleJia, Tony Z., and Yutetsu Kuruma. 2019. "Recent Advances in Origins of Life Research by Biophysicists in Japan" Challenges 10, no. 1: 28. https://doi.org/10.3390/challe10010028
APA StyleJia, T. Z., & Kuruma, Y. (2019). Recent Advances in Origins of Life Research by Biophysicists in Japan. Challenges, 10(1), 28. https://doi.org/10.3390/challe10010028