Biological Aspects of Juveniles of the Common Stingray, Dasyatis pastinaca (Linnaeus, 1758) (Elasmobranchii, Dasyatidae), from the Central Mediterranean Sea




Abstract
1. Introduction
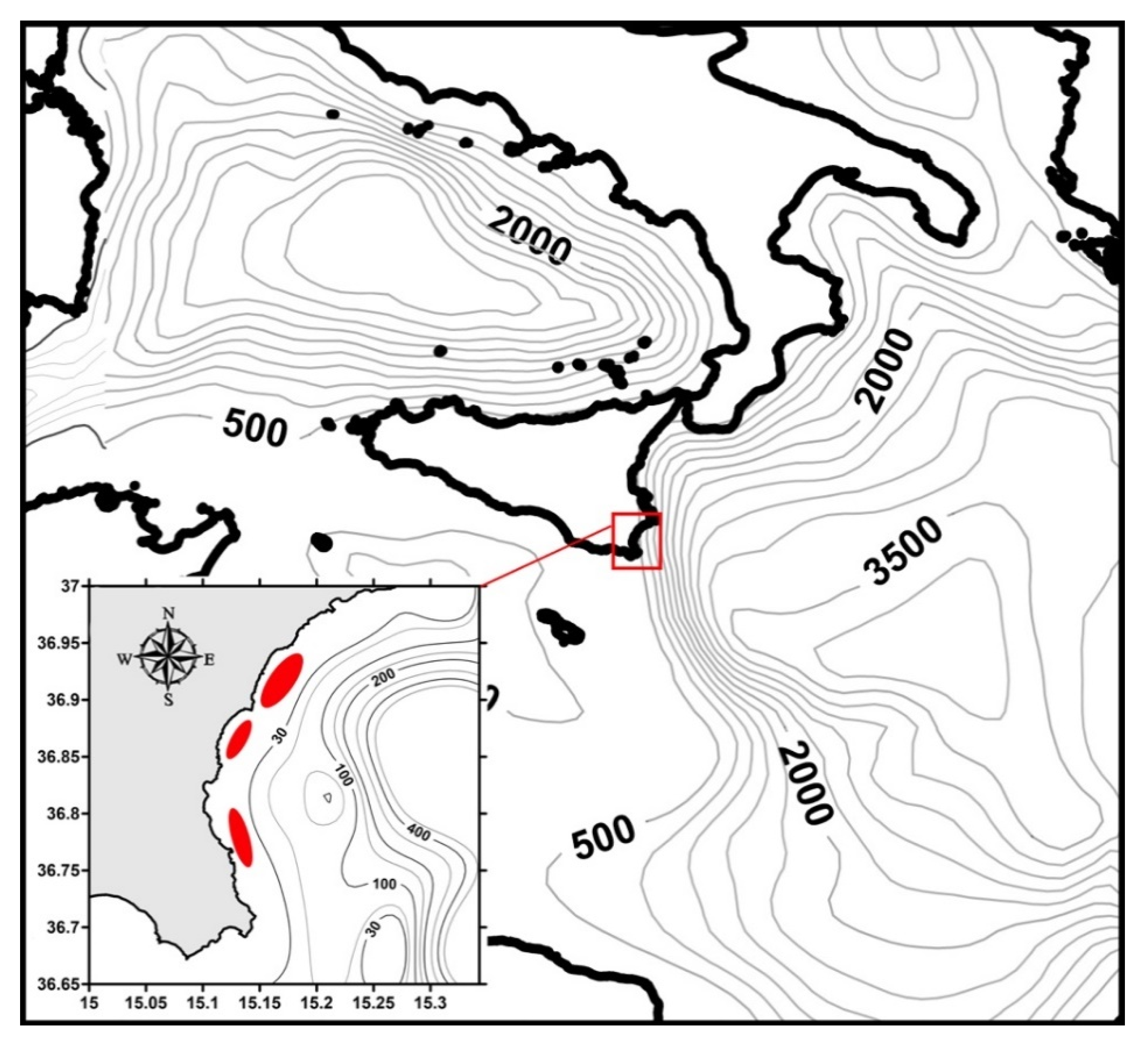
2. Materials and Methods
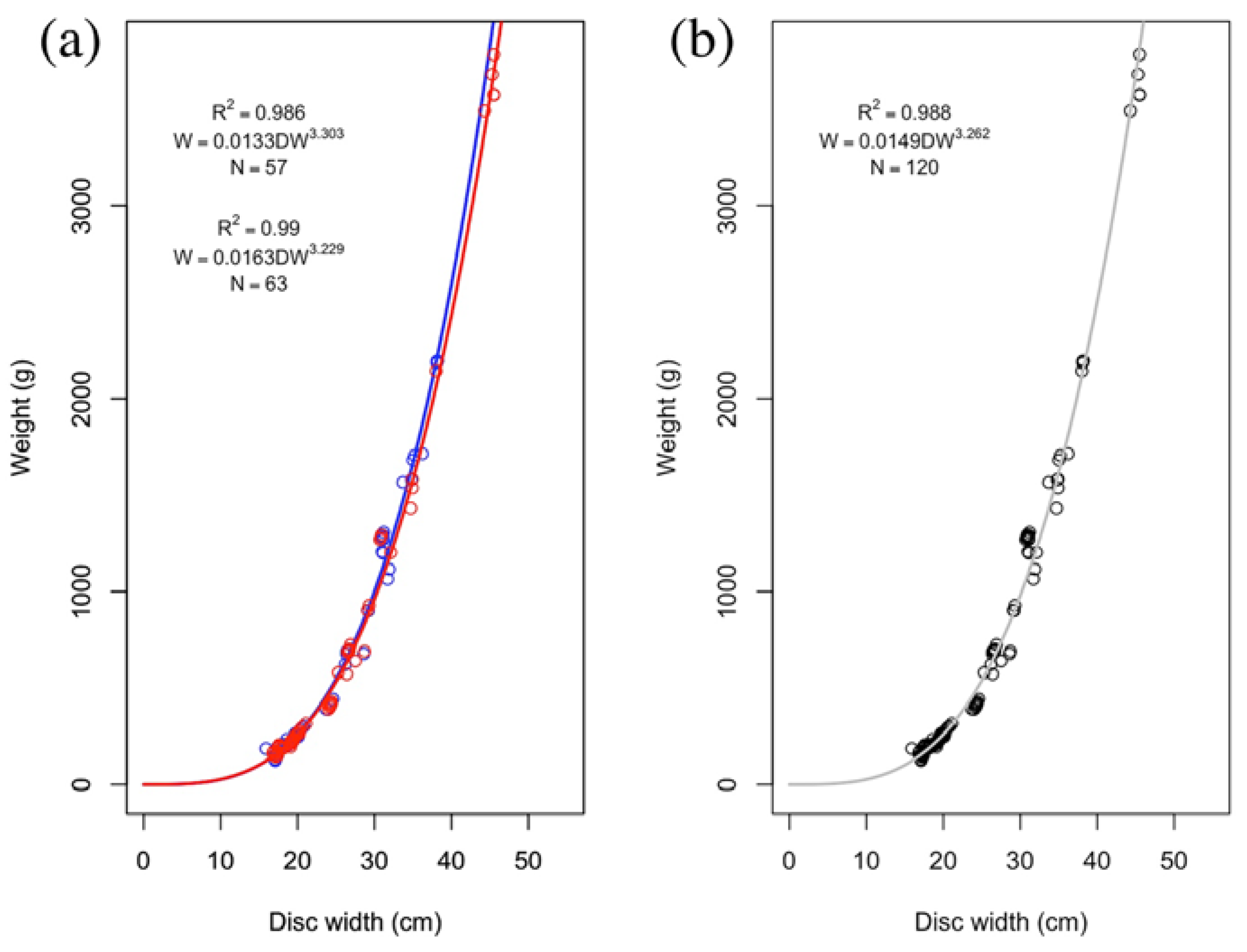
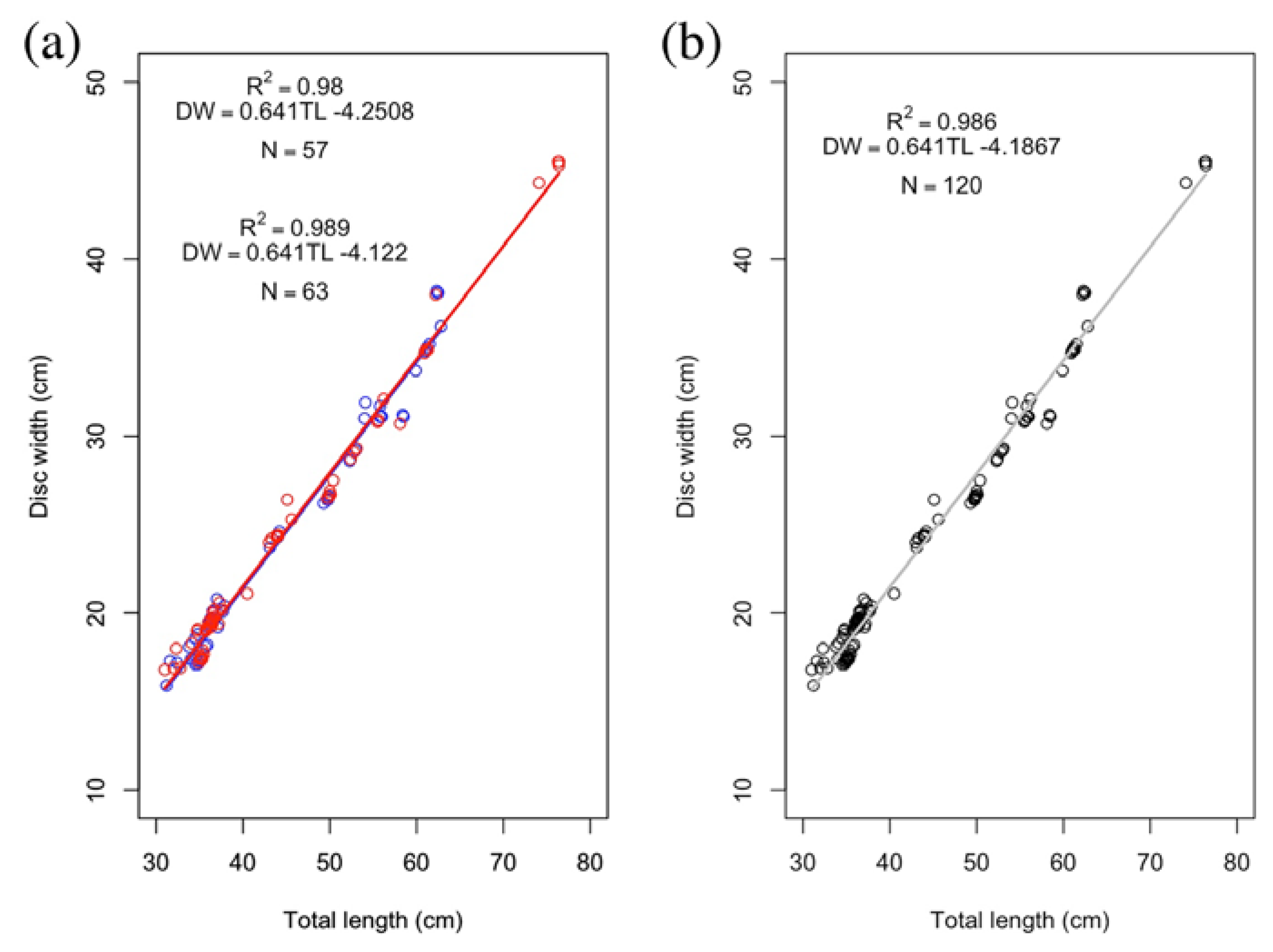
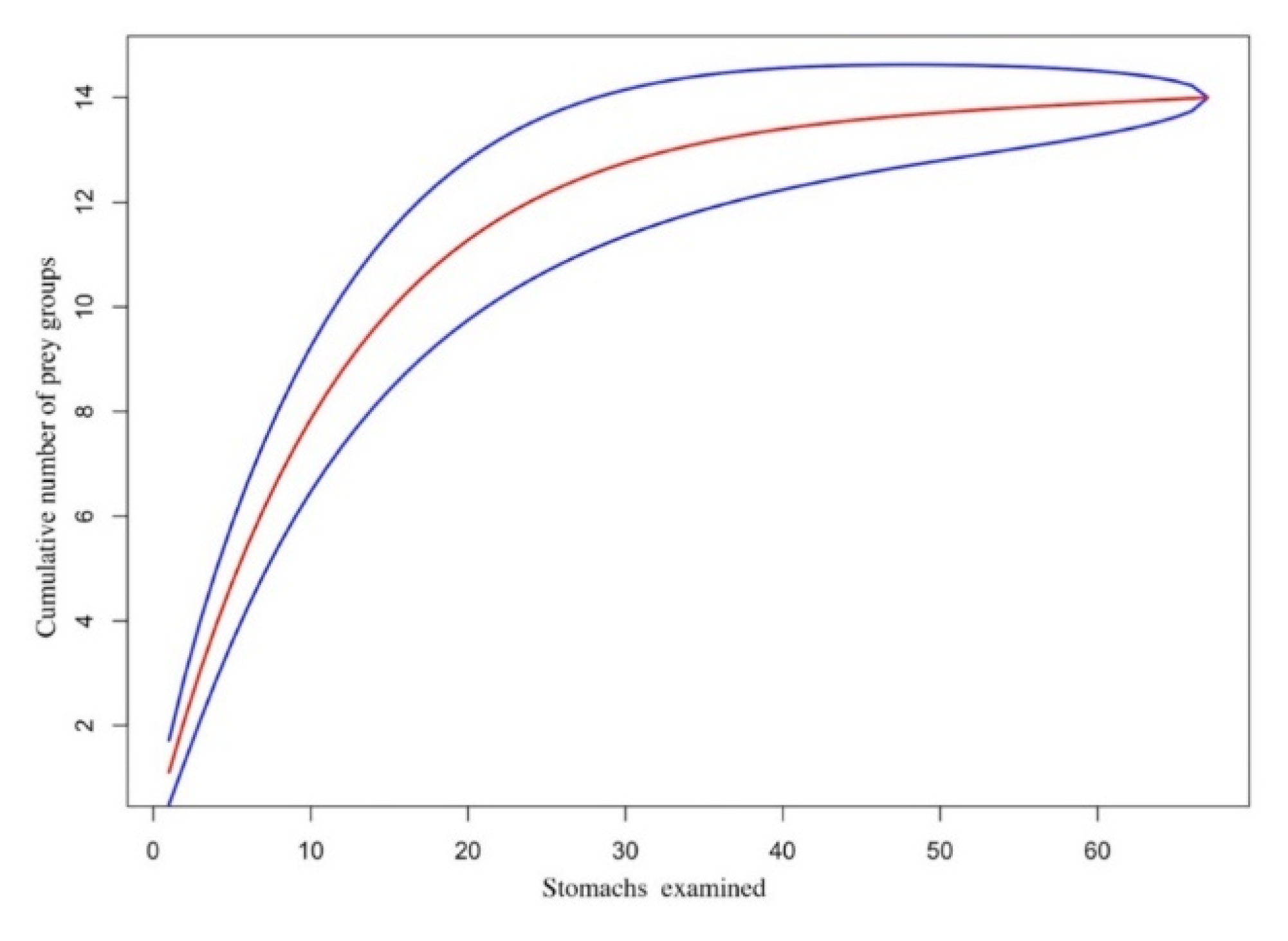
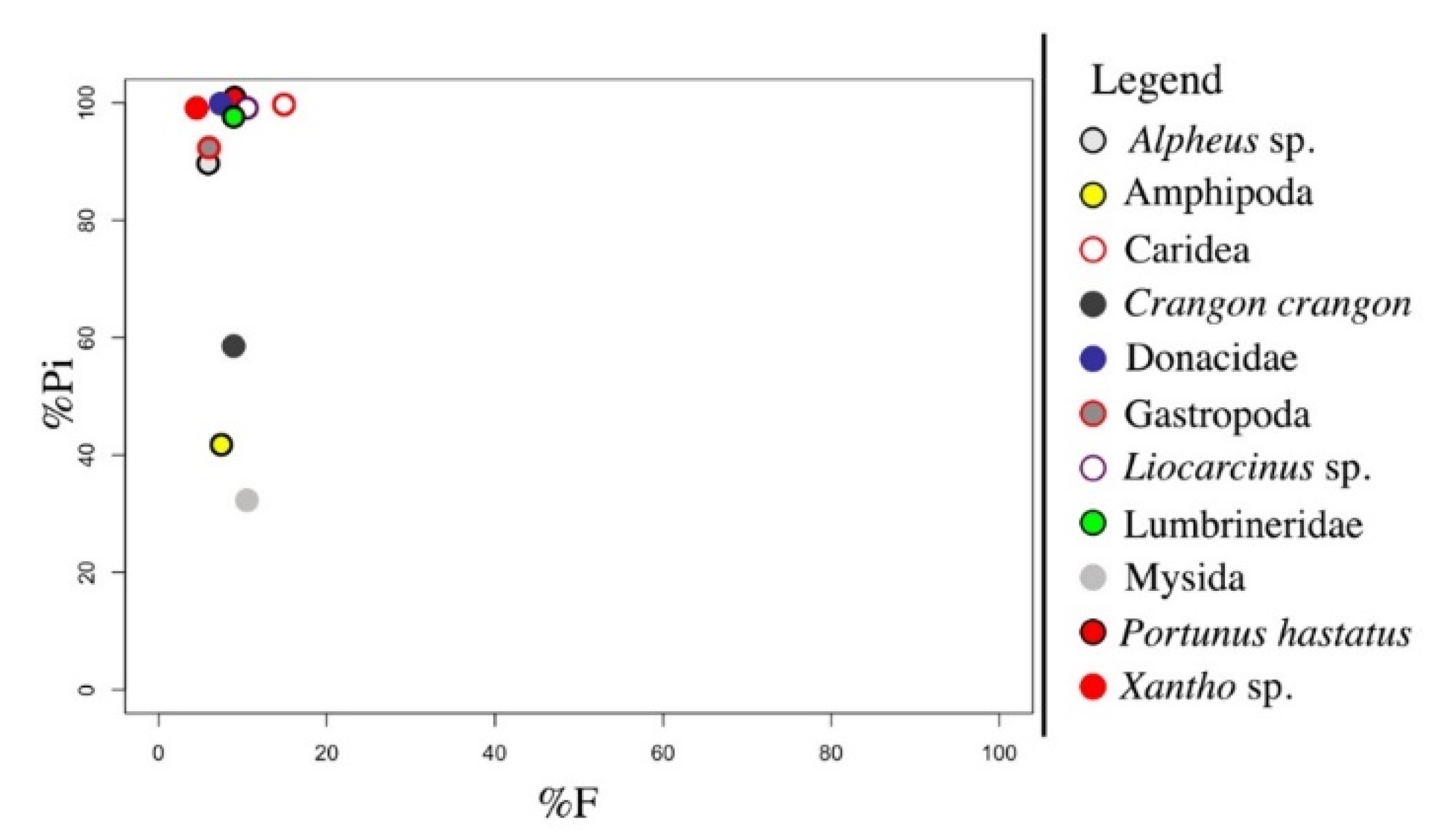
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Stevens, J.D.; Bonfil, R.; Dulvy, N.K.; Walker, P.A. The effects of fishing on sharks, rays and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES J. Mar. Sci. 2000, 57, 476–494. [Google Scholar] [CrossRef]
- Bornatowski, H.; Navia, A.F.; Braga, R.R.; Abilhoa, V.; Corrêa, M.F.M. Ecological importance of sharks and rays in a structural foodweb analysis in southern Brazil. ICES J. Mar. Sci. 2014, 71, 1586–1592. [Google Scholar] [CrossRef]
- Ragonese, S.; Cigala Fulgosi, F.; Bianchini, M.L.; Norrito, G.; Sinacori, G. Annotated check list of the skates (Chondrichthyes, Rajidae) in the Strait of Sicily (central Mediterranean Sea). Biol. Mar. Mediterr. 2003, 10, 874–881. [Google Scholar]
- White, W.T.; Blaber, S.J.M.; Craig, J.F. The current status of elasmobranchs: Biology, fisheries and conservation. J. Fish Biol. 2012, 80, 897–900. [Google Scholar] [CrossRef] [PubMed]
- Soldo, A. Status of sharks in the Mediterranean. Ann. Ser. Hist. Nat. 2003, 13, 191–200. [Google Scholar]
- Cavanagh, R.D.; Gibson, C. Overview of the Conservation Status of Cartilaginous Fishes (Chondrichthyans) in the Mediterranean Sea; IUCN: Gland, Switzerland; Malaga, Spain, 2007; Volume VI, p. 42. [Google Scholar]
- Bradai, M.N.; Saidi, B.; Enajjar, S. Elasmobranchs of the Mediterranean and Black Sea: Status, Ecology and Biology Bibliographic Analysis; FAO: Rome, Italy, 2012; Volume X, p. 103. [Google Scholar]
- Giovos, I.; Stoilas, V.O.; Al-Mabruk, S.A.A.; Doumpas, N.; Marakis, P.; Maximiadi, M.; Moutopoulos, D.; Kleitou, P.; Keramidas, I.; Tiralongo, F.; et al. Integrating local ecological knowledge, citizen science and long-term historical data for endangered species conservation: Additional records of angel sharks (Chondrichthyes: Squatinidae) in the Mediterranean Sea. Aquat. Conserv. 2019, 29, 881–890. [Google Scholar] [CrossRef]
- Giovos, I.; Arculeo, M.; Doumpas, N.; Katsada, D.; Maximiadi, M.; Mitsou, E.; Paravas, V.; Aga-Spyridopoulou, R.N.; Stoilas, V.O.; Tiralongo, F.; et al. Assessing multiple sources of data to detect illegal fishing, trade and mislabeling of elasmobranchs in Greek markets. Mar. Policy 2020, 112. [Google Scholar] [CrossRef]
- Vacchi, M.; Serena, F. Chondrichthyes. Biol. Mar. Mediterr. 2010, 17, 642–648. [Google Scholar]
- Notabartolo di Sciara, G.; Bianchi, I. Guida Degli Squali e Delle Razze del Mediterraneo; Franco Muzzio: Padova, Italy, 1998; p. 388. [Google Scholar]
- Özbek, E.Ö.; Çardak, M.; Kebapçioglu, T. Spatio-temporal patterns of abundance, biomass and length-weight relationships of Dasyatis species (Pisces: Dasyatidae) in the Gulf of Antalya, Turkey (Levantine Sea). J. Black Sea Medit. Environ. 2015, 21, 169–190. [Google Scholar]
- Serena, F.; Silvestri, R.; Voliani, A. Su una cattura accidentale di Taeniura grabata (E. Geoffroy Saint-Hilaire, 1817) (Chondrichthyes, Dasyatidae). Biol. Mar. Mediterr. 1999, 6, 617–618. [Google Scholar]
- Relini, G.; Biagi, F.; Serena, A.; Belluscio, M.T.; Spedicato, P.; Rinelli, P.; Follesa, M.C.; Piccinetti, C.; Ungaro, N.; Sion, L.; et al. I Selaci pescati con lo strascico nei mari italiani. Biol. Mar. Mediterr. 2000, 7, 347–384. [Google Scholar]
- Scacco, U.; Andaloro, F.; Campagnuolo, S.; Castriota, L.; Vacchi, M. Cartilaginous fishes as a component of trawl discard in Strait of Sicily (Elasmobranch Fisheries—Oral). NAFO SCR Doc 2002, 2, 87. [Google Scholar]
- Tiralongo, F.; Messina, G.; Lombardo, B.M. Discards of elasmobranchs in a trammel net fishery targeting cuttlefish, Sepia officinalis Linnaeus, 1758, along the coast of Sicily (central Mediterranean Sea). Reg. Stud. Mar. Sci. 2018, 20, 60–63. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase. Available online: http://www.fishbase.org (accessed on 15 January 2020).
- Serena, F.; Notabartolo di Sciara, G.; Ungaro, N. The IUCN Red List of Threatened Species. 2009. Available online: https://www.iucnredlist.org/species/161453/5427586 (accessed on 12 February 2020).
- Whitehead, P.J.P.; Bauchot, M.L.; Hureau, J.C.; Nielsen, J.; Tortonese, E. Fishes of the North-Eastern Atlantic and Mediterranean; Unesco: Paris, France, 1984; Volume 1-III, p. 1473. [Google Scholar]
- Fischer, W.; Bauchot, M.L.; Schneider, M. Fiches FAO D’indetification des Espèces Pour le Besoins de la Peche, Méditerranée et Mer Noire; FAO: Rome, Italy, 1987. [Google Scholar]
- Fuochi, V.; Li Volti, G.; Camiolo, G.; Tiralongo, F.; Giallongo, C.; Distefano, A.; Petronio, G.; Barbagallo, I.; Viola, M.; Furneri, P.M.; et al. Antimicrobial and anti-proliferative effects of skin mucus derived from Dasyatis pastinaca (Linnaeus, 1758). Mar. Drugs 2017, 15, 342. [Google Scholar] [CrossRef] [PubMed]
- Hyslop, E.J. Stomach content analysis, a review of methods and their application. J. Fish. Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Carrassòn, M.; Matallanas, J.; Casadevall, M. Feeding strategies of deep-water morids on the western Mediterranean slope. Deep Sea Res. 1997, 44, 1685–1699. [Google Scholar] [CrossRef]
- N’Da, K. Regime alimentaire du rouget de roche Mullus surmuletus (Mullidae) dans le nord du golfe de Gascogne. Cybium 1992, 16, 159–168. [Google Scholar]
- Costello, M.J. Predator feeding strategy and prey importance: A new geographical analysis. J. Fish. Biol. 1990, 36, 261–263. [Google Scholar] [CrossRef]
- Amundsen, P.A.; Gabler, H.M.; Staldvik, F.J. A new approach to graphical analysis of feeding strategy from stomach contents data-modification of the Costello (1990) method. J. Fish. Biol. 1996, 48, 607–614. [Google Scholar] [CrossRef]
- Krebs, J.C. Ecological Methodology; Harper & Row: New York, NY, USA, 1989; p. 620. [Google Scholar]
- Novakowski, G.C.; Hahn, N.S.; Fugi, R. Diet seasonality and food overlap of the fish assemblage in a pantanal pond. Neotrop. Ichthyol. 2008, 6, 567–576. [Google Scholar] [CrossRef]
- Brown, S.C.; Bizzarro, J.J.; Cailliet, G.M.; Ebert, D.A. Breaking with tradition: Redefining measures for diet description with a case study of the Aleutian skate Bathyraja aleutica (Gilbert 1869). Environ. Biol. Fish. 2012, 95, 3–20. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 30 November 2018).
- Yigin, C.C.; Ismen, A. Age, growth and reproduction of the common stingray, Dasyatis pastinaca from the North Aegean Sea. Mar. Biol. Res. 2012, 8, 644–653. [Google Scholar] [CrossRef]
- Jones, R.E.; Petrell, R.J.; Pauly, D. Using modified length-weight relationships to assess the condition of fish. Aquacult. Eng. 1999, 20, 261–276. [Google Scholar] [CrossRef]
- Moutopoulos, D.K.; Stergiou, K.I. Length-weight and length-length relationships of fish species from Aegean Sea (Greece). J. Appl. Ichthyol. 2002, 18, 200–203. [Google Scholar] [CrossRef]
- Vargas-Caro, C.; Bustamante, C.; Lamilla, J.; Bennett, M.B. A review of the longnose skates Zearaja chilensis and Dipturus trachyderma (Rajiformes: Rajidae). Univ. Sci. 2015, 20, 321–359. [Google Scholar] [CrossRef]
- Yeldan, H.; Avsar, D.; Manasirh, M. Age, growth and feeding of the common stingray (Dasyatis pastinaca, L. 1758) in the Cilician coastal basin, northeastern Mediterranean Sea. J. Appl. Ichthyol. 2009, 25, 98–102. [Google Scholar] [CrossRef]
- Yeldan, H.; Gundogdu, S. Morphometric relationships and growth of common stingray, Dasyatis pastinaca (Linnaeus, 1758) and marbled stingray, Dasyatis marmorata (Staindachner, 1892) in the northeastern Levantine Basin. J. Black Sea Medit. Environ. 2018, 24, 10–27. [Google Scholar]
- Tiralongo, F.; Messina, G.; Cazzolla Gatti, R.; Tibullo, D.; Lombardo, B.M. Some biological aspects of juveniles of the rough ray, Raja radula Delaroche, 1809 in Eastern Sicily (central Mediterranean Sea). J. Sea Res. 2018, 142, 174–179. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Brundo, M.V.; Lombardo, B.M. Biological aspects of the common torpedo, Torpedo torpedo (Linnaeus, 1758) (Elasmobranchii: Torpedinidae), in the central Mediterranean Sea (Sicily, Ionian Sea). Eur. Zool. J. 2019, 86, 488–496. [Google Scholar] [CrossRef]
- Romanelli, M.; Consalvo, I.; Vacchi, M.; Finoia, M.G. Diet of Torpedo torpedo and Torpedo marmorata in a coastal area of Central Western Italy (Mediterranean Sea). Mar. Life 2006, 16, 21–30. [Google Scholar]
- Saglam, H.; Ak, O.; Kutlu, S.; Aydin, I. Diet and feeding strategy of the common stingray Dasyatis pastinaca (Linnaeus, 1758) on the Turkish coast of southeastern Black Sea. Cah. Biol. Mar. 2010, 51, 37–44. [Google Scholar] [CrossRef]
- Santic, M.; Paladin, A.; Agovic, A. Diet of the common stingray, Dasyatis pastinaca (Chondrichthyes: Dasyatidae) in the eastern Adriatic Sea. Cah. Biol. Mar. 2011, 52, 349–356. [Google Scholar] [CrossRef]
- Ponte, D.; Barcelos, L.D.M.; Santos, C.; Medeiros, J.; Barreiros, P. Diet of Dasyatis pastinaca and Myliobatis aquila (Myliobatiformes) from the Azores, NE Atlantic. Cybium 2016, 40, 209–214. [Google Scholar]
- Compagno, L.J.V. Alternate life-history styles of cartilaginous fishes in time and space. Environ. Biol. Fish. 1990, 28, 33–75. [Google Scholar] [CrossRef]
- Motta, P.J. Prey capture behavior and feeding mechanisms of elasmobranchs. In Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: New York, NY, USA, 2004; pp. 165–202. [Google Scholar]
- Bezerra, M.F.; Lacerda, L.D.; Lai, C.-T. Trace metals and persistent organic pollutants contamination in batoids (Chondrichthyes: Batoidea): A systematic review. Environ. Pollut. 2019, 248, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.H.; Fodrie, F.J.; Summerson, H.C.; Powers, S.P. Site-specific and density-dependent extinction of prey by schooling rays: Generation of a population sink in top quality habitat for bay scallops. Oecologia 2001, 129, 349–356. [Google Scholar] [CrossRef]
- Thrush, S.F.; Pridmore, J.E.; Hewitt, J.E.; Cummings, V.J. The importance of predators on a sandflat: Interplay between seasonal changes in prey density and predator effects. Mar. Ecol. Prog. Ser. 1994, 107, 211–222. [Google Scholar] [CrossRef]
- Yemisken, E.; Forero, M.G.; Megalofonou, P.; Eryilmaz, L.; Navarro, J. Feeding habits of three Batoids in the Levantine Sea (north-eastern Mediterranean Sea) based on stomach content and isotopic data. J. Mar. Biol. Assoc. UK 2017, 98, 89–96. [Google Scholar] [CrossRef]
- Paterson, B.; Kainge, P. Rebuilding the Namibian hake fishery: A case for collaboration between scientists and fishermen. Ecol. Soc. 2014, 19, 49. [Google Scholar] [CrossRef]
- Tiralongo, F.; Lillo, A.O.; Tibullo, D.; Tondo, E.; Lo Martire, C.; D’Agnese, R.; Macali, A.; Mancini, E.; Giovos, I.; Coco, S.; et al. Monitoring uncommon and non-indigenous fishes in Italian waters: One year of results for the AlienFish project. Reg. Stud. Mar. Sci. 2018. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SEX | N | Total Length (TL) cm | Disc Width (DW) cm | Weight (W) g | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Range | Mean | SD | Range | Mean | SD | Range | Mean | SD | ||
| M | 57 | 31.2–62.8 | 44.62 | 10.72 | 15.9–38.2 | 24.36 | 6.94 | 125–2195 | 672.1 | 616.51 |
| F | 63 | 31–76.4 | 44.99 | 12.55 | 16.8–45.5 | 24.72 | 8.09 | 144–3781 | 750.1 | 952.76 |
| Combined | 120 | 31–76.4 | 44.82 | 11.67 | 15.9–45.5 | 24.55 | 7.53 | 125–3781 | 713 | 808.29 |
| SEX | N | a | C.I. a | b | C.I. b | R2 | p-Value |
|---|---|---|---|---|---|---|---|
| M | 57 | 0.0133 | 0.0095–0.0185 | 3.303 | 3.197–3.409 | 0.99 | <0.01 |
| F | 63 | 0.0163 | 0.0126–0.0217 | 3.229 | 3.147–3.311 | 0.99 | <0.01 |
| Combined | 120 | 0.0149 | 0.0121–0.0183 | 3.262 | 3.196–3.327 | 0.99 | <0.01 |
| Food Items | %F | %N | %W | IRI | %IRI |
|---|---|---|---|---|---|
| CRUSTACEA | |||||
| Alpheus sp. | 5.97 | 2.06 | 1.37 | 20.47 | 1.1 |
| Amphipoda | 7.46 | 11.86 | 2.07 | 103.91 | 5.6 |
| Caridea | 14.93 | 6.7 | 12.32 | 283.96 | 15.2 |
| Crangon crangon | 8.96 | 3.61 | 3.15 | 60.56 | 3.2 |
| Liocarcinus sp. | 10.45 | 3.61 | 20.2 | 248.81 | 13.3 |
| Mysida | 10.45 | 43.81 | 3.04 | 489.58 | 26.2 |
| Portunus hastatus | 8.96 | 3.09 | 17.44 | 183.94 | 9.8 |
| Xantho sp. | 4.48 | 1.55 | 8.74 | 46.09 | 2.5 |
| MOLLUSCA | |||||
| Donacidae | 7.46 | 3.09 | 2.32 | 40.35 | 2.2 |
| Gastropoda | 5.97 | 2.06 | 6.79 | 52.83 | 2.8 |
| POLYCHAETA | |||||
| Lumbrineridae | 8.96 | 11.34 | 15.5 | 240.48 | 12.9 |
| Polychaeta | 10.45 | 5.15 | 2.27 | 77.53 | 4.1 |
| TELEOSTEI | |||||
| Pomatoschistus sp. | 1.49 | 0.52 | 3.21 | 5.55 | 0.3 |
| Digested | 4.48 | 1.55 | 1.58 | 14.02 | 0.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiralongo, F.; Messina, G.; Lombardo, B.M. Biological Aspects of Juveniles of the Common Stingray, Dasyatis pastinaca (Linnaeus, 1758) (Elasmobranchii, Dasyatidae), from the Central Mediterranean Sea. J. Mar. Sci. Eng. 2020, 8, 269. https://doi.org/10.3390/jmse8040269
Tiralongo F, Messina G, Lombardo BM. Biological Aspects of Juveniles of the Common Stingray, Dasyatis pastinaca (Linnaeus, 1758) (Elasmobranchii, Dasyatidae), from the Central Mediterranean Sea. Journal of Marine Science and Engineering. 2020; 8(4):269. https://doi.org/10.3390/jmse8040269
Chicago/Turabian StyleTiralongo, Francesco, Giuseppina Messina, and Bianca Maria Lombardo. 2020. "Biological Aspects of Juveniles of the Common Stingray, Dasyatis pastinaca (Linnaeus, 1758) (Elasmobranchii, Dasyatidae), from the Central Mediterranean Sea" Journal of Marine Science and Engineering 8, no. 4: 269. https://doi.org/10.3390/jmse8040269
APA StyleTiralongo, F., Messina, G., & Lombardo, B. M. (2020). Biological Aspects of Juveniles of the Common Stingray, Dasyatis pastinaca (Linnaeus, 1758) (Elasmobranchii, Dasyatidae), from the Central Mediterranean Sea. Journal of Marine Science and Engineering, 8(4), 269. https://doi.org/10.3390/jmse8040269