Using Artificial-Reef Knowledge to Enhance the Ecological Function of Offshore Wind Turbine Foundations: Implications for Fish Abundance and Diversity




Abstract
1. Introduction
Objectives
2. Materials and Methods
2.1. Search Terms
2.2. Screening Process
2.3. Data Extraction
2.4. Data Analysis
3. Results
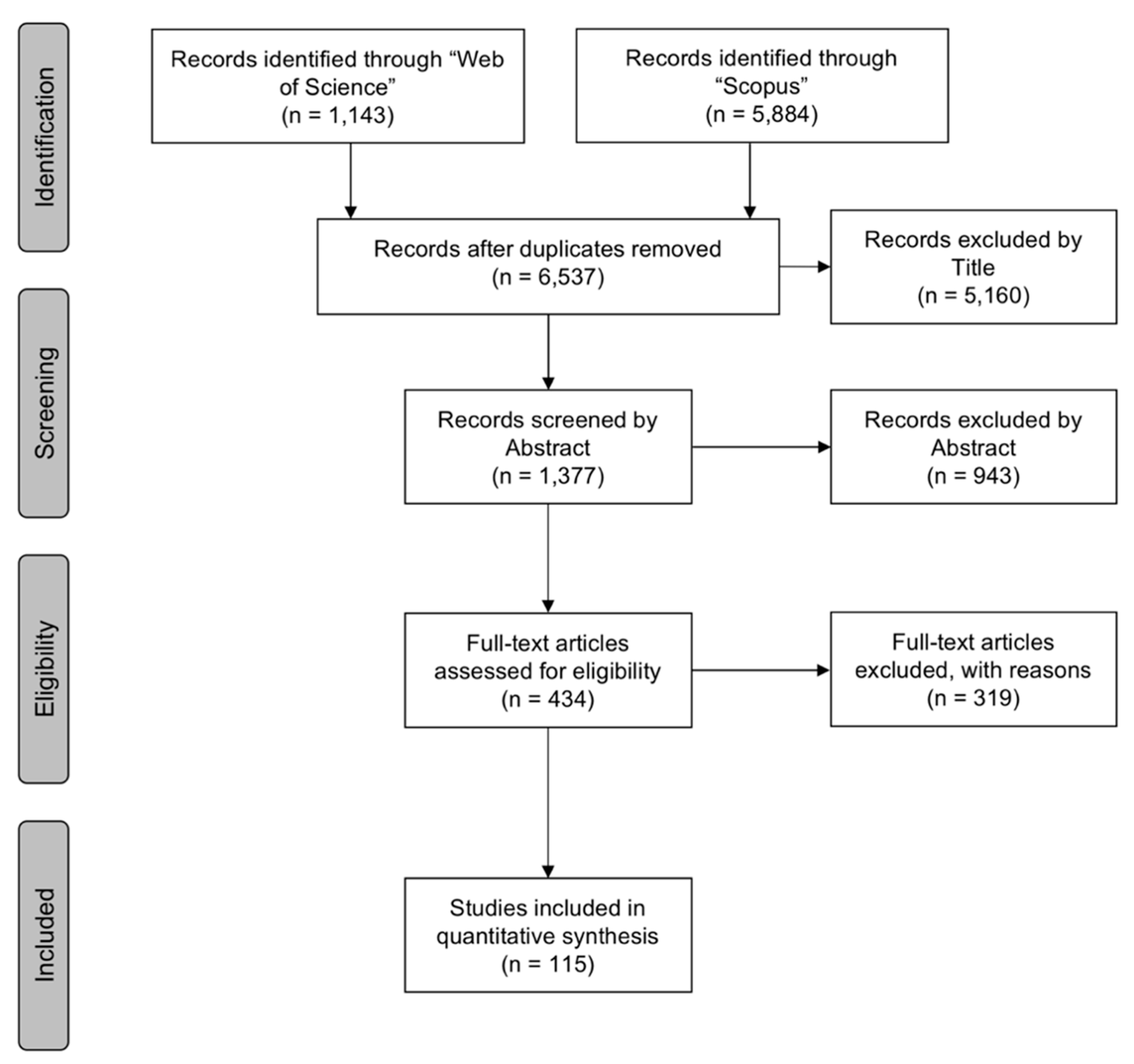
3.1. Selection Process
3.2. Temporal Research Trends
3.3. Location of Available Studies
3.4. Inconsistent Study Details
3.5. Overlapping Use of Materials for ARs and OWFs
3.6. Overlapping Water Depths Used for ARs and OWFs
3.7. Effects of ARs and OWFs on Fish Abundance
4. Discussion
4.1. Fish Abundance, Biodiversity and Distribution Associated with ARs and OWFs
4.2. Attraction Versus Production
4.3. Community Changes Associated with ARs and OWFs
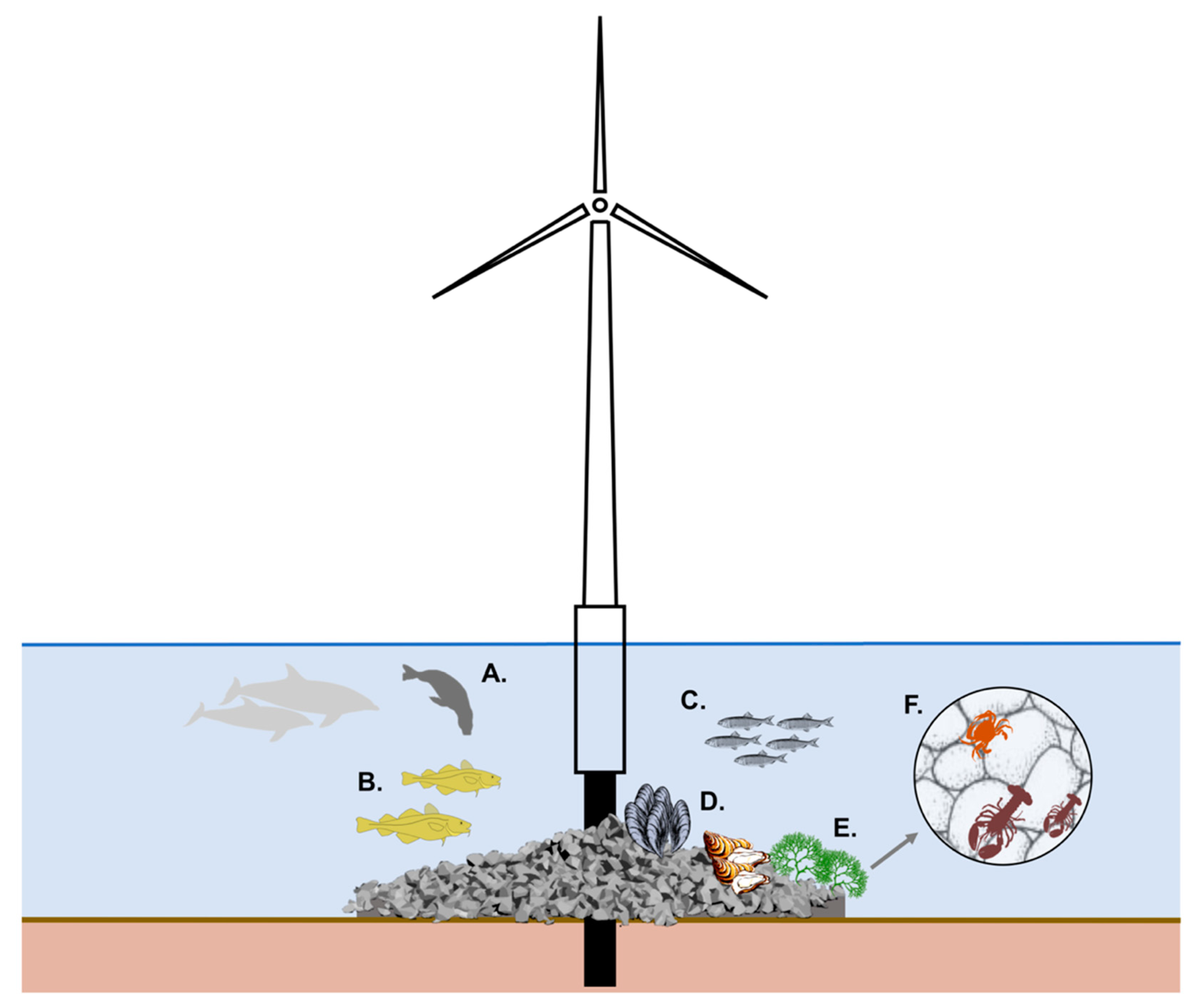
4.4. Scour Protection Functioning as An AR
4.4.1. Food Provision
4.4.2. Shelter
4.4.3. Nursery
4.4.4. Distance between Artificial Habitats
4.4.5. Reef Size
4.5. Optimizing Scour Protection
4.5.1. Structural Complexity
4.5.2. Materials
4.5.3. Enhancing Ecological Function
4.6. Advancing the Field
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Elbert, C.; Fox, H.E.; et al. A Global Map of Human Impact on Marine Ecosystems. Science 2008, 319, 948–953. [Google Scholar] [CrossRef]
- Halpern, B.S.; Longo, C.; Hardy, D.; McLeod, K.L.; Samhouri, J.F.; Katona, S.K.; Kleisner, K.; Lester, S.E.; Oleary, J.; Ranelletti, M.; et al. An index to assess the health and benefits of the global ocean. Nature 2012, 488, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Waltham, N.J.; Connolly, R.M. Global extent and distribution of artificial, residential waterways in estuaries. Estuar. Coast. Shelf Sci. 2011, 94, 192–197. [Google Scholar] [CrossRef]
- Dafforn, K.A.; Glasby, T.M.; Airoldi, L.; Rivero, N.K.; Mayer-Pinto, M.; Johnston, E.L. Marine urbanization: An ecological framework for designing multifunctional artificial structures. Front. Ecol. Environ. 2015, 13, 82–90. [Google Scholar] [CrossRef]
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.H.L.; Saksena-Taylor, P.; et al. Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Mar. Pollut. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef]
- Bulleri, F.; Chapman, M.G. The introduction of coastal infrastructure as a driver of change in marine environments. J. Appl. Ecol. 2010, 47, 26–35. [Google Scholar] [CrossRef]
- Bishop, M.J.; Mayer-Pinto, M.; Airoldi, L.; Firth, L.B.; Morris, R.L.; Loke, L.H.L.; Hawkins, S.J.; Naylor, L.A.; Coleman, R.A.; Chee, S.Y.; et al. Effects of ocean sprawl on ecological connectivity: Impacts and solutions. J. Exp. Mar. Biol. Ecol. 2017, 492, 7–30. [Google Scholar] [CrossRef]
- Heery, E.C.; Bishop, M.J.; Critchley, L.P.; Bugnot, A.B.; Airoldi, L.; Mayer-Pinto, M.; Sheehan, E.V.; Coleman, R.A.; Loke, L.H.L.; Johnston, E.L.; et al. Identifying the consequences of ocean sprawl for sedimentary habitats. J. Exp. Mar. Biol. Ecol. 2017, 492, 31–48. [Google Scholar] [CrossRef]
- Bortone, S.A.; Seaman, W. Technology for the creation of aquatic habitats and their evaluation in fisheries ecosystems. Fish. Sci. 2002, 68, 1677–1682. [Google Scholar] [CrossRef]
- Seaman, W. Artificial habitats and the restoration of degraded marine ecosystems and fisheries. Hydrobiologia 2007, 580, 143–155. [Google Scholar] [CrossRef]
- Firth, L.B.; Thompson, R.C.; Bohn, K.; Abbiati, M.; Airoldi, L.; Bouma, T.J.; Bozzeda, F.; Ceccherelli, V.U.; Colangelo, M.A.; Evans, A.; et al. Between a rock and a hard place: Environmental and engineering considerations when designing coastal defence structures. Coast. Eng. 2014, 87, 122–135. [Google Scholar] [CrossRef]
- Baine, M. Artificial reefs: A review of their design, application, management and performance. Ocean Coast. Manag. 2001, 44, 241–259. [Google Scholar] [CrossRef]
- Tessier, A.; Francour, P.; Charbonnel, E.; Dalias, N.; Bodilis, P.; Seaman, W.; Lenfant, P. Assessment of French artificial reefs: Due to limitations of research, trends may be misleading. Hydrobiologia 2015, 753, 1–29. [Google Scholar] [CrossRef]
- Reubens, J.T.; Braeckman, U.; Vanaverbeke, J.; Van Colen, C.; Degraer, S.; Vincx, M. Aggregation at windmill artificial reefs: CPUE of Atlantic cod (Gadus morhua) and pouting (Trisopterus luscus) at different habitats in the Belgian part of the North Sea. Fish. Res. 2013, 139, 28–34. [Google Scholar] [CrossRef]
- Fujii, T. Potential influence of offshore oil and gas platforms on the feeding ecology of fish assemblages in the North Sea. Mar. Ecol. Prog. Ser. 2016, 542, 167–186. [Google Scholar] [CrossRef]
- Fowler, A.M.; Jørgensen, A.; Coolen, J.W.P.; Brabant, R.; Rumes, B.; Degraer, S. The ecology of infrastructure decommissioning in the North Sea: What we need to know and how to achieve it. ICES J. Mar. Sci. 2019, 77, 1109–1126. [Google Scholar] [CrossRef]
- Edney, J.; Spennemann, D.H.R. Can Artificial Reef Wrecks Reduce Diver Impacts on Shipwrecks? The Management Dimension. J. Marit. Archaeol. 2015, 10, 141–157. [Google Scholar] [CrossRef]
- Petersen, J.K.; Malm, T. Offshore Windmill Farms: Threats to or Possibilities for the Marine Environment. Ambio 2006, 35, 75–80. [Google Scholar] [CrossRef]
- Ramírez, L.; Fraile, D.; Brindley, G. Offshore wind in Europe—Key trends and statistics 2019; WindEurope. 2020, pp. 1–38. Available online: https://windeurope.org/about-wind/statistics/offshore/european-offshore-wind-industry-key-trends-statistics-2019/ (accessed on 8 May 2020).
- Drewitt, A.L.; Langston, R.H.W. Assessing the impacts of wind farms on birds. Ibis 2006, 148, 29–42. [Google Scholar] [CrossRef]
- Thaxter, C.B.; Buchanan, G.M.; Carr, J.; Butchart, S.H.M.; Newbold, T.; Green, R.E.; Tobias, J.A.; Foden, W.B.; O’Brien, S.; Pearce-Higgins, J.W. Bird and bat species’ global vulnerability to collision mortality at wind farms revealed through a trait-based assessment. Proc. R. Soc. B 2017, 284, 20170829. [Google Scholar] [CrossRef]
- Wahlberg, M.; Westerberg, H. Hearing in fish and their reactions to sounds from offshore wind farms. Mar. Ecol. Prog. Ser. 2005, 288, 295–309. [Google Scholar] [CrossRef]
- Madsen, P.T.; Wahlberg, M.; Tougaard, J.; Lucke, K.; Tyack, P. Wind turbine underwater noise and marine mammals: Implications of current knowledge and data needs. Mar. Ecol. Prog. Ser. 2006, 309, 279–295. [Google Scholar] [CrossRef]
- Brandt, M.J.; Dragon, A.C.; Diederichs, A.; Bellmann, M.A.; Wahl, V.; Piper, W.; Nabe-Nielsen, J.; Nehls, G. Disturbance of harbour porpoises during construction of the first seven offshore wind farms in Germany. Mar. Ecol. Prog. Ser. 2018, 596, 213–232. [Google Scholar] [CrossRef]
- Öhman, M.C.; Sigray, P.; Westerberg, H. Offshore windmills and the effects of electromagnetic fields on fish. AMBIO 2007, 36, 630–633. [Google Scholar] [CrossRef]
- Bray, L.; Reizopoulou, S.; Voukouvalas, E.; Soukissian, T.; Alomar, C.; Vázquez-Luis, M.; Deudero, S.; Attrill, M.J.; Hall-Spencer, J.M. Expected effects of offshore wind farms on Mediterranean Marine Life. J. Mar. Sci. Eng. 2016, 4, 18. [Google Scholar] [CrossRef]
- Langhamer, O.; Wilhelmsson, D. Colonisation of fish and crabs of wave energy foundations and the effects of manufactured holes—A field experiment. Mar. Environ. Res. 2009, 68, 151–157. [Google Scholar] [CrossRef]
- Punt, M.J.; Groeneveld, R.A.; van Ierland, E.C.; Stel, J.H. Spatial planning of offshore wind farms: A windfall to marine environmental protection? Ecol. Econ. 2009, 69, 93–103. [Google Scholar] [CrossRef]
- Langhamer, O. Artificial reef effect in relation to offshore renewable energy conversion: State of the art. Sci. World J. 2012, 2012, 386713. [Google Scholar] [CrossRef]
- Ashley, M.C.; Mangi, S.C.; Rodwell, L.D. The potential of offshore windfarms to act as marine protected areas—A systematic review of current evidence. Mar. Policy 2014, 45, 301–309. [Google Scholar] [CrossRef]
- den Boon, J.; Sutherland, J.; Whitehouse, R. Scour behaviour and scour protection for monopile foundations of offshore wind turbines. In Proceedings of the European Wind Energy Conference & Exhibition (EWEC), London, UK, 22–25 November 2004; Volume 14, pp. 1–14. [Google Scholar]
- Sumer, B.M.; Fredsøe, J. The Mechanics of Scour in the Marine Environment; World Scientific: River Edge, NJ, USA, 2002; Volume 17, pp. 1–552. [Google Scholar]
- Atalah, J.; Fitch, J.; Coughlan, J.; Chopelet, J.; Coscia, I.; Farrell, E. Diversity of demersal and megafaunal assemblages inhabiting sandbanks of the Irish Sea. Mar. Biodivers. 2013, 43, 121–132. [Google Scholar] [CrossRef]
- Hammar, L.; Andersson, S.; Rosenberg, R. Adapting offshore wind power foundations to local environment; Swedish Environmental Protection Agency; 2010; Report no. 6367. pp. 1–86. Available online: http://www.naturvardsverket.se/documents/publikationer/978-91-620-6367-2.pdf (accessed on 8 May 2020).
- Bergström, L.; Sundqvist, F.; Bergström, U. Effects of an offshore wind farm on temporal and spatial patterns in the demersal fish community. Mar. Ecol. Prog. Ser. 2013, 485, 199–210. [Google Scholar] [CrossRef]
- De Vos, L.; De Rouck, J.; Troch, P.; Frigaard, P. Empirical design of scour protections around monopile foundations. Part 2: Dynamic approach. Coast. Eng. 2012, 60, 286–298. [Google Scholar] [CrossRef]
- Liang, B.; Du, S.; Pan, X.; Zhang, L. Local scour for vertical piles in steady currents: Review of mechanisms, influencing factors and empirical equations. J. Mar. Sci. Eng. 2020, 8, 4. [Google Scholar] [CrossRef]
- Whitehouse, R.J.S.; Harris, J.M.; Sutherland, J.; Rees, J. The nature of scour development and scour protection at offshore windfarm foundations. Mar. Pollut. Bull. 2011, 62, 73–88. [Google Scholar] [CrossRef]
- Steffen, W.; Crutzen, P.J.; Mcneill, J. The Anthropocene: Are Humans Now Overwhelming the Great Forces of Nature? AMBIO 2007, 36, 614–621. [Google Scholar] [CrossRef]
- Scheffers, B.R.; De Meester, L.; Bridge, T.C.L.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.M.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The broad footprint of climate change from genes to biomes to people. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Garnett, S.T.; Barlow, S.; Bekessy, S.A.; Bellairs, S.M.; Bishop, M.J.; Bradstock, R.A.; Jones, D.N.; Maxwell, S.L.; Pittock, J.; et al. Renewal ecology: Conservation for the Anthropocene. Restor. Ecol. 2017, 25, 674–680. [Google Scholar] [CrossRef]
- Maes, J.; Jacobs, S. Nature-Based Solutions for Europe’s Sustainable Development. Conserv. Lett. 2017, 10, 121–124. [Google Scholar] [CrossRef]
- Lenkeek, W.; Didderen, K.; Teunis, M.; Driessen, F.; Coolen, J.W.P.; Bos, O.G.; Vergouwen, S.A.; Raaijmakers, T.C.; de Vries, M.B.; van Koningsveld, M. Eco-friendly design of scour protection: potential enhancement of ecological functioning in offshore wind farms; Ministry of Economic Affairs, Department of Nature & Biodiversity; 2017; Report no. 17–001. pp. 1–28. Available online: https://edepot.wur.nl/411374 (accessed on 8 May 2020).
- Morris, R.L.; Porter, A.G.; Figueira, W.F.; Coleman, R.A.; Fobert, E.K.; Ferrari, R. Fish-smart seawalls: A decision tool for adaptive management of marine infrastructure. Front. Ecol. Environ. 2018, 16, 278–287. [Google Scholar] [CrossRef]
- Strain, E.M.A.; Olabarria, C.; Mayer-Pinto, M.; Cumbo, V.; Morris, R.L.; Bugnot, A.B.; Dafforn, K.A.; Heery, E.; Firth, L.B.; Brooks, P.R.; et al. Eco-engineering urban infrastructure for marine and coastal biodiversity: Which interventions have the greatest ecological benefit? J. Appl. Ecol. 2018, 55, 426–441. [Google Scholar] [CrossRef]
- Dafforn, K.A.; Mayer-Pinto, M.; Morris, R.L.; Waltham, N.J. Application of management tools to integrate ecological principles with the design of marine infrastructure. J. Environ. Manag. 2015, 158, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Pullin, A.S.; Stewart, G.B. Guidelines for systematic review in conservation and environmental management. Conserv. Biol. 2006, 20, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Papathanasopoulou, E.; Beaumont, N.; Hooper, T.; Nunes, J.; Queirós, A.M. Energy systems and their impacts on marine ecosystem services. Renew. Sustain. Energy Rev. 2015, 52, 917–926. [Google Scholar] [CrossRef]
- Flávio, H.M.; Ferreira, P.; Formigo, N.; Svendsen, J.C. Reconciling agriculture and stream restoration in Europe: A review relating to the EU Water Framework Directive. Sci. Total Environ. 2017, 596–597, 378–395. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Libersti, A.; Tetzlaff, J.; Altman, D.G.; Group, T.P. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Whitmarsh, S.K.; Fairweather, P.G.; Huveneers, C. What is Big BRUVver up to? Methods and uses of baited underwater video. Rev. Fish Biol. Fish. 2017, 27, 53–73. [Google Scholar] [CrossRef]
- Støttrup, J.G.; Dahl, K.; Niemann, S.; Stenberg, C.; Reker, J.; Stamphøj, E.M.; Göke, C.; Svendsen, J.C. Restoration of a boulder reef in temperate waters: Strategy, methodology and lessons learnt. Ecol. Eng. 2017, 102, 468–482. [Google Scholar] [CrossRef]
- Kristensen, L.D.; Støttrup, J.G.; Svendsen, J.C.; Stenberg, C.; Højbjerg Hansen, O.K.; Grønkjær, P. Behavioural changes of Atlantic cod (Gadus morhua) after marine boulder reef restoration: Implications for coastal habitat management and Natura 2000 areas. Fish. Manag. Ecol. 2017, 24, 353–360. [Google Scholar] [CrossRef]
- Kayano, Y. Effects of artificial reefs and the acoustic-sound feeding method on the colonization of released grouper seedlings. Fish. Sci. 2002, 68, 1683–1686. [Google Scholar] [CrossRef][Green Version]
- Castège, I.; Milon, E.; Fourneau, G.; Tauzia, A. First results of fauna community structure and dynamics on two artificial reefs in the south of the Bay of Biscay (France). Estuar. Coast. Shelf Sci. 2016, 179, 172–180. [Google Scholar] [CrossRef]
- Carral, L.; Alvarez-Feal, J.C.; Tarrio-Saavedra, J.; Rodriguez Guerreiro, M.J.; Fraguela, J.Á. Social interest in developing a green modular artificial reef structure in concrete for the ecosystems of the Galician rías. J. Clean. Prod. 2018, 172, 1881–1898. [Google Scholar] [CrossRef]
- Yang, Y.; Ji, T.; Lin, X.; Chen, C.; Yang, Z. Biogenic sulfuric acid corrosion resistance of new artificial reef concrete. Constr. Build. Mater. 2018, 158, 33–41. [Google Scholar] [CrossRef]
- Sánchez, S.; López-Gutiérrez, J.S.; Negro, V.; Esteban, M.D. Foundations in offshore wind farms: Evolution, characteristics and range of use. Analysis of main dimensional parameters in monopile foundations. J. Mar. Sci. Eng. 2019, 7, 441. [Google Scholar] [CrossRef]
- Abecasis, D.; Bentes, L.; Lino, P.G.; Santos, M.N.; Erzini, K. Residency, movements and habitat use of adult white seabream (Diplodus sargus) between natural and artificial reefs. Estuar. Coast. Shelf Sci. 2013, 118, 80–85. [Google Scholar] [CrossRef]
- Cresson, P.; Ruitton, S.; Harmelin-Vivien, M. Artificial reefs do increase secondary biomass production: Mechanisms evidenced by stable isotopes. Mar. Ecol. Prog. Ser. 2014, 509, 15–26. [Google Scholar] [CrossRef]
- La Mesa, G.; Longobardi, A.; Sacco, F.; Marino, G. First Release of Hatchery Juveniles of the Dusky Grouper Epinephelus marginatus (Lowe, 1834) (Serranidae: Teleostei) at Artificial Reefs in the Mediterranean: Results from a Pilot Study. Sci. Mar. 2008, 72, 743–756. [Google Scholar] [CrossRef]
- Hunter, W.R.; Sayer, M.D.J.J. The comparative effects of habitat complexity on faunal assemblages of northern temperate artificial and natural reefs. ICES J. Mar. Sci. 2009, 66, 691–698. [Google Scholar] [CrossRef]
- Wilhelmsson, D.; Malm, T. Fouling assemblages on offshore wind power plants and adjacent substrata. Estuar. Coast. Shelf Sci. 2008, 79, 459–466. [Google Scholar] [CrossRef]
- Wilhelmsson, D.; Malm, T.; Öhman, M.C. The influence of offshore windpower on demersal fish. ICES J. Mar. Sci. 2006, 63, 775–784. [Google Scholar] [CrossRef]
- Lindeboom, H.J.; Kouwenhoven, H.J.; Bergman, M.J.N.; Bouma, S.; Brasseur, S.; Daan, R.; Fijn, R.C.; De Haan, D.; Dirksen, S.; Van Hal, R.; et al. Short-term ecological effects of an offshore wind farm in the Dutch coastal zone; A compilation. Environ. Res. Lett. 2011, 6, 035101. [Google Scholar] [CrossRef]
- De Troch, M.; Reubens, J.T.; Heirman, E.; Degraer, S.; Vincx, M. Energy profiling of demersal fish: A case-study in wind farm artificial reefs. Mar. Environ. Res. 2013, 92, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Bergman, M.J.N.; Ubels, S.M.; Duineveld, G.C.A.; Meesters, E.W.G. Effects of a 5-year trawling ban on the local benthic community in a wind farm in the Dutch coastal zone. ICES J. Mar. Sci. 2015, 72, 962–972. [Google Scholar] [CrossRef]
- Krone, R.; Dederer, G.; Kanstinger, P.; Krämer, P.; Schneider, C.; Schmalenbach, I. Mobile demersal megafauna at common offshore wind turbine foundations in the German Bight (North Sea) two years after deployment—Increased production rate of Cancer pagurus. Mar. Environ. Res. 2017, 123, 53–61. [Google Scholar] [CrossRef]
- van Hal, R.; Griffioen, A.B.; van Keeken, O.A. Changes in fish communities on a small spatial scale, an effect of increased habitat complexity by an offshore wind farm. Mar. Environ. Res. 2017, 126, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Reubens, J.T.; Degraer, S.; Vincx, M. Aggregation and feeding behaviour of pouting (Trisopterus luscus) at wind turbines in the Belgian part of the North Sea. Fish. Res. 2011, 108, 223–227. [Google Scholar] [CrossRef]
- Reubens, J.T.; Pasotti, F.; Degraer, S.; Vincx, M. Residency, site fidelity and habitat use of atlantic cod (Gadus morhua) at an offshore wind farm using acoustic telemetry. Mar. Environ. Res. 2013, 90, 128–135. [Google Scholar] [CrossRef]
- Andersson, M.H.; Öhman, M.C. Fish and sessile assemblages associated with wind-turbine constructions in the Baltic Sea. Mar. Freshw. Res. 2010, 61, 642–650. [Google Scholar] [CrossRef]
- Stenberg, C.; Støttrup, J.G.; van Deurs, M.; Berg, C.W.; Dinesen, G.E.; Mosegaard, H.; Grome, T.M.; Leonhard, S.B. Long-term effects of an offshore wind farm in the North Sea on fish communities. Mar. Ecol. Prog. Ser. 2015, 528, 257–265. [Google Scholar] [CrossRef]
- van Deurs, M.; Grome, T.M.; Kaspersen, M.; Jensen, H.; Stenberg, C.; Sørensen, T.K.; Støttrup, J.; Warnar, T.; Mosegaard, H. Short- and long-term effects of an offshore wind farm on three species of sandeel and their sand habitat. Mar. Ecol. Prog. Ser. 2012, 458, 169–180. [Google Scholar] [CrossRef]
- Langhamer, O.; Dahlgren, T.G.; Rosenqvist, G. Effect of an offshore wind farm on the viviparous eelpout: Biometrics, brood development and population studies in Lillgrund, Sweden. Ecol. Indic. 2018, 84, 1–6. [Google Scholar] [CrossRef]
- Fabi, G.; Fiorentini, L. Comparison between an artificial reef and a control site in the Adriatic Sea: Analysis of four years of monitoring. Bull. Mar. Sci. 1994, 55, 538–558. [Google Scholar]
- Bombace, G.; Fabi, G.; Fiorentini, L.; Speranza, S. Analysis of The Efficacy of Artificial Reefs Located in Five Different Areas of the Adriatic Sea. Bull. Mar. Sci. 1994, 55, 559–580. [Google Scholar]
- Wilber, D.H.; Carey, D.A.; Griffin, M. Flatfish habitat use near North America’s first offshore wind farm. J. Sea Res. 2018, 139, 24–32. [Google Scholar] [CrossRef]
- Reubens, J.T.; Degraer, S.; Vincx, M. The ecology of benthopelagic fishes at offshore wind farms: A synthesis of 4 years of research. Hydrobiologia 2014, 727, 121–136. [Google Scholar] [CrossRef]
- Reubens, J.T.; Vandendriessche, S.; Zenner, A.N.; Degraer, S.; Vincx, M. Offshore wind farms as productive sites or ecological traps for gadoid fishes?—Impact on growth, condition index and diet composition. Mar. Environ. Res. 2013, 90, 66–74. [Google Scholar] [CrossRef]
- Bat, L.; Sezgin, M.; Sahin, F. Impacts of OWF installations on fisheries: A Literature Review. J. Coast. Life Med. 2013, 1, 241–252. [Google Scholar]
- Bohnsack, J.A.; Harper, D.E.; McClellan, D.B.; Hulsbeck, M. Effects of Reef Size on Colonization and Assemblage Structure of Fishes at Artificial Reefs Off Southeastern Florida, U.S.A. Bull. Mar. Sci. 1994, 55, 796–823. [Google Scholar]
- Methratta, E.T.; Dardick, W.R.; Methratta, E.T.; Dardick, W.R. Meta-Analysis of Finfish Abundance at Offshore Wind Farms. Rev. Fish. Sci. Aquac. 2019, 27, 242–260. [Google Scholar] [CrossRef]
- Yu, J.; Chen, P.; Tang, D.; Qin, C. Ecological effects of artificial reefs in Daya Bay of China observed from satellite and in situ measurements. Adv. Sp. Res. 2015, 55, 2315–2324. [Google Scholar] [CrossRef]
- Fabi, G.; Manoukian, S.; Spagnolo, A. Feeding behavior of three common fishes at an artificial reef in the northern Adriatic Sea. Bull. Mar. Sci. 2006, 78, 39–56. [Google Scholar]
- Aneer, G.; Nellbring, S. A SCUBA-diving investigation of Baltic herring (Clupea harengus membras L.) spawning grounds in the Asko-Landsort area, northern Baltic proper. J. Fish Biol. 1982, 21, 433–442. [Google Scholar] [CrossRef]
- Johannessen, A. Recruitment studies of herring (Clupea harengus L.) in Lindaaspollene, western Norway. Fisk. Skr. Ser. Havundersøkelser 1986, 18, 139–240. [Google Scholar]
- Kääriä, J.; Rajasilta, M.; Kurkilahti, M.; Soikkeli, M. Spawning bed selection by the Baltic herring (Clupea harengus membras) in the Archipelago of SW Finland. ICES J. Mar. Sci. 1997, 54, 917–923. [Google Scholar] [CrossRef]
- Šaškov, A.; Šiaulys, A.; Bučas, M.; Daunys, D. Baltic herring (Clupea harengus membras) spawning grounds on the Lithuanian coast: Current status and shaping factors. Oceanologia 2014, 56, 789–804. [Google Scholar] [CrossRef][Green Version]
- Brickhill, M.J.; Lee, S.Y.; Connolly, R.M. Fishes associated with artificial reefs: Attributing changes to attraction or production using novel approaches. J. Fish Biol. 2005, 67, 53–71. [Google Scholar] [CrossRef]
- Pickering, H.; Whitmarsh, D. Artificial reefs and fisheries exploitation: A review of the “attraction versus production” debate, the influence of design and its significance for policy. Fish. Res. 1997, 31, 39–59. [Google Scholar] [CrossRef]
- Granneman, J.E.; Steele, M.A. Fish Growth, Reproduction, and Tissue Production on Artificial Reefs Relative to Natural Reefs. ICES J. Mar. Sci. 2014, 71, 2494–2504. [Google Scholar] [CrossRef]
- Cresson, P.; Le Direach, L.; Rouanet, E.; Goberville, E.; Astruch, P.; Ourgaud, M.; Harmelin-Vivien, M. Functional traits unravel temporal changes in fish biomass production on artificial reefs. Mar. Environ. Res. 2019, 145, 137–146. [Google Scholar] [CrossRef]
- Roa-Ureta, R.H.; Santos, M.N.; Leitão, F. Modelling long-term fisheries data to resolve the attraction versus production dilemma of artificial reefs. Ecol. Modell. 2019, 407, 108727. [Google Scholar] [CrossRef]
- Wilson, J.C.; Elliott, M.; Cutts, N.D.; Mander, L.; Mendão, V.; Perez-Dominguez, R.; Phelps, A. Coastal and offshore wind energy generation: Is it environmentally benign? Energies 2010, 3, 1383–1422. [Google Scholar] [CrossRef]
- Osenberg, C.W.; St. Mary, C.M.; Wilson, J.A.; Lindberg, W.J. A quantitative framework to evaluate the attraction-production controversy. ICES J. Mar. Sci. 2002, 59, S214–S221. [Google Scholar] [CrossRef]
- Scarcella, G.; Grati, F.; Bolognini, L.; Domenichetti, F.; Malaspina, S.; Manoukian, S.; Polidori, P.; Spagnolo, A.; Fabi, G. Time-series analyses of fish abundance from an artificial reef and a reference area in the central-Adriatic Sea. J. Appl. Ichthyol. 2015, 31, 74–85. [Google Scholar] [CrossRef]
- Schwartzbach, A.; Behrens, J.; Svendsen, J. Atlantic cod Gadus morhua save energy on stone reefs: Implications for the attraction versus production debate in relation to reefs. Mar. Ecol. Prog. Ser. 2020, 635, 81–87. [Google Scholar] [CrossRef]
- Gill, A.B. Offshore renewable energy: Ecological implications of generating electricity in the coastal zone. J. Appl. Ecol. 2005, 42, 605–615. [Google Scholar] [CrossRef]
- Janßen, H.; Augustin, C.B.; Hinrichsen, H.H.; Kube, S. Impact of secondary hard substrate on the distribution and abundance of Aurelia aurita in the western Baltic Sea. Mar. Pollut. Bull. 2013, 75, 224–234. [Google Scholar] [CrossRef]
- Makabe, R.; Furukawa, R.; Takao, M.; Uye, S.I. Marine artificial structures as amplifiers of Aurelia aurita s.l. blooms: A case study of a newly installed floating pier. J. Oceanogr. 2014, 70, 447–455. [Google Scholar] [CrossRef]
- De Mesel, I.; Kerckhof, F.; Norro, A.; Rumes, B.; Degraer, S. Succession and seasonal dynamics of the epifauna community on offshore wind farm foundations and their role as stepping stones for non-indigenous species. Hydrobiologia 2015, 756, 37–50. [Google Scholar] [CrossRef]
- Tidbury, H.; Taylor, N.; Van der Molen, J.; Garcia, L.; Posen, P.; Gill, A.; Lincoln, S.; Judd, A.; Hyder, K. Social network analysis as a tool for marine spatial planning: Impacts of decommissioning on connectivity in the North Sea. J. Appl. Ecol. 2020, 57, 566–577. [Google Scholar] [CrossRef]
- Degraer, S.; Brabant, R. Offshore wind farms in the Belgian part of the North Sea: State of the art after two years of environmental monitoring; Royal Belgian Institute for Natural Sciences, Management Unit of the North Sea Mathematical Models, Marine Ecosystem Management Unit. 2009; pp. 1–287. Available online: https://tethys.pnnl.gov/publications/offshore-wind-farms-belgian-part-north-sea-state-art-after-two-years-environmental (accessed on 8 May 2020).
- Southward, A.J.; Hawkins, S.J.; Burrows, M.T. Seventy years’ observations of changes in distribution and abundance of zooplankton and intertidal organisms in the western English Channel in relation to rising sea temperature. J. Therm. Biol. 1995, 20, 127–155. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Terwin, J.R.; Whitlatch, R.B.; Osman, R.W. Linking climate change and biological invasions: Ocean warming facilitates nonindigenous species invasions. Proc. Natl. Acad. Sci. USA 2002, 99, 15497–15500. [Google Scholar] [CrossRef]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef]
- Coolen, J.W.P.; Lengkeek, W.; Degraer, S.; Kerckhof, F.; Kirkwood, R.J.; Lindeboom, H.J. Distribution of the invasive Caprella mutica Schurin, 1935 and native Caprella linearis (Linnaeus, 1767) on artificial hard substrates in the North Sea: Separation by habitat. Aquat. Invasions 2016, 11, 437–449. [Google Scholar] [CrossRef]
- Leonhard, S.B.; Pedersen, J. Benthic Communities at Horns Rev Before, During and After Construction of Horns Rev Offshore Wind Farm—Annual Report 2005. Vattenfall 2006, 84, 134. [Google Scholar]
- Adams, T.P.; Miller, R.G.; Aleynik, D.; Burrows, M.T. Offshore marine renewable energy devices as stepping stones across biogeographical boundaries. J. Appl. Ecol. 2014, 51, 330–338. [Google Scholar] [CrossRef]
- Maar, M.; Bolding, K.; Petersen, J.K.; Hansen, J.L.S.; Timmermann, K. Local effects of blue mussels around turbine foundations in an ecosystem model of Nysted off-shore wind farm, Denmark. J. Sea Res. 2009, 62, 159–174. [Google Scholar] [CrossRef]
- Wilson, J.C.; Elliott, M. The habitat-creation potential of offshore wind farms. Wind Energy 2009, 12, 203–212. [Google Scholar] [CrossRef]
- Reubens, J.T.; De Rijcke, M.; Degraer, S.; Vincx, M. Diel variation in feeding and movement patterns of juvenile Atlantic cod at offshore wind farms. J. Sea Res. 2014, 85, 214–221. [Google Scholar] [CrossRef]
- Kerckhof, F.; Rumes, B.; Jacques, T.; Degraer, S.; Norro, A. Early development of the subtidal marine biofouling on a concrete offshore windmill foundation on the Thornton Bank (southern North Sea): First monitoring results. Underw. Technol. 2010, 29, 137–149. [Google Scholar] [CrossRef]
- Hixon, M.A.; Brostoff, W.N. Substrate characteristics, fish grazing, and epibenthic assemblages off Hawaii. Bull. Mar. Sci. 1985, 37, 200–213. [Google Scholar]
- Beisiegel, K.; Tauber, F.; Gogina, M.; Zettler, M.L.; Darr, A. The potential exceptional role of a small Baltic boulder reef as a solitary habitat in a sea of mud. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 321–328. [Google Scholar] [CrossRef]
- Bortone, S.A.; Cody, R.P.; Turpin, R.K.; Bundrick, C.M. The impact of artificial-reef fish assemblages on their potential forage area. Ital. J. Zool. 1998, 65, 265–267. [Google Scholar] [CrossRef]
- Rosemond, R.C.; Paxton, A.B.; Lemoine, H.R.; Fegley, S.R.; Peterson, C.H. Fish use of reef structures and adjacent sand flats: Implications for selecting minimum buffer zones between new artificial reefs and existing reefs. Mar. Ecol. Prog. Ser. 2018, 587, 187–199. [Google Scholar] [CrossRef]
- Shen, W.Z.; Mikkelsen, R.F. Study on wind turbine arrangement for offshore wind farms. In Proceedings of the ICOWEOE-2011, Beijing, China, 31 October–2 November 2011. [Google Scholar]
- Leitão, F.; Santos, M.N.; Erzini, K.; Monteiro, C.C. Fish assemblages and rapid colonization after enlargement of an artificial reef off the Algarve coast (Southern Portugal). Mar. Ecol. 2008, 29, 435–448. [Google Scholar] [CrossRef]
- Ambrose, R.F.; Swarbrick, S.L. Comparison of fish assemblages on artificial and natural reefs off the coast of southern California. Bull. Mar. Sci. 1989, 44, 718–733. [Google Scholar]
- Champion, C.; Suthers, I.M.; Smith, J.A. Zooplanktivory is a key process for fish production on a coastal artificial reef. Mar. Ecol. Prog. Ser. 2015, 541, 1–14. [Google Scholar] [CrossRef]
- Ogawa, S.; Takeuchi, R.; Hattori, H. An estimate for the optimum size of artificial reefs. Bull. Jpn. Soc. Fish. Oceanogr. 1977, 30, 39–45. [Google Scholar]
- Bohnsack, J.A.; Sutherland, D.L. Artificial Reef Research: A Review with Recommendations for Future Priorities. Bull. Mar. Sci. 1985, 37, 11–39. [Google Scholar]
- Charbonnel, E.; Serre, C.; Ruitton, S.; Harmelin, J.G.; Jensen, A. Effects of increased habitat complexity on fish assemblages associated with large artificial reef units (French Mediterranean coast). ICES J. Mar. Sci. 2002, 59, 208–213. [Google Scholar] [CrossRef]
- Hixon, M.A.; Beets, J.P. Predation, Prey Refuges, and the Structure of Coral-Reef Fish Assemblages. Ecol. Monogr. 1993, 63, 77–101. [Google Scholar] [CrossRef]
- Crowder, L.B.; Cooper, W.E. Habitat Structural Complexity and the Interaction Between Bluegills and Their Prey. Ecology 1982, 63, 1802–1813. [Google Scholar] [CrossRef]
- Davis, K.L.; Coleman, M.A.; Connell, S.D.; Russell, B.D.; Gillanders, B.M.; Kelaher, B.P. Ecological performance of construction materials subject to ocean climate change. Mar. Environ. Res. 2017, 131, 177–182. [Google Scholar] [CrossRef]
- Walker, B.K.; Henderson, B.; Spieler, R.E.; Beach, M.; Walker, B.K.; Henderson, B.; Spieler, R.E. Fish Assemblages Associated with Artificial Reefs of Concrete Aggregates or Quarry Stone Offshore Miami Beach, Florida, USA. Aquat. Living Resour. 2002, 15, 95–105. [Google Scholar] [CrossRef]
- Carr, M.H.; Hixon, M.A. Artificial Reefs: The Importance of Comparisons with Natural Reefs. Fisheries 1997, 22, 28–33. [Google Scholar] [CrossRef]
- Brandt, M.J.; Diederichs, A.; Betke, K.; Nehls, G. Responses of harbour porpoises to pile driving at the Horns Rev II offshore wind farm in the Danish North Sea. Mar. Ecol. Prog. Ser. 2011, 421, 205–216. [Google Scholar] [CrossRef]
- Teilmann, J.; Carstensen, J. Negative long term effects on harbour porpoises from a large scale offshore wind farm in the Baltic—Evidence of slow recovery. Environ. Res. Lett. 2012, 7, 045101. [Google Scholar] [CrossRef]
- Furness, R.W.; Wade, H.M.; Masden, E.A. Assessing vulnerability of marine bird populations to offshore wind farms. J. Environ. Manag. 2013, 119, 56–66. [Google Scholar] [CrossRef]
- Russell, D.J.F.; Brasseur, S.M.J.M.; Thompson, D.; Hastie, G.D.; Janik, V.M.; Aarts, G.; McClintock, B.T.; Matthiopoulos, J.; Moss, S.E.W.; McConnell, B. Marine mammals trace anthropogenic structures at sea. Curr. Biol. 2014, 24, 638–639. [Google Scholar] [CrossRef]
- Spagnolo, A.; Fabi, G.; Manoukian, S.; Panfili, M. Benthic community settled on an artificial reef in the Western Adriatic Sea (Italy). Rapp. Comm. Int. Pour l’Exploration Sci. Mer Médititerranée 2004, 37, 552. [Google Scholar]
- Verdura, J.; Sales, M.; Ballesteros, E.; Cefalì, M.E.; Cebrian, E. Restoration of a Canopy-Forming Alga Based on Recruitment Enhancement: Methods and Long-Term Success Assessment. Front. Plant Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Fredriksen, S.; Filbee-Dexter, K.; Norderhaug, K.M.; Steen, H.; Bodvin, T.; Coleman, M.A.; Moy, F.; Wernberg, T. Green gravel: A novel restoration tool to combat kelp forest decline. Sci. Rep. 2020, 10, 1–7. [Google Scholar] [CrossRef]
- Scheidat, M.; Tougaard, J.; Brasseur, S.; Carstensen, J.; Van Polanen Petel, T.; Teilmann, J.; Reijnders, P. Harbour porpoises (Phocoena phocoena) and wind farms: A case study in the Dutch North Sea. Environ. Res. Lett. 2011, 6, 025102. [Google Scholar] [CrossRef]
- Hammill, M.O.; Stenson, G.B. Estimated Prey Consumption by Harp seals (Phoca groenlandica), Hooded seals (Cystophora cristata), Grey seals (Halichoerus grypus) and Harbour seals (Phoca vitulina) in Atlantic Canada. J. Northw. Atl. Fish. Sci. 1999, 26, 1–23. [Google Scholar] [CrossRef]
- Chouinard, G.A.; Swain, D.P.; Hammill, M.O.; Poirier, G.A. Covariation between grey seal (Halichoerus grypus) abundance and natural mortality of cod (Gadus morhua) in the southern Gulf of St. Lawrence. Can. J. Fish. Aquat. Sci. 2005, 62, 1991–2000. [Google Scholar] [CrossRef]
- Harlin, M.M.; Lindbergh, J.M. Selection of substrata by seaweeds: Optimal surface relief. Mar. Biol. 1977, 40, 33–40. [Google Scholar] [CrossRef]
- Beserra Azevedo, F.B.; Carloni, G.G.; Vercosa Carvalheira, L. Colonization of benthic organisms of different artificial substratum in Ilha Grande Bay, Rio de Janeiro. Braz. Arch. Biol. Technol. 2006, 49, 263–275. [Google Scholar] [CrossRef]
- Rilov, G.; Benayahu, Y. Vertical artificial structures as an alternative habitat for coral reef fishes in disturbed environments. Mar. Environ. Res. 1998, 45, 431–451. [Google Scholar] [CrossRef]
- De La Fuente, G.; Chiantore, M.; Asnaghi, V.; Kaleb, S.; Falace, A. First ex situ outplanting of the habitat-forming seaweed Cystoseira amentacea var. stricta from a restoration perspective. PeerJ 2019, 7, e7290. [Google Scholar] [CrossRef]
- Quaas, Z.; Harasti, D.; Gaston, T.F.; Platell, M.E.; Fulton, C.J. Influence of habitat condition on shallow rocky reef fish community structure around islands and headlands of a temperate marine protected area. Mar. Ecol. Prog. Ser. 2019, 626, 1–13. [Google Scholar] [CrossRef]
- Jensen, A.; Wickins, J.; Bannister, C. The Potential Use of Artificial Reefs to Enhance Lobster Habitat. In Artificial Reefs in European Seas; Springer: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Spanier, E. What Are the Characteristics of a Good Artificial Reef for Lobsters? Crustaceana 1994, 67, 173–186. [Google Scholar] [CrossRef]
- Cobb, J.S. The Shelter-Related Behavior of the Lobster, Homarus Americanus. Ecology 1971, 52, 108–115. [Google Scholar] [CrossRef]
- Barry, J.; Wickins, J.F. A model for the number and sizes of crevices that can be seen on the exposed surface of submerged rock reefs. Environmetrics 1992, 3, 55–69. [Google Scholar] [CrossRef]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological Consequences of Ecosystem Fragmentation—A Review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Ries, L.; Fletcher, R.J.; Battin, J.; Sisk, T.D. Ecological responses to habitat edges: Mechanisms, models, and variability explained. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 491–522. [Google Scholar] [CrossRef]
- Jelbart, J.E.; Ross, P.M.; Connolly, R.M. Edge effects and patch size in seagrass landscapes: An experimental test using fish. Mar. Ecol. Prog. Ser. 2006, 319, 93–102. [Google Scholar] [CrossRef]
- Selgrath, J.C.; Hovel, K.A.; Wahle, R.A. Effects of habitat edges on American lobster abundance and survival. J. Exp. Mar. Biol. Ecol. 2007, 353, 253–264. [Google Scholar] [CrossRef]
- Smith, T.M.; Hindell, J.S.; Jenkins, G.P.; Connolly, R.M. Edge effects on fish associated with seagrass and sand patches. Mar. Ecol. Prog. Ser. 2008, 359, 203–213. [Google Scholar] [CrossRef]
- Malcolm, H.A.; Jordan, A.; Smith, S.D.A. Biogeographical and cross-shelf patterns of reef fish assemblages in a transition zone. Mar. Biodivers. 2010, 40, 181–193. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Graham, N.A.J.; Calnan, J.M.; MacNeil, M.A. Toward Pristine Biomass: Reef Fish Recovery in Coral Reef Marine Protected Areas in Kenya. Ecol. Appl. 2007, 17, 1055–1067. [Google Scholar] [CrossRef]
- Fukami, T.; Nakajima, M. Community assembly: Alternative stable states or alternative transient states? Ecol. Lett. 2011, 14, 973–984. [Google Scholar] [CrossRef]
- Kayal, M.; Lenihan, H.S.; Brooks, A.J.; Holbrook, S.J.; Schmitt, R.J.; Kendall, B.E. Predicting coral community recovery using multi-species population dynamics models. Ecol. Lett. 2019, 22, 605–615. [Google Scholar] [CrossRef]
- Schoonees, T.; Gijón Mancheño, A.; Scheres, B.; Bouma, T.J.; Silva, R.; Schlurmann, T.; Schüttrumpf, H. Hard Structures for Coastal Protection, Towards Greener Designs. Estuaries Coasts 2019, 42, 1709–1729. [Google Scholar] [CrossRef]
- Hammar, L.; Perry, D.; Gullström, M. Offshore Wind Power for Marine Conservation. Open J. Mar. Sci. 2016, 6, 66–78. [Google Scholar] [CrossRef]
- Díaz, D.; Mallol, S.; Parma, A.M.; Goñi, R. A 25-year marine reserve as proxy for the unfished condition of an exploited species. Biol. Conserv. 2016, 203, 97–107. [Google Scholar] [CrossRef]
- Kough, A.S.; Belak, C.A.; Paris, C.B.; Lundy, A.; Cronin, H.; Gnanalingam, G.; Hagedorn, S.; Skubel, R.; Weiler, A.C.; Stoner, A.W. Ecological spillover from a marine protected area replenishes an over-exploited population across an island chain. Conserv. Sci. Pract. 2019, 1, e17. [Google Scholar] [CrossRef]
- Burkhard, B.; Gee, K. Establishing the resilience of a coastal-marine social-ecological system to the installation of offshore wind farms. Ecol. Soc. 2012, 17, 42. [Google Scholar] [CrossRef]
- Hewitt, J.E.; Thrush, S.F. Monitoring for tipping points in the marine environment. J. Environ. Manag. 2019, 234, 131–137. [Google Scholar] [CrossRef]
- Mavraki, N.; Degraer, S.; Moens, T.; Vanaverbeke, J. Functional differences in trophic structure of offshore wind farm communities: A stable isotope study. Mar. Environ. Res. 2020, 157, 104868. [Google Scholar] [CrossRef]
- Claisse, J.T.; Pondella, D.J.; Love, M.; Zahn, L.A.; Williams, C.M.; Williams, J.P.; Bull, A.S. Oil platforms off California are among the most productive marine fish habitats globally. Proc. Natl. Acad. Sci. USA 2014, 111, 15462–15467. [Google Scholar] [CrossRef]
- Ajemian, M.J.; Wetz, J.J.; Shipley-Lozano, B.; Dale Shively, J.; Stunz, G.W. An analysis of artificial reef fish community structure along the northwestern Gulf of Mexico shelf: Potential impacts of “rigs-to-reefs” Programs. PLoS ONE 2015, 10, 1–22. [Google Scholar] [CrossRef]
- Todd, V.L.G.; Lavallin, E.W.; Macreadie, P.I. Quantitative analysis of fish and invertebrate assemblage dynamics in association with a North Sea oil and gas installation complex. Mar. Environ. Res. 2018, 142, 69–79. [Google Scholar] [CrossRef]
- McLean, D.L.; Taylor, M.D.; Giraldo, A.; Partridge, J.C. An assessment of fish and marine growth associated with an oil and gas platform jacket using an augmented remotely operated vehicle. Cont. Shelf Res. 2019, 179, 66–84. [Google Scholar] [CrossRef]
- Inger, R.; Attrill, M.J.; Bearhop, S.; Broderick, A.C.; James Grecian, W.; Hodgson, D.J.; Mills, C.; Sheehan, E.; Votier, S.C.; Witt, M.J.; et al. Marine renewable energy: Potential benefits to biodiversity? An urgent call for research. J. Appl. Ecol. 2009, 46, 1145–1153. [Google Scholar] [CrossRef]
- Dons, S.; Jensen, D.J.; Struve, A.; Nielsen, B. Vindeby Havmøllepark; DONG Energy. 2016, pp. 1–61. Available online: https://ens.dk/sites/ens.dk/files/Vindenergi/miljoevurdering_for_nedtagning_af_vindeby_havmoellpark_inkl_bilag.pdf (accessed on 8 May 2020).
- Bull, A.S.; Love, M.S. Worldwide oil and gas platform decommissioning: A review of practices and reefing options. Ocean Coast. Manag. 2019, 168, 274–306. [Google Scholar] [CrossRef]
- Fowler, A.M.; Jørgensen, A.M.; Svendsen, J.C.; Macreadie, P.I.; Jones, D.O.B.; Boon, A.R.; Booth, D.J.; Brabant, R.; Callahan, E.; Claisse, J.T.; et al. Environmental benefits of leaving offshore infrastructure in the ocean. Front. Ecol. Environ. 2018, 16, 571–578. [Google Scholar] [CrossRef]
- Smyth, K.; Christie, N.; Burdon, D.; Atkins, J.P.; Barnes, R.; Elliott, M. Renewables-to-reefs?—Decommissioning options for the offshore wind power industry. Mar. Pollut. Bull. 2015, 90, 247–258. [Google Scholar] [CrossRef]
- Topham, E.; McMillan, D. Sustainable decommissioning of an offshore wind farm. Renew. Energy 2016, 102, 470–480. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TechnicalTerms | artificial reef *, reef effect *, fish attraction device *, artificial structure *, scour *, scour * protection *, scour control *, wind power foundation *, offshore wind, wind farm foundation *, turbine foundation *, offshore energy, offshore wind energy structure *, artificial offshore construction *, offshore wind farm *, wind turbine *, rock armour * |
| Ecosystem-Related Terms | ecological effect *, impact *, sanctuary *, spillover, habitat *, habitat change *, habitat restor *, habitat creat *, species abundance *, biomass *, biodiversity, species composition *, species densit *, nurser *, recruitment, coloni *, migrat *, food availab *, ecological function *, aggregate *, productive *, enhanced habitat *, heterogeneity, feeding, spawn * OR habitat creat *, habitat connectivity, habitat complexity, habitat enhancement *, no trawling zone *, hard bottom, invasive species, alien species, nonindigenous, habitat fragment *, habitat degrad *, habitat loss *, hard substrate *, substrate * |
| Target Species Group Term | fish * |
| Negative Terms | tropical, subtropical, Caribbean, Indian Ocean |
| Criteria | Include | Exclude |
|---|---|---|
| Peer-reviewing | Peer-reviewed studies | Reviews & everything else |
| Text Language | English | Everything else |
| Years | All years (1900–2018) | - |
| Location | Temperate seas | Everything else |
| Intervention | OWFs and designated ARs | Accidental ARs, such as shipwrecks |
| Subject | Fish and fisheries | Everything else |
| Variable | Examples | % of AR Studies Reported in (% Out of 89) | % of OWF Studies Reported in (% Out of 26) |
|---|---|---|---|
| General | |||
| Type of Study | Scientific Paper, Report | 100 | 100 |
| Year published | 2000, 2018 | 100 | 100 |
| Location | Italy, North Sea, UK | 100 | 100 |
| Study Design | |||
| Sampling method | Visual census, Fishing | 100 | 100 |
| Study design | BACI *, Impact only | 92 | 96 |
| Temporal Scale | 10 years after deployment | 79 | 69 |
| Sampling Season | Summer, Winter | 82 | 92 |
| Structure | |||
| Spatial Scale | 700 m2 | 44 | 54 |
| Depth | 30 m | 76 | 61 |
| Material | Concrete, rock | 84 | 61 |
| Shape of material | Cube, boulder | 79 | 0 |
| Volume of material | 1m3 | 52 | 4 |
| Effects on Fish | |||
| Overall impact | +, +/−, − | 94 | 88 |
| Biomass/Abundance | +, +/−, − | 72 | 73 |
| Diversity | +, +/−, − | 48 | 38 |
| Shelter (Complexity) | Yes, No, Possibly | 34 | 85 |
| Spawning/Settlement | Yes, No, Possibly | 22 | 31 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glarou, M.; Zrust, M.; Svendsen, J.C. Using Artificial-Reef Knowledge to Enhance the Ecological Function of Offshore Wind Turbine Foundations: Implications for Fish Abundance and Diversity. J. Mar. Sci. Eng. 2020, 8, 332. https://doi.org/10.3390/jmse8050332
Glarou M, Zrust M, Svendsen JC. Using Artificial-Reef Knowledge to Enhance the Ecological Function of Offshore Wind Turbine Foundations: Implications for Fish Abundance and Diversity. Journal of Marine Science and Engineering. 2020; 8(5):332. https://doi.org/10.3390/jmse8050332
Chicago/Turabian StyleGlarou, Maria, Martina Zrust, and Jon C. Svendsen. 2020. "Using Artificial-Reef Knowledge to Enhance the Ecological Function of Offshore Wind Turbine Foundations: Implications for Fish Abundance and Diversity" Journal of Marine Science and Engineering 8, no. 5: 332. https://doi.org/10.3390/jmse8050332
APA StyleGlarou, M., Zrust, M., & Svendsen, J. C. (2020). Using Artificial-Reef Knowledge to Enhance the Ecological Function of Offshore Wind Turbine Foundations: Implications for Fish Abundance and Diversity. Journal of Marine Science and Engineering, 8(5), 332. https://doi.org/10.3390/jmse8050332