The Northward Habitat Expansion of the Korean Top Shell Turbo sazae (Gastropoda: Vetigastropoda: Turbinidae) in the Korean Peninsula: Effects of Increasing Water Temperature



 ,
,
Abstract
1. Introduction
2. Materials and Methods
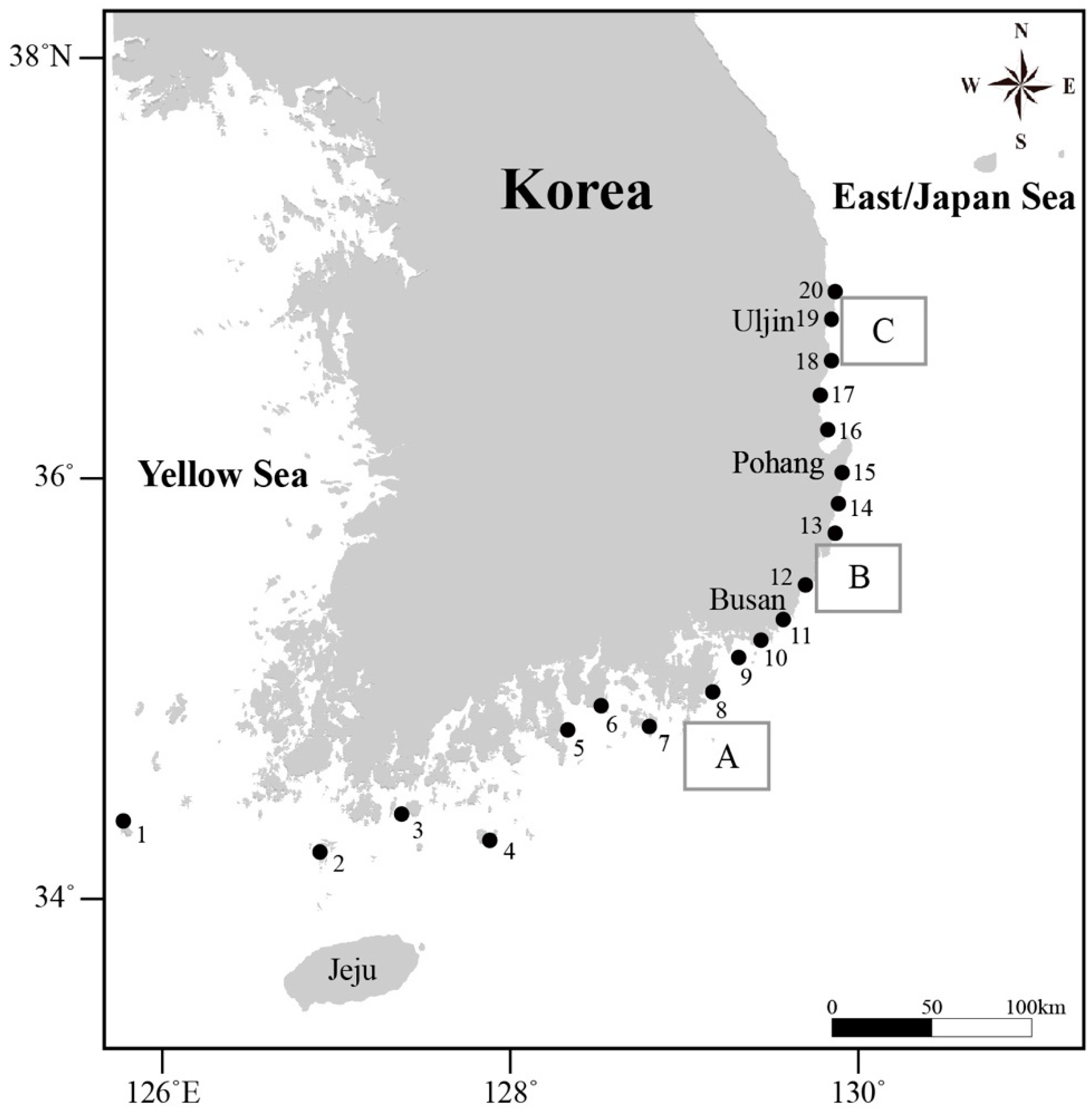
2.1. Data Collection
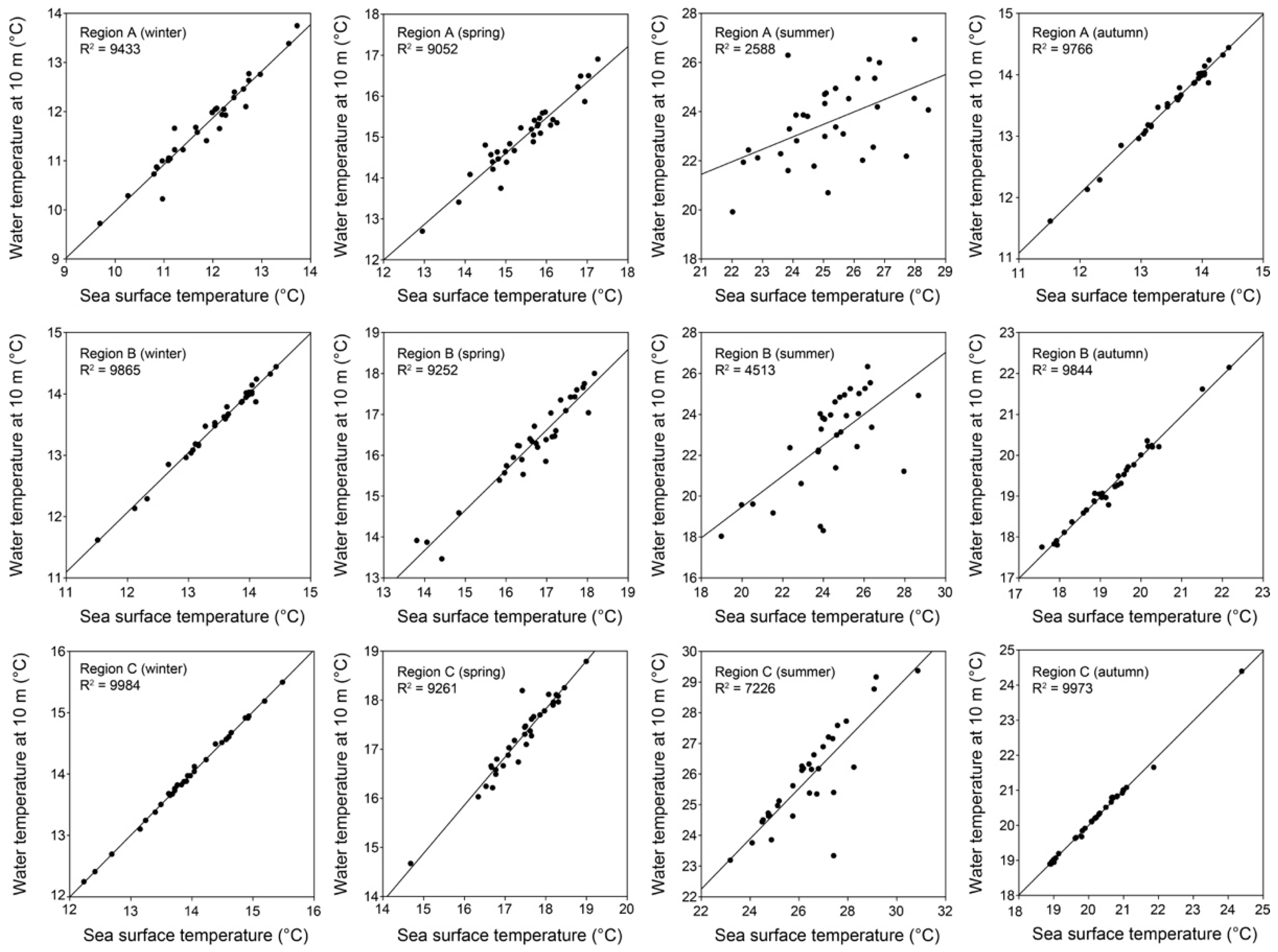
2.2. Seawater Temperature
3. Results
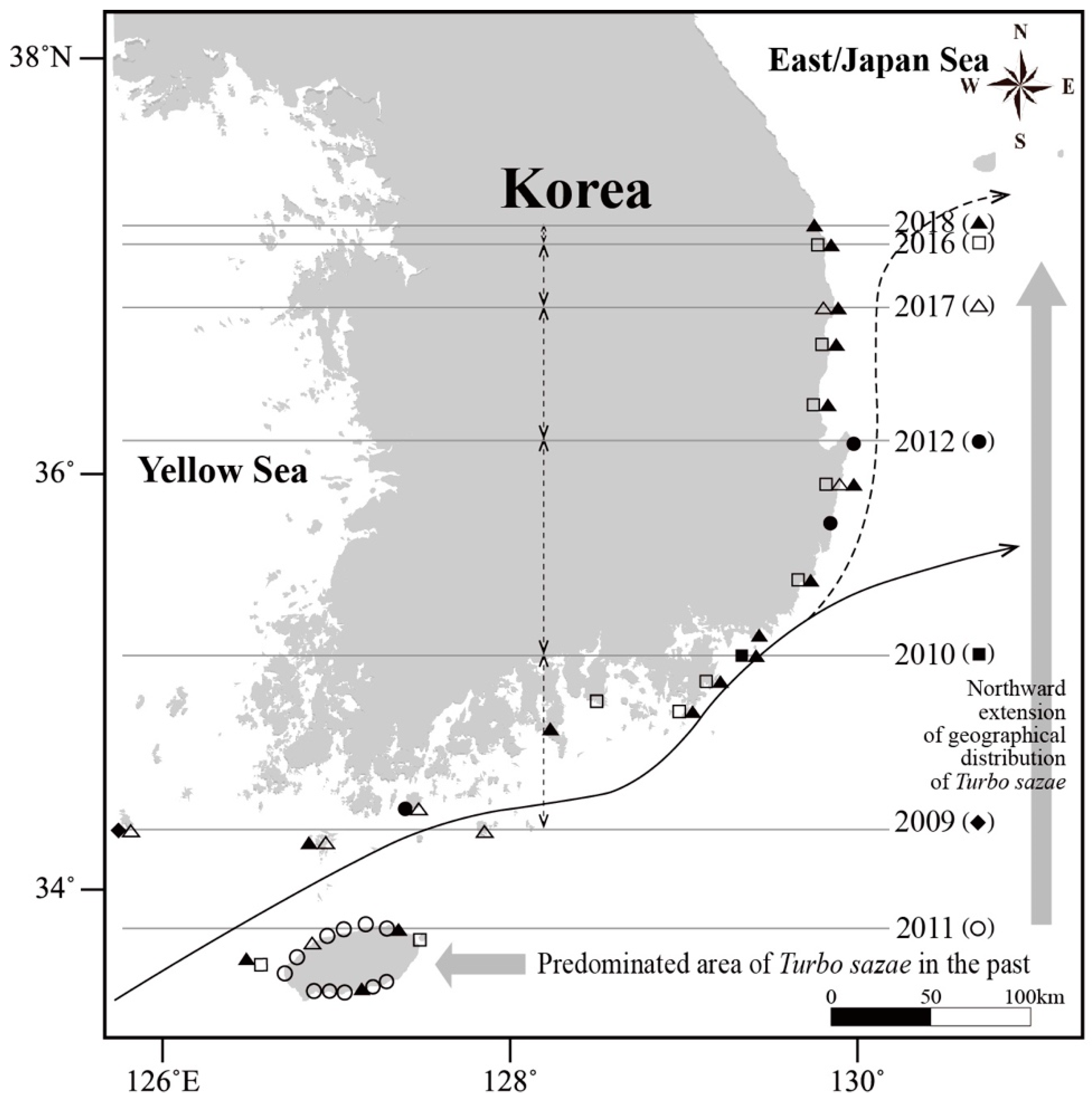
3.1. Occurrence and Geographical Distribution of Turbo Sazae
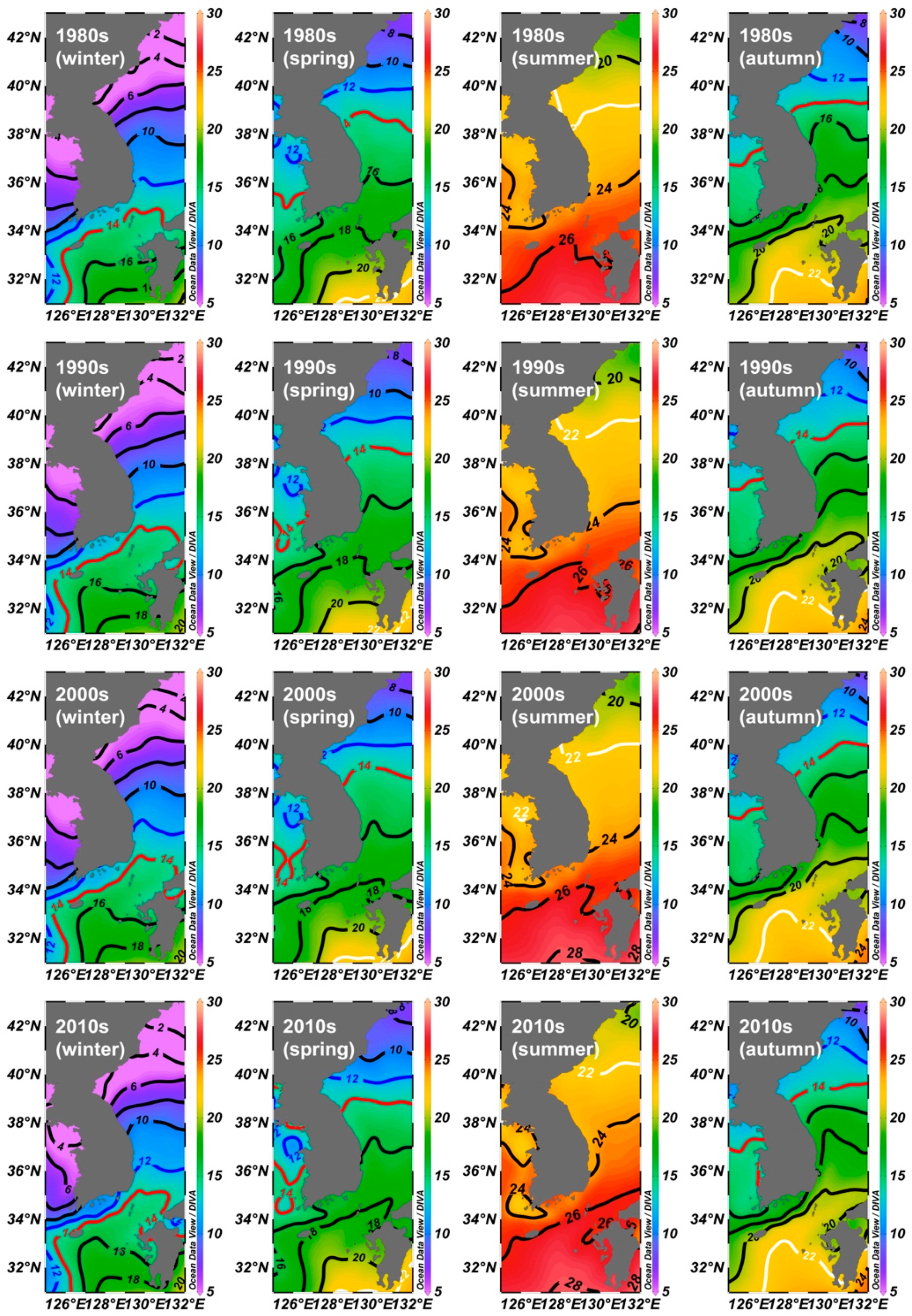
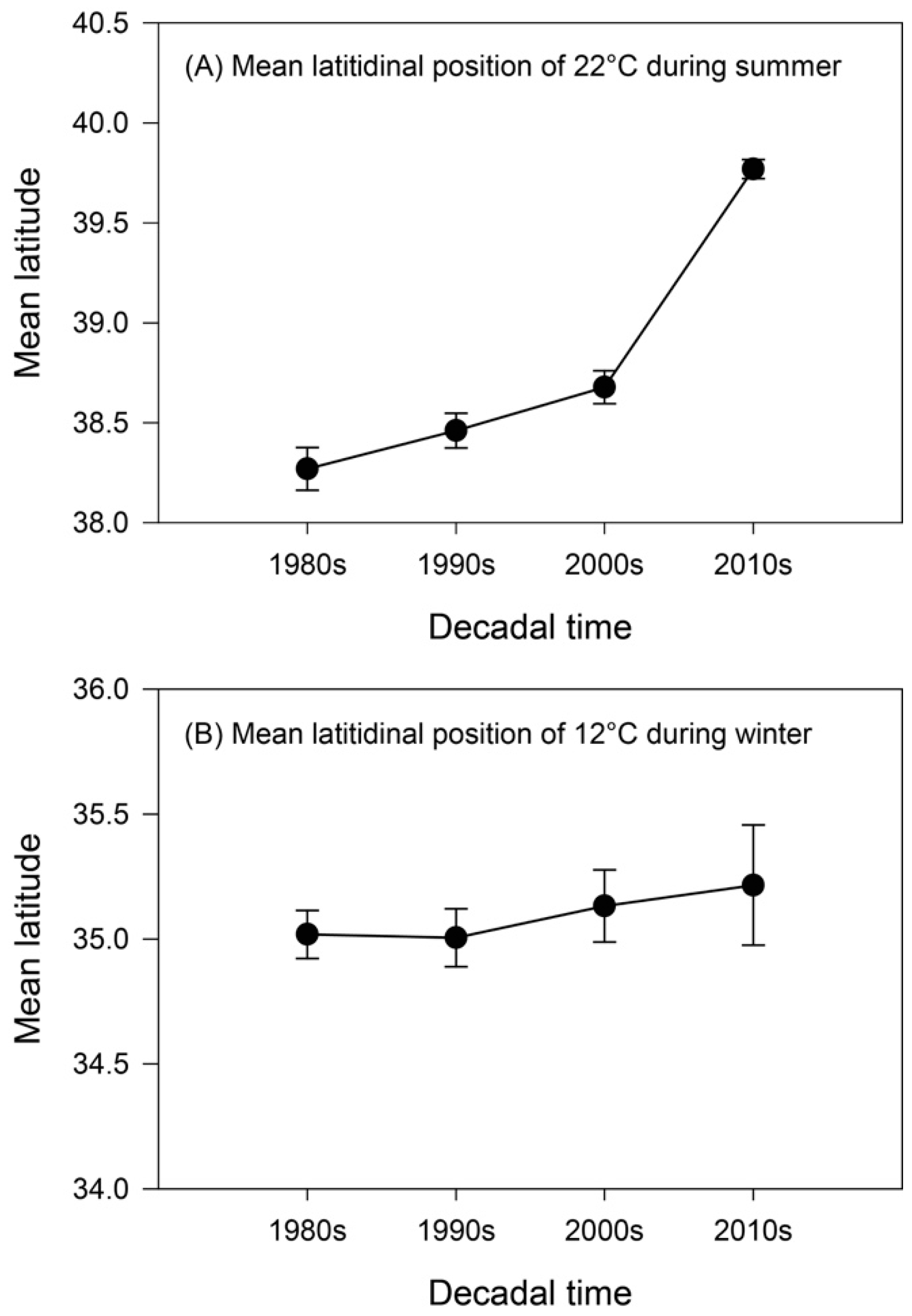
3.2. Variations of Seawater Temperatures
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liverman, D. From uncertain to unequivocal. Environ. Sci. Policy Sustain. Dev. 2007, 49, 28–32. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007; p. 996. [Google Scholar]
- Jung, H.K.; Rahman, S.M.; Kang, C.-K.; Park, S.-Y.; Lee, S.H.; Park, H.J.; Kim, H.-W.; Lee, C.I. The influence of climate regime shifts on the marine environment and ecosystems in the East Asian Marginal Seas and their mechanisms. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 143, 110–120. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Hawkins, S.J.; Southward, A.J.; Burrows, M.T. Modeling the response of populations of competing species to climate change. Ecology 2008, 89, 3138–3149. [Google Scholar] [CrossRef]
- Hawkins, S.; Sugden, H.; Mieszkowska, N.; Moore, P.; Poloczanska, E.; Leaper, R.; Herbert, R.J.; Genner, M.; Moschella, P.; Thompson, R. Consequences of climate-driven biodiversity changes for ecosystem functioning of North European rocky shores. Mar. Ecol. Prog. Ser. 2009, 396, 245–259. [Google Scholar] [CrossRef]
- Noke, R. What is Driving the Range Extension of Gibbula umbilicalis (Gastropda, Trochidae) in the Eastern English Channel? Master’s Thesis, Bournemouth University, Bournemouth, UK, 2016. [Google Scholar]
- Crickenberger, S. Range Limits, Range Shifts, and Lower Thermal Tolerance in the Tropical Barnacle Megabalanus coccopoma. Ph.D. Dissertation, Clemson University, Clemson, SC, USA, 2014. [Google Scholar]
- Hylleberg, J.; Siegismund, H.R. Niche overlap in mud snails (Hydrobiidae): Freezing tolerance. Mar. Biol. 1987, 94, 403–407. [Google Scholar] [CrossRef]
- Gaston, K.J. The Structure and Dynamics of Geographic Ranges; Oxford University Press on Demand: Oxford, UK, 2003; p. 266. [Google Scholar]
- Donn, T.E., Jr. Zonation patterns of Donax serra Röding (Bivalvia: Donacidae) in southern Africa. J. Coast. Res. 1990, 6, 903–911. [Google Scholar]
- Chown, S.L.; Nicolson, S. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: New York, NY, USA, 2004; p. 254. [Google Scholar]
- Southward, A.J.; Langmead, O.; Hardman-Mountford, N.J.; Aiken, J.; Boalch, G.T.; Dando, P.R.; Genner, M.J.; Joint, I.; Kendall, M.A.; Halliday, N.C. Long-term oceanographic and ecological research in the Western English Channel. Adv. Mar. Biol. 2005, 47, 1. [Google Scholar] [CrossRef]
- Helmuth, B.; Mieszkowska, N.; Moore, P.; Hawkins, S.J. Living on the edge of two changing worlds: Forecasting the responses of rocky intertidal ecosystems to climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 373–404. [Google Scholar] [CrossRef]
- Lima, F.P.; Queiroz, N.; Ribeiro, P.A.; Hawkins, S.J.; Santos, A.M. Recent changes in the distribution of a marine gastropod, Patella rustica Linnaeus, 1758, and their relationship to unusual climatic events. J. Biogeogr. 2006, 33, 812–822. [Google Scholar] [CrossRef]
- Herbert, R.; Southward, A.; Sheader, M.; Hawkins, S. Influence of recruitment and temperature on distribution of intertidal barnacles in the English Channel. J. Mar. Biol. Assoc. UK 2007, 87, 487–499. [Google Scholar] [CrossRef]
- Hawkins, S.J.; Moore, P.; Burrows, M.; Poloczanska, E.; Mieszkowska, N.; Herbert, R.; Jenkins, S.; Thompson, R.; Genner, M.; Southward, A. Complex interactions in a rapidly changing world: Responses of rocky shore communities to recent climate change. Clim. Res. 2008, 37, 123–133. [Google Scholar] [CrossRef]
- Mieszkowska, N.; Hawkins, S.; Burrows, M.; Kendall, M. Long-term changes in the geographic distribution and population structures of Osilinus lineatus (Gastropoda: Trochidae) in Britain and Ireland. J. Mar. Biol. Assoc. UK 2007, 87, 537–545. [Google Scholar] [CrossRef]
- Southward, A.; Hawkins, S.; Burrows, M. Seventy years’ observations of changes in distribution and abundance of zooplankton and intertidal organisms in the western English Channel in relation to rising sea temperature. J. Therm. Biol. 1995, 20, 127–155. [Google Scholar] [CrossRef]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Wethey, D.S.; Woodin, S.A. Ecological hindcasting of biogeographic responses to climate change in the European intertidal zone. Hydrobiologia 2008, 606, 139–151. [Google Scholar] [CrossRef]
- Cheung, W.W.; Lam, V.W.; Sarmiento, J.L.; Kearney, K.; Watson, R.; Pauly, D. Projecting global marine biodiversity impacts under climate change scenarios. Fish Fish. 2009, 10, 235–251. [Google Scholar] [CrossRef]
- Cheung, W.W.; Watson, R.; Pauly, D. Signature of ocean warming in global fisheries catch. Nature 2013, 497, 365–368. [Google Scholar] [CrossRef]
- Burrows, M.T.; Schoeman, D.S.; Buckley, L.B.; Moore, P.; Poloczanska, E.S.; Brander, K.M.; Brown, C.; Bruno, J.F.; Duarte, C.M.; Halpern, B.S. The pace of shifting climate in marine and terrestrial ecosystems. Science 2011, 334, 652–655. [Google Scholar] [CrossRef]
- Rubal, M.; Veiga, P.; Moreira, J.; Sousa-Pinto, I. The gastropod Phorcus sauciatus (Koch, 1845) along the north-west Iberian Peninsula: Filling historical gaps. Helgol. Mar. Res. 2014, 68, 169–177. [Google Scholar] [CrossRef]
- Brown, J.; Lomolino, M. Biogeography; Sinauer Associates, Inc.: Sunderland, UK, 1998; p. 624. [Google Scholar]
- Parmesan, C.; Gaines, S.; Gonzalez, L.; Kaufman, D.M.; Kingsolver, J.; Townsend Peterson, A.; Sagarin, R. Empirical perspectives on species borders: From traditional biogeography to global change. Oikos 2005, 108, 58–75. [Google Scholar] [CrossRef]
- Kordas, R.L.; Harley, C.D.; O’Connor, M.I. Community ecology in a warming world: The influence of temperature on interspecific interactions in marine systems. J. Exp. Mar. Biol. Ecol. 2011, 400, 218–226. [Google Scholar] [CrossRef]
- Team, G. GISS Surface Temperature Analysis (GISTEMP); NASA Goddard Institute for Space Studies: New York, NY, USA, 2017.
- Jung, S.; Pang, I.-C.; Lee, J.-H.; Choi, I.; Cha, H.K. Latitudinal shifts in the distribution of exploited fishes in Korean waters during the last 30 years: A consequence of climate change. Rev. Fish Biol. Fish. 2014, 24, 443–462. [Google Scholar] [CrossRef]
- KMA. Report of Global Atmosphere Watch 2018; National Institute of Meteorological Sciences: Seogwipo, Korea, 2019; p. 268.
- Seong, K.-T.; Hwang, J.-D.; Han, I.-S.; Go, W.-J.; Suh, Y.-S.; Lee, J.-Y. Characteristic for long-term trends of temperature in the Korean waters. J. Korean Soc. Mar. Environ. Saf. 2010, 16, 353–360. [Google Scholar]
- Rho, S. Studies on the propagation of top shell-Ⅰ. Spawning and early development of the top shell, Turbo cornutus SOLANDER. Korean J. Fish. Aquat. Sci. 1976, 9, 43–55. [Google Scholar]
- Cha, B.-Y.; Kim, D.-H.; Kim, B.-Y. Growth of Batillus cornutus by capture-recapture method. Korean J. Malacol. 2007, 23, 227–233. [Google Scholar]
- Lee, J.-Y.; Yang, H.-S.; Kang, D.-H.; Jeung, H.-D.; Hong, H.-K.; Lee, H.-J.; Kang, H.-S.; Choi, K.-S. Spatial variation in the annual reproductive cycle of Turbo (Batillus) cornutus (Gastropoda: Trochidae) at Jeju Island, Korea. Invertebr. Reprod. Dev. 2014, 58, 23–33. [Google Scholar] [CrossRef]
- Fukuda, H. Nomenclature of the horned turbans previously known as Turbo cornutus (Lightfoot, 1786) and Turbo chinensis Ozawa & Tomida, 1995 (Vetigastropoda: Trochoidea: Turbinidae) from China, Japan and Korea. Molluscan Res. 2017, 37, 268–281. [Google Scholar] [CrossRef]
- Choe, B. Illustrated Encyclopedia of Fauna and Flora of Korea, Vol. 33, Mollusca (II); National Textbook Company: Seoul, Korea, 1992; p. 860. [Google Scholar]
- Chan, B.K.; Lee, P.-F. Biogeography of intertidal barnacles in different marine ecosystems of Taiwan–potential indicators of climate change. In Global Advances in Biogeography; Stevens, L., Ed.; In Tech: Rijeka, Croatia, 2012; pp. 119–136. [Google Scholar]
- Kojima, S.; Segawa, R.; Hayashi, I. Genetic differentiation among populations of the Japanese turban shell Turbo (Batillus) cornutus corresponding to warm currents. Mar. Ecol. Prog. Ser. 1997, 150, 149–155. [Google Scholar] [CrossRef]
- Hellberg, M.E. Sympatric sea shells along the sea’s shore: The geography of speciation in the marine gastropod Tegula. Evolution 1998, 52, 1311–1324. [Google Scholar] [CrossRef]
- Donlon, C.J.; Martin, M.; Stark, J.; Roberts-Jones, J.; Fiedler, E.; Wimmer, W. The operational sea surface temperature and sea ice analysis (OSTIA) system. Remote Sens. Environ. 2012, 116, 140–158. [Google Scholar] [CrossRef]
- Chang, D.; Chung, S. Shell growth pattern of the top shell, Batillus cornutus in the coastal Jeju Island. J. Korean Soc. Fish. Resour. 2000, 3, 9–15. [Google Scholar]
- Pak, G.; Kim, Y.H.; Park, Y.-G. Lagrangian Approach for a New Separation Index of the East Korea Warm Current. Ocean Sci. J. 2019, 54, 29–38. [Google Scholar] [CrossRef]
- Dytham, C. Choosing and Using Statistics: A Biologist’s Guide; John Wiley & Sons: Chicester, UK, 2011; p. 303. [Google Scholar]
- Lubchenco, J.; Navarette, S.; Tissot, J.; Castilla, C. Possible ecological responses to global climate change: Nearshore benthic biota of northeastern Pacific coastal ecosystems. In Earth System Responses to Global Change; Mooney, H., Fuentes, E., Kronberg, B., Eds.; Academic Press: New York, NY, USA, 1993; pp. 147–166. [Google Scholar]
- Navarette, S.; Lubchenco, J.; Castilla, J. Pacific Ocean Ecosystems and global climate change. In Earth System Responses to Global Change; Mooney, H., Fuentes, E., Kronberg, B., Eds.; Academic Press: New York, NY, USA, 1993; pp. 189–193. [Google Scholar]
- Beaugrand, G.; Reid, P.C.; Ibanez, F.; Lindley, J.A.; Edwards, M. Reorganization of North Atlantic marine copepod biodiversity and climate. Science 2002, 296, 1692–1694. [Google Scholar] [CrossRef]
- Beaugrand, G.; Ibanez, F. Monitoring marine plankton ecosystems. II: Long-term changes in North Sea calanoid copepods in relation to hydro-climatic variability. Mar. Ecol. Prog. Ser. 2004, 284, 35–47. [Google Scholar] [CrossRef]
- Bianchi, C.; Morri, C. Southern species in the Ligurian Sea (northern Mediterranean): New records and a review. Boll. Mus. Ist. Biol. Univ. Genova 1994, 58, 181–197. [Google Scholar]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Zacherl, D.; Gaines, S.D.; Lonhart, S.I. The limits to biogeographical distributions: Insights from the northward range extension of the marine snail, Kelletia kelletii (Forbes, 1852). J. Biogeogr. 2003, 30, 913–924. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef]
- Precht, W.F.; Aronson, R.B. Climate flickers and range shifts of reef corals. Front. Ecol. Environ. 2004, 2, 307–314. [Google Scholar] [CrossRef]
- Stebbing, A.; Turk, S.; Wheeler, A.; Clarke, K. Immigration of southern fish species to south-west England linked to warming of the North Atlantic (1960–2001). J. Mar. Biol. Assoc. UK 2002, 82, 177–180. [Google Scholar] [CrossRef]
- Mieszkowska, N. Changes in the Biogeographic Distribution of the Trochid Gastropods Osilinus lineatus (da Costa) and Gibbula umbilicalis (da Costa) in Response to Global Climate Change: Range Dynamics and Physiological Mechanisms. Ph.D. Dissertation, University of Plymouth, Plymouth, UK, 2005. [Google Scholar]
- Grosholz, E.D. Contrasting rates of spread for introduced species in terrestrial and marine systems. Ecology 1996, 77, 1680–1686. [Google Scholar] [CrossRef]
- Kinlan, B.; Hastings, A. What exotic species tell us about rates of population spread and geographic range expansion. In Species Invasions: Insights to Ecology, Evolution and Biogeography; Sax, D., Stachowicz, J., Gaines, S., Eds.; Sinauer Associates: Sunderland, UK, 2005; pp. 381–419. [Google Scholar]
- Sorte, C.J.; Williams, S.L.; Zerebecki, R.A. Ocean warming increases threat of invasive species in a marine fouling community. Ecology 2010, 91, 2198–2204. [Google Scholar] [CrossRef]
- Kim, S.-J.; Woo, S.-H.; Kim, B.-M.; Hur, S.-D. Trends in sea surface temperature (SST) change near the Korean peninsula for the past 130 years. Ocean Polar Res. 2011, 33, 281–290. [Google Scholar] [CrossRef]
- Na, J.; Seo, J.; Lie, H.-J. Annual and seasonal variations of the sea surface heat fluxes in the East Asian marginal seas. J. Oceanogr. 1999, 55, 257–270. [Google Scholar] [CrossRef]
- Lim, S.; Jang, C.J.; Park, J. Climatology of the mixed layer depth in the East/Japan Sea. J. Mar. Syst. 2012, 96, 1–14. [Google Scholar] [CrossRef]
- Moriyasu, S. The Tsushima Current. In Kuroshio: Its Physical Aspects; Stommel, H.M., Yoshida, K., Eds.; University of Tokyo Press: Tokyo, Japan, 1972; pp. 353–369. [Google Scholar]
- Tian, Y.; Kidokoro, H.; Watanabe, T.; Iguchi, N. The late 1980s regime shift in the ecosystem of Tsushima warm current in the Japan/East Sea: Evidence from historical data and possible mechanisms. Prog. Oceanogr. 2008, 77, 127–145. [Google Scholar] [CrossRef]
- Hwang, J.; Suh, Y.; Ahn, J. Properties of Sea Surface Temperature variations derived from NOAA satellite in Northeastern Asian Waters from 1990 to 2008. Korean J. Nat. Conserv. 2012, 6, 130–136. [Google Scholar]
- Lim, B.-J.; Chang, Y.-S. Development of synthetic regression diagram for analyzing linear trend of sea surface height, temperature, and salinity around the Korean marginal seas. Sea 2016, 21, 67–77. [Google Scholar] [CrossRef][Green Version]
- Kim, J.-J.; Min, H.-S.; Kim, C.-H.; Yoon, J.-H.; Kim, S.-A. Prediction of the Spawning Ground of Todarodes pacificus under IPCC Climate A1B Scenario. Ocean Polar Res. 2012, 34, 253–264. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Jung, H.K.; Lee, C.I. Changes in the spawning ground environment of the common squid, Todarodes pacificus due to climate change. Ocean Polar Res. 2018, 40, 127–143. [Google Scholar] [CrossRef]
- Stauffer, J., Jr. Influence of temperature on fish behavior. In Power Plants: Effects on Fish and Shellfish Behavior; Hocutt, C., Stauffer, J., Jr., Edinger, J., Hall, L., Jr., Morgan, I., Eds.; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Giangrande, A.; Geraci, S.; Belmonte, G. Life-cycle and life-history diversity in marine invertebrates and the implications in community dynamics. Oceanogr. Mar. Biol. Ann. Rev. 1994, 32, 305–333. [Google Scholar]
- Keith, S.A.; Herbert, R.J.; Norton, P.A.; Hawkins, S.J.; Newton, A.C. Individualistic species limitations of climate-induced range expansions generated by meso-scale dispersal barriers. Divers. Distrib. 2011, 17, 275–286. [Google Scholar] [CrossRef]
- Vernberg, W.; Vernberg, F. Metabolic pattern of a trematode and its host: A study in the evolution of physiological responses. In Symbiosis in the Sea; Vernberg, W., Ed.; University of South Carolina Press: Columbia, SC, USA, 1974; pp. 161–172. [Google Scholar]
- Kim, S.-K. Growth of the Juvenile Top Shell, Batillus Cornutus in Food and Water Temperature; Jeju National University: Jeju, Korea, 1999. [Google Scholar]
- Son, M.; Hwang, C.; Kim, J. Beautiful Marine Organisms in the Vicinity of Nuclear Power Plant (Uljin), Korea; Dain Communications: Busan, Korea, 2015; p. 171. [Google Scholar]
- Naylor, E. Biological effects of a heated effluent in docks at Swansea, S. Wales. Proc. Zool. Soc. Lond. 1965, 144, 253–268. [Google Scholar] [CrossRef]
- Hayakawa, J.; Kawamura, T.; Ohashi, S.; Horii, T.; Watanabe, Y. Habitat selection of Japanese top shell (Turbo cornutus) on articulated coralline algae; combination of preferences in settlement and post-settlement stage. J. Exp. Mar. Biol. Ecol. 2008, 363, 118–123. [Google Scholar] [CrossRef]
- Kim, Y.-D.; Park, M.-S.; Yoo, H.-I.; Min, B.-H.; Jin, H.-J. Seasonal variations of seaweed community structure at the subtidal zone of Bihwa on the East coast of Korea. Korean J. Fish. Aquat. Sci. 2012, 45, 262–270. [Google Scholar] [CrossRef]
- Kim, C.; Kim, Y.S.; Choi, H.G.; Nam, K.W. Variations of seaweed community structure and distribution of crustose coralline algae at Gallam, Samchuk, eastern coast of Korea. Korean J. Environ. Ecol. 2014, 28, 10–23. [Google Scholar] [CrossRef]
- Han, S.J.; Hwang, Y.H.; Son, M.H.; Choi, H.G.; Jang, J.G. Seasonal Variation in Seaweed Community Structure in the Subtidal Zone of the Southern Part of the East Coast of Korea. Korean J. Fish. Aquat. Sci. 2018, 51, 571–578. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Year | |||||||
|---|---|---|---|---|---|---|---|---|
| No. | Name | 2009 | 2010 | 2012 | 2015 | 2016 | 2017 | 2018 |
| 1 | Gageodo | ● | ● | ● | ||||
| 2 | Chujado | ● | ● | ● | ||||
| 3 | Cheongsando | ● | ● | ● | ||||
| 4 | Geomundo | ● | ● | |||||
| 5 | Geumodo | ● | ||||||
| 6 | Mijodo | ● | ||||||
| 7 | Maemuldo | ● | ● | |||||
| 8 | Jisimdo | ● | ● | |||||
| 9 | Namuseom | ● | ● | |||||
| 10 | Dongseom | ● | ● | |||||
| 11 | Moondong | ● | ● | |||||
| 12 | Jeongjadong | ● | ||||||
| 13 | Oryu | ● | ● | ● | ||||
| 14 | Samjeong | ● | ||||||
| 15 | Bangeo | ● | ● | |||||
| 16 | Chuksan | ● | ● | |||||
| 17 | Geoil | ● | ● | |||||
| 18 | Jukbyeon | ● | ● | |||||
| 19 | Nagok | ● |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, M.H.; Lee, C.I.; Park, J.M.; Kim, H.J.; Riedel, R.; Hwang, I.; Kim, Y.-N.; Jung, H.K. The Northward Habitat Expansion of the Korean Top Shell Turbo sazae (Gastropoda: Vetigastropoda: Turbinidae) in the Korean Peninsula: Effects of Increasing Water Temperature. J. Mar. Sci. Eng. 2020, 8, 782. https://doi.org/10.3390/jmse8100782
Son MH, Lee CI, Park JM, Kim HJ, Riedel R, Hwang I, Kim Y-N, Jung HK. The Northward Habitat Expansion of the Korean Top Shell Turbo sazae (Gastropoda: Vetigastropoda: Turbinidae) in the Korean Peninsula: Effects of Increasing Water Temperature. Journal of Marine Science and Engineering. 2020; 8(10):782. https://doi.org/10.3390/jmse8100782
Chicago/Turabian StyleSon, Min Ho, Chung Il Lee, Joo Myun Park, Hyun Jung Kim, Ralf Riedel, Inseo Hwang, Young-Nam Kim, and Hae Kun Jung. 2020. "The Northward Habitat Expansion of the Korean Top Shell Turbo sazae (Gastropoda: Vetigastropoda: Turbinidae) in the Korean Peninsula: Effects of Increasing Water Temperature" Journal of Marine Science and Engineering 8, no. 10: 782. https://doi.org/10.3390/jmse8100782
APA StyleSon, M. H., Lee, C. I., Park, J. M., Kim, H. J., Riedel, R., Hwang, I., Kim, Y.-N., & Jung, H. K. (2020). The Northward Habitat Expansion of the Korean Top Shell Turbo sazae (Gastropoda: Vetigastropoda: Turbinidae) in the Korean Peninsula: Effects of Increasing Water Temperature. Journal of Marine Science and Engineering, 8(10), 782. https://doi.org/10.3390/jmse8100782