Varying Biological Activity and Wind Stress Affect the DMS Response during the SAGE Iron Enrichment Experiment

 , ,
, , 
Abstract
1. Introduction
2. Methods
2.1. Site Selection
2.2. The Preparation of the Iron and Dual Tracer Solutions
2.3. Wind Speed and SF6 Mapping
2.4. Phytoplankton Pigments
2.5. Microzooplankton Grazing
2.6. Underway DMS Measurements
2.7. Surface Seawater DMSP Concentrations
3. Results
3.1. SF6 Mapping and Dissolved Iron
3.2. Total Chlorophyll a and Photosynthetic Efficiency (Fv/Fm)
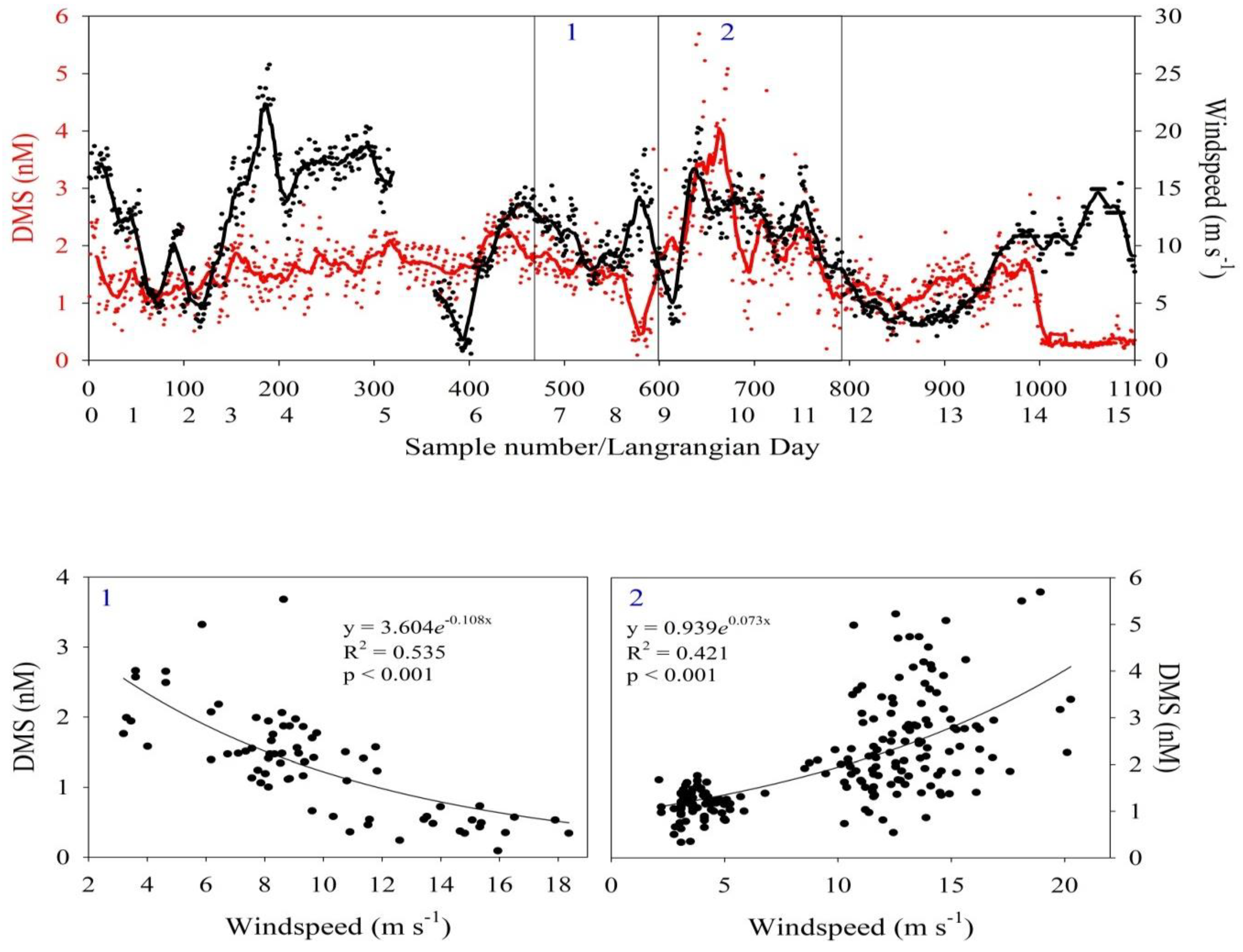
3.3. Continuous Underway Dissolved DMS Concentrations
3.4. Wind Speed and Correlation with Seawater DMS Concentrations
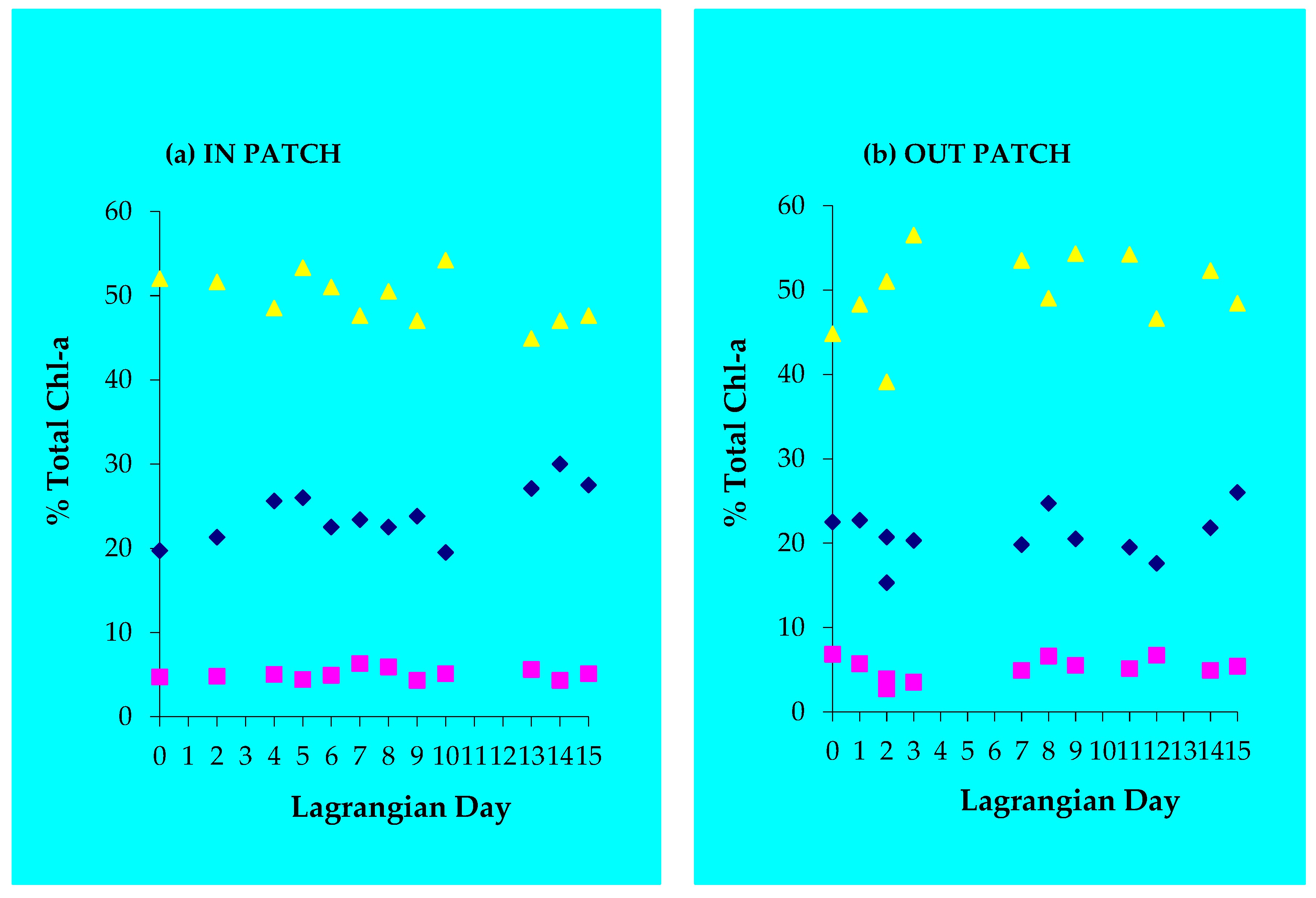
3.5. Variation of the Depth-Integrated DMSP and Chl a Fractions
3.6. Phytoplankton Pigments
3.7. DMSP Pigment Correlations
3.8. Dominant Phytoplankton Species, Growth Rates, and Microzooplankton Grazing
4. Discussion
4.1. Chlorophyll a and Phytoplankton Pigments
4.2. Microzooplankton Grazing
4.3. Effect of Increasing and Decreasing Wind Speed on Dissolved DMS
4.4. Comparison of DMS (P) with Other Iron Enrichment Experiments
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Martin, J.H. Glacial-interglacial CO2 change: The iron hypothesis. Paleoceanography 1990, 5, 1–13. [Google Scholar] [CrossRef]
- Boyd, P.W.; Watson, A.J.; Law, C.S.; Abraham, E.R.; Trull, T.; Murdoch, R.; Bakker, D.C.E.; Bowie, A.R.; Buesseler, K.O.; Chang, H.; et al. A mesoscale phytoplankton bloom in the polar Southern Ocean stimulated by iron fertilization. Nature 2000, 407, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef]
- Watson, A.J. Iron Limitation in the Oceans. In The Biogeochemistry of Iron in Seawater; Turner, D.R., Hunter, K.A., Eds.; John Wiley and Sons Ltd.: Chichester, UK, 2001; Volume 7, pp. 9–39. [Google Scholar]
- Charlson, R.J.; Lovelock, J.E.; Andreae, M.O.; Warren, S.G. Ocean phytoplankton, atmospheric sulphur, cloud albedo and climate. Nature 1987, 326, 655–661. [Google Scholar] [CrossRef]
- Jones, G.B.; Curran, M.A.J.; Swan, H.B.; Greene, R.M.; Griffiths, F.B.; Clemenston, L.A. Influence of different water masses and biological activity on dimethylsulphide and dimethylsulphoniopropionate in the subantarctic zone of the Southern Ocean during ACE 1. J. Geophys. Res. 1998, 103, 16691–16701. [Google Scholar] [CrossRef]
- Harvey, M.J.; Law, C.S.; Smith, M.J.; Hall, J.A.; Abraham, E.R.; Stevens, C.L.; Hadfield, M.G.; Ho, D.T.; Ward, B.; Archer, S.D.; et al. The SOLAS air-sea gas exchange experiment (SAGE) 2004. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 753–763. [Google Scholar] [CrossRef][Green Version]
- Coale, K.H.; Johnson, K.S.; Fitzwater, S.E.; Gordon, R.M.; Tanner, S.; Chavez, F.P.; Ferioli, L.; Sakamoto, C.; Rogers, P.; Millero, F.J.; et al. A massive phytoplankton bloom induced by an ecosystem-scale iron fertilization experiment in the equatorial Pacific Ocean. Nature 1996, 383, 495–501. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Bale, A.J.; Kolber, Z.S.; Aiken, J.; Falkowski, P.G. Confirmation of iron limitation of phytoplankton photosynthesis in the equatorial Pacific Ocean. Nature 1996, 383, 508–511. [Google Scholar] [CrossRef]
- Turner, S.M.; Nightingale, P.D.; Spokes, L.J.; Liddicoat, M.I.; Liss, P.S. Increased dimethyl sulfide concentrations in sea water from in situ iron enrichment. Nature 1996, 383, 513–517. [Google Scholar] [CrossRef]
- Turner, S.M.; Harvey, M.J.; Law, C.S.; Nightingale, P.D.; Liss, P.S. Iron-induced changes in oceanic sulfur biogeochemistry. Geophys. Res. Lett. 2004, 31, L14307. [Google Scholar] [CrossRef]
- Boyd, P.W.; Jickells, T.; Law, C.S.; Blain, S.; Boyle, E.; Buesseler, K.O.; Coale, K.H.; Cullen, J.; de Baar, H.; Follows, M.; et al. A synthesis of mesoscale iron-enrichment experiments 1993–2005: Key findings and implications for ocean biogeochemistry. Science 2007, 315, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Boyd, P.; Laroche, J.; Gall, M.; Frew, R.; McKay, R.M.L. Role of iron, light and silicate in controlling algal biomass in subantarctic waters southeast of New Zealand. J. Geophys. Res. 1999, 104, 13395–13408. [Google Scholar] [CrossRef]
- McKay, R.M.L.; Wilhelm, W.; Hall, J.; Hutchins, D.A.; Al-Rshaidat, M.M.D.; Mioni, C.E.; Pickmere, S.; Porta, D.; Boyd, P.W. Impact of phytoplankton on the biogeochemical cycling of iron in subantarctic waters southeast of New Zealand during FeCycle. Glob. Biogeochem. Cycles 2005, 19, GB4S24. [Google Scholar] [CrossRef]
- Bradford-Grieve, J.M.; Boyd, P.W.; Chang, F.H.; Chiswell, S.; Hadfield, M.; Hall, J.A.; James, M.R.; Nodder, S.D.; Shuskina, E.A. Pelagic ecosystem structure and functioning in the Subtropical Front region east of New Zealand in austral winter and spring 1993. J. Plankton Res. 1999, 21, 405–428. [Google Scholar] [CrossRef]
- Chang, F.H.; Gall, M. Phytoplankton assemblages and photosynthetic pigments during winter and spring in the Subtropical Convergence region near New Zealand. N. Z. J. Mar. Freshw. Res. 1998, 32, 515–530. [Google Scholar] [CrossRef]
- Hall, J.A.; James, M.R.; Bradford-Grieves, J.M. Structure and dynamics of the pelagic microbial food web of the Subtropical Convergence region east of New Zealand. Aquat. Microb. Ecol. 1999, 20, 95–105. [Google Scholar] [CrossRef]
- Peloquin, J.; Hall, J.; Safi, K.; Smith, W.O., Jr.; Wright, S.; Van den Enden, R. The response of phytoplankton to iron enrichment in Sub-Antarctic HNLCSi waters: Results from the SAGE experiment. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 808–823. [Google Scholar] [CrossRef]
- Peloquin, J.; Hall, J.; Safi, K.; Ellwood, M.; Law, C.S.; Thompson, K.; Kuparinen, J.; Harvey, M.; Pickmere, S. Control of the phytoplankton response during the SAGE experiment: A synthesis. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 824–838. [Google Scholar] [CrossRef]
- Law, C.S.; Smith, M.J.; Stevens, C.L.; Abraham, E.R.; Ellwood, M.J.; Hill, P.; Nodder, S.; Peloquin, J.; Pickmere, S.; Safi, K.; et al. Did dilution limit the phytoplankton response to iron addition in HNLCLSi Sub-Antarctic waters during the SAGE experiment? Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 786–799. [Google Scholar] [CrossRef]
- Wright, S.W.; Jeffrey, S.W.; Mantoura, R.F.C. Evaluation of methods and solvents for pigment extraction. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO: Paris, France, 1997; pp. 261–282. ISBN 92-3-103275-5. [Google Scholar]
- Zapata, M.; Rodriguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new method using reversed-phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef]
- Archer, S.D.; Safi, K.; Hall, A.; Cummings, D.G.; Harvey, M. Grazing suppression of dimethylsulphoniopropionate (DMSP) accumulation in iron-fertilised, Sub-Antarctic waters. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 839–850. [Google Scholar] [CrossRef]
- Turner, S.M.; Malin, G.; Liss, P.; Harbour, D.S.; Holligan, P.M. The seasonal variation of dimethyl sulfide and dimethylsulfoniopropionate concentrations in nearshore waters. Limnol. Oceanogr. 1988, 33, 364–375. [Google Scholar] [CrossRef]
- Curran, M.A.J.; Jones, G.B.; Burton, H. Spatial distribution of dimethylsulfide and dimethylsulfoniopropionate in the Australasian sector of the Southern Ocean. J. Geophys. Res. 1998, 103, 16677–616689. [Google Scholar] [CrossRef]
- Deschaseaux, E.S.M.; Beltran, V.H.; Jones, G.B.; Deseo, M.A.; Swan, H.B.; Harrison, P.L.; Eyre, B.D. Comparative response of DMS and DMSP concentrations in Symbiodinium clades C1 and D1 under thermal stress. J. Exp. Mar. Biol. Ecol. 2014, 459, 181–189. [Google Scholar] [CrossRef]
- Kiene, R.P.; Slezak, D. Low dissolved DMSP concentrations in seawater revealed by small-volume gravity filtration and dialysis sampling. Limnol. Oceanogr. Methods 2006, 4, 80–95. [Google Scholar] [CrossRef]
- Curran, M.A.J.; Jones, G.B. Dimethylsulfide in the Southern Ocean: Seasonality and flux. J. Geophys. Res. 2000, 105, 20451–20459. [Google Scholar] [CrossRef]
- Sutton, P.J.H. The Southland Current: A Sub Antarctic current. N. Z. J. Mar. Freshw. Res. 2003, 37, 645–652. [Google Scholar] [CrossRef]
- Zapata, M.; Jeffrey, S.W.; Wright, S.W.; Rodríguez, F.; Clementson, L.; Garrido, J.L. Photosynthetic pigments in 37 species (65 Strains) of Haptophyta: Implications for phylogeny and oceanography. Mar. Ecol. Prog. Ser. 2004, 270, 83–102. [Google Scholar] [CrossRef]
- Keller, M.D.; Bellows, W.K.; Guillard, R.R.L. Dimethyl sulfide production in marine phytoplankton. In Biogenic Sulfur in the Environment; Saltzman, E.S., Cooper, W.J., Eds.; American Chemical Society: New Orleans, LA, USA, 1989; pp. 167–182. [Google Scholar]
- Riseman, S.F.; DiTullio, G.R. Particulate dimethylsulfoniopropionate and dimethylsulfoxide in relation to iron availability and algal community structure in the Peru Upwelling System. Can. J. Fish. Aquat. Sci. 2004, 61, 721–735. [Google Scholar] [CrossRef]
- Ledyard, K.; Dacey, J.W.H. Microbial cycling of DMSP and DMS in coastal and oligotrophic seawater. Limnol. Oceanogr. 1996, 41, 33–40. [Google Scholar] [CrossRef]
- Levasseur, M.; Michaud., S.; Egge, J.; Cantin, G.; Nejstgaard., J.C.; Sanders, R.; Fernandez, E.; Solberg, P.T.; Heimdal, B.; Gosselin, M. Production of DMSP and DMS during a mesocosm study of an Emiliania huxleyi bloom: Influence of bacteria and Calanus finmarchicus grazing. Mar. Biol. 1996, 126, 609–618. [Google Scholar] [CrossRef]
- Zubkov, M.V.; Fuchs, B.M.; Archer, S.D.; Kiene, R.P.; Amann, R.; Burkill, P.H. Linking the composition of bacterioplankton to rapid turnover of dissolved dimethylsulfoniopropionate in an algal bloom in the North Sea. Environ. Microbiol. 2001, 3, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Burkill, P.H.; Archer, S.D.; Robinson, C.; Nightingale, P.D.; Groom, S.B.; Tarran, G.A.; Zubkov, M.V. Dimethyl sulfide biogeochemistry within a coccolithophore bloom (DISCO): An overview. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 2863–2885. [Google Scholar] [CrossRef]
- Brussaard, C.P.D. Viral control of phytoplankton populations: A review. J. Eukaryot. Microbiol. 2004, 52, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.; Geider, R.J. Impact of iron limitation on the photosynthetic apparatus of the diatom Chaetoceros meulleri (Bacillariophyceae). J. Phycol. 2001, 37, 987–1000. [Google Scholar] [CrossRef]
- Kuparinen, J.; Hall, J.; Ellwood, M.; Safi, K.; Peloquin, J.; Katz, D. Bacterioplankton responses to iron enrichment during the SAGE experiment. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 800–807. [Google Scholar] [CrossRef]
- Gabric, A.; Murray, N.; Stone, L.; Kohl, M. Modeling the production of dimethylsulfide during a phytoplankton bloom. J. Geophys. Res. 1993, 98, 22805–22816. [Google Scholar] [CrossRef]
- Currie, K.I.; Macaskill, B.; Reid, M.R.; Law, C.S. Processes governing the carbon chemistry during the SAGE experiment. Deep Sea Res. Part II 2011, 58, 851–860. [Google Scholar] [CrossRef]
- Smith, M.J.; Ho, D.T.; Law, C.S.; McGregor, J.; Popinet, S.; Schlosser, P. Uncertainties in gas exchange parameterization during the SAGE dual-tracer experiment. Depp Sea Res. II 2011, 58, 869–881. [Google Scholar] [CrossRef]
- Martin, J.H.; Coale, K.H.; Johnson, K.S.; Fitzwater, S.E.; Gordon, R.M.; Tanner, S.J.; Hunter, C.N.; Elrod, V.A.; Nowicki, J.L.; Coley, T.L.; et al. Testing the iron hypothesis in ecosystems of the equatorial Pacific Ocean. Nature 1994, 371, 123–129. [Google Scholar] [CrossRef]
- Boyd, P.W.; Abraham, E.R. Iron-mediated changes in phytoplankton photosynthetic competence during SOIREE. Deep Sea Res. Part II 2001, 48, 2529–2550. [Google Scholar] [CrossRef]
- Bonner-Knowles, D.; Jones, G.; Gabric, A. Dimethylsulphide production in the Southern Ocean using a nitrogen-based flow network model and field measurements from ACE-1. J. Atmos. Ocean Sci. 2005, 10, 95–122. [Google Scholar] [CrossRef]
- Scheffer, M.; Boer, R.J.D. Implications of spatial heterogeneity for the paradox of enrichment. Ecology 1995, 76, 2270–2277. [Google Scholar] [CrossRef]
- Quėguiner, B. Iron fertilization and the structure of planktonic communities in high nutrient regions of the Southern Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2012, 90, 43–54. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infusion Release | Date/Time | Time (days) | Tracer | DFe | |
|---|---|---|---|---|---|
| Area (km2) | (nmol L−1) | ||||
| 1. | 25 March 2004 (15:00–23:30) | 0 | SF6 | 33 | 3.03 |
| 2. | 31 March 2004 (00:00–06:00) | 5.2–5.45 | SF6 | 43 | 1.59 |
| 3. | 03 April 2004 (12:30–18:30) | 8.75–9.0 | SF6 | 72.5 | 0.55 |
| 4. | 06 April 2004 (22:20–03:30) | 12.14–12.35 | SF6 | 88 | 1.01 |
| Day/Time | SF6 (fmol/L) | In/Out Patch | Range DMS (nM) | Mean DMS (nM) |
|---|---|---|---|---|
| 8 (17:00–21:50) | nd | Out Patch | 0.09–0.73 | 0.46 (20) |
| 8.75–9 * | nd | In Patch | - | - |
| 9–10 (10:55–12:30) | 4–34 | In Patch | 1.12–5.7 | 2.69 (72) |
| 10 (12:45–18:10) | nd | Out Patch | 0.54–1.91 | 1.27 (12) |
| 10–11 (18:45–02:40) | 5–17 | In Patch | 0.81–4.7 | 2.21 (25) |
| 11 (03:50–04:30) | 0.7–3 | Out Patch | 0.97–2.03 | 1.70 (4) |
| 11 (04:45–10:35) | 4–16 | In Patch | 1.42–3.59 | 2.25 (18) |
| 11 (11:20–22:40) | 1–8 | Out Patch | 0.82–2.94 | 1.34 (19) |
| 11–12 (22:50–03:35) | 5–15 | In Patch | 0.2–1.53 | 1.29 (7) |
| 12 (01:15–02:55) | 0.3–0.4 | Out Patch | 0.56–1.29 | 0.89 (7) |
| 12 (01:15–02:55) | 3.2–9.3 | In Patch | 1.01–1.53 | 1.29 (18) |
| 12 (04:20–04:45) | 0.5–2.5 | Out Patch | 0.88–1.23 | 1.00 (3) |
| 12 (04:20–04:45) | 5–13 | In Patch | 0.8–2.15 | 1.29 (18) |
| 12 (05:55–10:25) | nd | Out Patch | 0.35–1.76 | 1.07 (44) |
| 12–13 ** (22:20–06:20) | nd | In Patch | 0.33–2.23 | 1.45(35) |
| 13 (06:35–08:40) | nd | Out Patch | 0.85–1.68 | 1.26 (10) |
| 13 (08:55–14:34) | 43–1720 | In Patch | 1.1–1.84 | 1.57 (9) |
| 13 (15:08–19:06) | 0.43–4.7 | Out Patch | 0.77–1.69 | 1.28(7) |
| 14 (06:30–11:15) | 13–21 | In Patch | 0.26–0.45 | 0.30 (19) |
| Correlation | Total Chl a | Fucoxanthin | 19′-Hexanoylloxy- Fucoxanthin | Peridin | β, β-Carotene |
|---|---|---|---|---|---|
| DMSPt | 0.64 * | 0.77 ** | 0.67 * | 0.74 ** | 0.77 ** |
| DMSPp | 0.03 ns | 0.06 ns | 0.002 ns | 0.12 ns | 0.08 ns |
| Experiment: | Start: | Interim | Final |
|---|---|---|---|
| IRONEX I | 2.7 (1) | 2.8 (3) | - |
| IRONEX II | 2.5 (−1) | 4.1 (6) | 4.2 (17) |
| SOIREE | 0.5 (1) | 0.9 (9) | 3.4 (13) |
| EisenEx | 1.9 (−1) | 3.1 (12) | 1.6 (19) |
| SAGE | 1.15 (1) | 2.7 (10–11) | 0.3 (16) |
| Experiment | Start | Interim | Final |
|---|---|---|---|
| (a) DMSPp (nM) (day of expt.) | |||
| IRONEX I | 30 (D1) | 56 (D3) | - |
| IRONEX II | 14 (D−1) | 36 (D6) | 16 (D17) |
| SOIREE | 27 (D1) | 56 (D9) | 34 (D13) |
| EisenEx | 45 (D−1) | 74 (D12) | 47 (D19) |
| SAGE a | 55 (D1) | 15 (D10–11) | 37 (D16) |
| (b) DMSPd (nM) (day of expt.) | |||
| SAGE b,c | 17 (D1) | 48 (D10–11) | 36 (D16) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, G.; Harvey, M.; King, S.; Schneider, A.; Wright, S.; Fortescue, D.; Swan, H.; Maher, D.T. Varying Biological Activity and Wind Stress Affect the DMS Response during the SAGE Iron Enrichment Experiment. J. Mar. Sci. Eng. 2020, 8, 268. https://doi.org/10.3390/jmse8040268
Jones G, Harvey M, King S, Schneider A, Wright S, Fortescue D, Swan H, Maher DT. Varying Biological Activity and Wind Stress Affect the DMS Response during the SAGE Iron Enrichment Experiment. Journal of Marine Science and Engineering. 2020; 8(4):268. https://doi.org/10.3390/jmse8040268
Chicago/Turabian StyleJones, Graham, Mike Harvey, Stacey King, Anke Schneider, Simon Wright, Darren Fortescue, Hilton Swan, and Damien T. Maher. 2020. "Varying Biological Activity and Wind Stress Affect the DMS Response during the SAGE Iron Enrichment Experiment" Journal of Marine Science and Engineering 8, no. 4: 268. https://doi.org/10.3390/jmse8040268
APA StyleJones, G., Harvey, M., King, S., Schneider, A., Wright, S., Fortescue, D., Swan, H., & Maher, D. T. (2020). Varying Biological Activity and Wind Stress Affect the DMS Response during the SAGE Iron Enrichment Experiment. Journal of Marine Science and Engineering, 8(4), 268. https://doi.org/10.3390/jmse8040268