1. Introduction
Artificial Reefs (ARs) started during the second half of the past century (in Europe between 1960–1970) and their deployments were intensified over the five last decades. They have been used around the world to create, protect or restore a rich and diverse ecosystem [
1,
2]. Their development has been intensified to elicit several ecological responses, i.e., settlement and colonization processes, in relation to benthic community succession in response to biotic and abiotic factors [
2,
3]. An AR can produce up to 100 times more biomass than the surrounding sandy-muddy bottom [
4]. Studies of ARs show an ecological succession in the mechanism of colonization; the first colonisers of ARs are benthic micro-organisms ranging from biofilms to microphytobenthos, followed by macroalgae as well as sessile and motile faunas [
5,
6,
7,
8].
Today, there are many submerged ARs of various shapes and materials [
1,
2,
3,
9,
10,
11]. These structures are submerged for diverse reasons such as improving fish production, as well as for the protection, conservation and restoration of habitats, or in relation to economic activities including fisheries, stock management, aquaculture, renewable energy, research and development of new materials and structures [
10,
12,
13,
14]. Historically, ARs have been deployed with the aim of increasing local fisheries production or protecting the coastal zone from the effects of trawling [
15]. Furthermore, recent research on ARs tends to focus on variations in the community structure or composition of biota, suggesting that the purpose of AR research has shifted from improving fishery resources to restoring marine ecosystems and developing marine structures to increase biodiversity, taking into account different scopes, such as biodeterioration of archaeological remains, protection against trawling, and bioremediation role [
2,
10,
12,
16,
17,
18,
19,
20]. However, the presence of such artificial structures in the marine environment modifies the seabed by adding areas of hard substrate to soft-bottom habitats [
10,
16,
17,
21].
One of the scientific questions concerning ARs is to establish an inventory of the species colonizing reefs, i.e., the taxonomic richness, the temporal succession of species and taxa during the immersion phase and the growth of existing species and taxa [
1,
10,
22,
23]. Most ARs are constructed to increase the production of fishes and invertebrates of commercial interest. However, it is important to monitor species colonizing the AR community since these species control the local diversity and could be potential prey for other species including commercial crabs and fishes [
24]. Moreover, the composition and heterogeneous nature, lithology, mineralogy, texture and porosity of the immersed artificial substrate play important roles during the early phases of colonization by fixed organisms [
1,
10,
22]. Rough textures induce greater microhabitat diversity than smooth surfaces [
15,
22]. Furthermore, the settlement, colonisation and succession of organisms on the surface of ARs are favoured by the water retention properties of the porous artificial reef material mainly in intertidal zones, while crevices and pits protect small organisms from predators [
15,
25]. Many motile taxa are also associated with the fouling community, which increases the attractiveness of the reef for predators [
25]. However, the nature of the substrate appears to be a determining factor in the abundance and species richness of colonizing organisms [
26]. Nevertheless, most of the studies of ARs consider only the taxa composition and succession of the main groups of macroalgae or macrofauna, which are mainly sessile organisms [
15,
16,
27,
28,
29]. Very few studies take into account all the species colonizing ARs [
30,
31]. Moreover, most of the studies were deployed over a short period, frequently one year [
15,
28,
29], and only few ARs have been studied over the medium term (2–3 years) [
27], or the long term (>3 years) [
16,
30,
31]. Some studies focused on the long-term changes in polychaetes [
18,
32] or molluscs [
33], while others compared the fauna colonizing ARs with natural rocky reefs [
10,
13,
16,
17,
31,
32,
33,
34,
35]. Studies have also been undertaken to compare the colonization of Non-Indigenous Species (NIS) on these two types of substrate [
36], or the role of ARs in the colonization by NIS [
37]. In these studies, species identification was mainly carried out by divers and photographers and concerned only the megafauna and fish, while only few studies have identified the small sessile and motile macrofauna.
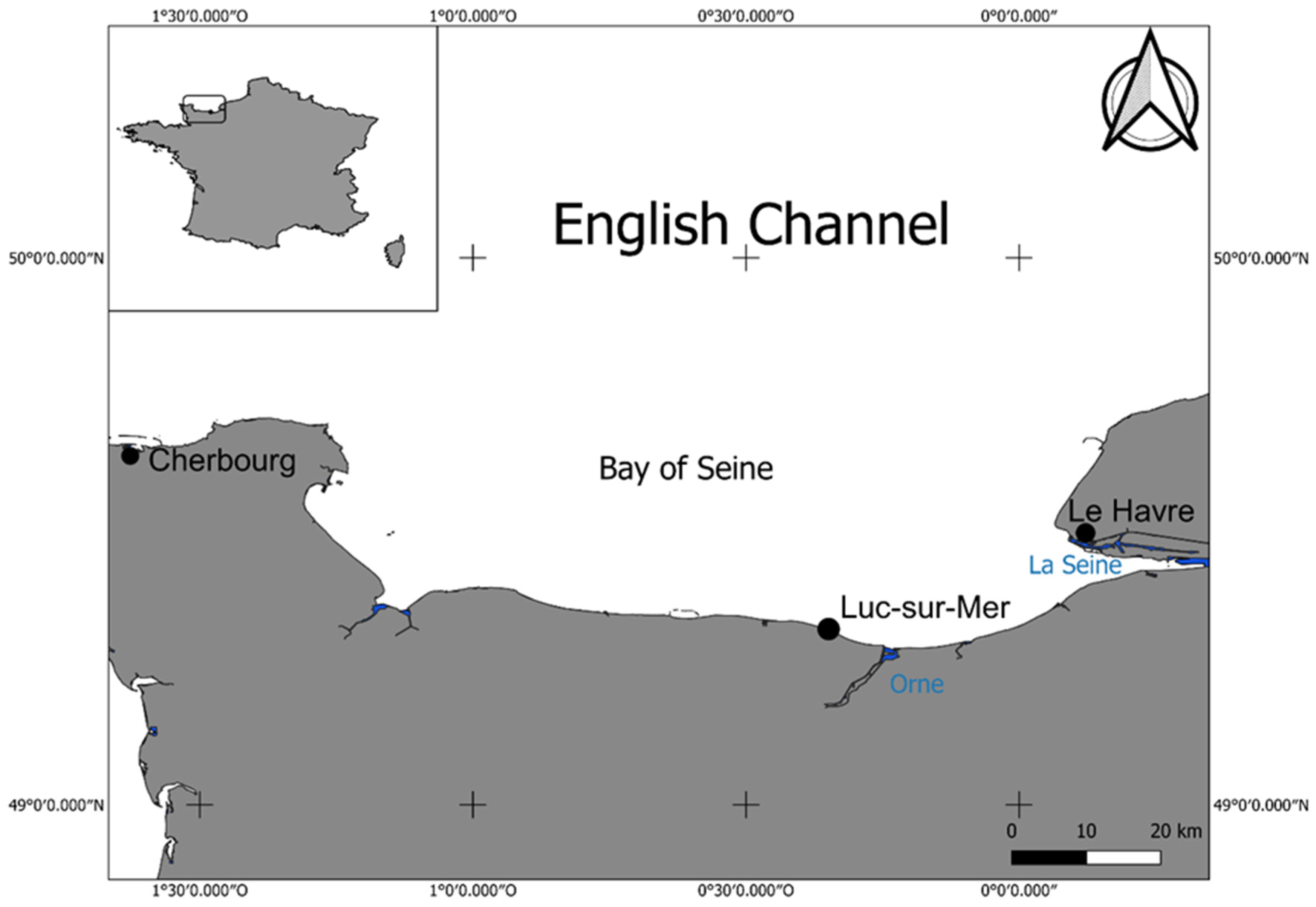
ARs have been deployed along the coast of metropolitan France since 1968 with the chief objective of enhancing the success and continuity of artisanal fishing mainly in the Mediterranean Sea [
3,
38]. Most of these ARs have been immerged recently—since 2000—and have been emplaced along the Atlantic coast, but only one AR has been deployed in the eastern part of the English Channel at Etretat [
39]. Fishermen opposed to the emplacement of ARs argued that numerous wrecks in the English Channel could be considered to act as ARs. These wrecks mainly resulted from Operation Overlord in Normandy in 1944. This may explain the low number of ARs currently emplaced in the English Channel.
At present, with the aim of maintaining or increasing biodiversity in coastal ecosystems including harbour basins, a new perception of ARs is spreading among the scientific community and users of the English Channel [
11,
39].
In the framework of the RECIF INTERREG project between France and the United Kingdom, concrete blocks have been deployed on the intertidal zone of the Bay of Seine (eastern part of the English Channel) since March 2014 and monitored each year at the beginning of the spring until April 2018. The objectives of our study are the first time for the English Channel, as an example of a megatidal sea with a large intertidal zone: (1) to provide a complete inventory of sessile and motile invertebrates colonizing artificial hard-bottom substrates, while most of the studies in such AR take into account only the large sessile species (megafauna), and (2) to describe the stages of colonization during the four-year study, and the succession of organisms on these blocks over the four years of the experiment, while most of the studies of AR colonization do not surpass one year.
4. Discussion
Our experiment illustrates the colonization of concreted blocks located in the intertidal zone of a megatidal sea ‘La Manche- the English Channel’ during four years. To our knowledge, this experiment is unique at the scale of the north-eastern Atlantic; however, the results are based on a small number of replicates (two concrete blocks per sampling date) and should be interpreted with caution. Moreover, our study considers all the identification of sessile and motile taxa including small organisms mainly at the species level. This identification protocol is only rarely applied since numerous AR studies are concerned solely with mega-zoological groups mainly composed of sessile organisms such as sponges, cnidarians, hydrozoans, mussels, etc., or motile fauna including decapods and fishes [
30,
31]. However, the sampling design of our experiment with the removal of blocks at low tide prevents the capture of fishes; hence, our study is focused on invertebrate macrofauna.
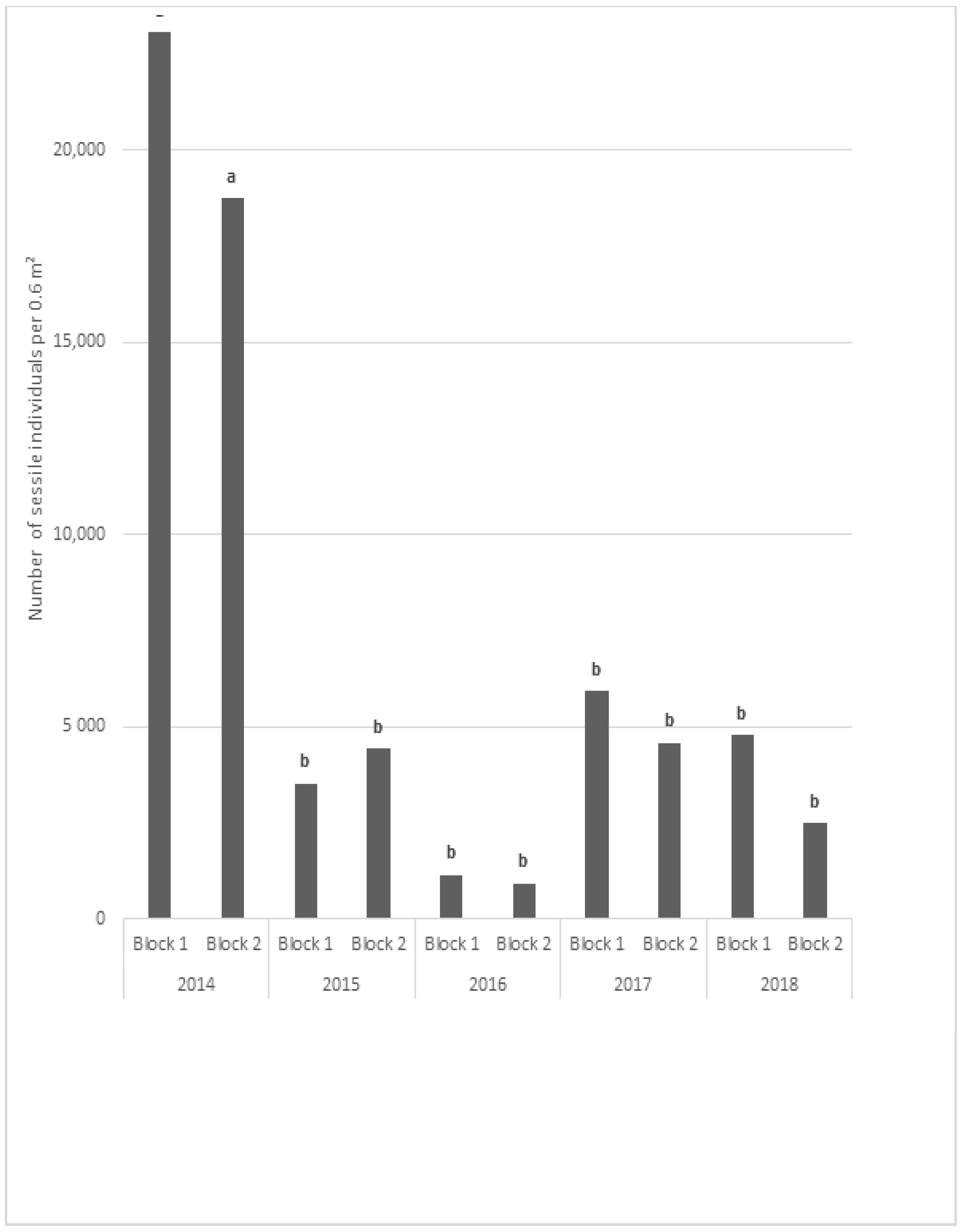
During the first year of the experiment, the main colonizing organisms were barnacles, hydrozoans, mussels, tunicates, and sessile annelids [
23]. Moreover, a biological succession is observed over time: a first assemblage mainly dominated by barnacles then a richer assemblage with the dominance of ascidians.
It is noteworthy that during the period of the study (2014–2018) there were no severe cold winters or warm summers. Neither there were there any floods of the Seine or Orne rivers which could have influenced the Luc-sur-Mer area during years with strong precipitation and high freshwater input (see:
https://www.somlit.fr/luc-sur-mer/ (accessed on: 18 October 2021)). At the SOMLIT (Service d’Observation en Milieu Littoral) Station at Luc-sur-Mer near the block immersion site, the sea water temperature at high tide varied from a minimum of 7 °C in winter (February-March) to a maximum of 21 °C at the end of the summer (September), and the salinity ranged from a minimum of 33 (November to March depending on the year) to a maximum of 34.5 at the end of the summer in September. Thus, normal seasonality occurred during our block study, and no environmental event needs to be evoked to interpret the long-term colonization of such ARs.
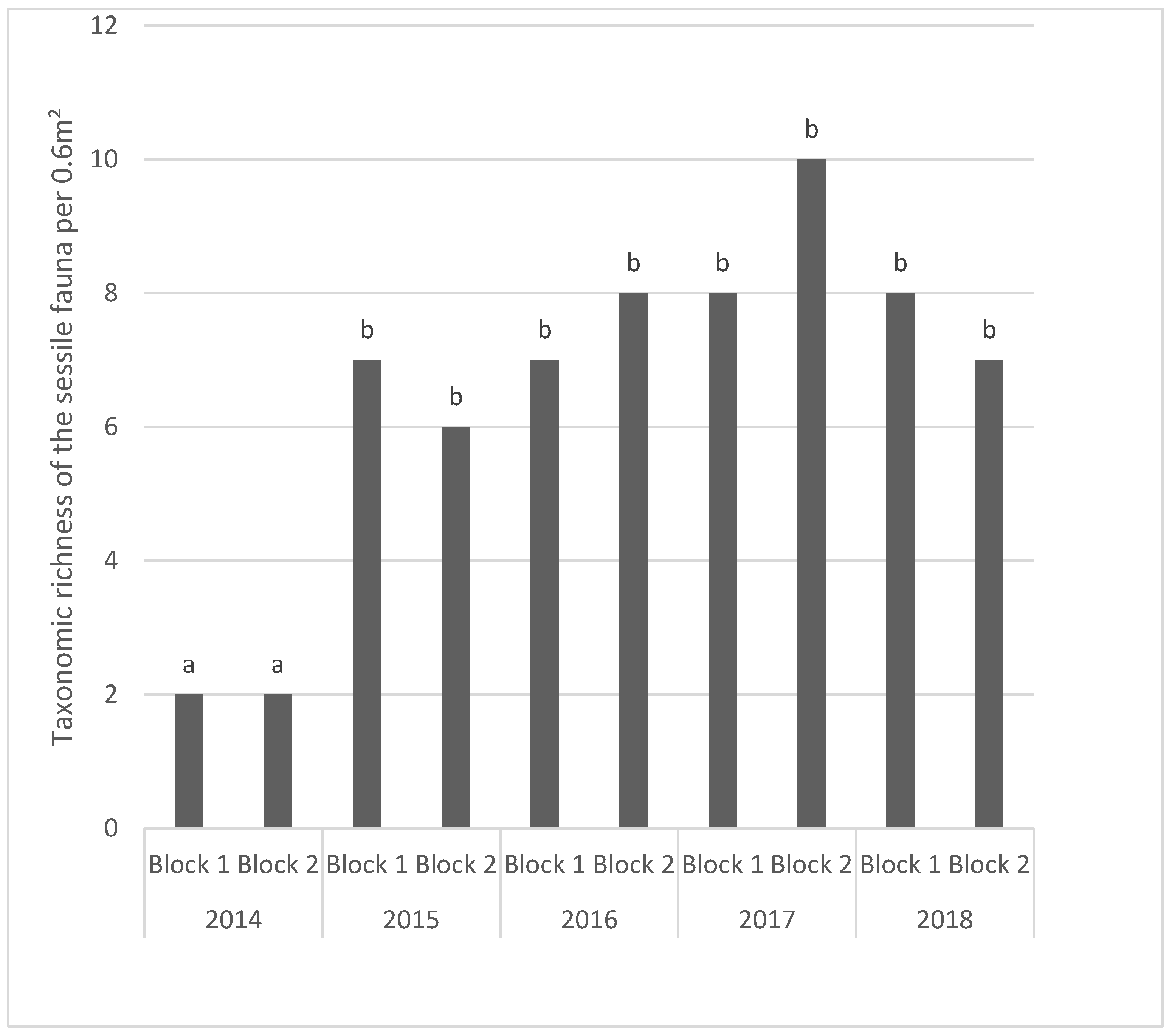
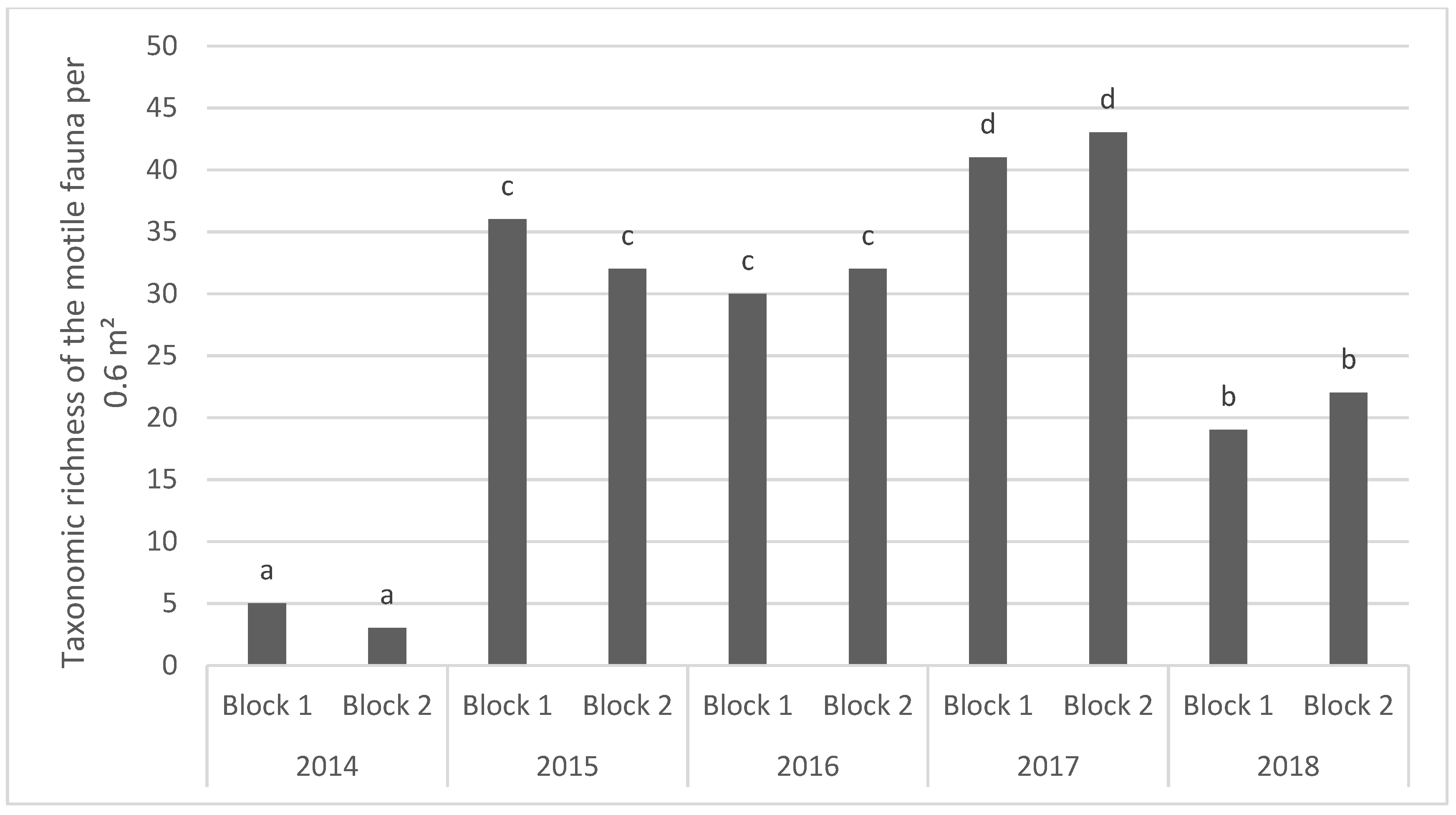
A total of 84 taxa were identified during this study. This high Taxonomic Richness is due to the effort for identification of the taxa, often at species level, while considering both the sessile and motile fauna. Sessile taxa make up 15% of the recorded fauna, while the motile taxa formed 85% of the fauna with numerous small organisms (
Appendix A). The most highly diversified zoological phylum is the Arthropoda (32 taxa; 38% of the fauna), then the Annelida (26; 31%), Mollusca (10; 12%), Chordata (5; 6%) and other groups (11; 13%). This taxonomic richness is on the same order that accounts for a geotextile artificial surf reef immerged at 2 m depth in the Poole Bay in the centre of the English coast in the English Channel and study during four years: 108 taxa ([
10]. Due to the sampling protocols, i.e., a combination of photography, underwater video using scuba diving and a mini-ROV (Remotely Operated Vehicle), and Baited Remote Underwater Video, and image analysis the number of sessile invertebrate (67 taxa, 66%) surpasses the number of motile fauna (41 taxa, 34%), which is different to the blocks of the intertidal zone of the Bay of Seine.
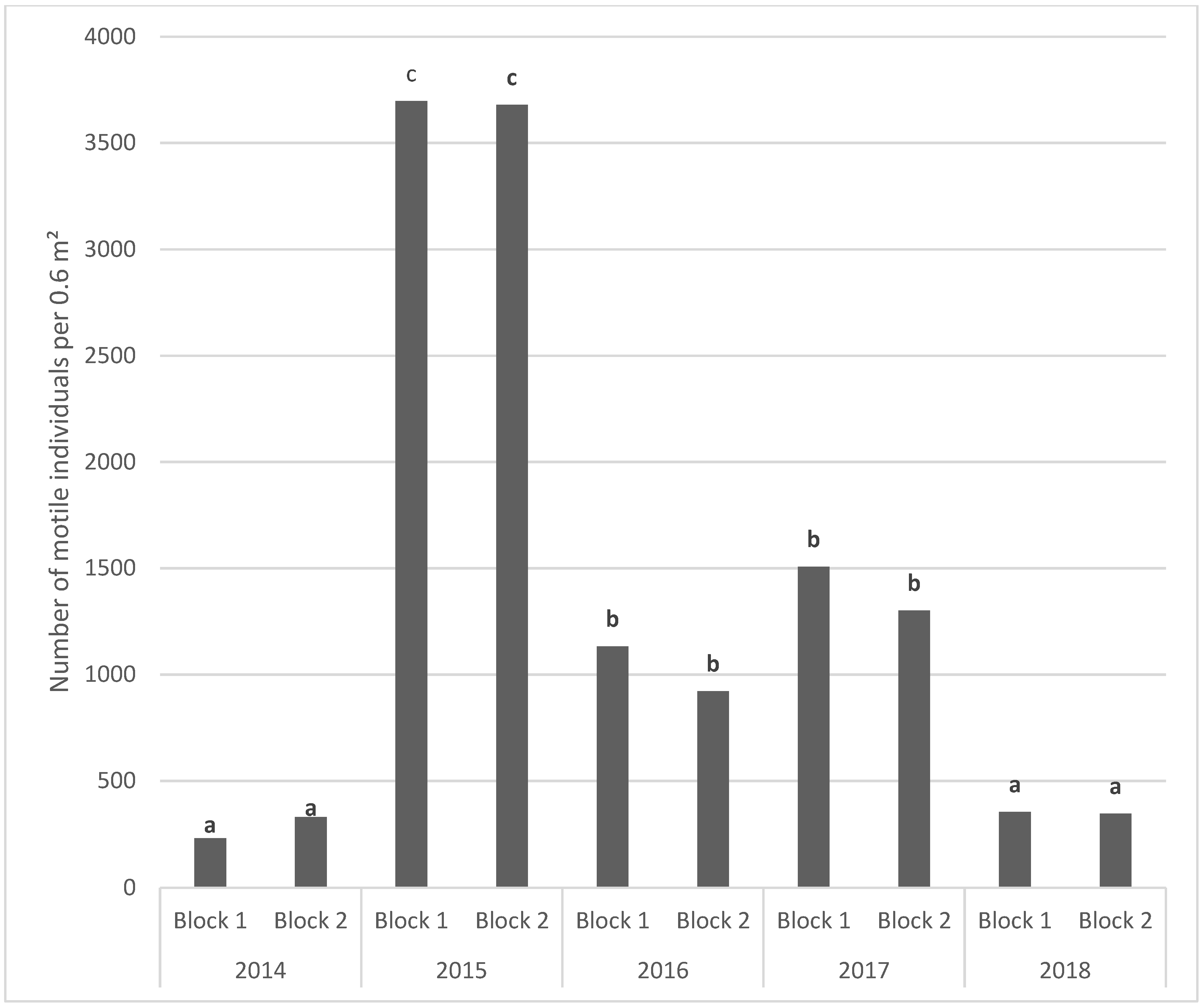
One month after the immersion, only seven taxa were found on the blocks. The colonisation of the blocks was rapid and one year after the beginning of the deployment of the blocks (in April 2015) the TR of the fauna reached 52. The TR reached a maximum in 2017 with 64 taxa observed on the two sampled blocks, decreasing to 45 in 2016 and only 37 at the end of the study in 2018. Our results illustrate that the colonization by sessile and motile species was relatively rapid. One year after the immersion, the TR was established at around 50. This was followed by a relatively stability of the TR, even though the blocks sampled in 2015 showed a significant decrease.
The number of non-indigenous species remained low both for sessile and motile fauna throughout the study, while only two sessile species and two motile taxa were recorded. This new artificial structure deployed in the intertidal zone of the Bay of Seine does not favour colonization by numerous pioneers NIS. This contrasts with the situation observed in other ARs where new habitats are found to facilitate biological invasions [
37]; but apart from the amphipod
Aoroides, the other NIS show very high abundances on blocks during the experiment.
Only one female of the amphipod
Aoroides was recorded in 2017; two species of this Pacific genus,
Aoroides longimerus Ren and Zheng, 1996 and
A. semicurvatus Ariyama, 2004 were recorded in 2019 for the first time along the Normandy coast of the English Channel associated with the brown macro-algae
Sargassum muticum as well as occurring on harbour pontoons [
45]. Both
A. semicurvatus and
A. longimerus show low abundances and were absent during a 2015 study carried out in Normandy marinas on associated species and the extent of biofouling “reefs” formed by the NIS polychaete
Ficopomatus enigmaticus (Fauvel, 1923) [
46]. Their introduction into Normandy waters, including the intertidal zone of Luc-sur-Mer, is probably recent.
Two NIS of the genus
Monocorophium colonized the blocks, attaining a maximum abundance only one year after the beginning of the experiment (3200 ind·m
−2 in 2015). Their abundance decreased rapidly thereafter and these taxa were no longer among the ten-dominant species in 2018 (
Table 5).
Monocorophium acherusicum (Costa, 1853) had been recorded at the beginning of the 20th century along the Brittany coast and identified in 1977 in Normandy in the Le Havre harbour basin [
47].
Monocorophium sextonae (Crawford, 1937) was described from New Zealand and recorded in European waters at the middle of the 20th century and recorded in 1976 in the Bay of Seine. These two
Monocorophium species are difficult to distinguish from each other; they are sympatric species with very high abundance and only mature males can be accurately identified. Both species were also reported in very high abundances in Normandy marinas during a two-year experiment of colonization of plates (unpublished data).
M. sextonae was reported in Normandy harbours by [
46].
The orange-tipped sea squirt
Corella eumyota (Traustedt, 1882) is a solitary tunicate native to the Southern Hemisphere and has been known in Normandy waters since 2007 in the Havre harbour basins and recorded along the Luc-sur-Mer intertidal zone in 2013 [
47]. It is known to be invasive and shows abundances higher than 680 ind·m
−2 in 2015; although its abundance subsequently decreased, the species remained among the most dominant sessile species on the blocks (
Table 4). The other tunicate
Perophora japonica (Oka, 1927) is a colonial sea squirt native to the North Indo-Pacific realm. It has colonized several other parts of the world including the south coast of England, France, the Netherlands and the west coast of the United States [
48], and was reported since 1982 in Normandy waters and later observed in 2013 at Luc-sur-Mer [
47]. Its abundance increased regularly from 2015 to 2017 when its abundance reached a maximum of 2750 ind·m
−2, then decreased in 2018 (
Table 4).
The tanaidacean
Zeuxo holdichi (Bamber, 1990) was described from the Bay of Arcachon in the south of the Bay of Biscay, France. Since its description, this small species (<6 mm total length) has been recorded in European waters, from the Iberian Peninsula to Germany [
49]. Along the French coast of the English Channel,
Z. holdichi remained rare at most of the locations where it was recorded except at Luc-sur-Mer where the species was very abundant in the blocks deployed during this experiment (Foveau et al., 2018). The reason for the high abundances (more than 100,000 individuals collected; up to 2500 ind·m
−2 in 2015) at this location remains enigmatic.
Z. holdichi can be considered as a pioneer invasive species. Since its first record in 2013 in Normandy waters in the Rade de Cherbourg [
49], it has become present in all soft-bottom and hard bottom habitats in Normandy (unpublished data). This shows that this species is ubiquitous in spite of its holobenthic development limiting its natural dispersal, which highlights its efficiency in engendering an abundant population in few months. Foveau et al. (2018) [
49] hypothesized that the spread of
Z. holdichi is in relation to aquaculture activities.
Depending on the fauna, two distinct patterns can be identified.
A rapid colonization by the pioneer barnacles was observed, favoured by the heterogeneous surface of the blocks [
42]. It is known that the complexity of the colonised surface influences micro-habitat selection and favours the settlement of barnacle cyprids [
13,
22,
26,
50].
Subsequently, the sessile fauna became characterized by a low biodiversity, with only few taxa colonizing the blocks, mainly barnacles and ascidians. These taxa show high abundances and great ability to cover the entire surface of the blocks, thus monopolizing the surface and preventing the establishment of other sessile taxa but favouring the presence of small motile organism. Such low biodiversity (one to four taxa according to the site) was also observed in Normandy marinas [
46], with the dominance of the NIS polychaete
Ficopomatus enigmaticus and the barnacle
Balanus improvisus (Darwin, 1854) with maximum densities reaching 10,000 ind·m
−2. The abundance of Balanus
crenatus in our blocks reached 33,000 ind·m
−2 in April 2014, then varied between 800 and 5500 ind·m
−2 from 2015 to 2018.
As regards the motile fauna, the opportunist amphipods and tanaids dominated the fauna in 2015, reaching a maximum abundance of around 5800 ind·m−2. Similarly, to the sessile fauna, the diversified motile fauna shows temporal fluctuations, with the lowest abundances observed during the last year of the study.
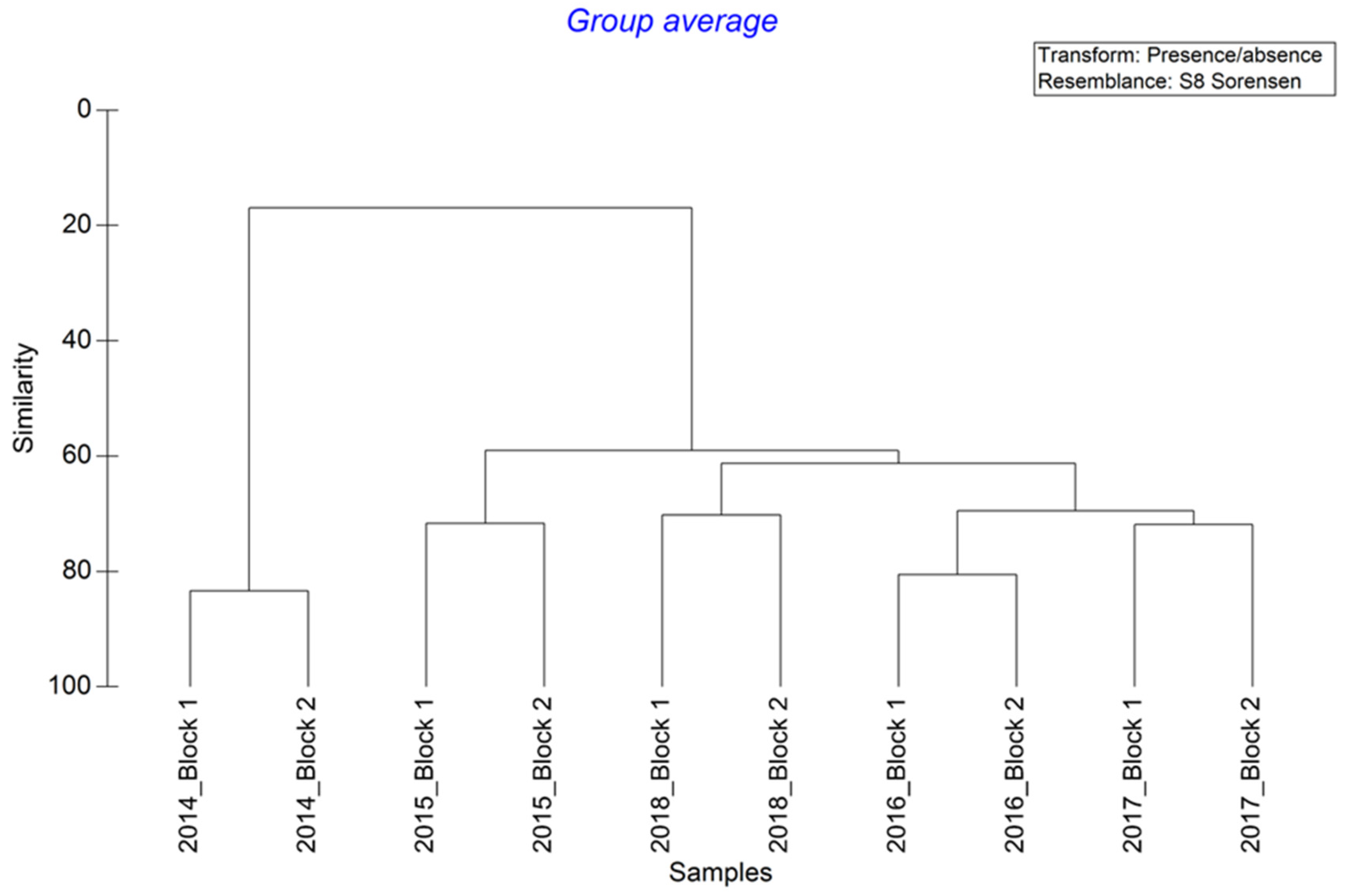
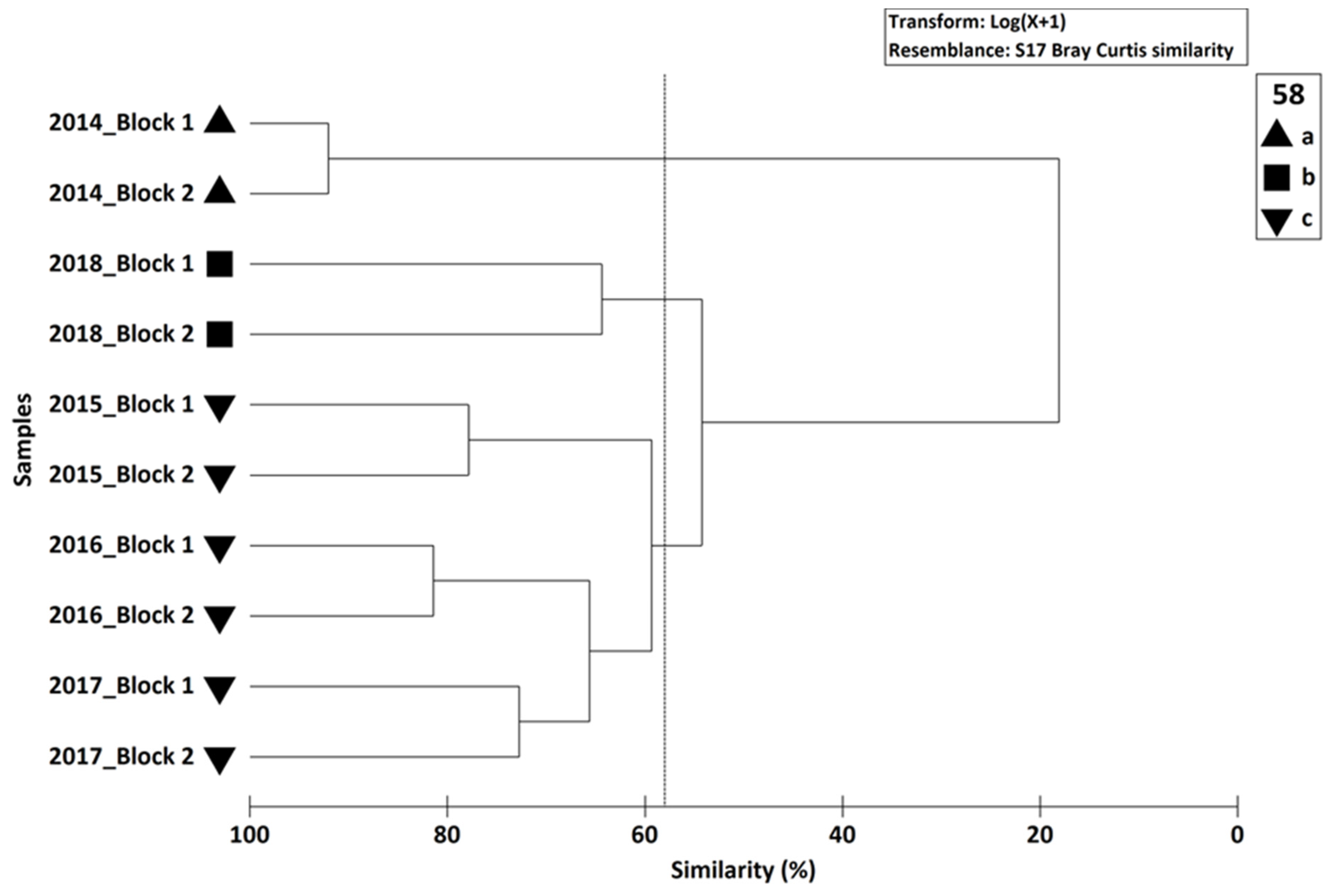
In summary, our experiment shows a very rapid colonisation of sessile fauna onto blocks deployed on oyster tables in the intertidal zone of the Bay of Seine, taking place over a period of one month. The colonization of motile fauna was also rapid, but took place over a period of one year. The type of observation used in this study was designed to test the efficiency of building materials favouring the colonisation of artificial structures on the lower part of the intertidal zone. Since this zone is accessible at each spring tide, it is easier to study than the subtidal zone, where the mobilization of divers needed to sample the blocks is dependent on the meteorological conditions. Two years after the beginning of the experiment, the blocks showed a relative stability. This process of colonisation is also observed on the geotextile artificial reef on the south coast of England, where the natural rock substrate where rare in the Poole Bay near the immersion zone of Boscombe (Herbert et al. 2017). At Luc-sur-Mer, the presence of natural hard substratum is not a limiting factor for the block colonisation by both sessile and motile surrounding macrofauna. An analysis of the main environmental factors controlling settlement, colonization and competition between species during the first year (2014–2015) of the block’s colonization shows that the dominant factors explaining biological processes are light plus hydrodynamics and larval behaviour [
23,
41,
42]. The main factor affecting the long-term pattern of colonisation is a biological pattern with succession of sessile taxa from pioneer and opportunist species such as the barnacles (100% of block cover), and then the settlement of the blue mussel, and ascidians and those of motile fauna with both pioneer taxa the amphipod
Monocorophium and the tanaid
Zeuxo holdichi, and a taxonomic diversity associated with a drastic reduction of abundance at the end of the study in 2018.
To our knowledge, the single study which had examined the faunal succession of artificial structure colonization in the English Channel were those of Herbert et al. (2017) and Taormina et al. (2020) [
10,
51]. Herbert et al. (2017) [
10] observed different stages in colonisation beginning with bryozoans and green algae which are replaced by red algae, hydroids and ascidians. Taormina et al. (2020) [
51] monitored the epibenthic colonization of artificial structures in a subtidal (18–20 m depth) high-energy hydrodynamic environment in the western part of the English Channel at a site planned for the deployment of a tidal turbine. Using four years of image-based underwater studies, they identified a rich fauna (28 taxa) and characterized changes of the epibenthic communities which reached a mature stage at the end of their study. However, their study suggested that the ecological succession was still in progress five years after the deployment of artificial structures.
However, it was difficult to compare the colonization of intertidal and subtidal structures with different designs and influences, because of the size of the blocks, i.e., it was known that for the sessile organisms the surface of potential colonised substrates was a limiting factor. However, it appeared noticeable differences in the colonisation patterns of intertidal and subtidal zones, with low diversity and rapid colonization in the intertidal zone, and high diversity and low colonization of subtidal artificial structures. It should be useful to measure the biomass of the colonizing fauna to estimate the role of artificial structures in the secondary production of artificial hard bottoms and compare them with natural hard bottoms, and thus assess their potential for increasing biodiversity in coastal ecosystems.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}