The Effect of Glacier Recession on Benthic and Pelagic Communities: Case Study in Herve Cove, Antarctica

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Glacier’s Range
- Aerial photography taken on 20 December 1956 with a scale of 1:28,000 during the Falkland Island Dependency Aerial Survey Expedition (FIDASE);
- Aerial photography taken on 04 January 1979 with a scale of 1:3600 during the Second Polish Antarctic Expedition to H. Arctowski Polish Antarctic Station;
- Landsat 4 satellite images taken on 16 February 1990 with a resolution of 15 m.
2.3. Bathymetry
2.4. Sediments
2.5. Zoobenthos
2.6. Zooplankton
3. Results
3.1. Glacier Range, Bathymetry, Sediments
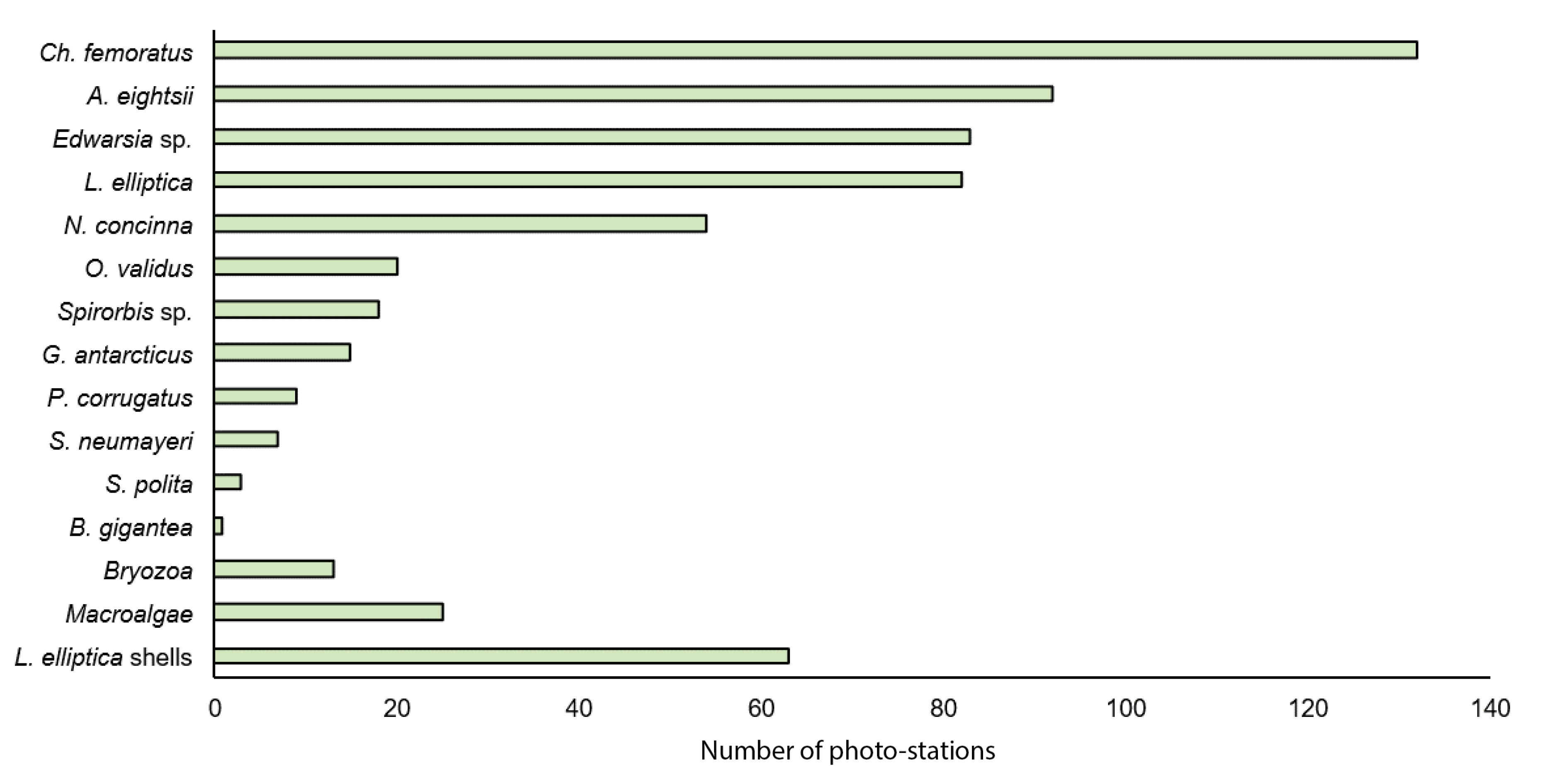
3.2. Zoobenthos
3.3. Zooplankton
4. Discussion
5. Conclusions
- Early colonization: Glacier retreated but still had contact with water; new bottom area opened but was still influenced by glacier discharge/meltwater/mineral suspension. Benthic animals can cross the submerged moraine and enter the lagoon, but conditions inside the lagoon are still unfavorable for settlement. Only tolerant species can survive in these conditions, such as anemones, amphipods and bivalves. This stage of colonization was most likely observed by Siciński et al. [25] in 1993.
- Colonization: Glacier retreated, and no contact with the water. The whole lagoon is open, and the influence of the glacier was less significant. Meltwater discharge and mineral suspension are significant seasonally and only affect small areas of the cove. Planktonic larvae of benthic species can enter the cove and settle on the bottom. Macrofauna is also present at the far end of the cove, which provided favorable conditions for its larvae. There was higher biodiversity, but still a lack of many species. This stage of colonization was observed in 2008.
- Late colonization/climax: Glacier has retreated far up the hill-side, no influence of meltwater. Colonization of more sensitive species. Biodiversity almost the same as that in the open water of the Ezcurra Inlet. The available climatic scenarios suggest that this bay should be an ideal place to analyze this stage of colonization in the near future, perhaps within 10–20 years.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Taxa | Our Station | Siciński et al., 1996 | Kittel et al., 2001 |
|---|---|---|---|
| Siphonophorae | - | + | + |
| Nematoda | + | + | + |
| Polychaeta | |||
| Pelagobia longicirrata Greeff, 1879 | - | + | + |
| Polychaeta larvae | + | + | + |
| Gastropoda larvae | + | - | - |
| Gastropoda/Pteropoda | |||
| Limacina rangii f. antarctica Woodward, 1854 | - | + | + |
| Ostracoda | |||
| Alacia belgicae Müller, G.W. (1906) | - | + | + |
| Alacia hettacra Müller, G.W. (1906) | - | + | + |
| Copepoda | |||
| Calanus/Calanoides (nauplii and copepodit I-III) | + | ||
| Calanus propinqus Brady, 1883 | + | + | + |
| Calanoides acutus Giesbrecht, 1902 | + | + | + |
| Ctenocalanus citer Heron et Bowman, 1971 | + | + | + |
| Microcalanus pygmaeus G.O. Sars, 1903 | + | + | + |
| Euchaeta antarctica Giesbrecht, 1902 | + | + | + |
| Rhincalanus gigas Brady, 1883 | - | + | + |
| Racovitzanus antarcticus Giesbrecht, 1902 | - | + | + |
| Scaphocalanus spp. | - | + | + |
| Scolecithricella glacialis Giesbrecht, 1902 | + | + | + |
| Stephos longipes Giesbrecht, 1902 | - | + | + |
| Metridia gerlachei Giesbrecht, 1902 | + | + | + |
| Oithona frigida Giesbrecht, 1902 | + | + | + |
| Oithona similis Claus, 1863 | + | + | + |
| Oncaea antarctica Heron, 1977 | + | + | + |
| Oncaea curvata Giesbrecht, 1902 | + | + | + |
| Oncaea parila Heron, 1977 | + | - | - |
| Oncea sp. | + | - | - |
| Harpacticoida | + | + | + |
| Isopoda | + | - | - |
| Cumacea | |||
| Eudorella splendida Zimmer, 1902 | + | - | - |
| Euphausiacea | |||
| Thysanoessa macrura G.O. Sars, 1883 | + | - | - |
| T.macrura – larvae | + | - | - |
| Euphausia crystallorophias Holt et Tattersall, 1906 | + | - | - |
| Chaetognatha | |||
| Eukrohnia hamata Mobius, 1875 | + | + | + |
| Pseudosagitta gazelae Ritter-Záhony, 1909 | + | + | + |
| Solidosagitta marri David, 1956 | + | + | + |
| Chaetognatha uind. | + | ||
| Appendiculariae | |||
| Frittilaria sp. | + | - | - |
| Ascidiacea larvae | + | + | - |
References
- Hoegh-Guldberg, O.; Bruno, J.F. The impact of climate change on the world’s marine ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Pasotti, F.; Manini, E.; Giovannelli, D.; Wölfl, A.C.; Monien, D.; Verleyen, E.; Braeckman, U.; Abele, D.; Vanreusel, A. Antarctic shallow water benthos in an area of recent rapid glacier retreat. Mar. Ecol. 2015, 36, 716–733. [Google Scholar] [CrossRef]
- Sahade, R.; Lagger, C.; Torre, L.; Momo, F.; Monien, P.; Schloss, I.; Barnes, D.K.; Servetto, N.; Tarantelli, S.; Tatián, M.; et al. Climate change and glacier retreat drive shifts in an Antarctic benthic ecosystem. Sci. Adv. 2015, 1, e1500050. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.W.; Wan Hussin, W.M.R.; Kim, H.C.; Ahn, I.Y. The impacts of climate change on Antarctic nearshore mega-epifaunal benthic assemblages in a glacial fjord on King George Island: Responses and implications. Ecol. Indic. 2015, 57, 280–292. [Google Scholar] [CrossRef]
- Simoes, C.L.; Rosa, K.K.D.; Czapela, F.F.; Vieira, R.; Simoes, J.C. Collins Glacier retreat process and regional climatic variations, King George Island, Antarctica. Geogr. Rev. 2015, 105, 462–471. [Google Scholar] [CrossRef]
- Sobota, I.; Kejna, M.; Araźny, A. Short-term mass changes and retreat of the Ecology and Sphinx glacier system, King George Island, Antarctic Peninsula. Antarct. Sci. 2015, 27, 500–510. [Google Scholar] [CrossRef]
- Oliva, M.; Ruiz-Fernández, J. Geomorphological processes and frozen ground conditions in Elephant Point (Livingston Island, South Shetland Islands, Antarctica). Geomorphology 2017, 293, 368–379. [Google Scholar] [CrossRef]
- Pętlicki, M.; Sziło, J.; MacDonell, S.; Vivero, S.; Bialik, R.J. Recent deceleration of the ice elevation change of ecology glacier (King George Island, Antarctica). Remote Sens. 2017, 9, 520. [Google Scholar] [CrossRef]
- Pudełko, R.; Angiel, P.J.; Potocki, M.; Jędrejek, A.; Kozak, M. Fluctuation of Glacial Retreat Rates in the Eastern Part of Warszawa Icefield, King George Island, Antarctica, 1979–2018. Remote Sens. 2018, 10, 892. [Google Scholar] [CrossRef]
- Thatje, S.; Hillenbrand, C.D.; Larter, R. On the origin of Antarctic marine benthic community structure. Trends Ecol. Evol. 2005, 20, 534–540. [Google Scholar] [CrossRef]
- Quartino, M.L.; Deregibus, D.; Campana, G.L.; Latorre, G.E.J.; Momo, F.R. Evidence of macroalgal colonization on newly ice-free areas following glacial retreat in Potter Cove (South Shetland Islands), Antarctica. PLoS ONE 2013, 8, e58223. [Google Scholar] [CrossRef] [PubMed]
- Włodarska-Kowalczuk, M.; Węsławski, J.M. Impact of climate warming on Arctic benthic biodiversity: A case study of two Arctic glacial bays. Clim. Res. 2001, 18, 127–132. [Google Scholar] [CrossRef]
- Husmann, G.; Abele, D.; Monien, D.; Monien, P.; Kriews, M.; Philipp, E.E.R. The influence of sedimentation on metal accumulation and cellular oxidative stress markers in the Antarctic bivalve Laternula elliptica. Estuar. Coast. Shelf Sci. 2012, 111, 48–59. [Google Scholar] [CrossRef]
- Siciński, J.; Pabis, K.; Jażdżewski, K.; Konopacka, A.; Błażewicz-Paszkowycz, M. Macrozoobenthos of two Antarctic glacial coves: A comparison with non-disturbed bottom areas. Polar Biol. 2012, 35, 355–367. [Google Scholar] [CrossRef]
- Zajączkowski, M. Sediment supply and fluxes in glacial and outwash fjords, Kongsfjorden and Adventfjorden, Svalbard. Pol. Polar Res. 2008, 29, 59–72. [Google Scholar]
- Sziło, J.; Bialik, R.J. Bedload transport in two creeks at the ice-free area of the Baranowski Glacier, King George Island, West Antarctica. Pol. Polar Res. 2017, 38, 21–39. [Google Scholar] [CrossRef]
- Sziło, J.; Bialik, R.J. Recession and Ice Surface Elevation Changes of Baranowski Glacier and Its Impact on Proglacial Relief (King George Island, West Antarctica). Geosciences 2018, 8, 355. [Google Scholar] [CrossRef]
- Smale, D.A. Continuous benthic community change along a depth gradient in Antarctic shallows: Evidence of patchiness but not zonation. Polar Biol. 2008, 31, 189–198. [Google Scholar] [CrossRef]
- Kejna, M.; Araźny, A.; Sobota, I. Climatic change on King George Island in the years 1948–2011. Pol. Polar Res. 2013, 34, 213–235. [Google Scholar] [CrossRef]
- Oliva, M.; Navarro, F.; Hrbáˇcek, F.; Hernández, A.; Nyvlt, D.; Pereira, P.; Ruiz-Fernández, J.; Trigo, R. Recent regional climate cooling on the Antarctic Peninsula and associated impacts on the cryosphere. Sci. Total Environ. 2017, 580, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Nonato, E.F.; Brito, T.A.S.; Paiva, P.C.; Petti, M.A.V.; Corbisier, T.N. Benthic megafauna of the nearshore zone of Martel Inlet (King George Island, South Shetland Islands, Antarctica): Depth zonation and underwater observations. Polar Biol. 2000, 23, 580–588. [Google Scholar] [CrossRef]
- Jażdzewski, K.; De Broyer, C.; Pudlarz, M.; Zielinski, D. Seasonal fluctuations of vagile benthos in the uppermost sublittoral of a maritime Antarctic fjord. Polar Biol. 2001, 24, 910–917. [Google Scholar] [CrossRef]
- Siciński, J.; Jażdżewski, K.; De Broyer, C.; Presler, P.; Ligowski, R.; Nonato, E.F.; Corbisier, T.N.; Petti, M.A.; Brito, T.A.; Lavrado, H.P.; et al. Admiralty Bay Benthos Diversity—A census of a complex polar ecosystem. Deep Sea Res. Part 2 Top. Stud. Oceanogr. 2010, 58, 30–48. [Google Scholar] [CrossRef]
- Gaździcki, A.; Majewski, W. Recent Foraminifera from Goulden Cove of King George Island, Antarctica. Pol. Polar Res. 2003, 24, 3–12. [Google Scholar]
- Siciński, J.; Rożycki, O.; Kittel, W. Zoobenthos and zooplankton of Herve Cove, King George Island, South Shetland Islands, Antarctic. Pol. Polar Res. 1996, 17, 221–238. [Google Scholar]
- Rakusa-Suszczewski, S. The hydrography of Admiralty Bay and its inlets, coves and lagoons (King George Island, Antarctica). Pol. Polar Res. 1995, 16, 61–70. [Google Scholar]
- Pabis, K.; Siciński, J.; Krymarys, M. Distribution patterns in the biomass of macrozoobenthic communities in Admiralty Bay (King George Island, South Shetlands, Antarctic). Polar Biol. 2011, 34, 489–500. [Google Scholar] [CrossRef][Green Version]
- Lagger, C.; Nime, M.; Torre, L.; Servetto, N.; Tatián, M.; Sahade, R. Climate change, glacier retreat and a new ice-free island offer new insights on Antarctic benthic responses. Ecography 2017, 41, 579–591. [Google Scholar] [CrossRef]
- Markowska, M.; Kidawa, A. Encounters between Antarctic limpets, Nacella concinna, and predatory sea stars, Lysasterias sp., in laboratory and field experiments. Mar. Biol. 2007, 151, 1959–1966. [Google Scholar] [CrossRef]
- Kittel, W.; Siciński, J.; Żmijewska, M.I.; Bielecka, L.; Blachowiak-Samołyk, K. Antarctic neritic zooplankton community (Admiralty Bay, King George Island, South Shetland Islands). Pol. Polar Res. 2001, 22, 11–33. [Google Scholar]
- Jażdżewski, K.; Jurasz, W.; Kittel, W.; Presler, E.; Presler, P.; Siciński, J. Abundance and biomass estimates of the benthic fauna in Admiralty Bay, King George Island, South Shetland Islands. Polar Biol. 1986, 61, 5–16. [Google Scholar] [CrossRef]
- Stanwell-Smith, D.; Barnes, D.K.A. Benthic community development in Antarctica: Recruitment and growth on settlement panels at Signy Island. J. Exp. Mar. Biol. Ecol. 1997, 212, 61–79. [Google Scholar] [CrossRef]
- Strathmann, R.R.; Kendall, L.R.; Marsh, A.G. Embryonic and larval development of a cold adapted Antarctic ascidian. Polar Biol. 2006, 29, 495–501. [Google Scholar] [CrossRef]
- Segelken-Voigt, A.; Bracher, A.; Dorschel, B.; Gutt, J.; Huneke, W.; Link, H.; Piepenburg, D. Spatial distribution patterns of ascidians (Ascidiacea: Tunicata) on the continental shelves off the northern Antarctic Peninsula. Polar Biol. 2016, 39, 863–879. [Google Scholar] [CrossRef]
- Netto, S.A.; Fonseca, G. Regime shifts in coastal lagoons: Evidence from free-living marine nematodes. PLoS ONE 2017, 12, e0172366. [Google Scholar] [CrossRef] [PubMed]
- Obolewski, K.; Glińska-Lewczuk, K.; Bąkowska, M. From isolation to connectivity: The effect of floodplain lake restoration on sediments as habitats for macroinvertebrate communities. Aquat. Sci. 2018, 80, 4. [Google Scholar] [CrossRef]
- Peck, L.S.; Brockington, S.; Vanhove, S.; Beghyn, M. Community recovery following catastrophic iceberg impacts in a soft-sediment shallow-water site at Signy Island, Antarctica. Mar. Ecol. Prog. Ser. 1999, 186, 1–8. [Google Scholar] [CrossRef]
- Loque, C.P.; Medeiros, A.O.; Pellizzari, F.M.; Oliveira, E.C.; Rosa, C.A.; Rosa, L.H. Fungal community associated with marine macroalgae from Antarctica. Polar Biol. 2010, 33, 641–648. [Google Scholar] [CrossRef]
- Cacabelos, E.; Olabarria, C.; Incera, M.; Troncoso, J.S. Effects of habitat structure and tidal height on epifaunal assemblages associated with macroalgae. Estuar. Coast. Shelf Sci. 2010, 89, 43–52. [Google Scholar] [CrossRef]
- Torres, A.C.; Veiga, P.; Rubal, M.; Sousa-Pinto, I. The role of annual macroalgal morphology in driving its epifaunal assemblages. J. Exp. Mar. Biol. Ecol. 2015, 464, 96–106. [Google Scholar] [CrossRef]
- Mercuri, G.; Tatián, M.; Momo, F.; Fuentes, V.; Sahade, R. Massive input of terrigenous sediment into Potter Cove during austral summer and the effects on the bivalve Laternula elliptica: A laboratory experiment. Berichte Polar Meeresforsch. 2008, 571, 111–117. [Google Scholar]
- Stockton, W.L. The biology and ecology of the epifaunal scallop Adamussium colbecki on the west side of McMurdo Sound, Antarctica. Mar. Biol. 1984, 78, 171–178. [Google Scholar] [CrossRef]
- Barnes, D.K.; Conlan, K.E. Disturbance, colonization and development of Antarctic benthic communities. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Davenport, J. Feeding, oxygen uptake, ventilation rate and shell growth in the Antarctic protobranch bivalve mollusc Yoldia eightsi (Courthony). In Proceedings of the British Antarctic Survey Antarctic Special Topic Award Scheme Symposium; Heywood, R.B., Ed.; British Antarctic Survey: Cambridge, UK, 1989; pp. 57–63. [Google Scholar]
- Peck, L.S.; Ansell, A.D.; Webb, K.E.; Hepburn, L.; Burrows, M. Movements and burrowing activity in the Antarctic bivalve molluscs Laternula elliptica and Yoldia eightsi. Polar Biol. 2004, 27, 357–367. [Google Scholar] [CrossRef]
- Philipp, E.E.; Husmann, G.; Abele, D. The impact of sediment deposition and iceberg scour on the Antarctic soft shell clam Laternula elliptica at King George Island, Antarctica. Antarct. Sci. 2011, 23, 127–138. [Google Scholar] [CrossRef]
- Momo, F.R.; Sahade, R.; Tatián, M. Benthic animal communities of Potter Cove (King George Island, Antarctica): Observed patterns and explanatory models. Berichte Polar Meeresforsch. 2008, 571, 162–167. [Google Scholar]



| Transects Length (m) | No. of Photo Stations | Intervals (m) | |
|---|---|---|---|
| H1-H2 | 57 | 12 | 5 |
| H3-H4 | 55 | 8 | 7 |
| H5-H6 | 299 | 18 | 15 |
| H7-H8 | 400 | 42 | 10 |
| H9-H10 | 268 | 24 | 10 |
| H11-H12 | 261 | 35 | 7 |
| H13-H14 | 93 | 15 | 6 |
| H15-H16 | 121 | 17 | 7 |
| Phylum | Order | Genus/Species | Assemblage | |||
|---|---|---|---|---|---|---|
| A | B | B1 | C | |||
| Mollusca | Gastropoda | Nacella concinna | - | + | + | - |
| Bivalvia | Laternula elliptica | + | + | - | + | |
| Aequiyoldia eightsii | + | + | - | + | ||
| Echinodermata | Echinoidea | Sterechinus neumayeri | + | + | + | - |
| Asteroidea | Odontaster validus | + | + | + | - | |
| Cnidaria | Acinaria | Edwarsia sp. | + | + | + | + |
| Nemertea | Heteronemertea | Parborlasia corrugata | - | + | + | - |
| Arthropoda | Isopoda | Glyptonotus antarcticus | + | + | + | - |
| Serolis polita | - | + | - | - | ||
| Amphipoda | Cheirimedon femoratus | + | + | + | + | |
| Bovallia gigantea | - | - | + | - | ||
| Annelida | Polychaeta | Spirorbis sp. | - | + | + | - |
| Bryozoa | Bryozoa uind. | - | + | + | - | |
| Macroalgae | - | + | + | - | ||
| Periphyton | + | + | + | - | ||
| Other Notothenia coriiceps | + | - | - | - | ||
| Phylum | Taxon | 13.01.2009 | 22.01.2009 | 05.02.2009 | 20.02.2009 | 05.03.2009 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A | % | A | % | A | % | A | % | A | % | ||
| Nematoda | Nematoda | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 475.3 | 0.4 | 0.0 | 0.0 |
| Annelida | Polychaeta larvae | 502.7 | 0.1 | 1025.6 | 0.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Mollusca | Gastropoda larvae | 0.0 | 0.0 | 256.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Arthropoda | Copepoda | ||||||||||
| Calanoida | 15,932.8 | 3.7 | 38,974.4 | 4.8 | 8422.6 | 23.6 | 4039.7 | 3.7 | 14,545.1 | 3.8 | |
| Cyclopoida | 139,422.9 | 32.7 | 178,461.5 | 22.0 | 20,620.8 | 57.7 | 87,446.9 | 79.8 | 97,810.5 | 25.3 | |
| Harpacticoida | 268,738.1 | 63.1 | 583,333.3 | 72.0 | 1452.2 | 4.1 | 11,406.1 | 10.4 | 274,038.2 | 70.8 | |
| Isopoda | 0.0 | 0.0 | 0.0 | 0.0 | 290.4 | 0.8 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Cumacea | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 237.6 | 0.2 | 0.0 | 0.0 | |
| Euphausiacea | |||||||||||
| Thysanoessa macrura | 128.1 | 0.0 | 256.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 210.8 | 0.1 | |
| Euphausia crystallorophias | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 210.8 | 0.1 | |
| Chaetognatha | Eukrohnia hamata | 0.0 | 0.0 | 2564.1 | 0.3 | 290.4 | 0.8 | 0.0 | 0.0 | 210.8 | 0.1 |
| Pseudosagitta gazelae | 0.0 | 0.0 | 256.4 | 0.0 | 290.4 | 0.8 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Solidosagitta marri | 0.0 | 0.0 | 256.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Chaetognatha uind. | 630.8 | 0.1 | 1794.9 | 0.2 | 1161.7 | 3.3 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Chordata | Appendiculariae | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 950.5 | 0.9 | 0.0 | 0.0 |
| Ascidiacea larvae | 379.5 | 0.1 | 2820.5 | 0.3 | 3194.8 | 8.9 | 4990.2 | 4.6 | 210.8 | 0.1 | |
| Total abundance (ind.∙1000 m−3) | 425,734.9 | 810,000.0 | 35,723.3 | 10,9546.3 | 387,237.0 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potocka, M.; Kidawa, A.; Panasiuk, A.; Bielecka, L.; Wawrzynek-Borejko, J.; Patuła, W.; Wójcik, K.A.; Plenzler, J.; Janecki, T.; Bialik, R.J. The Effect of Glacier Recession on Benthic and Pelagic Communities: Case Study in Herve Cove, Antarctica. J. Mar. Sci. Eng. 2019, 7, 285. https://doi.org/10.3390/jmse7090285
Potocka M, Kidawa A, Panasiuk A, Bielecka L, Wawrzynek-Borejko J, Patuła W, Wójcik KA, Plenzler J, Janecki T, Bialik RJ. The Effect of Glacier Recession on Benthic and Pelagic Communities: Case Study in Herve Cove, Antarctica. Journal of Marine Science and Engineering. 2019; 7(9):285. https://doi.org/10.3390/jmse7090285
Chicago/Turabian StylePotocka, Marta, Anna Kidawa, Anna Panasiuk, Luiza Bielecka, Justyna Wawrzynek-Borejko, Weronika Patuła, Kornelia A. Wójcik, Joanna Plenzler, Tomasz Janecki, and Robert J. Bialik. 2019. "The Effect of Glacier Recession on Benthic and Pelagic Communities: Case Study in Herve Cove, Antarctica" Journal of Marine Science and Engineering 7, no. 9: 285. https://doi.org/10.3390/jmse7090285
APA StylePotocka, M., Kidawa, A., Panasiuk, A., Bielecka, L., Wawrzynek-Borejko, J., Patuła, W., Wójcik, K. A., Plenzler, J., Janecki, T., & Bialik, R. J. (2019). The Effect of Glacier Recession on Benthic and Pelagic Communities: Case Study in Herve Cove, Antarctica. Journal of Marine Science and Engineering, 7(9), 285. https://doi.org/10.3390/jmse7090285