
Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae)




Abstract
1. Introduction
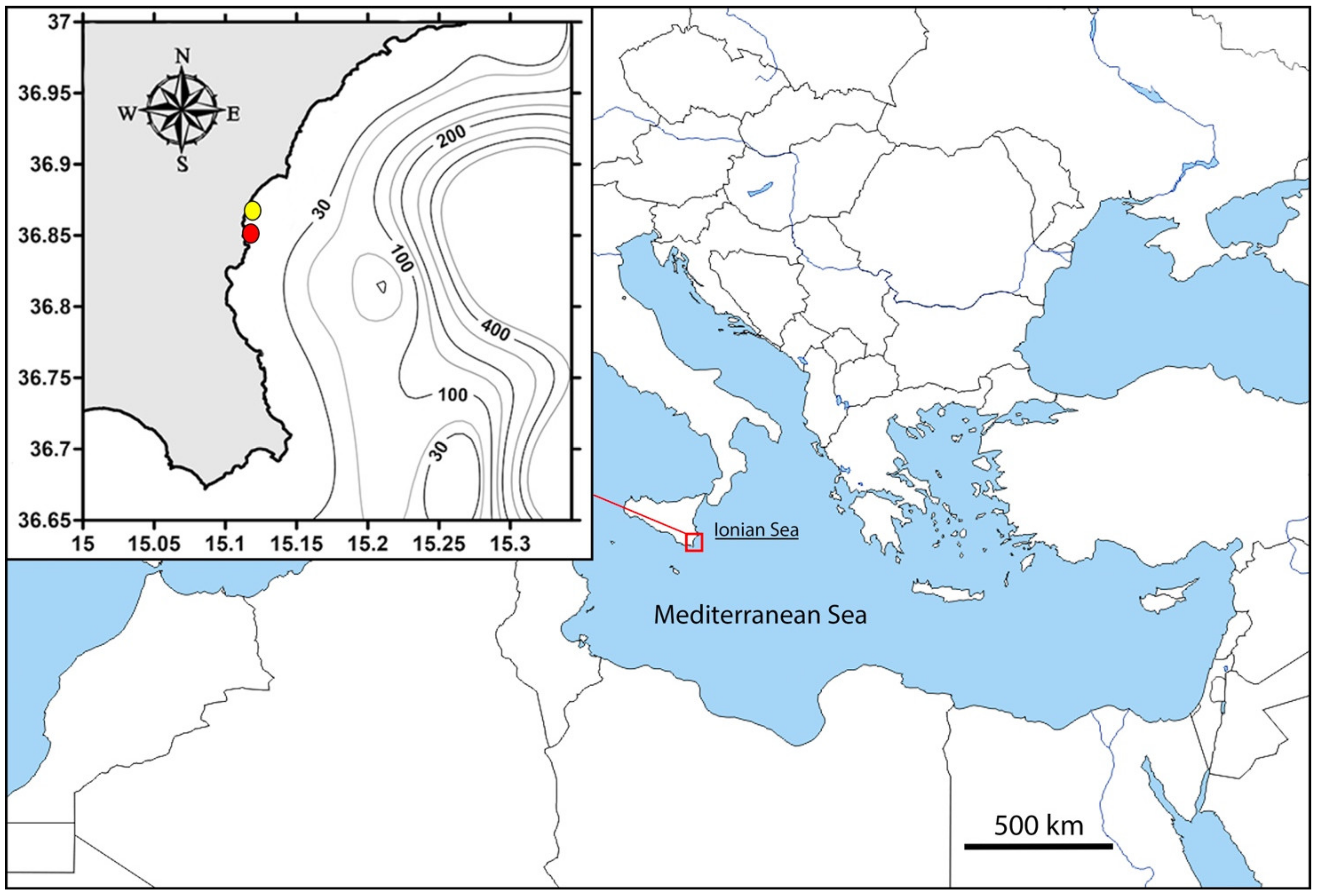
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosholz, E. Ecological and evolutionary consequences of coastal invasions. Trends Ecol. Evol. 2002, 17, 22–27. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef]
- Ricciardi, A.; Palmer, M.E.; Yan, N.D. Should biological invasions be managed as natural disasters? BioScience 2011, 61, 312–317. [Google Scholar] [CrossRef]
- Bailey, S.; Brown, L.; Campbell, M.; Canning-Clode, J.; Carlton, J.; Castro, N.; Chainho, P.; Chan, F.T.; Creed, J.C.; Curd, A.; et al. Trends in the detection of aquatic non-indigenous species across global marine, estuarine and freshwater ecosystems: A 50-year perspective. Divers. Distrib. 2020, 26, 1780–1797. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Noè, S.; Gianguzza, P.; di Trapani, F.; Badalamenti, F.; Vizzini, S.; Fernández, T.V.; Bonaviri, C. Native predators control the population of an invasive crab in no-take marine protected areas. Aquatic Conserv. Mar. Freshw. Ecosyst. 2018, 28, 1229–1237. [Google Scholar] [CrossRef]
- Madenjian, C.P.; Stapanian, M.A.; Witzel, L.D.; Einhouse, D.W.; Pothoven, S.A.; Whitford, H.L. Evidence for predatory control of the invasive round goby. Biol. Invasions 2011, 13, 987–1002. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Species by Family/Subfamily. California Academy of Sciences, San Francisco, USA. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 17 November 2020).
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley and Sons: Hoboken, NJ, USA, 2016; p. 752. [Google Scholar]
- Kovačić, M. Checklist of gobies (Teleostei: Gobiidae) of the Mediterranean Sea and a key for species identification. Zootaxa 2020, 4877, 75–101. [Google Scholar] [CrossRef]
- Hajji, F.; Ouannes-Ghorbel, A.; Ghorbel, M.; Jarboui, O. Diet shifts of Gobius paganellus (Teleostei, Gobiidae) from the Gulf of Gabes (Central Mediterranean). Vie Milieu 2012, 62, 69–76. [Google Scholar]
- Hajji, F.; Ouannes-Ghorbel, A.; Ghorbel, M.; Jarboui, O. Reproductive biology of the rock goby, Gobius paganellus (Actinopterygii: Perciformes: Gobiidae), on the southern Tunisian coast (Gulf of Gabes). Cienc. Mar. 2012, 38, 505–515. [Google Scholar] [CrossRef][Green Version]
- Kovačić, M.; la Mesa, A. Feeding ecology of the Buen’s goby, Buenia affinis, in the Kvarner area (Adriatic Sea). Vie Milieu 2008, 58, 277–281. [Google Scholar]
- Tiralongo, F.; Messina, G.; Lombardo, B.M. First data on habitat preference, diet and length-weight relationship of Gobius incognitus Kovačić & Šanda, 2016 (Pisces: Gobiidae). Acta Adriat. 2020, 61, 67–78. [Google Scholar] [CrossRef]
- Iglésias, S.P.; Vukić, J.; Sellos, D.Y.; Soukupová, T.; Šanda, R. Gobius xoriguer, a new offshore Mediterranean goby (Gobiidae), and phylogenetic relationships within the genus Gobius. Ichthyol. Res. 2021, 1–15. [Google Scholar] [CrossRef]
- Kovačić, M.; Šanda, R. A new species of Gobius (Perciformes: Gobiidae) from the Mediterranean Sea and the redescription of Gobius bucchichi. J. Fish Biol. 2016, 88, 1104–1124. [Google Scholar] [CrossRef]
- Kovačić, M.; Patzner, R.; Schliewen, U. A first quantitative assessment of the ecology of cryptobenthic fishes in the Mediterranean Sea. Mar. Biol. 2012, 159, 2731–2742. [Google Scholar] [CrossRef]
- Miller, P.J. Gobiidae. In Fishes of the North-Eastern Atlantic and Mediterranean; Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; pp. 1019–1085. [Google Scholar]
- Tortonese, E. Mediterranean fishes present in the Red Sea: Pan-oceanic and anti-lessepsian species. Cybium 1984, 8, 99–102. [Google Scholar]
- Engin, S.; Seyhan, K. Biological Characteristics of rock goby, Gobius paganellus (Actinopterygii: Perciformes: Gobiidae), in the South-Eastern Black Sea. Acta Ichthyol. Pisc. 2009, 39, 111–118. [Google Scholar] [CrossRef]
- Miller, P.J. Age, growth, and reproduction of the rock goby, Gobius paganellus L., in the Isle of Man. J. Mar. Biol. Assoc. UK 1961, 41, 737–769. [Google Scholar] [CrossRef]
- Wilkins, H.K.A.; Myers, A.A. Microhabitat utilisation by an assemblage of temperate Gobiidae (Pisces: Teleostei). Mar. Ecol. Prog. Ser. 1992, 90, 103–112. [Google Scholar] [CrossRef]
- Azevedo, J.M.N.; Simas, A.M.V. Age and growth, reproduction and diet of a sublittoral population of the rock goby Gobius paganellus (Teleostei, Gobiidae). Hydrobiologia 2000, 440, 129–135. [Google Scholar] [CrossRef]
- Louiz, I.; Attia, M.B.; Hassine, O.K.B. Some aspects of reproductive biology of Gobius paganellus (Gobiidae) on the north-eastern coasts of Tunisia (Bizerta lagoon). J. Mar. Biol. Assoc. UK 2013, 93, 2235–2246. [Google Scholar] [CrossRef]
- Mazé, R.A. Seasonal and ontogenetic diet shift in an intertidal population of Gobius paganellus (Teleostei, Gobiidae) from the Cantabrian Coast. Vie Milieu 2004, 54, 1–6. [Google Scholar]
- Katsanevakis, S.; Poursanidis, D.; Yokes, M.B.; Mačić, V.; Beqiraj, S.; Kashta, L.; Sghaier, Y.R.; Zakhama-Sraieb, R.; Benamer, I.; Bitar, G.; et al. Twelve years after the first report of the crab Percnon gibbesi (H. Milne Edwards, 1853) in the Mediterranean: Current distribution and invasion rates. J. Biol. Res. 2011, 16, 224–236. [Google Scholar]
- Relini, M.; Orsi, L.; Puccio, V.; Azzurro, E. The exotic crab Percnon gibbesi (H. Milne Edwards, 1853) (Decapoda, Grapsidae) in the Central Mediterranean. Sci. Mar. 2000, 64, 337–340. [Google Scholar] [CrossRef]
- Suaria, G.; Pierucci, A.; Zanello, P.; Fanelli, E.; Chiesa, S.; Azzurro, E. Percnon gibbesi (H. Milne Edwards, 1853) and Callinectes sapidus (Rathbun, 1896) in the Ligurian Sea: Two additional invasive species detection made in collaboration with local fishermen. BioInvasions Rec. 2017, 6, 147–151. [Google Scholar] [CrossRef]
- Pipitone, C.; Badalamenti, F.; Sparrow, A. Contribution to the knowledge of Percnon gibbesi (Decapoda, Grapsidae), an exotic species spreading rapidly in Sicilian waters. Crustaceana 2001, 74, 1009–1017. [Google Scholar]
- Sciberras, M.; Schembri, P.J. Biology and interspecific interactions of the alien crab Percnon gibbesi in the Maltese Islands. Mar. Biol. Res. 2008, 4, 321–332. [Google Scholar] [CrossRef]
- Crocetta, F.; Shokouros-Oskarsson, M.; Doumpas, N.; Giovos, I.; Kalogirou, S.; Langeneck, J.; Tanduo, V.; Tiralongo, F.; Virgili, R.; Kleitou, P. Protect the natives to combat the aliens: Could Octopus vulgaris Cuvier, 1797 be a natural agent for the control of the lionfish invasion in the Mediterranean Sea? J. Mar. Sci. Eng. 2021, 9, 308. [Google Scholar] [CrossRef]
- Kaldonski, N.; Lagrue, C.; Motreuil, S.; Rigaud, T.; Bollache, L. Habitat segregation mediates predation by the benthic fish Cottus gobio on the exotic amphipod species Gammarus roeseli. Naturwissenschaften 2008, 95, 839–844. [Google Scholar] [CrossRef]
- Maljković, A.; van Leeuwen, T.E.; Cove, S.N. Predation on the invasive red lionfish, Pterois volitans (Pisces: Scorpaenidae), by native groupers in the Bahamas. Coral Reefs 2008, 27, 501. [Google Scholar] [CrossRef]
- Louette, G. Use of a native predator for the control of an invasive amphibian. Wildl. Res. 2012, 39, 271–278. [Google Scholar] [CrossRef]
- Giakoumi, S.; Pey, A.; Thiriet, P.; Francour, P.; Guidetti, P. Patterns of predation on native and invasive alien fish in Mediterranean protected areas. Mar. Environ. Res. 2019, 150, 104792. [Google Scholar] [CrossRef]
- Yokes, B.; Galil, B.S. Touchdown—First record of Percnon gibbesi (H. Milne Edwards, 1853) (Crustacea: Decapoda: Grapsidae) from the Levantine coast. Aquat. Invasions 2006, 1, 130–132. [Google Scholar] [CrossRef]
- Galil, B.; Froglia, C.; Noel, P.Y. Atlas of Exotic Species in the Mediterranean. 2008. Available online: www.ciesm.org/atlas/appendix2.html (accessed on 10 December 2020).
- Deudero, S.; Frau, A.; Cerda, M.; Hampel, H. Distribution and densities of the decapod crab Percnon gibbesi, an invasive Grapsidae, in western Mediterranean waters. Mar. Ecol. Prog. Ser. 2005, 285, 151–156. [Google Scholar] [CrossRef]
- Cannicci, S.; Badalamenti, F.; Milazzo, M.; Gomei, M.; Baccarella, A.; Vannini, M. Unveiling the secrets of a successful invader: Preliminary data on the biology and the ecology of the crab Percnon gibbesi (H. Milne Edwards, 1853). Rapp. Com. Int. Mer Médit. 2004, 37, 326. [Google Scholar]
- Jones, R.E.; Petrell, R.J.; Pauly, D. Using modified length-weight relationships to assess the condition of fish. Aquac. Eng. 1999, 20, 261–276. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Riedl, R. Fauna e Flora del Mediterraneo; Franco Muzzio Editore: Padua, Italy, 2010; p. 777. [Google Scholar]
- Hayward, P.J.; Ryland, J.S. Handbook of the Marine Fauna of North-West Europe; Oxford University Press: Oxford, UK, 1990; p. 816. [Google Scholar]
- Hyslop, E.J. Stomach content analysis, a review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Carrassòn, M.; Matallanas, J.; Casadevall, M. Feeding strategies of deep-water morids on the western Mediterranean slope. Deep Sea Res. 1997, 44, 1685–1699. [Google Scholar] [CrossRef]
- N’da, K. Regime alimentaire du rouget de roche Mullus surmuletus (Mullidae) dans le nord du golfe de Gascogne. Cybium 1992, 16, 159–168. [Google Scholar]
- Krebs, J.C. Ecological Methodology; Harper & Row: New York, NY, USA, 1989; p. 620. [Google Scholar]
- Novakowski, G.C.; Hahn, N.S.; Fugi, R. Diet seasonality and food overlap of the fish assemblage in a pantanal pond. Neotrop. Ichthyol. 2008, 6, 567–576. [Google Scholar] [CrossRef]
- Brown, S.C.; Bizzarro, J.J.; Cailliet, G.M.; Ebert, D.A. Breaking with tradition: Redefining measures for diet description with a case study of the Aleutian skate Bathyraja aleutica (Gilbert 1869). Environ. Biol. Fish. 2012, 95, 3–20. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2018. Available online: http://www.R-project.org (accessed on 10 December 2020).
- Zander, C.D.; Berg, J. Feeding ecology of littoral gobiid and blennioid fishes of the Banyuls area (Mediterranean Sea) II. Prey selection and size preference. Vie Milieu 1984, 34, 149–157. [Google Scholar]
- Cubedo-Verdiell, D.; Oliva-Paterna, F.J.; Torralva, M. Length-weight relationships for 22 fish species of the Mar Menor coastal lagoon (western Mediterranean Sea). J. Appl. Ichthyol. 2006, 22, 293–294. [Google Scholar] [CrossRef]
- Giakoumi, S.; Katsanevakis, S.; Albano, P.G.; Azzurro, E.; Cardoso, A.C.; Cebrian, E.; Deidun, A.; Edelist, D.; Francour, P.; Jimenez, C.; et al. Management priorities for marine invasive species. Sci. Total Environ. 2019, 688, 976–982. [Google Scholar] [CrossRef]
- Byun, C.; Lee, E.J. Ecological application of biotic resistance to control the invasion of an invasive plant, Ageratina altissima. Ecol. Evol. 2017, 7, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Funk, J.L.; Cleland, E.E.; Suding, K.N.; Zavaleta, E.S. Restoration trough reassembly: Plant traits and invasion resistance. Trends Ecol. Evol. 2008, 23, 695–703. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; The University of Chicago Press: Chicago, IL, USA, 1958. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | N | N Avola | N Noto | N Prey | TL (cm) | SL (cm) | W (g) |
|---|---|---|---|---|---|---|---|
| 4 September 2020 | 15 | 5 | 10 | 11 | 9.52 | 7.89 | 11.45 |
| 14 September 2020 | 11 | 2 | 9 | 7 | 9.24 | 7.68 | 10.52 |
| 21 September 2020 | 12 | 4 | 8 | 9 | 9.54 | 8 | 11.26 |
| 28 September 2020 | 16 | 8 | 8 | 12 | 9.50 | 7.99 | 11.78 |
| 2 October 2020 | 12 | 5 | 7 | 10 | 9.82 | 8.20 | 12.91 |
| 9 October 2020 | 15 | 10 | 5 | 13 | 9.83 | 8.25 | 12.09 |
| 16 October 2020 | 13 | 3 | 10 | 9 | 9.09 | 7.57 | 10.05 |
| 27 October 2020 | 14 | 6 | 8 | 6 | 9.56 | 7.96 | 11.80 |
| 3 November 2020 | 12 | 8 | 4 | 8 | 9.59 | 8.06 | 11.48 |
| 10 November 2020 | 15 | 8 | 7 | 10 | 9.19 | 7.71 | 10.69 |
| 22 November 2020 | 14 | 9 | 5 | 11 | 9.8 | 8.19 | 12.85 |
| 5 December 2020 | 13 | 4 | 9 | 9 | 9.49 | 7.95 | 11.26 |
| Prey types | %F | %N | %W | IRI | %IRI |
|---|---|---|---|---|---|
| Mollusca | 11.31 | 9.36 | 10.24 | 49.916 | 1.62 |
| Aplysia sp. | 0.87 | 0.72 | 1.63 | 2.0435 | 0.07 |
| Bittium lacteum | 0.87 | 0.72 | 0.54 | 1.0956 | 0.04 |
| Patella caerulea | 1.74 | 1.44 | 1.01 | 4.2610 | 0.14 |
| Tritia cuvierii | 1.74 | 1.44 | 1.90 | 5.8116 | 0.19 |
| Gastropoda n.i. | 4.35 | 3.60 | 3.67 | 31.6087 | 1.02 |
| Trochidae n.i. | 1.74 | 1.44 | 1.49 | 5.0956 | 0.16 |
| Polychaeta | 2.61 | 2.16 | 0.95 | 8.113 | 0.26 |
| Polychaetes n.i. | 2.61 | 2.16 | 0.95 | 8.113 | 0.26 |
| Crustacea | 89.59 | 77.73 | 79.65 | 2925.5660 | 94.42 |
| Clibanarius erythropus | 6.96 | 6.47 | 4.27 | 74.7130 | 2.41 |
| Eriphia verrucosa | 4.35 | 3.60 | 3.49 | 30.8415 | 1.00 |
| Pachygrapsus marmoratus | 5.22 | 5.04 | 5.03 | 52.5391 | 1.70 |
| Percnon gibbesi | 33.91 | 30.22 | 40.15 | 2386.4608 | 77.01 |
| Pisidia sp. | 6.09 | 5.04 | 12.42 | 106.2782 | 3.43 |
| Xantho sp. | 6.96 | 5.76 | 4.17 | 69.0783 | 2.23 |
| Amphipoda n.i. | 6.09 | 5.04 | 1.22 | 38.1234 | 1.23 |
| Brachyura n.i. | 6.96 | 5.76 | 4.61 | 69.3753 | 2.24 |
| Caridea n.i. | 6.96 | 5.76 | 1.49 | 50.4347 | 1.63 |
| Isopoda n.i. | 6.09 | 5.04 | 2.80 | 47.7217 | 1.54 |
| Algae | 6.09 | 5.04 | 3.53 | 52.1652 | 1.68 |
| Seagrass (Posidonia oceanica) | 0.87 | 0.72 | 0.43 | 1.0005 | 0.03 |
| Digested | 6.09 | 5.04 | 5.19 | 62.2695 | 2.01 |
| Total Length (TL)-cm | Standard Length (SL)-cm | Weight (W)-g | ||||||
|---|---|---|---|---|---|---|---|---|
| Range | Mean | SD | Range | Mean | SD | Range | Mean | SD |
| 7.1–11.6 | 9.5 | 1.11 | 5.9–9.6 | 7.9 | 0.97 | 4.98–20.14 | 11.53 | 3.80 |
| a | C.I. a | b | C.I. b | r2 | p-value | |||
| 0.014 | 0.011–0.018 | 2.959 | 2.849–3.068 | 0.95 | >0.05 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiralongo, F.; Messina, G.; Lombardo, B.M. Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae). J. Mar. Sci. Eng. 2021, 9, 393. https://doi.org/10.3390/jmse9040393
Tiralongo F, Messina G, Lombardo BM. Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae). Journal of Marine Science and Engineering. 2021; 9(4):393. https://doi.org/10.3390/jmse9040393
Chicago/Turabian StyleTiralongo, Francesco, Giuseppina Messina, and Bianca Maria Lombardo. 2021. "Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae)" Journal of Marine Science and Engineering 9, no. 4: 393. https://doi.org/10.3390/jmse9040393
APA StyleTiralongo, F., Messina, G., & Lombardo, B. M. (2021). Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae). Journal of Marine Science and Engineering, 9(4), 393. https://doi.org/10.3390/jmse9040393