
DNA Sequence Analyses Reveal Two New Species of Caloglossa (Delesseriaceae, Rhodophyta) from the Skin of West Indian Manatees


Abstract
1. Introduction
2. Materials and Methods
3. Results
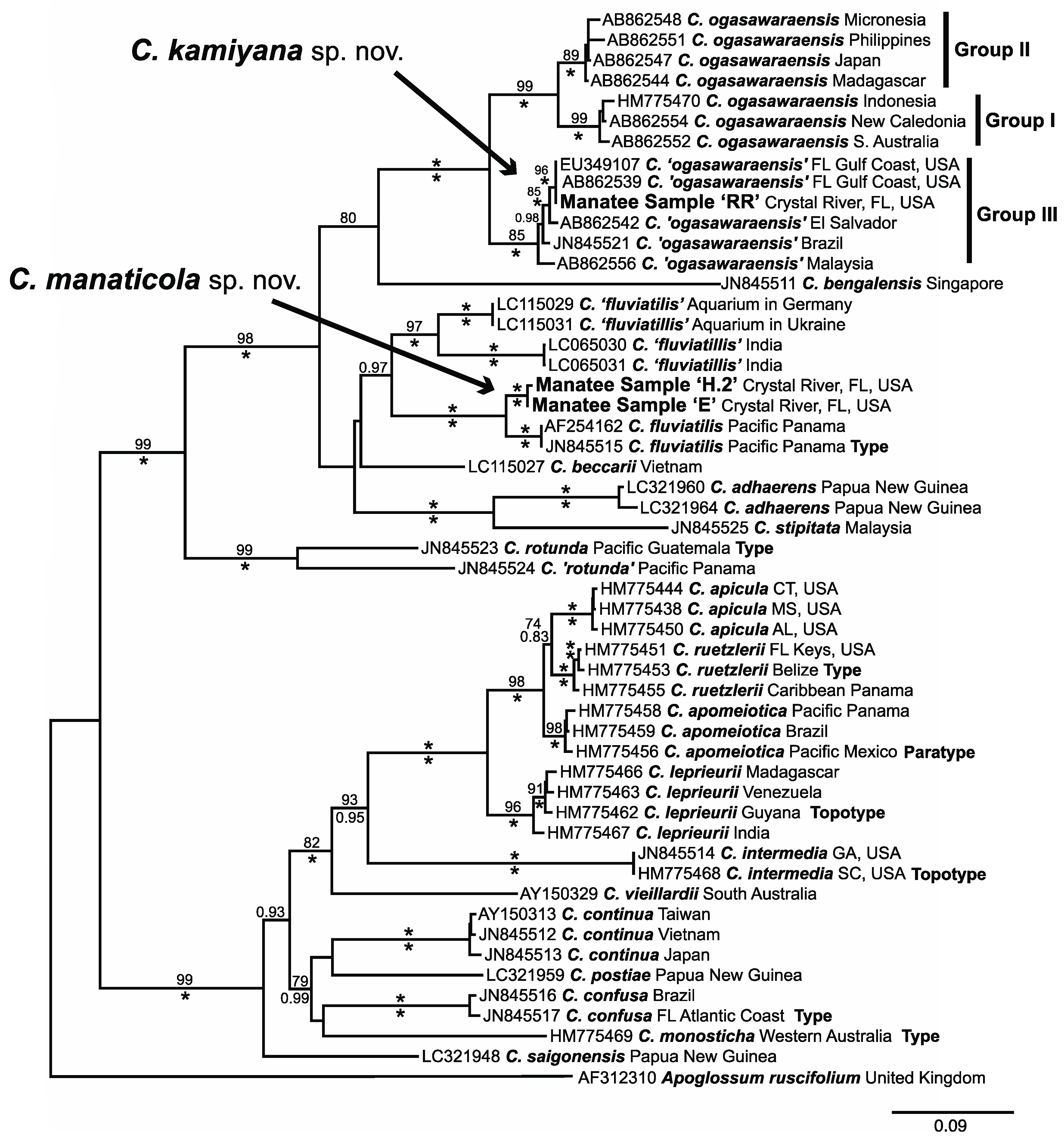
3.1. Molecular Analyses
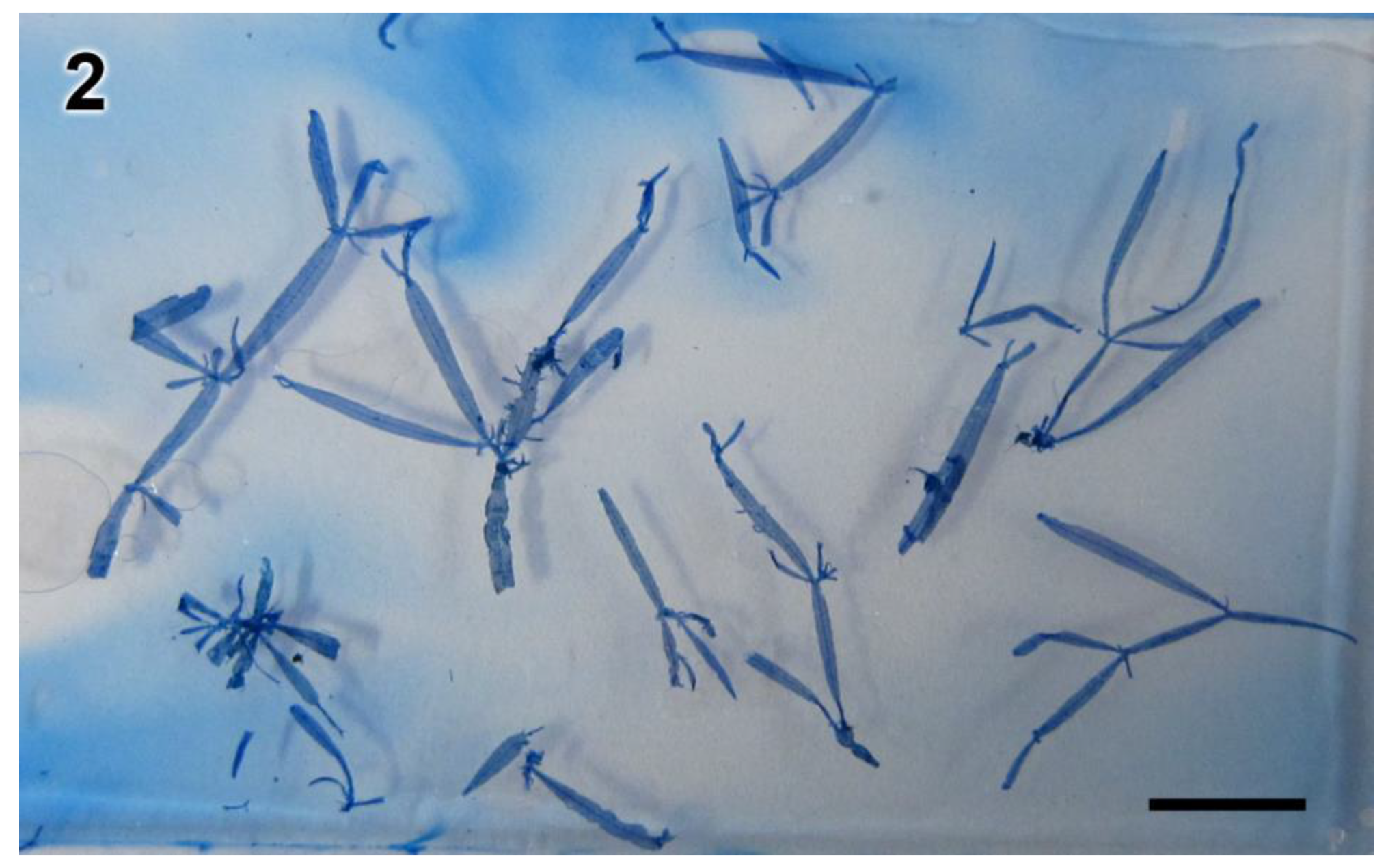
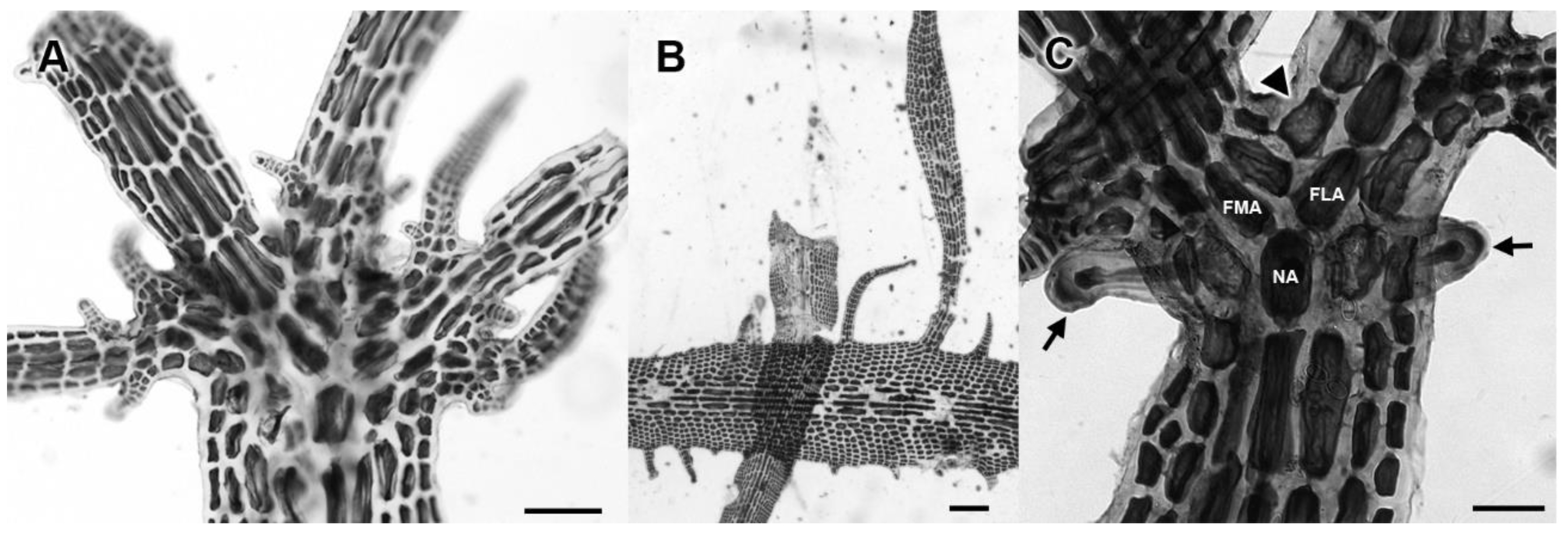
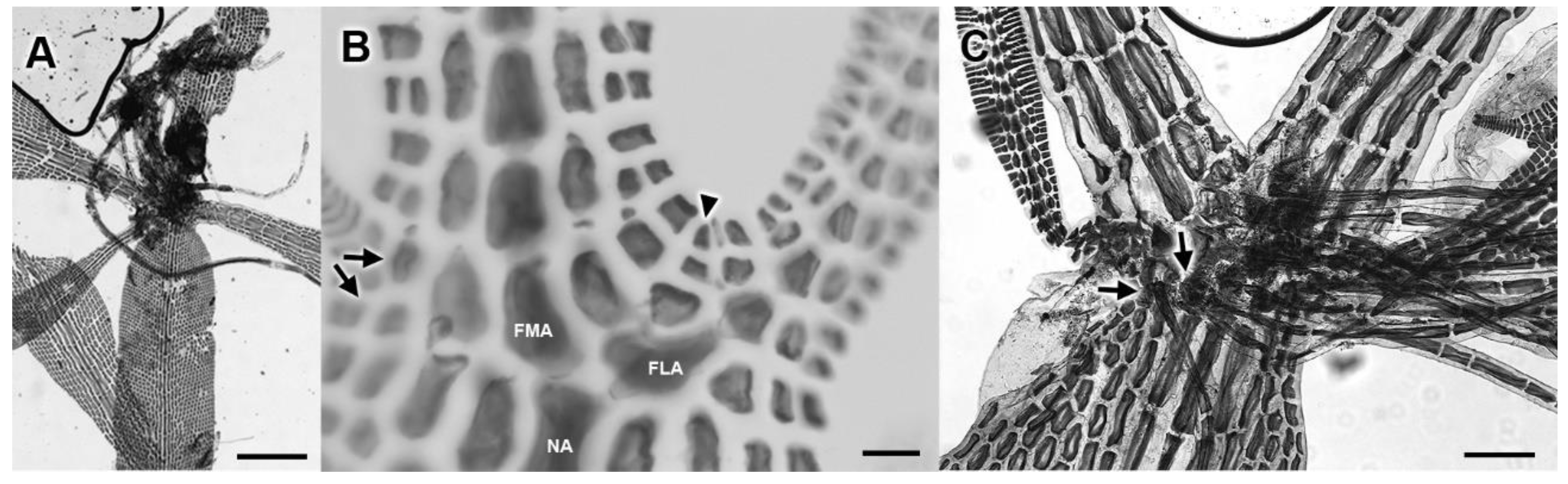
3.2. Morphological Analyses
3.3. New Species
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheang, C.C.; Tsang, L.M.; Chu, K.H.; Chen, I.-J.; Chan, B.K.K. Host-specific phenotypic plasticity of the turtle barnacle Chelonibia testudinaria: A widespread generalist rather than a specialist. PLoS ONE 2013, 8, e57592. [Google Scholar] [CrossRef] [PubMed]
- Ewers-Saucedo, C.; Chan, B.K.K.; Zardus, J.D.; Wares, J.P. Parallel patterns of host-specific morphology and genetic admixture in sister lineages of a commensal barnacle. Biol. Bull. 2017, 232, 171–185. [Google Scholar] [CrossRef]
- Siciliano, S.; Cardoso, J.; Francisco, A.; De Souza, S.P.; Hauser-Davis, R.A.; Iwasa-Arai, T. Epizoic barnacle (Xenobalanus globicipitis) infestations in several cetacean species in south-eastern Brazil. Mar. Biol. Res. 2020, 16, 356–368. [Google Scholar] [CrossRef]
- Denys, L. Morphology and taxonomy of epizoic diatoms (Epiphalaina and Tursiocola) on a sperm whale (Physeter macrocephalus) stranded on the coast of Belgium. Diatom Res. 1997, 12, 1–18. [Google Scholar] [CrossRef]
- Holmes, R.W. The morphology of diatoms epizoic on cetaceans and their transfer from Cocconeis to two new genera, Benettella and Epipellis. Br. Phycol. J. 1985, 20, 43–57. [Google Scholar] [CrossRef]
- Holmes, R.W.; Nagasawa, S.; Takano, H. The morphology and geographic distribution of epidermal diatoms of the Dall’s porpoise (Phocoenoides dalli True) in the Northern Pacific Ocean. Bull. Natl. Sci. Mus. Tokyo Ser. B 1993, 19, 1–18. [Google Scholar]
- Nemoto, T. On the diatoms of the skin film of the whales in the Northern Pacific. Sci. Rep. Whales Res. Inst. Tokyo 1956, 11, 99–132. [Google Scholar]
- Frankovich, T.A.; Sullivan, M.J.; Stacy, N.I. Three new species of Tursiocola (Bacillariophyta) from the skin of the West Indian manatee. Phytotaxa 2015, 204, 33–48. [Google Scholar] [CrossRef][Green Version]
- Frankovich, T.A.; Ashworth, M.P.; Sullivan, M.J.; Theriot, E.C.; Stacy, N.I. Epizoic and apochlorotic Tursiocola species (Bacillariophyta) from the skin of Florida manatees (Trichechus manatus latirostris). Protist 2018, 169, 539–568. [Google Scholar] [CrossRef]
- Irvine, A. Manatee metabolism and its influence on distribution in Florida. Biol. Conserv. 1983, 25, 315–334. [Google Scholar] [CrossRef]
- Deutsch, C.; Reid, J.; Bonde, R.K.; Easton, D.; Kochman, H.; O’Shea, T. Seasonal movements, migratory behavior and site fidelity of West Indian Manatee along the Atlantic coast of the United States. Wildl. Monogr. 2003, 151, 1–77. [Google Scholar]
- Meyer, W.; Seegers, U. A preliminary approach to epidermal antimicrobial defense in the Delphinidae. Mar. Biol. 2004, 144, 841–844. [Google Scholar] [CrossRef]
- Baum, C.; Meyer, W.; Stelzer, R.; Feischer, L.G.; Siebers, D. Average nanorough skin surface of the pilot whale (Globicephala melas, Delphinidae): Considerations of the self-cleaning abilities based on nanoroughness. Mar. Biol. 2002, 140, 653–657. [Google Scholar] [CrossRef]
- Bledsoe, E.L.; Harr, K.E.; Cichra, M.F.; Philips, E.J.; Bonde, R.K.; Lowe, M. A comparison of biofouling communities associated with free-ranging and captive Florida manatees (Trichechus manatus latirostris). Mar. Mammal Sci. 2006, 22, 997–1003. [Google Scholar] [CrossRef]
- Majewska, R.; Goosen, W.E. For better, for worse: Manatee-associated Tursiocola (Bacillariophyta) remain faithful to their host. J. Phycol. 2020, 56, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Sentíes, A.; Espinoza-Avalos, J.; Zurita, J.C. Epizoic algae of nesting sea turtles Caretta caretta (L.) and Chelona mydas (L.) from the Mexican Caribbean. Bull. Mar. Sci. 1999, 64, 185–188. [Google Scholar]
- Serio, D.; Furnari, G.; Moro, I.; Sciuto, K. Molecular and morphological characterisation of Melanothamnus testudinis sp. nov. (Rhodophyta, Rhodomelaceae) and its distinction from Polysiphonia carettia. Phycologia 2020, 59, 281–291. [Google Scholar] [CrossRef]
- Woodworth, K.A.; Frankovich, T.A.; Freshwater, D.W. Melanothamnus maniticola sp. nov. (Ceramiales, Rhodophyta): An epizoic species evolved for living on the West Indian manatee. J. Phycol. 2019, 55, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.J.K.; Wynne, M.J. Patulophycus eclipes gen. et sp. nov. (Delesseriaceae, Rhodophyta) from the southwestern Pacific. Syst. Bot. 1992, 17, 409–416. [Google Scholar] [CrossRef]
- Taylor, R.L.; Bailey, J.C.; Freshwater, D.W. Systematics of Cladophora spp. (Chlorophyta) from North Carolina, USA, based upon morphology and DNA sequence data with a description of Cladophora subtilissima sp. nov. J. Phycol. 2017, 53, 541–556. [Google Scholar] [CrossRef]
- Freshwater, D.W.; Rueness, J. Phylogenetic relationships of some European Gelidium (Gelidiales, Rhodophyta) species, based on rbcL nucleotide sequence analysis. Phycologia 1994, 33, 187–194. [Google Scholar] [CrossRef]
- Stuercke, B.; Freshwater, D.W. Consistency of morphological characters used to delimit Polysiphonia sensu lato species (Ceramiales, Florideophyceae): Analyses of North Carolina specimens. Phycologia 2008, 47, 541–559. [Google Scholar] [CrossRef]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Stamatakis, R.; Ludwig, T.; Meier, H. RAxML-III: A fast program for maximum likelihood-based inference of large phylogenetic trees. Bioinformatics 2005, 21, 456–463. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Kamiya, M.; West, J.A. Cryptic diversity in the euryhaline red alga Caloglossa ogasawaraensis (Delesseriacea, Ceramiales). Phycologia 2014, 53, 374–382. [Google Scholar] [CrossRef]
- Kamiya, M.; West, J.A.; Karsten, U.; Ganesan, E.K. Molecular and morphological delineation of Caloglossa beccarii and related species (Delesseriaceae, Rhodophyta). Phycologia 2016, 55, 640–649. [Google Scholar] [CrossRef]
- Okamura, K. On the algae from Ogasawara-jima (Bonin Islands). Bot. Mag. Tokyo 1897, 11, 1–17. [Google Scholar] [CrossRef][Green Version]
- Tanaka, J.; Kamiya, M. Reproductive structure of Caloglossa ogasawaraensis Okamura (Ceramiales, Rhodophyceae) in nature and culture. Jpn. J. Phycol. 1993, 41, 113–121. [Google Scholar]
- Kamiya, M.; Zuccarello, G.C.; West, J.A. Evolutionary relationships of the genus Caloglossa (Delesseriaceae, Rhodophyta) inferred from large-subunit ribosomal RNA gene sequences, morphological evidence and reproductive compatability, with description of a new species from Guatemala. Phycologia 2003, 42, 478–497. [Google Scholar] [CrossRef]
- Krayesky, D.M.; Norris, J.N.; West, J.A.; Kamiya, M.; Viguerie, M.; Wysor, B.S.; Fredericq, S. Two new species of Caloglossa (Delesseriaceae, Rhodophyta) from the Americas, C. confusa and C. fluviatilis. Phycologia 2012, 51, 513–530. [Google Scholar] [CrossRef][Green Version]
- Kamiya, M.; West, J.A.; King, R.J.; Zuccarello, G.C.; Tanaka, J.; Hara, Y. Evolutionary divergence in the red algae Caloglossa leprieurii and C. apomeiotica. J. Phycol. 1998, 34, 361–370. [Google Scholar] [CrossRef]
- Kamiya, M.; Tanaka, J.; King, R.J.; West, J.A.; Zuccarello, G.C.; Kawai, H. Reproductive and genetic distinction between broad and narrow entities of Caloglossa continua (Delesseriaceae, Rhodophyta). Phycologia 1999, 38, 356–367. [Google Scholar] [CrossRef]
- Kamiya, M.; West, J.A.; Zuccarello, G.C.; Kawai, H. Caloglossa intermedia sp. nov. (Rhodophyta) from the western Atlantic coast: Molecular and morphological analyses with special reference to C. leprieurii and C. monosticha. J. Phycol. 2000, 36, 411–420. [Google Scholar] [CrossRef]
- Zuccarello, G.C.; Bartlett, J.; Yeates, P.H. Differentiation of Caloglossa leprieurii (Rhodophyta) populations in northern and eastern Australia using plastid haplotypes. Eur. J. Phycol. 2000, 35, 357–363. [Google Scholar] [CrossRef]
- Gabrielson, P.W.; Hughey, J.R.; Diaz-Pulido, G. Genomics reveals abundant speciation in the coral reef building alga Porolithon onkodes (Corallinales, Rhodophyta). J. Phycol. 2018, 54, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Jesus, P.B.; Costa, A.L.; Castro Nunes, J.M.; Manghisi, A.; Genovese, G.; Morabito, M.; Schnadelbach, A.S. Species delimitation methods reveal cryptic diversity in the Hypnea cornuta complex (Cystocloniaceae, Rhodophyta). Eur. J. Phycol. 2019, 54, 135–153. [Google Scholar] [CrossRef]
- Hughey, J.R.; Leister, G.L.; Gabrielson, P.W.; Hommersand, M.H. Sarcopeltis gen. nov. (Gigartinaceae, Rhodophyta), with S. skottsbergii comb. nov. from southern South America and S. antarctica sp. nov. from the Antarctic Peninsula. Phytotaxa 2020, 468, 75–88. [Google Scholar] [CrossRef]
- Krayesky, D.M.; Norris, J.; West, J.A.; Fredercq, S. The Caloglossa leprieurii-complex (Delesseriaceae, Rhodphyta) in the Americas: The elucidation of overlooked species based on molecular and morphological evidence. Cryptogam. Algol. 2011, 32, 37–62. [Google Scholar] [CrossRef]
- Kamiya, M.; West, J.A.; Zuccarello, G.C. High species richness and genetic diversity of the genus Caloglossa (Delesseriaceae, Rhodophyta) in New Ireland, Papua New Guinea. Phycol. Res. 2018, 66, 218–230. [Google Scholar] [CrossRef]
- Seto, R. Typification of Caloglossa ogasawaraensis Okamura (Ceramiales, Rhodophyta). Jpn. J. Phycol. 1985, 33, 317–319. [Google Scholar]
- Hughey, J.; Gabrielson, P.W. Comment on “Acquiring DNA sequence data from dried archival red algae (Florideophyceae) for the purpose of applying available names to contemporary genetic species: A critical assessment”. Botany 2012, 90, 1191–1194. [Google Scholar] [CrossRef][Green Version]
- Hughey, J.R.; Gabrielson, P.W.; Rohmer, L.; Tortolani, J.; Silva, M.; Miller, K.A.; Young, J.D.; Martell, C.; Ruediger, E. Minimally destructive sampling of type specimens of Pyropia (Bangiales, Rhodophyta) recovers complete plastid and mitochondrial genomes. Sci. Rep. 2014, 4, 5113. [Google Scholar] [CrossRef]
- Lin, S.-M.; Fredericq, S.; Hommersand, M.H. Systematics of the Delesseriaceae (Ceramiales, Rhodophyta) based on large subunit rDNA and rbcL sequences, including the Phycodryoideae, subfam. nov. J. Phycol. 2001, 37, 881–899. [Google Scholar] [CrossRef]
- West, J.A.; Kamiya, M.; Ganesan, E.K.; Louiseaux-de Goër, S.; Jose, L. Caloglossa beccarii (Delesseriaceae, Rhodophyta) from freshwater rivers in Kerala, India, a critical new record. Algae 2015, 30, 207–216. [Google Scholar] [CrossRef]
- Børgesen, F. The marine algae of the Danish West Indies. Part 3. Rhodophyceae (4). Dansk Bot. Ark. 1918, 3, 241–304. [Google Scholar]
- Taylor, W.R. Marine Algae of the Eastern Tropical and Subtopical Coasts of the Americas; The University of Michigan Press: Ann Arbor, MI, USA, 1960; p. 870. [Google Scholar]
- Vianna, J.A.; Bonde, R.K.; Caballero, S.; Giraldo, J.P.; Lima, R.P.; Clark, A.; Marmontel, M.; Morales-Vela, B.; Joséde Souza, M.; Parr, L.; et al. Phylogeography, phylogeny and hybridization in trichechid sirenians: Implications for manatee conservation. Mol. Ecol. 2006, 15, 433–447. [Google Scholar] [CrossRef]
- Hunter, M.E.; Auil-Gomez, N.E.; Tucker, K.P.; Bonde, R.K.; Powell, J.; McGuire, P.M. Low genetic variation and evidence of limited dispersal in the regionally important Belize manatee. Anim. Conserv. 2010, 13, 592–602. [Google Scholar] [CrossRef]
- Santizábal, P.; Mignucci-Giannoni, A.A.; Duchene, S.; Caicedo-Herrera, D.; Perea-Sicchar, C.M.; García-Dávila, C.R.; Trujillo, F.; Caballero, S.J. Phylogeography and sex-biased dispersal across riverine manatee populations (Trichechus inunguis and Trichechus manatus) in South America. PLoS ONE 2012, 7, e52468. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group I | Group II | Group III: C. kamiyana sp. nov. | |
|---|---|---|---|
| Group I (n = 13) | 0.0–0.9% | ||
| Group II (n = 10) | 2.9–3.6% | 0.0–1.2% | |
| Group III = C. kamiyana sp. nov. (n = 8) | 4.9–5.3% | 4.6–5.3% | 0.0–1.4% |
| C. fluviatilis | Manatee Samples | C. ‘fluviatilis’ European Aquaria | C. ‘fluviatilis’ India | |
|---|---|---|---|---|
| C. fluviatilis (n = 2) | 0.0% | |||
| Manatee samples (n = 2) | 2.0–2.4% | 0.5% | ||
| C. ‘fluviatilis’ European aquaria (n = 2) | 6.0–6.1% | 5.9–6.1% | 0.0% | |
| C. ‘fluviatilis’ India (n = 2) | 6.9% | 6.5% | 4.8% | 0.0% |
| C. apicula | C. apomeiotica | C. ruetzleri | |
|---|---|---|---|
| C. apicula (n = 14) | 0.0–0.4% | ||
| C. apomeiotica (n = 4) | 2.7–3.3% | 0.0–0.7% | |
| C. ruetzleri (n = 6) | 2.8–3.6% | 2.4–3.3% | 0.0–1.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freshwater, D.W.; Miller, C.E.; Frankovich, T.A.; Wynne, M.J. DNA Sequence Analyses Reveal Two New Species of Caloglossa (Delesseriaceae, Rhodophyta) from the Skin of West Indian Manatees. J. Mar. Sci. Eng. 2021, 9, 163. https://doi.org/10.3390/jmse9020163
Freshwater DW, Miller CE, Frankovich TA, Wynne MJ. DNA Sequence Analyses Reveal Two New Species of Caloglossa (Delesseriaceae, Rhodophyta) from the Skin of West Indian Manatees. Journal of Marine Science and Engineering. 2021; 9(2):163. https://doi.org/10.3390/jmse9020163
Chicago/Turabian StyleFreshwater, D. Wilson, Cathryn E. Miller, Thomas A. Frankovich, and Michael J. Wynne. 2021. "DNA Sequence Analyses Reveal Two New Species of Caloglossa (Delesseriaceae, Rhodophyta) from the Skin of West Indian Manatees" Journal of Marine Science and Engineering 9, no. 2: 163. https://doi.org/10.3390/jmse9020163
APA StyleFreshwater, D. W., Miller, C. E., Frankovich, T. A., & Wynne, M. J. (2021). DNA Sequence Analyses Reveal Two New Species of Caloglossa (Delesseriaceae, Rhodophyta) from the Skin of West Indian Manatees. Journal of Marine Science and Engineering, 9(2), 163. https://doi.org/10.3390/jmse9020163