Experimental Assessment of a Conducting Polymer (PEDOT) and Microbial Biofilms as Deterrents and Facilitators of Macro-Biofouling: Larval Settlement of the Barnacle Notobalanus flosculus (Darwin, 1854) from Central Chile

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species and Sampling Area
2.2. Settlement Substrates
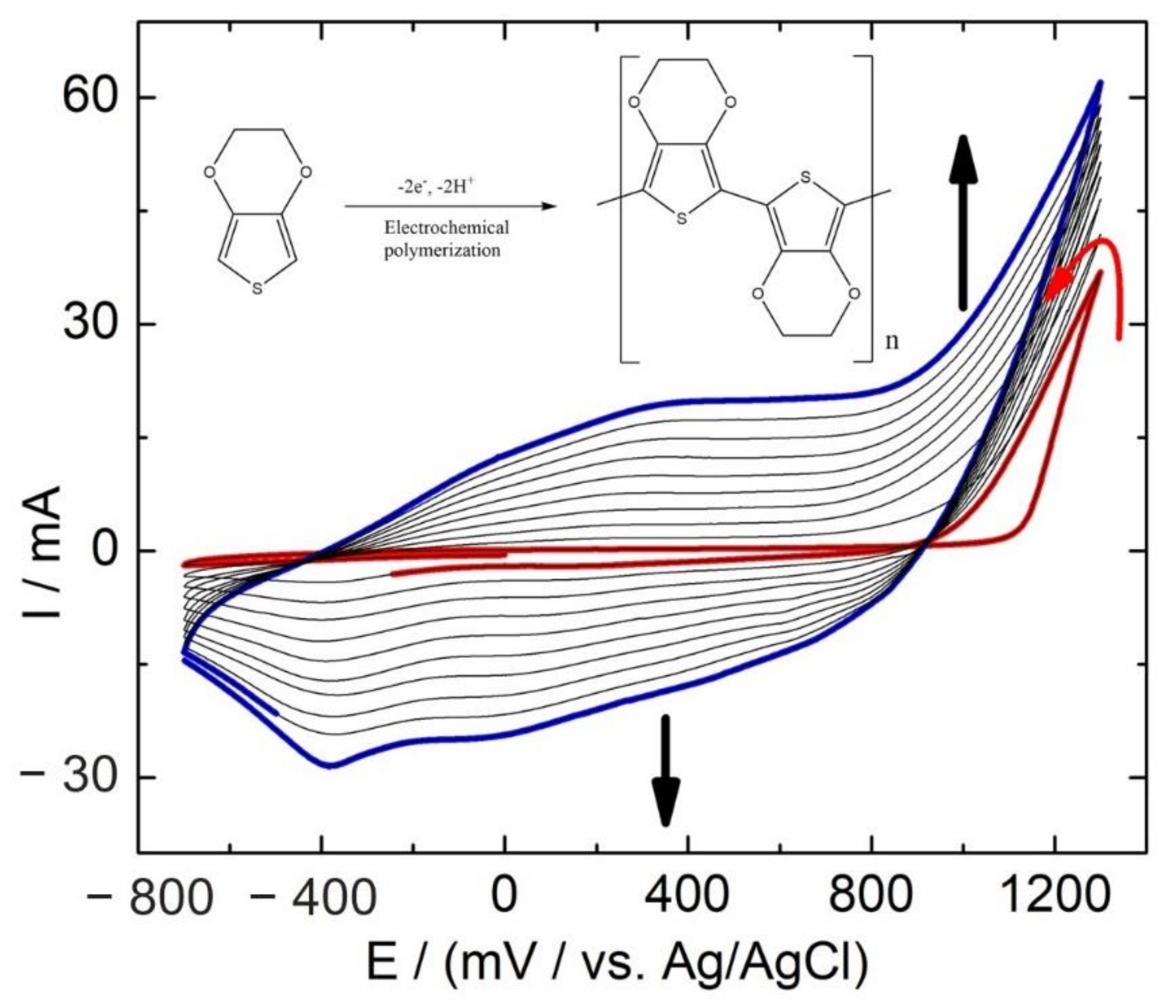
2.3. Preparation of PEDOT Coated Stainless Steel
2.4. Surface Roughness and Wettability of PEDOT, Acrylic and AF Coating
2.5. Biofilm Deposition and Characterization
2.6. Barnacle Collection, Spawning and Larval Rearing
2.7. Settlement Choice Experiments
2.7.1. Experimental Set-Up
2.7.2. Laboratory Trials
3. Results
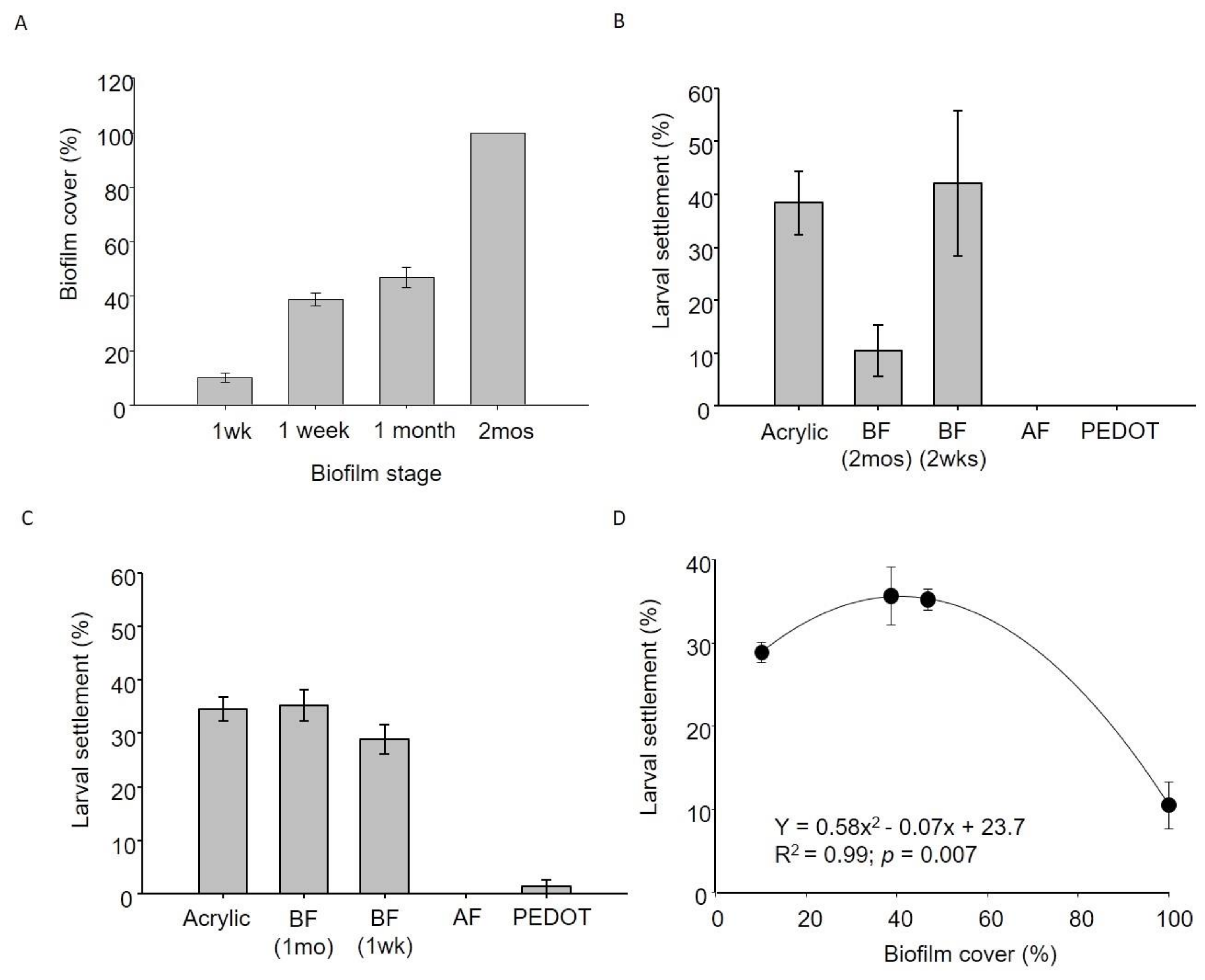
3.1. Physical Properties of Materials and Biofilm Coverage
3.2. Settlement Experiments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayers, J.; Turner, H. The Principal Fouling Organisms. In Marine Fouling and Its Prevention; Redfield, A., Ketchum, B., Eds.; United States Naval Institute: Annapolis, MD, USA, 1952. [Google Scholar]
- Callow, J.A.; Callow, M.E. Trends in the development of environmentally friendly fouling-resistant marine coatings. Nat. Commun. 2011, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Want, A.; Crawford, R.; Kakkonen, J.; Kiddie, G.; Miller, S.; Harris, R.E.; Porter, J.S. Biodiversity characterisation and hydrodynamic consequences of marine fouling communities on marine renewable energy infrastructure in the Orkney Islands Archipelago, Scotland, UK. Biofouling 2017, 33, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, P.A.; Simas, T.; Cruz, E.; Pinori, E.; Svenson, J. Marine biofouling: A European database for the marine renewable energy sector. J. Mar. Sci. Eng. 2020, 8, 495. [Google Scholar] [CrossRef]
- Davidson, I.; Scianni, C.; Hewitt, C.; Everett, R.; Holm, E.; Tamburri, M.; Ruiz, G. Mini-review: Assessing the drivers of ship biofouling management—Aligning industry and biosecurity goals. Biofouling 2016, 32, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Loxton, J.; Macleod, A.K.; Nall, C.R.; McCollin, T.; Machado, I.; Simas, T.; Vance, T.; Kenny, C.; Want, A.; Miller, R.G. Setting an agenda for biofouling research for the marine renewable energy industry. Int. J. Mar. Energy 2017, 19, 292–303. [Google Scholar] [CrossRef]
- Arinaga, R.A.; Cheung, K.F. Atlas of global wave energy from 10 years of reanalysis and hindcast data. Renew. Energy 2012, 39, 49–64. [Google Scholar] [CrossRef]
- López, I.; Andreu, J.; Ceballos, S.; De Alegría, M.I.; Kortabarria, I. Review of wave energy technologies and the necessary power-equipment. Renew. Sustain. Energy Rev. 2013, 413–434. [Google Scholar] [CrossRef]
- Gentry, R.R.; Froehlich, H.E.; Grimm, D.; Kareiva, P.; Parke, M.; Rust, M.; Gaines, S.D.; Halpern, B.S. Mapping the global potential for marine aquaculture. Nat. Ecol. Evol. 2017, 1, 1317–1324. [Google Scholar] [CrossRef]
- Lacoste, E.; Gaertner-Mazouni, N. Biofouling impact on production and ecosystem functioning: A Review for bivalve aquaculture. Rev. Aquac. 2015, 7, 187–196. [Google Scholar] [CrossRef]
- Macleod, A.K.; Stanley, M.S.; Day, J.G.; Cook, E.J. Biofouling community composition across a range of environmental conditions and geographical locations suitable for floating marine renewable energy generation. Biofouling 2016, 32, 261–276. [Google Scholar] [CrossRef]
- Clare, A.S.; Aldred, N. Surface colonisation by marine organisms and its impact on antifouling research. In Advances in Marine Antifouling Coatings and Technologies; Elsevier Ltd.: Amsterdam, The Netherlands, 2009; pp. 46–79. [Google Scholar] [CrossRef]
- Qian, P.Y.; Dahms, H.-U. A triangle model: Environmental changes affect biofilms that affect larval settlement. In Marine and Industrial Biofouling; Springer: Berlin/Heidelberg, Germany, 2008; pp. 315–328. [Google Scholar] [CrossRef]
- Hadfield, M.G. Biofilms and marine invertebrate larvae: What bacteria produce that larvae use to choose settlement sites. Ann. Rev. Mar. Sci. 2011, 3, 453–470. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Rittschof, D. Love at first taste: Induction of larval settlement by marine microbes. Int. J. Mol. Sci. 2020, 21, 731. [Google Scholar] [CrossRef] [PubMed]
- Aldred, N.; Nelson, A. Microbiome acquisition during larval settlement of the barnacle Semibalanus Balanoides. Biol. Lett. 2019, 15, 20180763. [Google Scholar] [CrossRef] [PubMed]
- Maan, A.M.C.; Hofman, A.H.; Vos, W.M.; Kamperman, M. Recent developments and practical feasibility of polymer-based antifouling coatings. Adv. Funct. Mater. 2020, 30, 2000936. [Google Scholar] [CrossRef]
- Salta, M.; Wharton, J.A.; Blache, Y.; Stokes, K.R.; Briand, J.-F. Marine biofilms on artificial surfaces: Structure and dynamics. Environ. Microbiol. 2013. [Google Scholar] [CrossRef]
- Salta, M.; Wharton, J.A.; Stoodley, P.; Dennington, S.P.; Goodes, L.R.; Werwinski, S.; Mart, U.; Wood, R.J.K.; Stokes, K.R. Designing biomimetic antifouling surfaces. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2010, 4729–4754. [Google Scholar] [CrossRef]
- Liu, L.; Li, W.; Liu, Q. Recent development of antifouling polymers: Structure, evaluation, and biomedical applications in nano/micro-structures. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2014, 6, 599–614. [Google Scholar] [CrossRef]
- Gomez-Carretero, S.; Nybom, R.; Richter-Dahlfors, A. Electroenhanced antimicrobial coating based on conjugated polymers with covalently coupled silver nanoparticles prevents Staphylococcus Aureus biofilm formation. Adv. Healthc. Mater. 2017, 6, 1700435. [Google Scholar] [CrossRef]
- Mathiazhagan, A.; Joseph, R. Nanotechnology-A new prospective in organic coating—Review. Int. J. Chem. Eng. Appl. 2011, 225–237. [Google Scholar] [CrossRef]
- Yang, M.C.; Tsou, H.M.; Hsiao, Y.S.; Cheng, Y.W.; Liu, C.C.; Huang, L.Y.; Peng, X.Y.; Liu, T.Y.; Yung, M.C.; Hsu, C.C. Electrochemical polymerization of PEDOT-graphene oxide-heparin composite coating for anti-fouling and anti-clotting of cardiovascular stents. Polymers 2019, 11, 1520. [Google Scholar] [CrossRef]
- Li, Y.; Ning, C. Latest research progress of marine microbiological corrosion and bio-fouling, and new approaches of marine anti-corrosion and anti-fouling. Bioact. Mater. 2019, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Armelin, E.; Meneguzzi, Á.; Ferreira, C.A.; Alemán, C. Polyaniline, polypyrrole and poly (3,4-Ethylenedioxythiophene) as additives of organic coatings to prevent corrosion. Surf. Coat. Technol. 2009, 203, 3763–3769. [Google Scholar] [CrossRef]
- Zhu, G.; Hou, J.; Zhu, H.; Qiu, R.; Xu, J. Electrochemical synthesis of poly(3,4-Ethylenedioxythiophene) on stainless steel and its corrosion inhibition performance. J. Coat. Technol. Res. 2013, 10, 659–668. [Google Scholar] [CrossRef]
- Aguirre, J.; Daille, L.; Fischer, D.A.; Galarce, C.; Pizarro, G.; Vargas, I.; Walczak, M.; de la Iglesia, R.; Armijo, F. Study of poly (3,4-Ethylendioxythiphene) as a coating for mitigation of biocorrosion of AISI 304 stainless steel in natural seawater. Prog. Org. Coat. 2017, 113, 175–184. [Google Scholar] [CrossRef]
- Smart Polymers and Their Applications | ScienceDirect. Available online: https://www.sciencedirect.com/book/9780857096951/smart-polymers-and-their-applications (accessed on 2 November 2020).
- Del Valle, M.A.; Ramírez, A.M.; Hernández, L.A.; Armijo, F.; Díaz, F.R.; Arteaga, G.C. Influence of the supporting electrolyte on the electrochemical polymerization of 3,4-ethylenedioxythiophene. Effect on p- and n-doping/undoping, conductivity and morphology. Int. J. Electrochem. Sci. 2016, 11, 7048–7065. [Google Scholar] [CrossRef]
- Romero, M.; del Valle, M.A.; del Río, R.; Díaz, F.R.; Armijo, F.; Dalchiele, E.A. Temperature effect on nucleation and growth mechanism of poly (o-anisidine) and poly (aniline) electro-synthesis. J. Electrochem. Soc. 2013, 160, G125–G134. [Google Scholar] [CrossRef]
- Salgado, R.; del Rio, R.; del Valle, M.A.; Armijo, F. Selective electrochemical determination of dopamine, using a poly (3,4-Ethylenedioxythiophene)/polydopamine hybrid film modified electrode. J. Electroanal. Chem. 2013, 704, 130–136. [Google Scholar] [CrossRef]
- Darmanin, T.; Guittard, F. Wettability of conducting polymers: From superhydrophilicity to superoleophobicity. Prog. Polym. Sci. 2014, 656–682. [Google Scholar] [CrossRef]
- Tapia, F.J.; Largier, J.L.; Castillo, M.; Wieters, E.A.; Navarrete, S.A. Latitudinal discontinuity in thermal conditions along the nearshore of central-northern Chile. PLoS ONE 2014, 9, e110841. [Google Scholar] [CrossRef]
- Cifuentes, M.; Kamlah, C.; Thiel, M.; Lenz, M.; Wahl, M. Effects of temporal variability of disturbance on the succession in marine fouling communities in northern-central Chile. J. Exp. Mar. Biol. Ecol. 2007, 352, 280–294. [Google Scholar] [CrossRef]
- Cifuentes, M.; Krueger, I.; Dumont, C.P.; Lenz, M.; Thiel, M. Does primary colonization or community structure determine the succession of fouling communities? J. Exp. Mar. Biol. Ecol. 2010, 395, 10–20. [Google Scholar] [CrossRef]
- Navarrete, S.A.; Parragué, M.; Osiadacz, N.; Rojas, F.; Bonicelli, J.; Fernández, M.; Arboleda-Baena, C.; Perez-Matus, A.; Finke, R. Abundance, composition and succession of sessile subtidal assemblages in high wave-energy environments of central Chile: Temporal and Depth Variation. J. Exp. Mar. Biol. Ecol. 2019, 512, 51–62. [Google Scholar] [CrossRef]
- Broitman, B.; Navarrete, S.; Smith, F.; Gaines, S. Geographic variation of southeastern Pacific intertidal communities. Mar. Ecol. Prog. Ser. 2001, 224, 21–34. [Google Scholar] [CrossRef]
- Pacheco, A.S.; Laudien, J.; Thiel, M.; Oliva, M.; Heilmayer, O. Succession and seasonal onset of colonization in subtidal hard-bottom communities off northern Chile. Mar. Ecol. 2011, 32, 75–87. [Google Scholar] [CrossRef]
- Darwin, C. A monograph of the subclass cirripedia, with figures of all the species. The lepadidæ or pedunculated cirripedes. Ann. Mag. Nat. Hist. 1853, 12, 444–448. [Google Scholar] [CrossRef]
- Valdivia, N.; Heidemann, A.; Thiel, M.; Molis, M.; Wahl, M. Effects of disturbance on the diversity of hard-bottom macrobenthic communities on the coast of Chile. Mar. Ecol. Prog. Ser. 2005, 299, 45–54. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Otsu, N. Threshold selection method from gray-level histograms. IEEE Trans. Syst. Man Cybern. 1979, 9, 62–66. [Google Scholar] [CrossRef]
- Jonsson, P.R.; Wrange, A.L.; Lind, U.; Abramova, A.; Ogemark, M.; Blomberg, A. The barnacle balanus improvisus as a marine model—Culturing and gene expression. J. Vis. Exp. 2018, 2018, 57825. [Google Scholar] [CrossRef]
- Pansch, C.; Jonsson, P.R.; Berglin, M.; Pinori, E.; Wrange, A.L. A new flow-through bioassay for testing low-emission antifouling coatings. Biofouling 2017, 33, 613–623. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar] [CrossRef]
- Dahlström, M.; Jonsson, H.; Jonsson, P.R.; Elwing, H. Surface wettability as a determinant in the settlement of the barnacle balanus improvisus (DARWIN). J. Exp. Mar. Biol. Ecol. 2004, 305, 223–232. [Google Scholar] [CrossRef]
- Petrone, L.; di Fino, A.; Aldred, N.; Sukkaew, P.; Ederth, T.; Clare, A.S.; Liedberg, B. Effects of surface charge and gibbs surface energy on the settlement behaviour of barnacle cyprids (Balanus Amphitrite). Biofouling 2011, 27, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Aldred, N.; Ista, L.K.; Callow, M.E.; Callow, J.A.; Lopez, G.P.; Clare, A.S. Mussel (Mytilus Edulis) byssus deposition in response to variations in surface wettability. J. R. Soc. Interface 2006, 3, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Callow, J.; Callow, M.; Ista, L.; Lopez, G.; Chaudhury, M. The influence of surface energy on the wetting behaviour of the spore adhesive of the marine alga Ulva Linza (Synonym Enteromorpha Linza). J. R. Soc. Interface 2005, 2, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, S.; Parragué, M.; Osiadacz, N.; Rojas, F.; Bonicelli, J.; Fernández, M.; Arboleda-Baena, C.; Finke, R.; Baldanzi, S. Susceptibility of different materials and antifouling coating to macrofouling organisms in a high wave-energy environment. J. Ocean Technol. 2020, 15, 70–91. [Google Scholar]
- Roberts, D.; Rittschof, D.; Holm, E.; Schmidt, A.R. Factors influencing initial larval settlement: Temporal, spatial and surface molecular components. J. Exp. Mar. Biol. Ecol. 1991, 150, 203–221. [Google Scholar] [CrossRef]
- Wieczorek, S.; Clare, A.; Todd, C. Inhibitory and facilitatory effects of microbial films on settlement of balanus amphitrite amphitrite larvae. Mar. Ecol. Prog. Ser. 1995, 119, 221–228. [Google Scholar] [CrossRef]
- Huggett, M.J.; Nedved, B.T.; Hadfield, M.G. Effects of initial surface wettability on biofilm formation and subsequent settlement of hydroides elegans. Biofouling 2009, 25, 387–399. [Google Scholar] [CrossRef]
- Toupoint, N.; Mohit, V.; Linossier, I.; Bourgougnon, N.; Myrand, B.; Olivier, F.; Lovejoy, C.; Tremblay, R. Effect of biofilm age on settlement of mytilus edulis. Biofouling 2012, 28, 985–1001. [Google Scholar] [CrossRef]
- Mos, B.; Cowden, K.L.; Nielsen, S.J.; Dworjanyn, S.A. Do cues matter? Highly inductive settlement cues don’t ensure high post-settlement survival in sea urchin aquaculture. PLoS ONE 2011, 6, e28054. [Google Scholar] [CrossRef]
- Daille, L.K.; Aguirre, J.; Fischer, D.; Galarce, C.; Armijo, F.; Pizarro, G.E.; Walczak, M.; de la Iglesia, R.; Vargas, I.T. Effect of tidal cycles on bacterial biofilm formation and biocorrosion of stainless steel AISI 316L. J. Mar. Sci. Eng. 2020, 8, 124. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plate | Surface Roughness (µm2) | Wettability (WCA) | Hydrophobicity |
|---|---|---|---|
| Acrylic | 0.332 | 81.9 | Intermediate |
| PEDOT | 0.118 | 103.4 | Hydrophobic |
| AF | 0.243 | 81.13 | Intermediate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldanzi, S.; Vargas, I.T.; Armijo, F.; Fernández, M.; Navarrete, S.A. Experimental Assessment of a Conducting Polymer (PEDOT) and Microbial Biofilms as Deterrents and Facilitators of Macro-Biofouling: Larval Settlement of the Barnacle Notobalanus flosculus (Darwin, 1854) from Central Chile. J. Mar. Sci. Eng. 2021, 9, 82. https://doi.org/10.3390/jmse9010082
Baldanzi S, Vargas IT, Armijo F, Fernández M, Navarrete SA. Experimental Assessment of a Conducting Polymer (PEDOT) and Microbial Biofilms as Deterrents and Facilitators of Macro-Biofouling: Larval Settlement of the Barnacle Notobalanus flosculus (Darwin, 1854) from Central Chile. Journal of Marine Science and Engineering. 2021; 9(1):82. https://doi.org/10.3390/jmse9010082
Chicago/Turabian StyleBaldanzi, Simone, Ignacio T. Vargas, Francisco Armijo, Miriam Fernández, and Sergio A. Navarrete. 2021. "Experimental Assessment of a Conducting Polymer (PEDOT) and Microbial Biofilms as Deterrents and Facilitators of Macro-Biofouling: Larval Settlement of the Barnacle Notobalanus flosculus (Darwin, 1854) from Central Chile" Journal of Marine Science and Engineering 9, no. 1: 82. https://doi.org/10.3390/jmse9010082
APA StyleBaldanzi, S., Vargas, I. T., Armijo, F., Fernández, M., & Navarrete, S. A. (2021). Experimental Assessment of a Conducting Polymer (PEDOT) and Microbial Biofilms as Deterrents and Facilitators of Macro-Biofouling: Larval Settlement of the Barnacle Notobalanus flosculus (Darwin, 1854) from Central Chile. Journal of Marine Science and Engineering, 9(1), 82. https://doi.org/10.3390/jmse9010082