An Invasive Mussel (Arcuatula senhousia, Benson 1842) Interacts with Resident Biota in Controlling Benthic Ecosystem Functioning

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
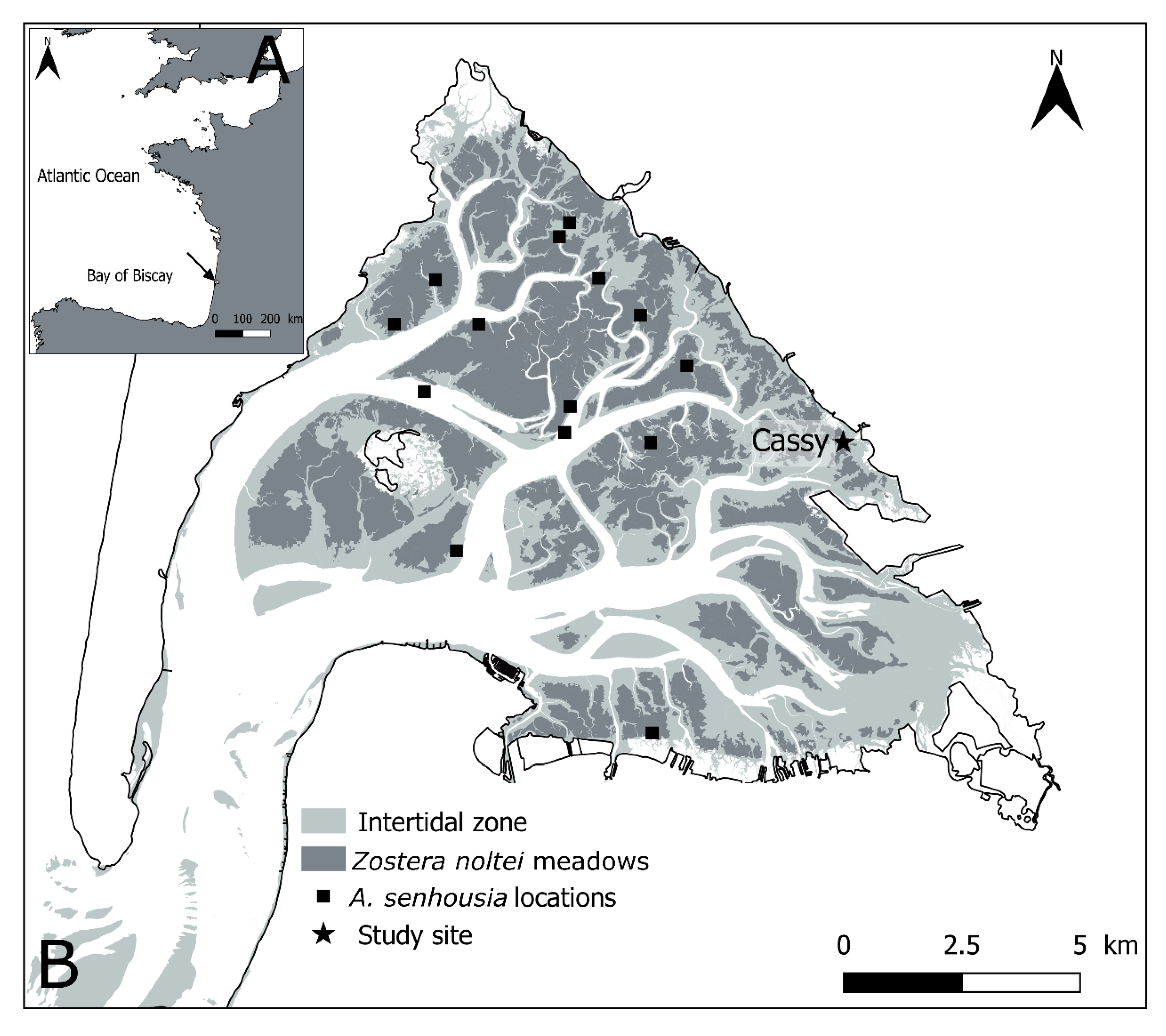
2.1. Study Site
2.2. Field Sampling and Experimental Procedures
2.2.1. General Strategy
2.2.2. Animal and Sediment Cores Collection
2.2.3. Experimental Setup
2.2.4. Experimental Treatments
2.2.5. Bioturbation Measurements
2.2.6. Nutrient Flux Measurements
2.2.7. Macrofauna and Seagrass Characteristics
2.3. Statistical Analyses
3. Results
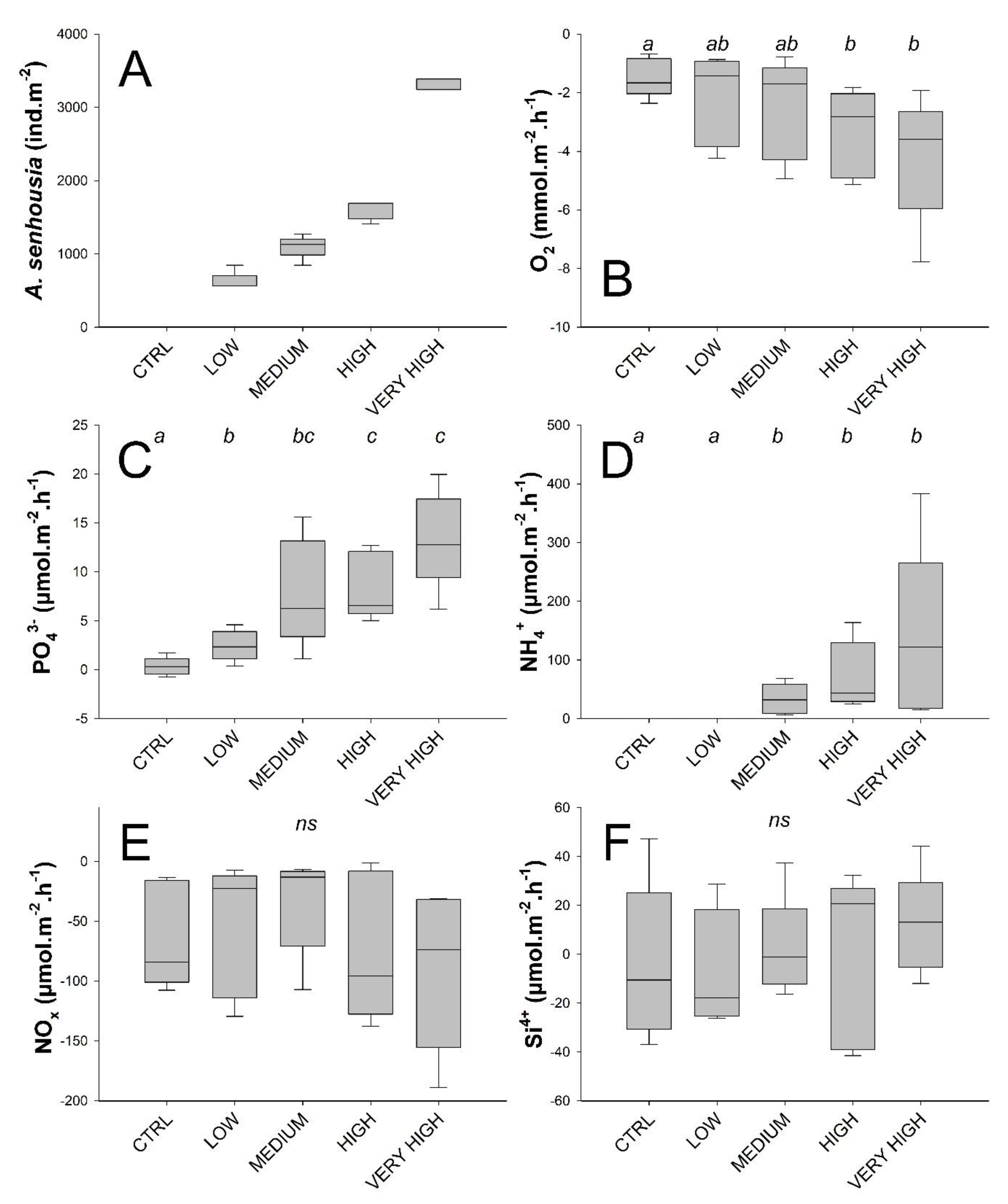
3.1. Environmental Variables
3.2. Solute Fluxes
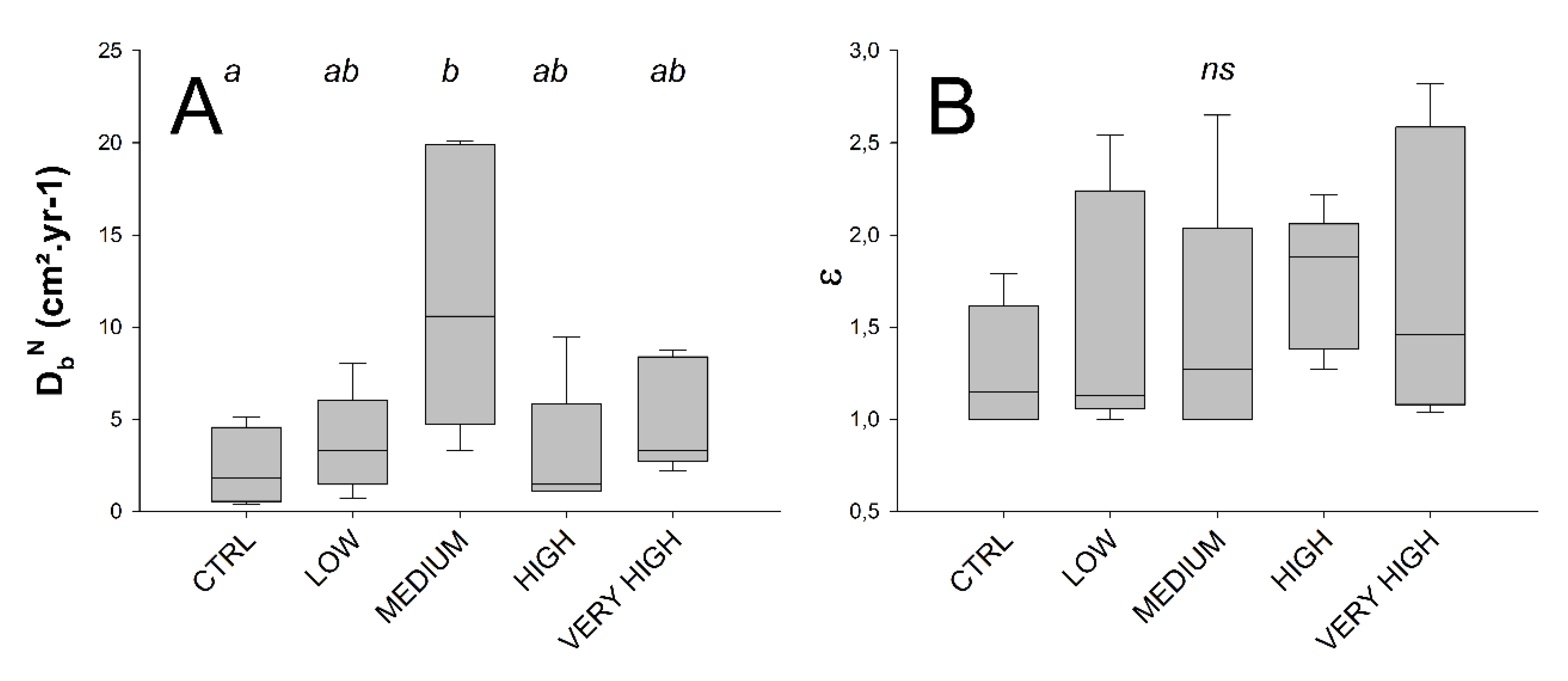
3.3. Bioturbation
3.4. Contextualization of A. senhousia Effects
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hulme, P.H. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Williams, S.L.; Grosholz, E.D. The Invasive Species Challenge in Estuarine and Coastal Environments: Marrying Management and Science. Estuaries Coasts 2008, 31, 3–20. [Google Scholar] [CrossRef]
- Chauvaud, L.; Jean, J.; Ragueneau, O.; Thouzeau, G. Long-term variation of the Bay of Brest ecosystem: Benthic–pelagic coupling revisited. Mar. Ecol. Prog. Ser. 2000, 200, 35–48. [Google Scholar] [CrossRef]
- Kauppi, L.; Norkko, J.; Ikonen, J.; Norkko, A. Seasonal variability in ecosystem functions: Quantifying the contribution of invasive species to nutrient cycling in coastal ecosystems. Mar. Ecol. Prog. Ser. 2017, 572, 193–207. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Crooks, J. Lag times and exotic species: The ecology and management of biological invasions in slow-motion. Ecoscience 2005, 12, 316–329. [Google Scholar] [CrossRef]
- Bachelet, G.; Blanchet, H.; Cottet, M.; Dang, C.; de Montaudouin, X.; de Moura Queiros, A.; Gouillieux, B.; Lavesque, N. A round-the-world tour almost completed: First records of the invasive mussel Musculista senhousia in the North-east Atlantic (southern Bay of Biscay). Mar. Biodivers. Rec. 2009, 2, 1–4. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Bacher, S.; Blackburn, T.M.; Dick, J.T.A.; Essl, F.; Evans, T.; Gaertner, M.; Hulme, E.; Kühn, I.; Mrugała, A.; et al. Defining the impact of non-native species. Conserv. Biol. 2014, 28, 1188–1194. [Google Scholar] [CrossRef]
- Giakoumi, S.; Katsanevakis, S.; Albano, P.G.; Azzurro, E.; Cardoso, A.C.; Cebrian, E.; Deidun, A.; Edelist, D.; Francour, P.; Jimenez, C.; et al. Management priorities for marine invasive species. Sci. Total Environ. 2019, 688, 976–982. [Google Scholar] [CrossRef]
- Bouma, T.J.; Ortells, V.; Ysebaert, T. Comparing biodiversity effects among ecosystem engineers of contrasting strength: Macrofauna diversity in Zostera noltii and Spartina anglica vegetations. Helgol. Mar. Res. 2009, 63, 3–18. [Google Scholar] [CrossRef]
- Gestoso, I.; Arenas, F.; Rubal, M.; Veiga, P.; Peña, P.; Olabarria, C. Shifts from native to non-indigenous mussels: Enhanced habitat complexity and its effects on faunal assemblages. Mar. Environ. Res. 2013, 90, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, J.E.; Norkko, J.; Kauppi, L.; Villnäs, A.; Norkko, A. Species and functional trait turnover in response to broad-scale change and an invasive species. Ecosphere 2016, 7, e01289. [Google Scholar] [CrossRef]
- De Montaudouin, X.; Blanchet, H.; Hippert, B. Relationship between the invasive slipper limpet Crepidula fornicata and benthic megafauna structure and diversity, in Arcachon bay. J. Mar. Biol. Assoc. UK 2018, 98, 2017–2028. [Google Scholar] [CrossRef]
- Galimany, E.; Freeman, C.J.; Lunt, J.; Domingos, A.; Sacks, P.; Walters, L. Feeding competition between the native oyster Crassostrea virginica and the invasive mussel Mytella charruana. Mar. Ecol. Prog. Ser. 2017, 564, 57–66. [Google Scholar] [CrossRef]
- Grosholz, E.D.; Ruiz, G.M.; Dean, C.A.; Shirley, K.A.; Maron, J.L.; Connors, P.G. The impacts of a nonindigenous marine predator in a Californian bay. Ecology 2000, 81, 1206–1224. [Google Scholar] [CrossRef]
- Guy-Haim, T.; Lyons, D.A.; Kotta, J.; Ojaveer, H.; Queirós, A.M.; Chatzinikolaou, E.; Arvanitidis, C.; Como, S.; Magni, P.; Blight, A.J.; et al. Diverse effects of invasive ecosystem engineers on marine biodiversity and ecosystem functions: A global review and meta-analysis. Glob. Chang. Biol. 2018, 24, 906–924. [Google Scholar] [CrossRef]
- Reise, K.; Buschbaum, C.; Büttger, H.; Wegner, M. Invading oysters and native mussels: From hostile takeover to compatible bedfellows. Ecosphere 2017, 8, e01949. [Google Scholar] [CrossRef]
- Kauppi, L.; Bernard, G.; Bastrop, R.; Norkko, A.; Norkko, J. Increasing densities of an invasive polychaete enhance bioturbation with variable effects on solute fluxes. Sci. Rep. 2018, 8, 7619. [Google Scholar] [CrossRef]
- Crespo, D.; Solan, M.; Leston, S.; Pardal, M.A.; Dolbeth, M. Ecological consequences of invasion across the freshwater-water transition in a warming world. Ecol. Evol. 2018, 8, 1807–1817. [Google Scholar] [CrossRef]
- Como, S.; Pais, A.; Rumolo, P.; Saba, S.; Sprovieri, M.; Magni, P. Effects of an invasive mussel, Arcuatula senhousia, on local benthic consumers: A laboratory 13C-labeling study. Mar. Biol. 2016, 163, 140. [Google Scholar] [CrossRef]
- Bartoli, M.; Nizzoli, D.; Viaroli, P.; Turolla, E.; Castaldelli, G.; Fano, E.A.; Rossi, R. Impact of Tapes philippinarum farming on nutrient dynamics and benthic respiration in the Sacca di Goro. Hydrobiologia 2001, 455, 203–212. [Google Scholar] [CrossRef]
- Kim, S.L.; Lee, H.G.; Kang, S.M.; Yu, O.H. The influence of Manilla Clam (Tapes philippinarum) on Macrobenthos Communities in a Korean Tidal Ecosystem. Sustainability 2020, 12, 4205. [Google Scholar] [CrossRef]
- Anton, A.; Geraldi, N.R.; Lovelock, C.E.; Apostolaki, E.T.; Bennet, S.; Cebrian, J.; Krause-Jensen, D.; Marbà, N.; Martinetto, P.; Pandolfi, J.M.; et al. Global ecological impacts of marine exotic species. Nat. Ecol. Evol. 2019, 3, 787–800. [Google Scholar] [CrossRef]
- Queiros, A.D.; Hiddink, J.G.; Johnson, G.; Cabral, H.N.; Kaiser, M.J. Context dependence of marine ecosystem engineer invasion impacts on benthic ecosystem functioning. Biol. Invas. 2011, 13, 1059–1075. [Google Scholar] [CrossRef]
- Thomsen, M.; Wernberg, T.; Olden, J.D.; Griffin, J.N.; Silliman, B.R. A framework to study the context-dependent impacts of marine invasions. J. Exp. Mar. Biol. Ecol. 2011, 400, 322–327. [Google Scholar] [CrossRef]
- Sorte, C.J.B.; Williams, S.L.; Zerebecki, R.A. Ocean warming increases threat of invasive species in a marine fouling community. Ecology 2010, 91, 2198–2204. [Google Scholar] [CrossRef]
- Geraldi, N.R.; Anton, A.; Santana-Garcon, J.; Bennett, S.; Marbà, N.; Lovelock, C.E.; Apostolaki, E.T.; Cebrian, J.; Krause-Jensen, D.; Martinetto, P.; et al. Ecological effects of non-native species in marine ecosystems relate to co-occurring anthropogenic pressures. Glob. Chang. Biol. 2019, 26, 1248–1258. [Google Scholar] [CrossRef]
- Crooks, J.A. The population ecology of an exotic mussel Musculista senhousia, in a southern California bay. Estuaries 1996, 19, 42–45. [Google Scholar] [CrossRef]
- Creese, R.; Hooker, S.; DeLuca, S.; Wharton, W. Ecology and environmental impact of Musculista senhousia (Mollusca: Bivalvia: Mytilidae) in Tamaki estuary, Auckland, New Zealand. N. Z. J. Mar. Freshw. Res. 1997, 31, 225–236. [Google Scholar] [CrossRef]
- Mistri, M. Ecological characteristics of the invasive Asian date mussel, Musculista senhousia, in the Sacca di Goro (Adriatic Sea, Italy). Estuaries 2002, 25, 431–440. [Google Scholar] [CrossRef]
- Barfield, P.; Holmes, A.; Watson, G.; Rowe, G. First evidence of Arcuatula senhousia (Benson, 1842), the Asian date mussel in UK waters. J. Conchol. 2018, 43, 43–217. [Google Scholar]
- Faasse, M. A record of the Asian mussel Arcuatula senhousia (Benson in Cantor, 1842) from NW Europe (the Netherlands). Spirula 2018, 416, 14–15. [Google Scholar]
- Lourenço, P.; Henriques, M.; Catry, I.; Granadeiro, J.; Catry, T. First record of the invasive Asian date mussel Arcuatula senhousia (Benson, 1842) (Mollusca: Bivalvia: Mytilidae) in West Africa. J. Nat. Hist. 2018, 52, 2567–2571. [Google Scholar] [CrossRef]
- Kovalev, E.; Zhivoglyadova, L.; Revkov, N.; Frolenko, L.; Afanasyev, D. First record of the bivalve Arcuatula senhousia (Benson, 1842) in the Russian part of the Azov-Black Sea basin. Russ. J. Biol. Invasions 2017, 8, 316–320. [Google Scholar] [CrossRef]
- Magni, P.; Como, S.; Montani, S.; Tsutsumi, H. Interlinked temporal changes in environmental conditions, chemical characteristics of sediments and macrofaunal assemblages in an estuarine intertidal sandflat (Seto Inland Sea, Japan). Mar. Biol. 2006, 149, 1185–1197. [Google Scholar] [CrossRef]
- Yamamuro, M.; Hiratsuka, J.; Ishitobi, Y. Seasonal change in a filter-feeding bivalve Musculista senhousia population of a eutrophic estuarine lagoon. J. Mar. Syst. 2000, 26, 117–126. [Google Scholar] [CrossRef]
- Takenaka, R.; Komorita, T.; Tsutsumi, H. Accumulation of organic matter within a muddy carpet created by the Asian date mussel, Arcuatula senhousia, on the Midori River tidal flats, Japan. Plankton Benthos Res. 2018, 13, 1–9. [Google Scholar] [CrossRef][Green Version]
- Mistri, M.; Munari, C. The invasive bag mussel Arcuatula senhousia is a CO2 generator in near-shore coastal ecosystems. J. Exp. Mar. Biol. Ecol. 2013, 440, 164–168. [Google Scholar] [CrossRef]
- Kellogg, M.L.; Cornwell, J.C.; Owens, M.S.; Paynter, K.T. Denitrification and nutrient assimilation on a restored oyster reef. Mar. Ecol. Prog. Ser. 2013, 480, 1–19. [Google Scholar] [CrossRef]
- Smyth, A.R.; Piehler, M.F.; Grabowski, J.H. Habitat context influences nitrogen removal by restored oyster reefs. J. Appl. Ecol. 2015, 52, 716–725. [Google Scholar] [CrossRef]
- Smyth, A.R.; Geraldi, N.R.; Thompson, S.P.; Piehler, M.F. Biological activity exceeds biogenic structure in influencing sediment nitrogen cycling in experimental oyster reefs. Mar. Ecol. Prog. Ser. 2016, 560, 173–183. [Google Scholar] [CrossRef]
- Crooks, J.A. Habitat alteration and community-level effects of an exotic mussel, Musculista senhousia. Mar. Ecol. Prog. Ser. 1998, 162, 137–152. [Google Scholar] [CrossRef]
- Yamamuro, M.; Hiratsuka, J.; Ishitobi, Y. What prevents Musculista senhousia from constructing byssal thread mats in estuarine environments? A case study focusing on Lake Shinji and nearby estuarine waters. Landsc. Ecol. Eng. 2010, 6, 23–28. [Google Scholar] [CrossRef]
- Cohen, A.N. The Exotics Guide: Non-native Marine Species of the North American Pacific Coast. Center for Research on Aquatic Bioinvasions, Richmond, CA, and San Francisco Estuary Institute: Oakland, CA, USA Revised September 2011. Available online: http://www.exoticsguide.org (accessed on 15 October 2020).
- Crooks, J.A. Predators of the invasive mussel Musculista senhousia (Mollusca; Mytilidae). Pac. Sci. 2002, 56, 49–56. [Google Scholar] [CrossRef]
- Taylor, D.I.; Wood, S.A.; McNabb, P.; Ogilvie, S.; Cornelisen, C.; Walker, J.; Khor, S.; Craig Cary, S. Favcilitation effects of invasive and farmed bivalves on native populations of the sea slug Pleurobranchaea maculata. Mar. Ecol. Prog. Ser. 2015, 537, 39–48. [Google Scholar] [CrossRef]
- Marshall, D.J. Predatory and reproductive responses of the estuarine whelk Thais gradata (Caenogastropoda: Muricidae) to novel colonization by Musculista senhousia (Bivalvia: Mytilidae). J. Mar. Biol. Assoc. UK 2009, 87, 1387–1393. [Google Scholar] [CrossRef]
- Magni, P.; Montani, S.; Takada, C.; Tsutsumi, H. Temporal scaling and relevance of bivalve nutrient excretion on a tidal flat of the Seto Island Sea, Japan. Mar. Ecol. Prog. Ser. 2000, 198, 139–155. [Google Scholar] [CrossRef]
- Mistri, M.; Modugno, S.; Rossi, R. Sediment Organic Matter and its Nutritional Quality: A Short-Term Experiment with Two Exotic Bivalve Species. Chem. Ecol. 2003, 19, 225–231. [Google Scholar] [CrossRef]
- Reusch, T.B.H.; Williams, S.L. Variable responses of native eelgrass Zost, era marina to a non-indigenous bivalve Musculista senhousia. Oecologia 1998, 113, 428–441. [Google Scholar] [CrossRef]
- Crooks, J.A.; Khim, H.S. Architectural vs. biological effects of a habitat-altering, exotic mussel, Musculista senhousia. J. Exp. Mar. Biol. Ecol. 1999, 240, 53–75. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Staehr, P.A.; Nejrup, L.B.; Schiel, D.R. Effects of the invasive macroalga Gracilaria vermiculophylla on two co-occurring foundation species and associated invertebrates. Aquat. Invasions 2013, 8, 133–145. [Google Scholar] [CrossRef]
- Como, S.; Floris, A.; Pais, A.; Rumolo, P.; Saba, S.; Sprovieri, M.; Magni, P. Assessing the impact of the Asian mussel Arcuatula senhousia in the recently invaded Oristano Lagoon-Gulf system (W Sardinia, Italy). Estuar. Coast. Shelf Sci. 2018, 201, 123–131. [Google Scholar] [CrossRef]
- Bongiorni, L.; Fiorentino, F.; Auriemma, R.; Aubry, F.B.; Camatti, E.; Camin, F.; Nasi, F.; Pansera, M.; Ziller, L.; Grall, J. Food web of a confined and anthropogenically affected coastal basin (the Mar Piccolo of Taranto) revealed by carbon and nitrogen stable isotopes analyses. Environ. Sci. Pollut. Res. 2016, 23, 12725–12738. [Google Scholar] [CrossRef] [PubMed]
- Do, V.T.; Blanchet, H.; de Montaudouin, X.; Lavesque, N. Limited consequences of seagrass decline on benthic macrofauna and associated biotic indicators. Estuaries Coast 2013, 36, 795–807. [Google Scholar] [CrossRef]
- Dubois, S.; Blanchet, H.; Garcia, A.; Masse, M.; Galois, R.; Grémare, A.; Charlier, K.; Guillou, G.; Richard, P.; Savoye, N. Trophic resource use by macrozoobenthic primary consumers within a semi-enclosed coastal ecosystem: Stable isotope and fatty acid assessment. J. Sea Res. 2014, 88, 87–99. [Google Scholar] [CrossRef]
- Bernard, G. (CNRS-University of Bordeaux UMR 5805 EPOC, France). Personal communication, 2020.
- Plus, M.; Dalloyau, S.; Trut, G.; Auby, I.; de Montaudouin, X.; Emery, E.; Noël, C.; Viala, C. Long-term evolution (1988–2008) of Zostera spp. meadows in Arcachon Bay (Bay of Biscay). Estuar. Coast. Shelf Sci. 2010, 87, 357–366. [Google Scholar]
- Auby, I.; Labourg, P.J. Seasonal dynamics of Zostera noltei Hornem. in the Bay of Arcachon (France). J. Sea Res. 1996, 35, 269–277. [Google Scholar] [CrossRef]
- Deborde, J.; Abril, G.; Mouret, A.; Jézéquel, D.; Thouzeau, G.; Clavier, J.; Bachelet, G.; Anschutz, P. Effects of seasonal dynamics of a Zostera noltii meadow on phosphorus and iron cycles in a tidal mudflat (Arcachon Bay, France). Mar. Ecol. Prog. Ser. 2008, 355, 59–71. [Google Scholar] [CrossRef]
- Presley, B.; Claypool, G. Techniques for Analyzing Interstitial Water Samples. Part 1: Determination of Selected Minor and Major Inorganic Constituents. In Initial Reports of the DeepSea Drilling Project, Volume VII; Tracey, J.I., Jr., Sutton, G.H., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1971; pp. 1749–1755. [Google Scholar]
- Andersson, J.H.; Middelburg, J.J.; Soetaert, K. Identifiability and uncertainty analysis of bio-irrigation rates. J. Mar. Res. 2006, 64, 407–429. [Google Scholar] [CrossRef]
- Maire, O.; Duchêne, J.C.; Rosenberg, R.; Braga de Mendonça, J., Jr.; Grémare, A. Effects of food availability on sediment reworking in Abra ovata and A. nitida. Mar. Ecol. Prog. Ser. 2006, 319, 135–153. [Google Scholar] [CrossRef][Green Version]
- Meysman, F.J.R.; Malyuga, V.S.; Boudreau, B.P.; Middelburg, J.J. A generalized stochastic approach to particle dispersal in soils and sediments. Geochim. Cosmochim. Acta 2008, 72, 3460–3478. [Google Scholar] [CrossRef]
- Maire, O.; Duchêne, J.C.; Grémare, A.; Malyuga, V.S.; Meysman, F.J.R. A comparison of sediment reworking rates by the surface deposit-feeding bivalve Abra ovata during summertime and wintertime, with a comparison between two models of sediment reworking. J. Exp. Mar. Biol. Ecol. 2007, 343, 21–36. [Google Scholar] [CrossRef]
- Bernard, G.; Delgard, M.L.; Maire, O.; Ciutat, A.; Lecroart, P.; Deflandre, B.; Duchêne, J.C.; Grémare, A. Comparative study of sediment particle mixing in a Zostera noltei meadow and a bare sediment mudflat. Mar. Ecol. Prog. Ser. 2014, 514, 71–86. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA + for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Delgard, M.L.; Deflandre, B.; Deborde, J.; Richard, M.; Charbonnier, C.; Anschutz, P. Changes in nutrient biogeochemistry in response to the regression of Zostera Noltii meadows in the Arcachon Bay (France). Aquat. Geochem. 2013, 19, 241–259. [Google Scholar] [CrossRef]
- Delgard, M.L.; Deflandre, B.; Bernard, G.; Richard, M.; Kochoni, E.; Charbonnier, C.; Anschutz, P. Benthic oxygen exchange over a heterogeneous Zostera noltei meadow in a temperate coastal ecosystem. Mar. Ecol. Prog. Ser. 2016, 543, 55–71. [Google Scholar] [CrossRef]
- Snelgrove, P.V.R.; Thrush, S.F.; Wall, D.H.; Norkko, A. Real world biodiversity-ecosystem functioning: A seafloor perspective. Trends Ecol. Evol. 2014, 29, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, D.; Solan, M.; Godbold, J.A. Specific arrangements of species dominance can be more influential than evenness in maintaining ecosystem process and function. Sci. Rep. 2016, 6, 39325. [Google Scholar] [CrossRef]
- Bernard, G.; Gammal, J.; Järnström, M.; Norkko, J.; Norkko, A. Quantifying bioturbation across coastal seascapes: Habitat characteristics modify effects of macrofaunal communities. J. Sea Res. 2019, 152, 101766. [Google Scholar] [CrossRef]
- Mistri, M. Effect of Musculista senhousia mats on clam mortality and growth: Much ado about nothing? Aquaculture 2004, 241, 207–218. [Google Scholar] [CrossRef]
- Wrede, A.; Andresen, H.; Asmus, R.; Wiltshire, K.H.; Brey, T. Macrofaunal irrigation traits enhance predictability of nutrient fluxes across the sediment-water interface. Mar. Ecol. Prog. Ser. 2019, 632, 27–42. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Rosenberg, R.; François-Carcaillet, F.; Norling, K.; Mauclaire, L. Influence of bioturbation by three benthic infaunal species on microbial communities and biogeochemical processes in marine sediment. Aquat. Microb. Ecol. 2004, 36, 271–284. [Google Scholar] [CrossRef]
- Lavoie, M.F.; McKindsey, C.W.; Pearce, C.M.; Archambault, P. Influence of intertidal Manila clam Venerupis philippinarum aquaculture on biogeochemical fluxes. Aquacult. Environ. Interact. 2016, 8, 117–130. [Google Scholar] [CrossRef]
- Welsh, D.T.; Nizzoli, D.; Fano, E.A.; Viaroli, P. Direct contribution of clams (Ruditapes philippinarum) to benthic fluxes, nitrification, denitrification and nitrous oxide emission in a farmed sediment. Estuar. Coast. Shelf Sci. 2015, 154, 84–93. [Google Scholar] [CrossRef]
- Magni, P.; Como, S.; Montani, S.; Tsutsumi, H. Interlinked seasonal variation in biogenic nutrient fluxes and pore-water nutrient concentrations in intertidal sediments. Mar. Biol. 2014, 161, 2767–2779. [Google Scholar] [CrossRef]
- Sundbäck, K.; Granéli, W. Influence of microphytobenthos on the nutrient flux between sediment and water: A laboratory analysis. Mar. Ecol. Prog. Ser. 1988, 43, 63–69. [Google Scholar] [CrossRef]
- Douglas, E.J.; Haggitt, T.R.; Rees, T.A.V. Supply- and demand-driven phosphate uptake and tissue phosphorus in temperate seaweeds. Aquat. Biol. 2014, 23, 49–60. [Google Scholar] [CrossRef]
- Lee, K.S.; Park, S.R.; Kim, Y.K. Effects of irradiance, temperature, and nutrients on growth dynamics of seagrasses: A review. J. Exp. Mar. Biol. Ecol. 2007, 350, 144–175. [Google Scholar] [CrossRef]
- Bartoli, M.; Cattadori, M.; Giordani, G.; Viaroli, P. Benthic oxygen respiration, ammonium and phosuphorus regeneration in surficial sediments of the Sacca di Goro (Northern Italy) and two French coastal lagoons: A comparative study. In Coastal Lagoon Eutrophication and ANaerobic Processes (C.L.E.AN.); Developments in, Hydrobiology; Caumette, P., Castel, J., Herbert, R., Eds.; Springer: Dordrecht, The Netherlands, 1996; Volume 117, pp. 143–159. [Google Scholar] [CrossRef]
- Deborde, J. Processus Biogéochimiques des Zones Intertidales des Systems Lagunaires: Le Bassin D’Arcachon (SW, France). Ph.D. Thesis, Université Bordeaux 1, Talence, France, 6 December 2007. [Google Scholar]
- Rigaud, S.; Deflandre, B.; Maire, O.; Bernard, G.; Duchêne, J.C.; Poirier, D.; Anschutz, P. Transient biogeochemistry in intertidal sediments: New insights from tidal pools in Zostera noltei meadows of Arcachon Bay (France). Mar. Chem. 2018, 200, 1–13. [Google Scholar] [CrossRef]
- Delgard, M.L.; Deflandre, B.; Kochoni, E.; Avaro, J.; Cesbron, F.; Bichon, S.; Poirier, D.; Anschutz, P. Biogeochemistry of dissolved inorganic carbon, nitrogen and phosphorus in seagrass (Zostera noltei) sediments at high and low biomass. Estuar. Coast. Shelf Sci. 2016, 179, 12–22. [Google Scholar] [CrossRef]
- Abreu, M.H.; Pereira, R.; Sousa-Pinto, I.; Yarish, C. Eco-physiological studies of the non-indigenous species Gracilaria vermiculophylla (Rhodophyta) and its abundance patterns in Ria de Aveiro lagoon, Portugal. Eur. J. Phycol. 2011, 46, 453–464. [Google Scholar] [CrossRef]
- Welsh, D.T.; Bartoli, M.; Nizzoli, D.; Castaldelli, G.; Riou, S.A.; Viaroli, P. Denitrification, nitrogen fixation, community primary productivity and inorganic-N and oxygen fluxes in an intertidal Zostera noltii meadow. Mar. Ecol. Prog. Ser. 2000, 208, 65–77. [Google Scholar] [CrossRef]
- Gonzalez, D.J.; Smyth, A.R.; Piehler, M.F.; McGlathery, K.J. Mats of the nonnative macroalga, Gracilaria vermiculophylla, alter net denitrification rates and nutrient fluxes on intertidal muflats. Limnol. Oceanogr. 2013, 58, 2101–2108. [Google Scholar] [CrossRef]
- Cacabelos, E.; Engelen, A.H.; Mejia, A.; Arenas, F. Comparison of the assemblage functioning of estuary systems dominated by the seagrass Nanozostera noltii versus the invasive drift seaweed Gracilaria vermiculophylla. J. Sea Res. 2012, 72, 99–105. [Google Scholar] [CrossRef]
- Hosozawa, T.; Kunii, H.; Nakamura, M.; Ojima, T.; Sugiyama, Y.; Yamaguchi, K. Spatial, temporal and vertical variation of distribution and major habitats in Asian mussel (Arcuatula senhousia) in a brackish river along Sea of Japan. Plankton Benthos Res. 2020, 15, 121–131. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Macrofauna (AB ind.m−2; DW g·m−2) | Environment | Bioturbation | Fluxes (µmol.m−2·h−1) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Arc AB | Arc DW | Filt biv DW | Burr poly DW | Sp DW | Sp AB | As DW | Tb DW | Pu DW | C/Nsurf | Zn DW above | Zn DW below | DbN (cm²·yr−1) | ε | PO43− | NH4+ | NOx | Si | O2 (mmol m−2 h−1) | ||
| Control | Mean | 0 | 0.00 | 113.28 | 0.00 | 27.24 | 198 | 0.62 | 0.14 | 16.07 | 8.72 | 1.41 | 7.05 | 2.41 | 1.28 | 0.34 | 0 | −63.50 | −4.36 | −1.48 |
| Min | 0 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0.00 | 0.00 | 4.61 | 8.57 | 0.00 | 0.00 | 0.37 | 1.00 | −0.74 | 0 | −107.53 | −36.92 | −2.36 | |
| Max | 0 | 0.00 | 317.73 | 0.00 | 74.97 | 282 | 1.31 | 0.52 | 51.54 | 8.80 | 9.87 | 12.70 | 5.11 | 1.79 | 1.74 | 0 | −13.26 | 47.27 | −0.68 | |
| SD | 0 | 0.00 | 157.02 | 0.00 | 30.29 | 126 | 0.66 | 0.21 | 19.95 | 0.10 | 4.23 | 7.05 | 2.07 | 0.34 | 0.93 | 0 | 44.36 | 32.50 | 0.66 | |
| Low | Mean | 621 | 269.69 | 87.50 | 0.35 | 6.96 | 113 | 1.41 | 0.05 | 22.11 | 8.89 | 2.46 | 21.16 | 3.65 | 1.54 | 2.48 | 0 | −54.89 | −6.37 | −2.19 |
| Min | 564 | 232.66 | 0.00 | 0.00 | 0.00 | 0 | 0.00 | 0.00 | 5.76 | 7.79 | 0.00 | 16.93 | 0.73 | 1.00 | 0.40 | 0 | −129.41 | −26.10 | −4.23 | |
| Max | 846 | 315.22 | 370.25 | 1.75 | 19.37 | 141 | 4.09 | 0.14 | 43.76 | 10.06 | 3.37 | 26.80 | 8.03 | 2.54 | 4.59 | 0 | −7.12 | 28.75 | −0.87 | |
| SD | 126 | 34.09 | 160.72 | 0.78 | 7.78 | 63 | 1.98 | 0.07 | 14.00 | 0.81 | 1.40 | 4.23 | 2.74 | 0.67 | 1.56 | 0 | 55.31 | 23.87 | 1.54 | |
| Medium | Mean | 1100 | 507.56 | 370.28 | 0.17 | 32.25 | 113 | 1.70 | 0.03 | 14.76 | 9.01 | 4.23 | 8.46 | 11.97 | 1.47 | 7.88 | 33.33 | −34.25 | 2.34 | −2.52 |
| Min | 846 | 414.37 | 0.00 | 0.00 | 0.00 | 0 | 0.00 | 0.00 | 7.00 | 8.25 | 0.00 | 0.00 | 3.29 | 1.00 | 1.12 | 6.67 | −107.03 | −16.40 | −4.94 | |
| Max | 1270 | 577.24 | 1425.74 | 0.83 | 78.98 | 282 | 7.25 | 0.05 | 20.73 | 9.92 | 21.16 | 15.52 | 20.08 | 2.65 | 15.62 | 68.75 | −6.54 | 37.43 | −0.77 | |
| SD | 155 | 62.75 | 605.29 | 0.37 | 34.63 | 118 | 3.12 | 0.02 | 5.69 | 0.67 | 8.46 | 5.64 | 7.68 | 0.68 | 5.50 | 25.87 | 42.14 | 20.68 | 1.72 | |
| High | Mean | 1608 | 780.63 | 20.72 | 0.31 | 20.51 | 85 | 1.20 | 0.09 | 16.61 | 8.76 | 4.23 | 4.11 | 3.07 | 1.75 | 8.45 | 71.90 | −73.41 | −0.74 | −3.34 |
| Min | 1411 | 714.61 | 0.00 | 0.00 | 0.00 | 0 | 0.00 | 0.00 | 5.76 | 8.36 | 0.00 | 0.00 | 1.10 | 1.27 | 5.05 | 25.17 | −137.69 | −41.56 | −5.14 | |
| Max | 1693 | 868.69 | 103.60 | 1.54 | 69.38 | 282 | 4.06 | 0.34 | 25.34 | 9.39 | 18.34 | 29.63 | 9.49 | 2.22 | 12.68 | 163.81 | −1.15 | 32.25 | −1.82 | |
| SD | 126 | 70.99 | 46.33 | 0.69 | 30.87 | 126 | 1.70 | 0.15 | 8.31 | 0.39 | 8.46 | 11.29 | 3.62 | 0.37 | 3.39 | 58.01 | 61.62 | 35.36 | 1.49 | |
| Very high | Mean | 3329 | 1621.89 | 53.12 | 0.87 | 61.14 | 141 | 0.59 | 0.00 | 17.29 | 8.74 | 5.64 | 14.11 | 5.11 | 1.76 | 13.29 | 137.50 | −89.65 | 12.21 | −4.16 |
| Min | 3245 | 1561.22 | 0.00 | 0.00 | 0.00 | 0 | 0.00 | 0.00 | 2.30 | 7.87 | 0.00 | 0.00 | 2.19 | 1.04 | 6.21 | 15.38 | −188.84 | −11.97 | −7.77 | |
| Max | 3386 | 1654.97 | 265.60 | 2.99 | 227.97 | 423 | 1.64 | 0.01 | 58.82 | 10.10 | 19.75 | 31.04 | 8.76 | 2.82 | 19.97 | 383.40 | −30.91 | 44.24 | −1.93 | |
| SD | 77 | 38.99 | 118.78 | 1.32 | 95.91 | 173 | 0.64 | 0.01 | 23.48 | 0.84 | 8.46 | 11.29 | 3.04 | 0.79 | 4.95 | 149.72 | 66.71 | 20.82 | 2.18 | |
| Response | Predictor | AIC | SS (Trace) | Pseudo-F | P | Prop. | Cumul. | res.df |
|---|---|---|---|---|---|---|---|---|
| PO43− | A. senhousia DW | 69.42 | 466.18 | 31.33 | 0.00 | 0.58 | 0.58 | 23 |
| S. plana DW | 62.93 | 98.53 | 8.89 | 0.01 | 0.12 | 0.70 | 22 | |
| A. segmentum DW | 58.96 | 51.78 | 5.67 | 0.04 | 0.06 | 0.76 | 21 | |
| NH4+ | A. senhousia DW | 213.90 | 64,463.00 | 13.68 | 0.00 | 0.37 | 0.37 | 23 |
| Z. noltei aboveground DW | 211.48 | 15,579 | 3.69 | 0.08 | 0.09 | 0.46 | 22 | |
| S. plana DW | 210.95 | 8927 | 2.24 | 0.15 | 0.05 | 0.51 | 21 | |
| Burrowing polychaete DW | 210.63 | 7420 | 1.94 | 0.18 | 0.04 | 0.55 | 20 | |
| Nox | Z. noltei aboveground DW | 196.5 | 13,540 | 5.64 | 0.01 | 0.20 | 0.20 | 23 |
| A. segmentum DW | 196.1 | 5038 | 2.21 | 0.17 | 0.07 | 0.27 | 22 | |
| Si | S. plana DW | 156.7 | 4763 | 9.74 | 0.01 | 0.30 | 0.30 | 23 |
| Filtering bivalve DW | 156.6 | 895 | 1.90 | 0.17 | 0.05 | 0.35 | 22 | |
| A. Segmentum DW | 154.9 | 1421 | 3.34 | 0.07 | 0.09 | 0.44 | 21 | |
| A. senhousia DW | 152.4 | 1494 | 4.02 | 0.06 | 0.09 | 0.53 | 20 | |
| O2 | Z. noltei aboveground DW | 19.6 | 25 | 12.61 | 0.01 | 0.35 | 0.35 | 23 |
| A. senhousia DW | 13.4 | 13 | 8.60 | 0.01 | 0.18 | 0.53 | 22 | |
| S. plana DW | 10.5 | 6 | 4.59 | 0.04 | 0.08 | 0.61 | 21 | |
| ε | Burrowing polychaete DW | −36 | 2.9 | 13.04 | 0.00 | 0.36 | 0.36 | 23 |
| Z. noltei belowground DW | −37 | 0.7 | 3.43 | 0.07 | 0.09 | 0.45 | 22 | |
| DbN | S. plana DW | 83.11 | 84.04 | 3.27 | 0.08 | 0.12 | 0.12 | 23 |
| A. segmentum DW | 83.05 | 46.94 | 1.90 | 0.17 | 0.07 | 0.19 | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernard, G.; Kauppi, L.; Lavesque, N.; Ciutat, A.; Grémare, A.; Massé, C.; Maire, O. An Invasive Mussel (Arcuatula senhousia, Benson 1842) Interacts with Resident Biota in Controlling Benthic Ecosystem Functioning. J. Mar. Sci. Eng. 2020, 8, 963. https://doi.org/10.3390/jmse8120963
Bernard G, Kauppi L, Lavesque N, Ciutat A, Grémare A, Massé C, Maire O. An Invasive Mussel (Arcuatula senhousia, Benson 1842) Interacts with Resident Biota in Controlling Benthic Ecosystem Functioning. Journal of Marine Science and Engineering. 2020; 8(12):963. https://doi.org/10.3390/jmse8120963
Chicago/Turabian StyleBernard, Guillaume, Laura Kauppi, Nicolas Lavesque, Aurélie Ciutat, Antoine Grémare, Cécile Massé, and Olivier Maire. 2020. "An Invasive Mussel (Arcuatula senhousia, Benson 1842) Interacts with Resident Biota in Controlling Benthic Ecosystem Functioning" Journal of Marine Science and Engineering 8, no. 12: 963. https://doi.org/10.3390/jmse8120963
APA StyleBernard, G., Kauppi, L., Lavesque, N., Ciutat, A., Grémare, A., Massé, C., & Maire, O. (2020). An Invasive Mussel (Arcuatula senhousia, Benson 1842) Interacts with Resident Biota in Controlling Benthic Ecosystem Functioning. Journal of Marine Science and Engineering, 8(12), 963. https://doi.org/10.3390/jmse8120963