
Assessment of Water Quality and Phytoplankton Structure of Eight Alexandria Beaches, Southeastern Mediterranean Sea, Egypt


 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
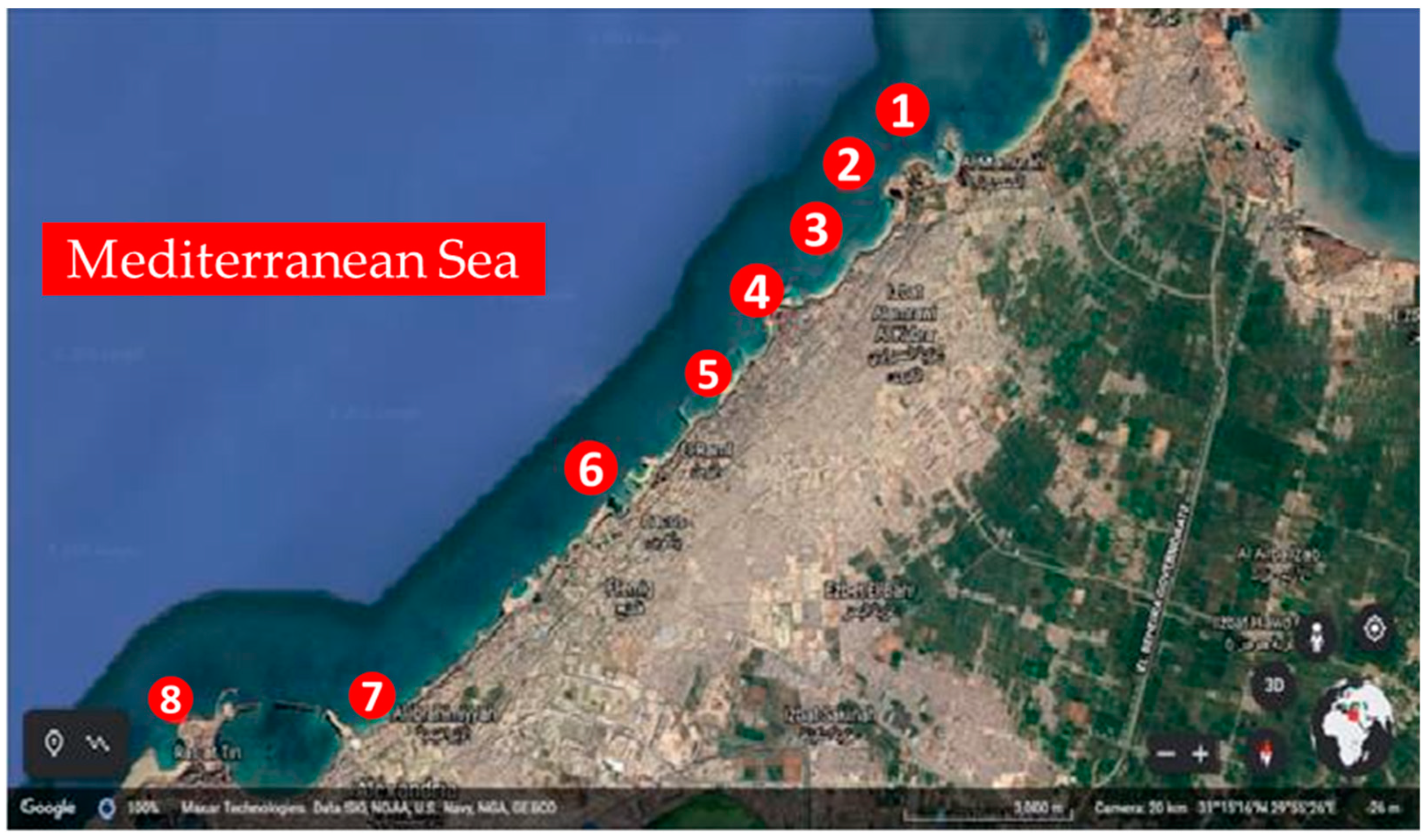
2.1. Study Area
2.2. Sampling and Laboratory Analysis
2.3. Water Quality Index (WQI)
2.4. Species Diversity Index
2.5. Pielou’s Evenness Index (J)
2.6. Statistical Analysis
3. Results
3.1. Physicochemical Conditions
3.2. Phytoplankton Community Structure and Composition
3.3. Seasonal Variation of Phytoplankton
3.4. Correlation Analysis between Biotic and Abiotic Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azov, Y. Eastern Mediterranean—A marine desert? Mar. Pollut. Bull. 1991, 23, 225–232. [Google Scholar] [CrossRef]
- Abo-Taleb, H.; Ashour, M.; El-Shafei, A.; Alataway, A.; Maaty, M.M.; Abo-Taleb, H.A.; Ashour, M.; El-Shafei, A.; Alataway, A.; Maaty, M.M. Biodiversity of Calanoida Copepoda in Different Habitats of the North-Western Red Sea (Hurghada Shelf). Water 2020, 12, 656. [Google Scholar] [CrossRef]
- Dorgham, M.M. Phytoplankton dynamics and ecology in a polluted area on the Alexandria Mediterranean coast. In Proceedings of the 3rd International Conference on Mediterranean Coastal Environment, Qawra, Malta, 11–14 November 1997; Volume 1, pp. 151–160. [Google Scholar]
- Alprol, A.E.; Heneash, A.M.M.; Soliman, A.M.; Ashour, M.; Alsanie, W.F.; Gaber, A.; Mansour, A.T. Assessment of Water Quality, Eutrophication, and Zooplankton Community in Lake Burullus, Egypt. Diversity 2021, 13, 268. [Google Scholar] [CrossRef]
- Zaghloul, F.A.-R.; Khairy, H.M.; Hussein, N.R. Assessment of phytoplankton community structure and water quality in the Eastern Harbor of Alexandria, Egypt. Egypt. J. Aquat. Res. 2020, 46, 145–151. [Google Scholar] [CrossRef]
- El-Sherif, Z.; Mikhail, S.K. Phytoplankton dynamics in the southwestern part of Abu Qir Bay, Alexandria, Egypt. Egypt. J. Aquat. Biol. Fish 2003, 7, 219–239. [Google Scholar]
- Ismael, A.A.; Dorgham, M.M. Ecological indices as a tool for assessing pollution in El-Dekhaila Harbour (Alexandria, Egypt). Oceanologia 2003, 45, 121–131. [Google Scholar]
- Gharib, S.M.; Dorgham, M.M. Eutrophication stress on phytoplankton community in the Western Harbour of Alexandria, Egypt. Int. J. Ocean. Oceanogr. 2006, 1, 261–273. [Google Scholar]
- Dorgham, M.M.; Abdel-Aziz, N.E.; El-Deeb, K.Z.; Okbah, M.A. Eutrophication problems in the western harbour of Alexandria, Egypt. Oceanologia 2004, 46, 25–44. [Google Scholar]
- Heneash, A.M.M.; Tadrose, H.R.Z.; Hussein, M.M.A.; Hamdona, S.K.; Abdel-Aziz, N.; Gharib, S.M. Potential effects of abiotic factors on the abundance and distribution of the plankton in the Western Harbour, south-eastern Mediterranean Sea, Egypt. Oceanologia 2015, 57, 61–70. [Google Scholar] [CrossRef]
- Zingone, A.; Escalera, L.; Aligizaki, K.; Fernández-tejedor, M.; Ismael, A.; Montresor, M.; Mozetič, P.; Taş, S.; Totti, C. Toxic marine microalgae and noxious blooms in the Mediterranean Sea: A contribution to the Global HAB Status Report. Harmful Algae 2020, 101843. [Google Scholar] [CrossRef] [PubMed]
- Shams El-Din, N.; Abel Halim, A. Changes in phytoplankton community structure at three touristic sites at western Alexandria Beach. Egypt. J. Aquat. Biol. Fish. 2008, 12, 85–118. [Google Scholar] [CrossRef][Green Version]
- Abbas, E.M.; Ali, F.S.; Desouky, M.G.; Ashour, M.; El-Shafei, A.; Maaty, M.M.; Sharawy, Z.Z. Novel comprehensive molecular and ecological study introducing coastal mud shrimp (Solenocera crassicornis) recorded at the Gulf of suez, Egypt. J. Mar. Sci. Eng. 2021, 9, 9. [Google Scholar] [CrossRef]
- Mabrouk, M.M.; Ashour, M.; Labena, A.; Zaki, M.A.A.; Abdelhamid, A.F.; Gewaily, M.S.; Dawood, M.A.O.; Abualnaja, K.M.; Ayoub, H.F. Nanoparticles of Arthrospira platensis improves growth, antioxidative and immunological responses of Nile tilapia (Oreochromis niloticus) and its resistance to Aeromonas hydrophila. Aquac. Res. 2021, 1–11. [Google Scholar] [CrossRef]
- Ashour, M.; Abo-Taleb, H.; Abou-Mahmoud, M.; El-Feky, M.M.M. Effect of the integration between plankton natural productivity and environmental assessment of irrigation water, El-Mahmoudia Canal, on aquaculture potential of Oreochromis niloticus. Turk. J. Fish. Aquat. Sci. 2018, 18, 1163–1175. [Google Scholar] [CrossRef]
- Metwally, S.A.; El-Naggar, H.A.; El-Damhougy, K.A.; Bashar, M.A.E.; Ashour, M.; Abo-Taleb, H.A.H. GC-MS analysis of bioactive components in six different crude extracts from the Soft Coral (Sinularia maxim) collected from Ras Mohamed, Aqaba Gulf, Red Sea, Egypt. Egypt. J. Aquat. Biol. Fish. 2020, 24, 425–434. [Google Scholar] [CrossRef]
- Ashour, M.; Mabrouk, M.M.; Ayoub, H.F.; El-Feky, M.M.M.; Sharawy, Z.Z.; Hoseinifar, S.H.; Rossi, W.; Van Doan, H.; El-Haroun, E.; Goda, A.M.-S. Effect of dietary seaweed extract supplementation on growth, feed utilization, hematological indices, and non-specific immunity of Nile Tilapia, Oreochromis niloticus challenged with Aeromonas hydrophila. J. Appl. Phycol. 2020, 32, 3467–3479. [Google Scholar] [CrossRef]
- El-Shenody, R.A.; Ashour, M.; Ghobara, M.M.E. Evaluating the chemical composition and antioxidant activity of three Egyptian seaweeds: Dictyota dichotoma, Turbinaria decurrens, and Laurencia obtusa. Braz. J. Food Technol. 2019, 22. [Google Scholar] [CrossRef]
- Elshobary, M.E.; El-Shenody, R.A.; Ashour, M.; Zabed, H.M.; Qi, X. Antimicrobial and antioxidant characterization of bioactive components from Chlorococcum minutum. Food Biosci. 2020, 35, 100567. [Google Scholar] [CrossRef]
- Abualnaja, K.M.; Alprol, A.E.; Abu-Saied, M.A.; Mansour, A.T.; Ashour, M. Studying the Adsorptive Behavior of Poly (Acrylonitrile- co -Styrene) and Carbon Nanotubes (Nanocomposites) Impregnated with Adsorbent Materials towards Methyl Orange Dye. Nanomaterials 2021, 11, 1144. [Google Scholar] [CrossRef]
- Abualnaja, K.M.; Alprol, A.E.; Ashour, M. Removing of Anionic Dye from Aqueous Solutions by Adsorption Using of Multiwalled Carbon Nanotubes and Poly (Acrylonitrile-styrene) Impregnated with Activated Carbon. Sustainability 2021, 13, 7077. [Google Scholar] [CrossRef]
- Abualnaja, K.M.; Alprol, A.E.; Ashour, M.; Mansour, A.T. Influencing Multi-Walled Carbon Nanotubes for the Removal of Ismate Violet 2R Dye from Wastewater: Isotherm, Kinetics, and Thermodynamic Studies. Appl. Sci. 2021, 11, 4786. [Google Scholar] [CrossRef]
- Ashour, M.; Mabrouk, M.; Abo-Taleb, H.A.; Sharawy, Z.Z.; Ayoub, H.F.; Van Doan, H.; Davies, S.J.; El-Haroun, E.; Goda, A.A. A liquid seaweed extract (TAM®) improves aqueous rearing environment, diversity of zooplankton community, whilst enhancing growth and immune response of Nile tilapia, Oreochromis niloticus, challenged by Aeromonas hydrophila. Aquaculture 2021, 543, 736915. [Google Scholar] [CrossRef]
- Magouz, F.I.; Essa, M.A.; Matter, M.; Tageldein Mansour, A.; Alkafafy, M.; Ashour, M. Population dynamics, fecundity and fatty acid composition of Oithona nana (Cyclopoida, Copepoda), fed on different diets. Animals 2021, 11, 1188. [Google Scholar] [CrossRef]
- Abo-Taleb, H.A.; Ashour, M.; Elokaby, M.A.; Mabrouk, M.M.; El-feky, M.M.M.; Abdelzaher, O.F.; Gaber, A.; Alsanie, W.F.; Mansour, A.T. Effect of a New Feed Daphnia magna (Straus, 1820), as a Fish Meal Substitute on Growth, Feed Utilization, Histological Status, and Economic Revenue of Grey Mullet, Mugil cephalus (Linnaeus 1758). Sustainability 2021, 13, 7093. [Google Scholar] [CrossRef]
- Abo-Taleb, H.A.; El-feky, M.M.M.; Azab, A.M.; Mabrouk, M.M.; Elokaby, M.A.; Ashour, M.; Mansour, A.T.; Abdelzaher, O.F.; Abualnaja, K.M.; Sallam, A.E. Growth Performance, Feed Utilization, Gut Integrity, and Economic Revenue of Grey Mullet, Mugil cephalus, Fed an Increasing Level of Dried Zooplankton Biomass Meal as Fishmeal Substitutions. Fishes 2021, 6, 38. [Google Scholar] [CrossRef]
- Magouz, F.I.; Essa, M.A.; Matter, M.; Mansour, A.T.; Gaber, A.; Ashour, M. Effect of different salinity levels on Copepoda (Oithona nana) population dynamics, production, and composition. Diversity 2021, 30, 190. [Google Scholar] [CrossRef]
- Abo-Taleb, H.A.; Zeina, A.F.; Ashour, M.; Mabrouk, M.M.; Sallam, A.E.; El-Feky, M.M. Isolation and cultivation of the freshwater amphipod Gammarus pulex (Linnaeus, 1758), with an evaluation of its chemical and nutritional content. Egypt. J. Aquat. Biol. Fish. 2020, 24, 69–82. [Google Scholar] [CrossRef]
- Heneash, A.M.M.; Alprol, A.E. Monitoring of Water Quality and Zooplankton Community in Presence of Different Dietary Levels of Commercial Wood Charcoal of Red Tilapia. J. Aquac. Res. Dev. 2020, 11, 1–6. [Google Scholar] [CrossRef]
- Zaki, M.A.; Ashour, M.; Heneash, A.M.M.; Mabrouk, M.M.; Alprol, A.E.; Khairy, H.M.; Nour, A.M.; Mansour, A.T.; Hassanien, H.A.; Gaber, A. Potential Applications of Native Cyanobacterium Isolate (Arthrospira platensis NIOF17/003) for Biodiesel Production and Utilization of Its Byproduct in Marine Rotifer (Brachionus plicatilis) Production. Sustainability 2021, 13, 1769. [Google Scholar] [CrossRef]
- Alprol, A.E.; Ashour, M.; Abualnaja, K.M.; Alhashmialameer, D.; Mansour, A.T.; Sharawy, Z.Z.; Abu-Saied, M.A.; Abomohra, A.E.-F. Potential Applications of Arthrospira platensis Lipid-Free Biomass in Bioremediation of Organic Dye from Industrial Textile Effluents and Its Influence on Marine Rotifer (Brachionus plicatilis). Materials 2021, 14, 4446. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.; Alprol, A.E.; Heneash, A.M.M.; Saleh, H.; Abualnaja, K.M.; Alhashmialameer, D.; Mansour, A.T. Ammonia Bioremediation from Aquaculture Wastewater Effluents Using Arthrospira platensis NIOF17/003: Impact of Biodiesel Residue and Potential of Ammonia-Loaded Biomass as Rotifer Feed. Materials 2021, 14, 5460. [Google Scholar] [CrossRef]
- Parsons, T.R. A Manual of Chemical & Biological Methods for Seawater Analysis; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 1483293394. [Google Scholar]
- Elhdad, A.M.A. Assessment of Surface Water Quality, Raw versus Treated, for Different Uses at Dakahlia Governorate, Egypt. Egypt. J. Chem. 2019, 62, 1117–1129. [Google Scholar] [CrossRef]
- Sargaonkar, A.; Deshpande, V. Development of an overall index of pollution for surface water based on a general classification scheme in Indian context. Environ. Monit. Assess. 2003, 89, 43–67. [Google Scholar] [CrossRef]
- El-Hamid, H.T.A.; Hegazy, T.A.; Ibrahim, M.S.; Khalid, M. Assessment of water quality of the Northern Delta Lakes, Egypt. J. Environ. Sci. 2017, 46, 21–34. [Google Scholar]
- Tiwari, T.N.; Manzoor, A. Water quality index for Indian rivers. In Ecology and Pollution of Indian River; Ashish Publishing House: New Delhi, India, 1988; pp. 271–286. [Google Scholar]
- Pielou, E.C. Ecological Diversity; John Wiley & Sons: New York, NY, USA, 1975; ISBN 0471689254. [Google Scholar]
- Wilhm, J.L.; Dorris, T.C. Biological Parameters for Water Quality Criteria. BioScience 1968, 18, 477–481. [Google Scholar] [CrossRef]
- Staub, R.; Appling, J.N.; Hotsteiler, A.M.; Hass, I.J. The effects of industrial wastes of Memphis and Shelby county on primary planktonic producers. BioScience 1970, 20, 905–912. [Google Scholar] [CrossRef]
- Gharib, S.M.; El-Sherif, Z.M.; Abdel-Halim, A.M.; Radwan, A.A. Phytoplankton and environmental variables as a water quality indicator for the beaches at Matrouh, south-eastern Mediterranean Sea, Egypt: An assessment. Oceanologia 2011, 53, 819–836. [Google Scholar] [CrossRef]
- Dango, E.A.; Ibrahim, M.S.; Hussein, N.R.; El Gammal, M.I.; Okbah, M.A. Spatial and temporal variations of phytoplankton communities and environmental conditions along the coastal area of Alexandria. Sci. Res. 2015, 3, 273–282. [Google Scholar] [CrossRef]
- Khairy, H.M.; Faragallah, H.M.; Hussein, N.R.; Dorgham, M.M. Environmental characteristics and nutritional level of chronically eutrophic bay on Alexandria Sea coast, Egypt. Indian J. Mar. Sci. 2014. Available online: http://www.niscair.res.in/jinfo/ijms/ijms-forthcoming-articles/BKP-IJMS-PR-Aug 2014/MS 2166 Edited.pdf (accessed on 30 October 2021).
- Fepa, A. Guidelines and Standards for Environmental Pollution Control in Nigeria; Federal Environmental Protection Agency (FEPA): Lagos, Nigeria, 1991.
- Tadros, H.R.Z.; Ibrahim, G.H.; El Zokm, G.M. Multivariate analysis to investigate the organization of physicochemical Multivariate analysis to investigate the organization of physicochemical parameters in the Eastern Harbour Alexandria, Egypt. Int. J. Contemp. Appl. Sci. 2016, 3, 33–45. [Google Scholar]
- Adenan, N.S.; Yusoff, F.M.; Shariff, M. Effect of salinity and temperature on the growth of diatoms and green algae. J. Fish. Aquat. Sci. 2013, 8, 397. [Google Scholar] [CrossRef]
- Oczkowski, A.; Nixon, S. Increasing nutrient concentrations and the rise and fall of a coastal fishery; a review of data from the Nile Delta, Egypt. Estuar. Coast. Shelf Sci. 2008, 77, 309–319. [Google Scholar] [CrossRef]
- Seroka, G. The Relationship between Dissolved Oxygen, Nitrate and Phosphate Concentrations and Chlorophyll-a Concentration in Rhode River, a Subestuary of Chesapeake Bay; Thomas Jefferson High School of Science and Technology: Alexandria, VA, USA, 2004. [Google Scholar]
- James, B.K.; Adejare, L.I. Nutrients and phytoplankton production dynamics of a tropical harbor in relation to water quality indices. J. Am. Sci. 2010, 6, 261–275. [Google Scholar]
- Redfield, A.C. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 205–221. [Google Scholar]
- Krom, M.D.; Kress, N.; Brenner, S.; Gordon, L.I. Phosphorus limitation of primary productivity in the eastern Mediterranean Sea. Limnol. Oceanogr. 1991, 36, 424–432. [Google Scholar] [CrossRef]
- Bethoux, J.P.; Morin, P.; Madec, C.; Gentili, B. Phosphorus and nitrogen behaviour in the Mediterranean Sea. Deep. Sea Res. Part A Oceanogr. Res. Pap. 1992, 39, 1641–1654. [Google Scholar] [CrossRef]
- Khairy, H.M.; Hussein, N.R.; Faragallah, H.M.; Dorgham, M.M. The phytoplankton communities in two eutrophic areas on the Alexandria coast, Egypt. Rev. Biol. Mar. Oceanogr. 2014, 49, 267–277. [Google Scholar] [CrossRef]
- El-Sherif, Z.M.; Nassar, M.Z.; Fahmy, M.A. Phytoplankton distribution in the southeastern Mediterranean Sea (Egyptian waters) in summer and winter 2005. Egypt. J. Aquat. Res. 2010, 36, 609–621. [Google Scholar]
- Turkoglu, M.; Koray, T. Phytoplankton species’ succession and nutrients in the southern Black Sea (Bay of Sinop). Turk. J. Bot. 2002, 26, 235–252. [Google Scholar]
- Lundholm, N.; Andersen, P.; Jørgensen, K.; Thorbjørnsen, B.R.; Cembella, A.; Krock, B. Domoic acid in Danish blue mussels due to a bloom of Pseudo-nitzschia seriata. Harmful Algae News 2005, 29, 8–10. [Google Scholar]
- Liefer, J.D.; MacIntyre, H.L.; Novoveská, L.; Smith, W.L.; Dorsey, C.P. Temporal and spatial variability in Pseudo-nitzschia spp. in Alabama coastal waters: A “hot spot” linked to submarine groundwater discharge? Harmful Algae 2009, 8, 706–714. [Google Scholar] [CrossRef]
- Develİ, E.E.; Kideyş, A.E. Weekly variations in phytoplankton structure of a harbour in Mersin Bay (north-eastern Mediterranean). Turk. J. Bot. 2000, 24, 13–24. [Google Scholar]
- Ouyang, Z.; Wen, X.; Geng, Y.; Mei, H.; Hu, H.; Zhang, G.; Li, Y. The effects of light intensities, temperatures, pH and salinities on photosynthesis of Chlorella. J. Wuhan Bot. Res. 2010, 28, 49–55. [Google Scholar] [CrossRef]
- Mohanty, A.K.; Satpathy, K.K.; Sahu, G.; Hussain, K.J.; Prasad, M.V.R.; Sarkar, S.K. Bloom of Trichodesmium erythraeum (Ehr.) and its impact on water quality and plankton community structure in the coastal waters of southeast coast of India. Indian J. Mar. Sci. 2010, 39, 323–333. [Google Scholar]
- Asha, P.S.; Diwakar, K.; Sivanesh, H.; Kaladharan, P. Bloom of micro alga Chlorella marina (Butcher) in the Karapad lagoon, Gulf of Mannar, southeast coast of India. J. Mar. Biol. Assoc. India 2015, 57, 31–35. [Google Scholar] [CrossRef]
- Olli, K.; Heiskanen, A.-S.; Seppälä, J. Development and fate of Eutreptiella gymnastica bloom in nutrient-enriched enclosures in the coastal Baltic Sea. J. Plankton Res. 1996, 18, 1587–1604. [Google Scholar] [CrossRef][Green Version]
- Khairy, H.M.; Gharib, S.M. Factors regulating composition and abundance of phytoplankton in El Dekhaila Harbor, South-Eastern Mediterranean Sea, Egypt. Asian J. Biol. Sci. 2017, 10, 27–37. [Google Scholar] [CrossRef][Green Version]
- Sánchez, E.; Colmenarejo, M.F.; Vicente, J.; Rubio, A.; García, M.G.; Travieso, L.; Borja, R. Use of the water quality index and dissolved oxygen deficit as simple indicators of watersheds pollution. Ecol. Indic. 2007, 7, 315–328. [Google Scholar] [CrossRef]
- Turkoglu, M. Winter bloom and ecological behaviors of coccolithophore Emiliania huxleyi (Lohmann) Hay & Mohler, 1967 in the Dardanelles (Turkish Straits System). Hydrol. Res. 2010, 41, 104–114. [Google Scholar]
- Turkoglu, M. Temporal variations of surface phytoplankton, nutrients and chlorophyll-a in the Dardanelles (Turkish Straits System): A coastal station sample in weekly time intervals. Turk. J. Biol. 2010, 34, 319–333. [Google Scholar]



{kind=link}
{kind=link}
| No | Sampling Site | N | E |
|---|---|---|---|
| 1 | El-Montaza | 31 17 21 | 30 00 42 |
| 2 | El-Mandra | 31 16 51 | 30 00 36 |
| 3 | El-Asafra | 31 16 25 | 30 00 01 |
| 4 | Miamy | 31 16 12 | 29 59 29 |
| 5 | Sidi Bisher | 31 15 45 | 29 59 03 |
| 6 | Al-saraya | 31 13 20 | 29 55 51 |
| 7 | El-Shatby | 31 12 38 | 29 54 29 |
| 8 | El-Anfoshy | 31 12 46 | 29 52 54 |
| Beaches * | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| Parameter | (Min–Max) | (Min–Max) | (Min–Max) | (Min–Max) | (Min–Max) | (Min–Max) | (Min–Max) | (Min–Max) |
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | |
| Temperature (°C) | 18.6–27.0 | 18.6–27.0 | 18.7–27.1 | 18.7–27.3 | 18.7–27.2 | 18.8–27.2 | 18.8–27.2 | 18.8–27.3 |
| 23 ± 3.31 | 23.1 ± 3.25 | 23.2 ± 3.21 | 23.2 ± 3.28 | 23.2 ± 3.26 | 23.3 ± 3.29 | 23.3 ± 3.28 | 23.3 ± 3.3 | |
| pH | 7.23–8.20 | 7.54 −8.20 | 7.67–8.30 | 7.45–8.54 | 7.42–8.34 | 7.26–8.32 | 7.43–8.34 | 7.38–8.44 |
| 7.88 ± 0.27 | 7.92 ± 0.17 | 7.94 ± 0.24 | 7.93 ± 0.25 | 7.98 ± 0.27 | 7.96 ± 0.28 | 7.97 ± 0.26 | 8.03 ± 0.32 | |
| Salinity (ppt) | 34.80–38.8 | 33.4–38.2 | 33.60 −38.3 | 32.5–38.1 | 32.40–38.1 | 32.3–38.2 | 32.0–38.1 | 27.50–38.1 |
| 37.53 ± 1.00 | 35.43 ± 1.55 | 35.57 ± 1.54 | 35.27 ± 1.69 | 35.24 ± 1.69 | 35.06 ± 1.70 | 35.04 ± 1.75 | 34.31 ± 2.75 | |
| DO (mg L−1) | 4.23–5.80 | 4.12–6.40 | 4.65–6.23 | 4.80–6.40 | 4.87–6.10 | 4.12–6.10 | 4.54–6.40 | 4.10–7.50 |
| 5.00 ± 0.62 | 5.60 ± 0.66 | 5.42 ± 0.45 | 5.49 ± 0.41 | 5.59 ± 0.36 | 5.42 ± 0.50 | 5.67 ± 0.48 | 6.39 ± 1.06 | |
| NO2-N (µg L−1) | 0.01–0.93 | 0.01–0.95 | 0.04 −1.21 | 0.06–1.76 | 0.03–1.98 | 0.07–1.87 | 0.28–1.98 | 0.42–1.50 |
| 0.23 ± 0.29 | 0.26 ± 0.28 | 0.32 ± 0.34 | 0.49 ± 0.47 | 0.51 ± 0.52 | 0.61 ± 0.48 | 0.71 ± 0.45 | 0.74 ± 0.30 | |
| NO3-N (µg L−1) | 2.36–9.04 | 1.54–10.54 | 1.98–10.65 | 2.65–16.76 | 3.43–19.76 | 3.54–33.21 | 2.65–23.76 | 3.54–13.78 |
| 3.84 ± 2.22 | 4.06 ± 2.52 | 4.23 ± 2.37 | 5.40 ± 3.78 | 6.05 ± 4.43 | 7.71 ± 8.17 | 7.79 ± 5.65 | 6.79 ± 2.59 | |
| NH4 (µg L−1) | 0.12–7.55 | 0.04–6.34 | 0.23–5.87 | 0.23–7.34 | 0.04–7.54 | 0.10–7.39 | 0.04–9.45 | 0.26–7.98 |
| 2.48 ± 2.85 | 2.19 ± 2.31 | 2.51 ± 2.28 | 2.74 ± 2.92 | 2.89 ± 2.89 | 2.70 ± 2.75 | 3.54 ± 3.82 | 3.23 ± 3.13 | |
| PO4-P (µg L−1) | 0.02–0.34 | 0.01–0.07 | 0.03–0.45 | 0.02–0.93 | 0.02–0.72 | 0.03–1.05 | 0.03–1.54 | 0.02–1.05 |
| 0.09 ± 0.09 | 0.02 ± 0.02 | 0.15 ± 0.16 | 0.28 ± 0.40 | 0.20 ± 0.23 | 0.30 ± 0.34 | 0.42 ± 0.54 | 0.29 ± 0.37 | |
| SiO2 (µg L−1) | 9.08–29.53 | 5.91–25.21 | 6.98–16.09 | 0.67–23.41 | 6.14–22.05 | 4.05–23.12 | 3.98–25.67 | 3.57–25.70 |
| 16.29 ± 6.03 | 14.23 ± 6.48 | 11.04 ± 3.37 | 11.58 ± 5.88 | 14.19 ± 4.88 | 14.25 ± 6.38 | 15.28 ± 5.69 | 13.86 ± 7.21 | |
| Chl-a (µg L−1) | 0.16–4.74 | 0.12–4.65 | 0.22–4.60 | 0.12 −12.31 | 0.47–5.65 | 0.16–4.12 | 0.13–5.73 | 0.18–5.23 |
| 1.57 ± 1.33 | 2.16 ± 1.34 | 2.37 ± 1.41 | 3.19 ± 3.34 | 2.64 ± 1.59 | 2.52 ± 1.41 | 2.76 ± 1.76 | 2.59 ± 1.59 | |
| WQI | 66–90 | 82–92 | 75–90 | 70–89 | 69–89 | 69–86 | 64–90 | 68–91 |
| 82.65 ± 6.87 | 86.52 ± 2.53 | 83.03 ± 4.73 | 80.66 ± 5.12 | 81.19 ± 6.09 | 78.68 ± 5.83 | 79.30 ± 8.76 | 80.53 ± 7.81 |
| Class | Richness (Number of Taxa) |
|---|---|
| Bacillariophyceae | 136 |
| Dinoflagellates | 36 |
| Chlorophyceae | 28 |
| Cyanophyceae | 19 |
| Euglenophyta | 6 |
| Rhodophyta | 1 |
| Raphidophyceae | 1 |
| Silicoflagellates | 1 |
| Total phytoplankton richness | 228 |
| Beach 1 * | % | Beach 2 | % | Beach 3 | % | Beach 4 | % |
| Pseudo-nitzschia delicatissima | 12.27 | Pseudo-nitzschia delicatissima | 15.44 | Skeletonema costatum | 23.30 | Chlorella marina | 44.44 |
| Skeletonema costatum | 11.13 | Skeletonema costatum | 11.16 | Leptocylindrus danicus | 14.47 | Skeletonema costatum | 19.25 |
| Navicula tripunctata | 11.10 | Akashiwo sanguinea | 5.12 | Pseudo-nitzschia delicatissima | 12.89 | Pseudo-nitzschia delicatissima | 5.78 |
| Prorocentrum triestinum | 10.31 | Cyclotella kützingiana | 5.06 | Chlorella marina | 7.97 | Karenia mikimotoi | 3.67 |
| Cyclotella kützingiana | 7.84 | Cyclotella comta | 4.60 | Thalassiosira decipiens | 3.89 | Protoperidinium conicum | 2.82 |
| Cyclotella comta | 7.53 | Prorocentrum triestinum | 3.82 | Cyclotella kützingiana | 3.00 | Chaetoceros constrictus | 2.28 |
| Licmophora ehrenbergii | 4.45 | Navicula tripunctata | 3.59 | Cyclotella comta | 2.66 | Thalassiosira decipiens | 2.19 |
| Crucigenia rectangularis | 3.55 | Chlorella marina | 2.97 | Prorocentrum triestinum | 2.59 | Cyclotella kützingiana | 1.90 |
| Prorocentrum micans | 3.42 | Crucigenia rectangularis | 2.96 | Chaetoceros decipiens | 2.02 | Cyclotella comta | 1.77 |
| Leptocylindrus danicus | 2.84 | Nitzschia longissima | 2.16 | Navicula tripunctata | 1.95 | Eutreptiella braarudii | 1.62 |
| Beach 5 | % | Beach 6 | % | Beach 7 | % | Beach 8 | % |
| Chlorella marina | 29.72 | Chlorella marina | 38.06 | Chlorella marina | 79.03 | Chlorella marina | 1.62 |
| Pseudo-nitzschia delicatissima | 10.94 | Pseudo-nitzschia delicatissima | 13.08 | Pseudo-nitzschia delicatissima | 7.33 | Pseudo-nitzschia delicatissima | 58.67 |
| Skeletonema costatum | 9.68 | Skeletonema costatum | 10.28 | Eutreptiella braarudii | 1.52 | Eutreptiella braarudii | 5.95 |
| Thalassiosira decipiens | 6.77 | Cyclotella kützingiana | 5.20 | Skeletonema costatum | 1,21 | Karenia mikimotoi | 5.33 |
| Leptocylindrus danicus | 5.28 | Leptocylindrus danicus | 4.64 | Protoperidinium pellucidum | 1.07 | Skeletonema costatum | 4.67 |
| Karenia mikimotoi | 3.91 | Gonyaulax spinifera | 3.67 | Chaetoceros constrictus | 0.89 | Cyclotella kützingiana | 4.47 |
| Chaetoceros constrictus | 3.67 | Nitzschia longissima | 2.29 | Cyclotella kützingiana | 0.76 | Leptocylindrus danicus | 3.22 |
| Cyclotella comta | 3.47 | Protoperidinium cerasus | 1.89 | Exuviaella marina | 0.76 | Euglena granulata | 2.60 |
| Prorocentrum triestinum | 2.43 | Prorocentrum triestinum | 1.84 | Leptocylindrus danicus | 0.74 | Prorocentrum lima | 1.39 |
| Protoperidinium cerasus | 2.33 | Gyrodinium spirale | 1.45 | Pseudo-nitzschia seriata | 0.36 | Eutreptiella gymnastica | 1.28 |
| Nutrients Salts | Alexandria Beaches | Matrouh Beaches | Touristic Village Marakia |
|---|---|---|---|
| Present Study | Gharib et al. [41] | Shams El-Din and Abdel Halim [12] | |
| Phosphate | 0.01–1.54 | 0.01–7.30 | 0.00–2.50 |
| Silicate | 0.67–29.53 | 0.20–4.49 | 0.11–3.08 |
| Nitrate | 1.54–33.21 | 0.13–5.10 | 0.10–4.01 |
| Nitrite | 0.01–1.98 | 0.01–0.30 | 0.00–0.30 |
| Ammonia | 0.04–9.45 | 0.18–16.83 | 0.14–24.25 |
| Group/Source | Bacillariophyta | Dinoflagellates | Total Phytoplankton Species | |
|---|---|---|---|---|
| Alexandria Beaches | Present Study | 136 | 36 | 228 |
| Matrouh beaches | [41] | 120 | 52 | 203 |
| Alexandria coast | [42] | 85 | 31 | 153 |
| Eastern Harbor | [43] | 100 | 32 | 162 |
| Kayet Bay | 64 | 21 | 110 | |
| El Mohandessin village | [12] | 61 | 11 | 90 |
| Marakia village | 56 | 12 | 83 | |
| Marabella village | 34 | 9 | 54 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alprol, A.E.; Ashour, M.; Mansour, A.T.; Alzahrani, O.M.; Mahmoud, S.F.; Gharib, S.M. Assessment of Water Quality and Phytoplankton Structure of Eight Alexandria Beaches, Southeastern Mediterranean Sea, Egypt. J. Mar. Sci. Eng. 2021, 9, 1328. https://doi.org/10.3390/jmse9121328
Alprol AE, Ashour M, Mansour AT, Alzahrani OM, Mahmoud SF, Gharib SM. Assessment of Water Quality and Phytoplankton Structure of Eight Alexandria Beaches, Southeastern Mediterranean Sea, Egypt. Journal of Marine Science and Engineering. 2021; 9(12):1328. https://doi.org/10.3390/jmse9121328
Chicago/Turabian StyleAlprol, Ahmed E., Mohamed Ashour, Abdallah Tageldein Mansour, Othman M. Alzahrani, Samy F. Mahmoud, and Samiha M. Gharib. 2021. "Assessment of Water Quality and Phytoplankton Structure of Eight Alexandria Beaches, Southeastern Mediterranean Sea, Egypt" Journal of Marine Science and Engineering 9, no. 12: 1328. https://doi.org/10.3390/jmse9121328
APA StyleAlprol, A. E., Ashour, M., Mansour, A. T., Alzahrani, O. M., Mahmoud, S. F., & Gharib, S. M. (2021). Assessment of Water Quality and Phytoplankton Structure of Eight Alexandria Beaches, Southeastern Mediterranean Sea, Egypt. Journal of Marine Science and Engineering, 9(12), 1328. https://doi.org/10.3390/jmse9121328