Seasonal Changes in the Intertidal and Subtidal Algal Communities of Extremely and Moderately Polluted Coastal Regions of Sanya Bay (Hainan Island, China)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site, Time, and Conditions
2.1.1. Hainan Island
2.1.2. Time and Sites of Samplings
2.2. Collection, Conservation, and Identification of Marine Algae
2.3. Statistical Analysis
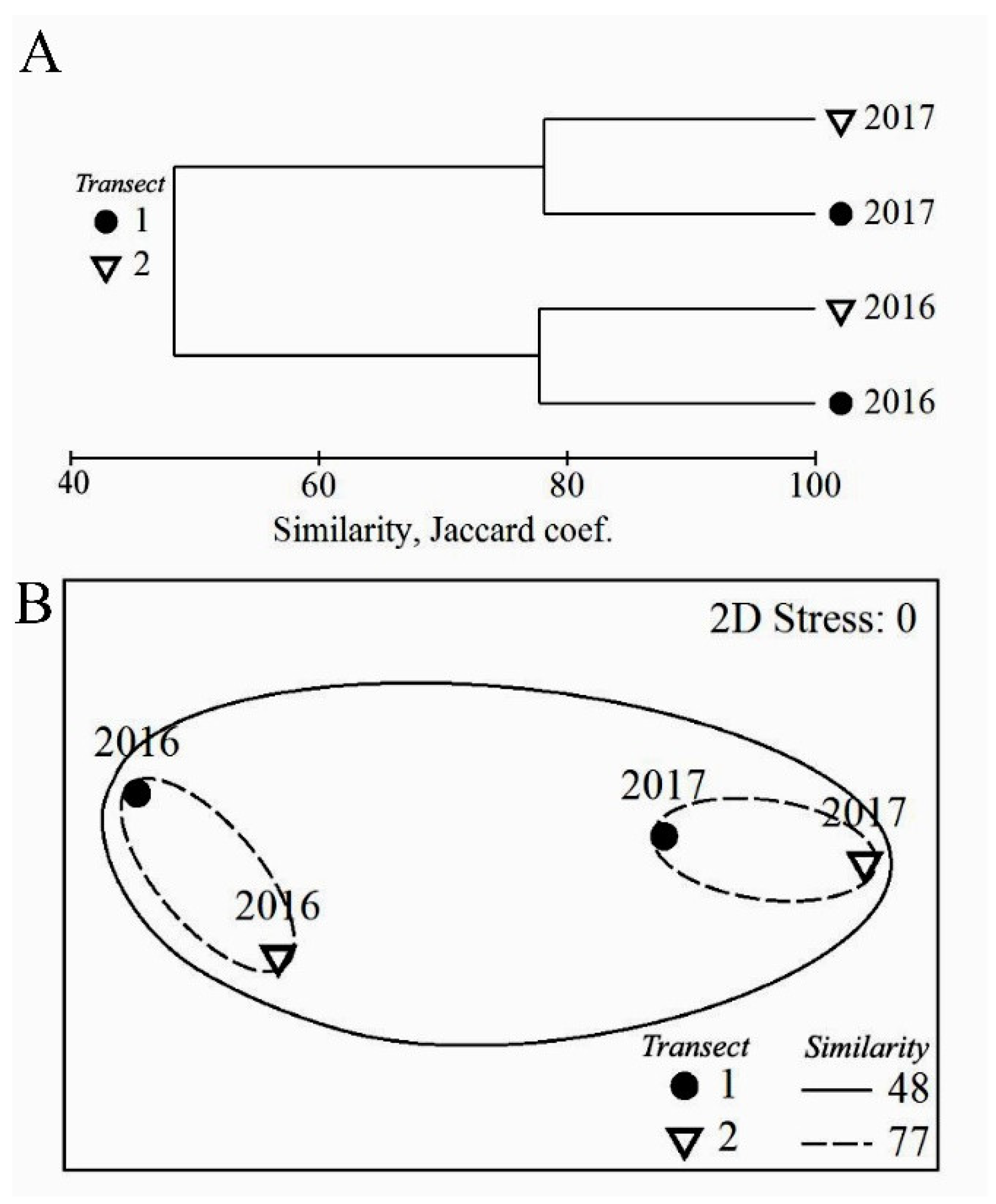
3. Results
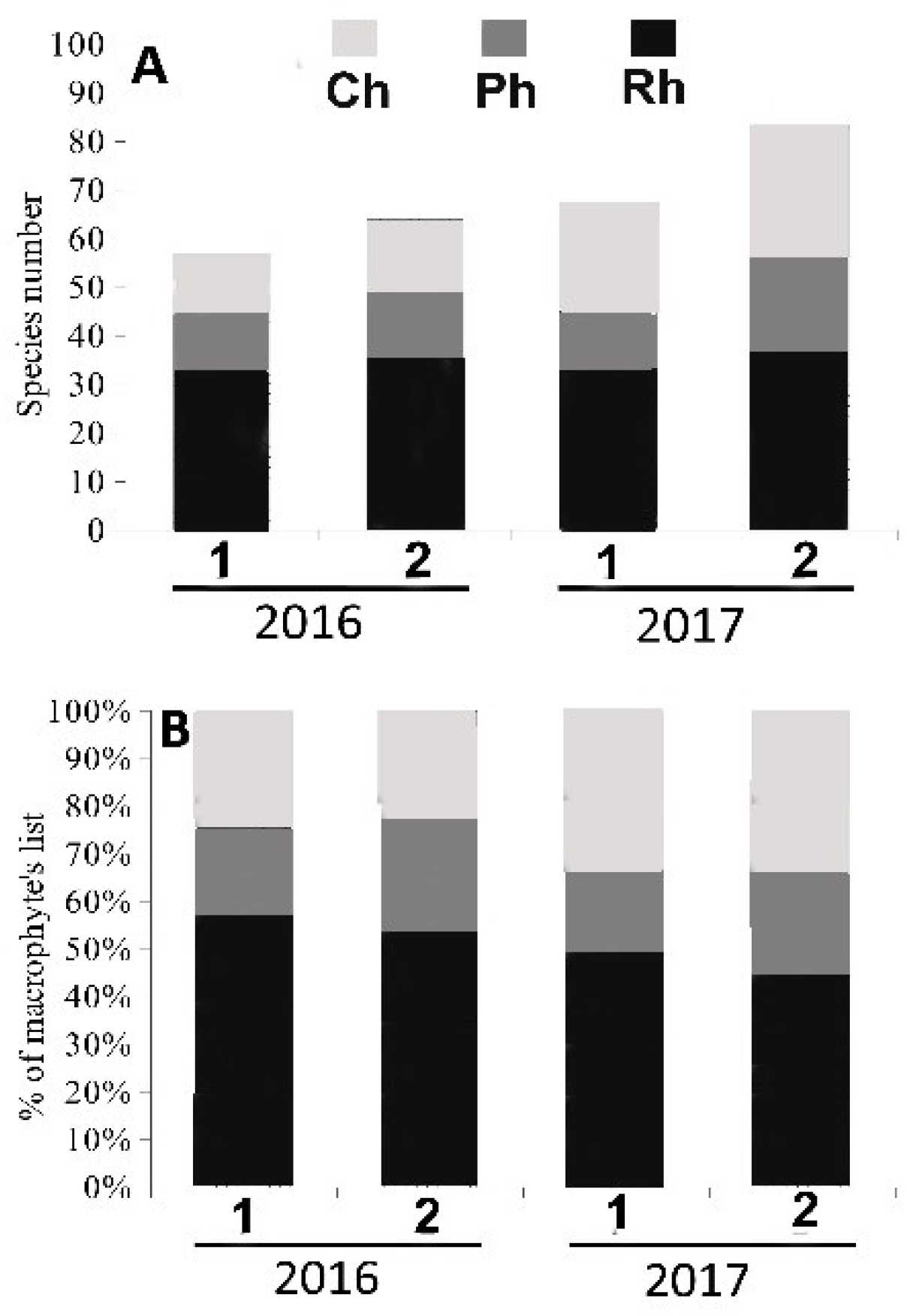
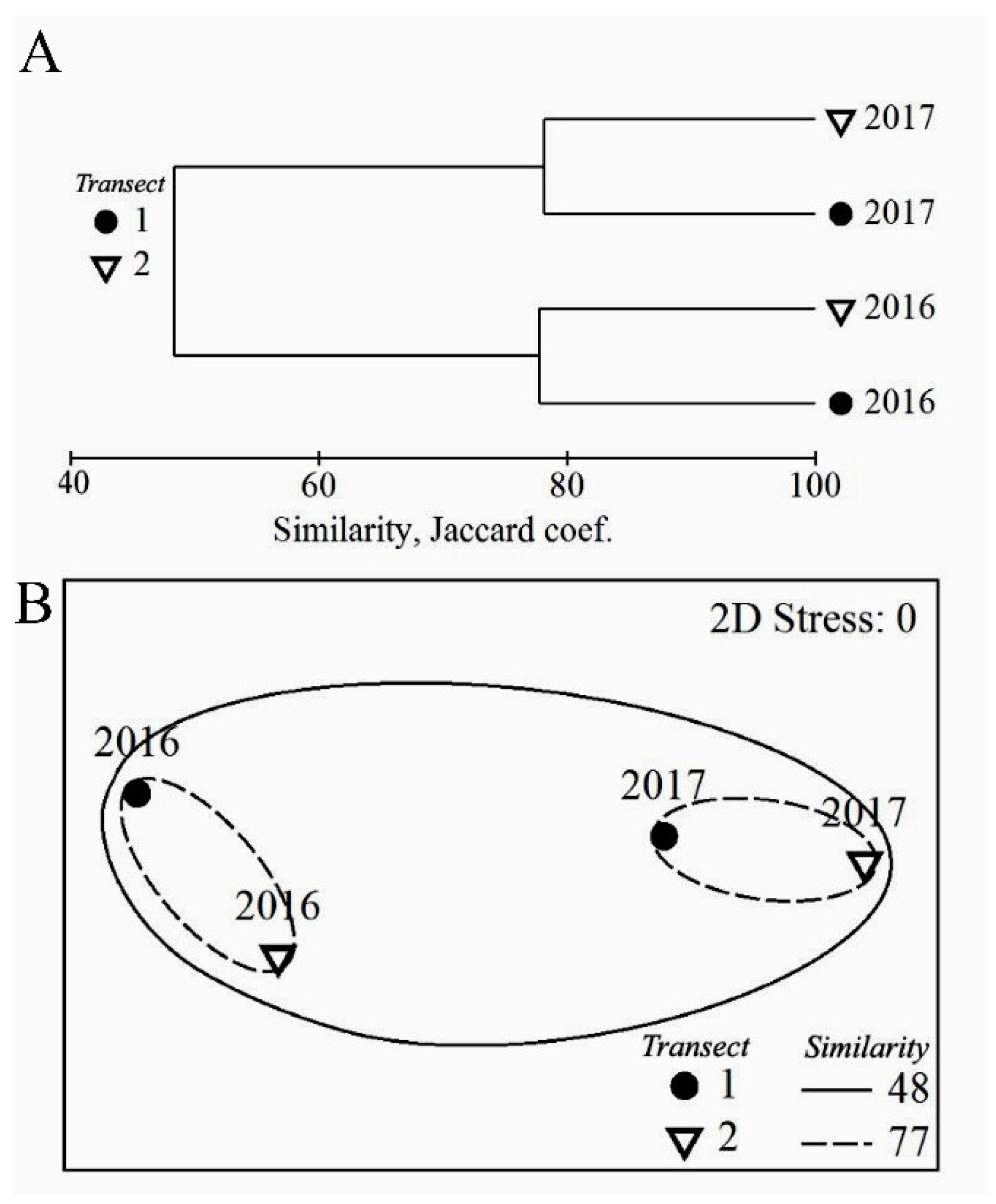
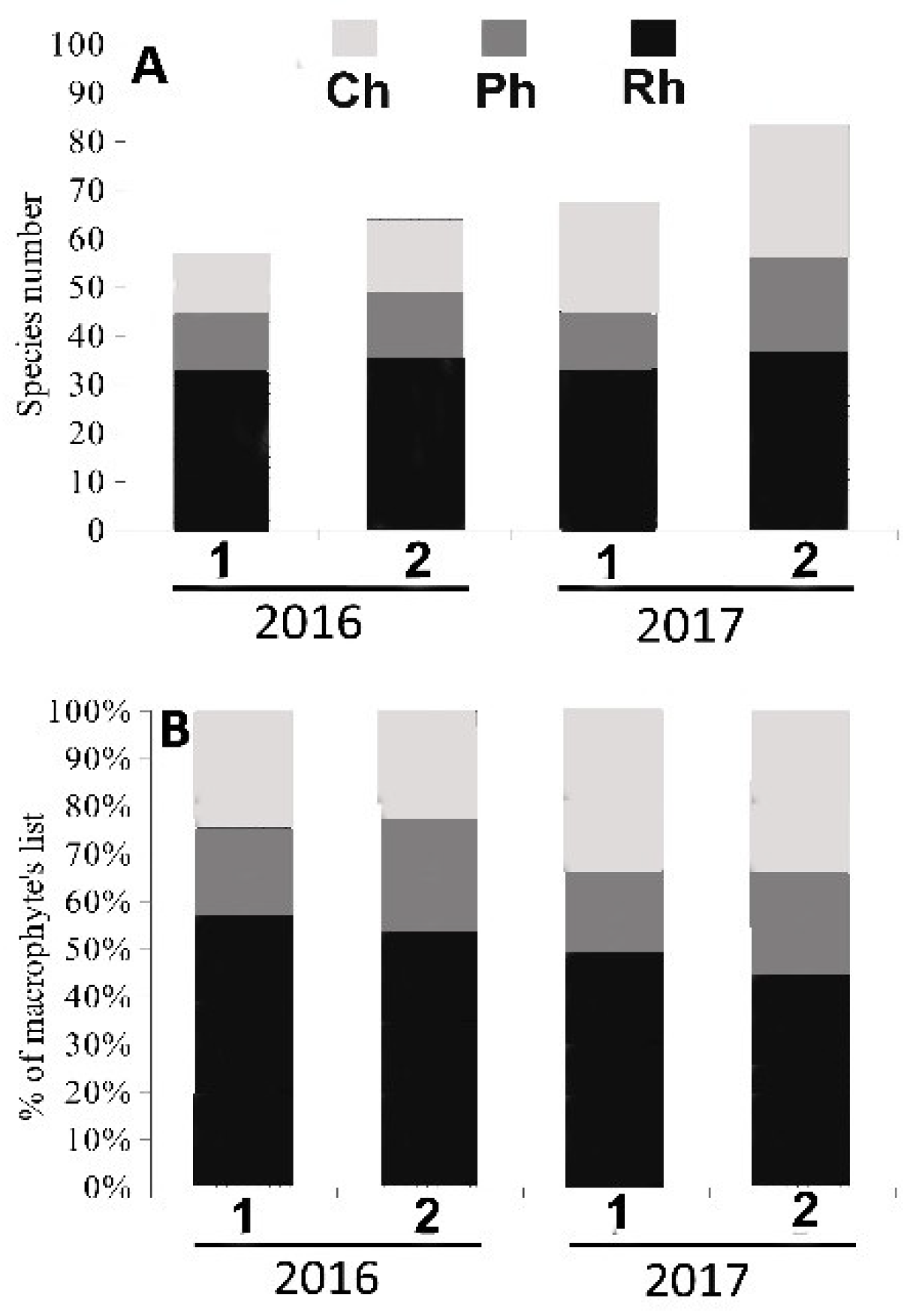
3.1. Algal Species Diversity, Taxonomic Composition and the Structure of Algal Communities in Heavily and Moderately Polluted Areas at the End of the Rainy Season and at the End of the Dry Season
Species Diversity and Taxonomic Composition
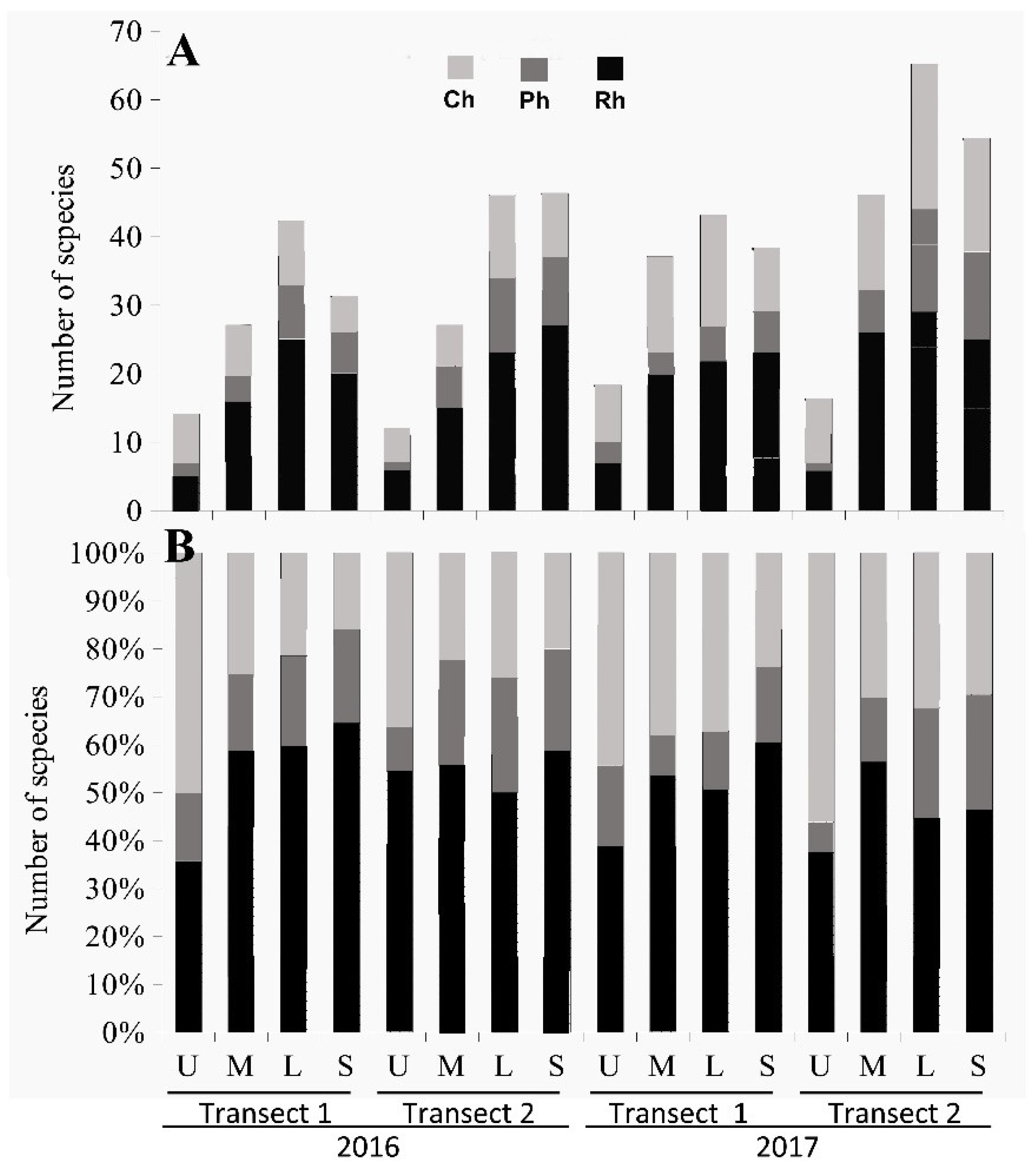
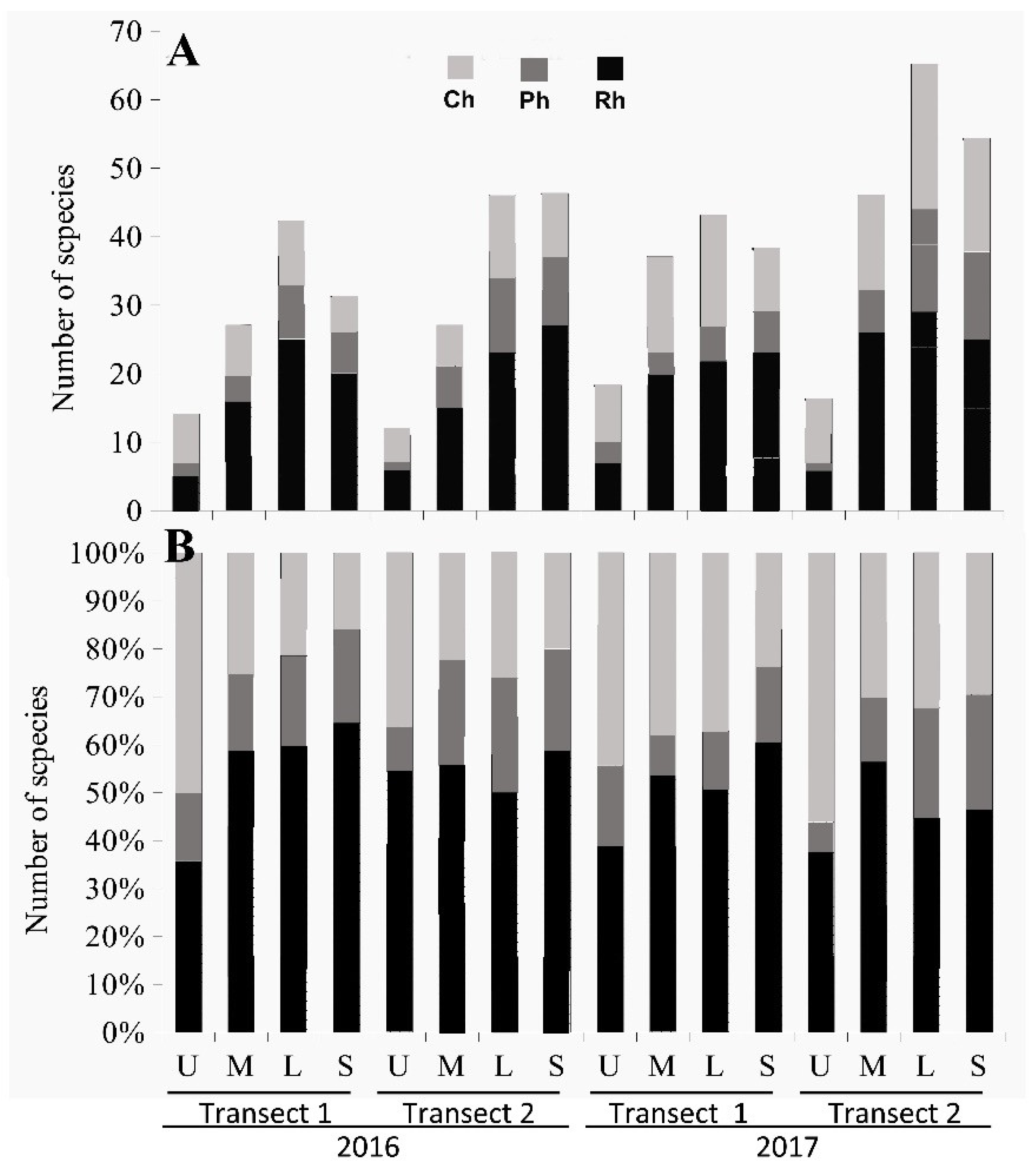
3.2. The Structure of Algal Communities and Their Distribution in Tidal Zones
3.2.1. The End of the Rainy Season
Heavily Polluted Area (Transect 1)
Moderately Polluted Area (Transect 2)
3.2.2. The End of the Dry Season
Heavily Polluted Area (Transect 1)
Moderately Polluted Area (Transect 2).
4. Discussion
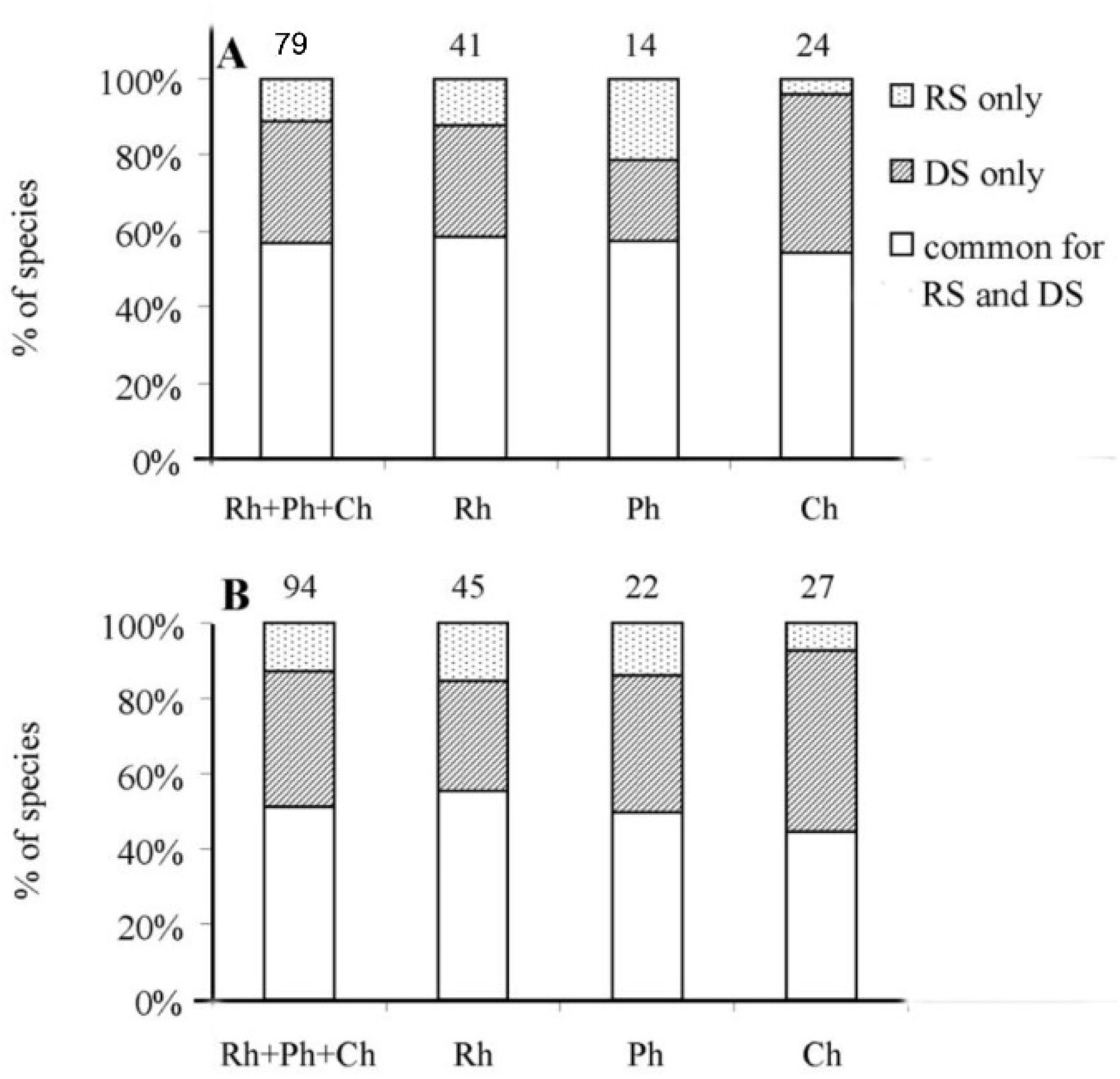
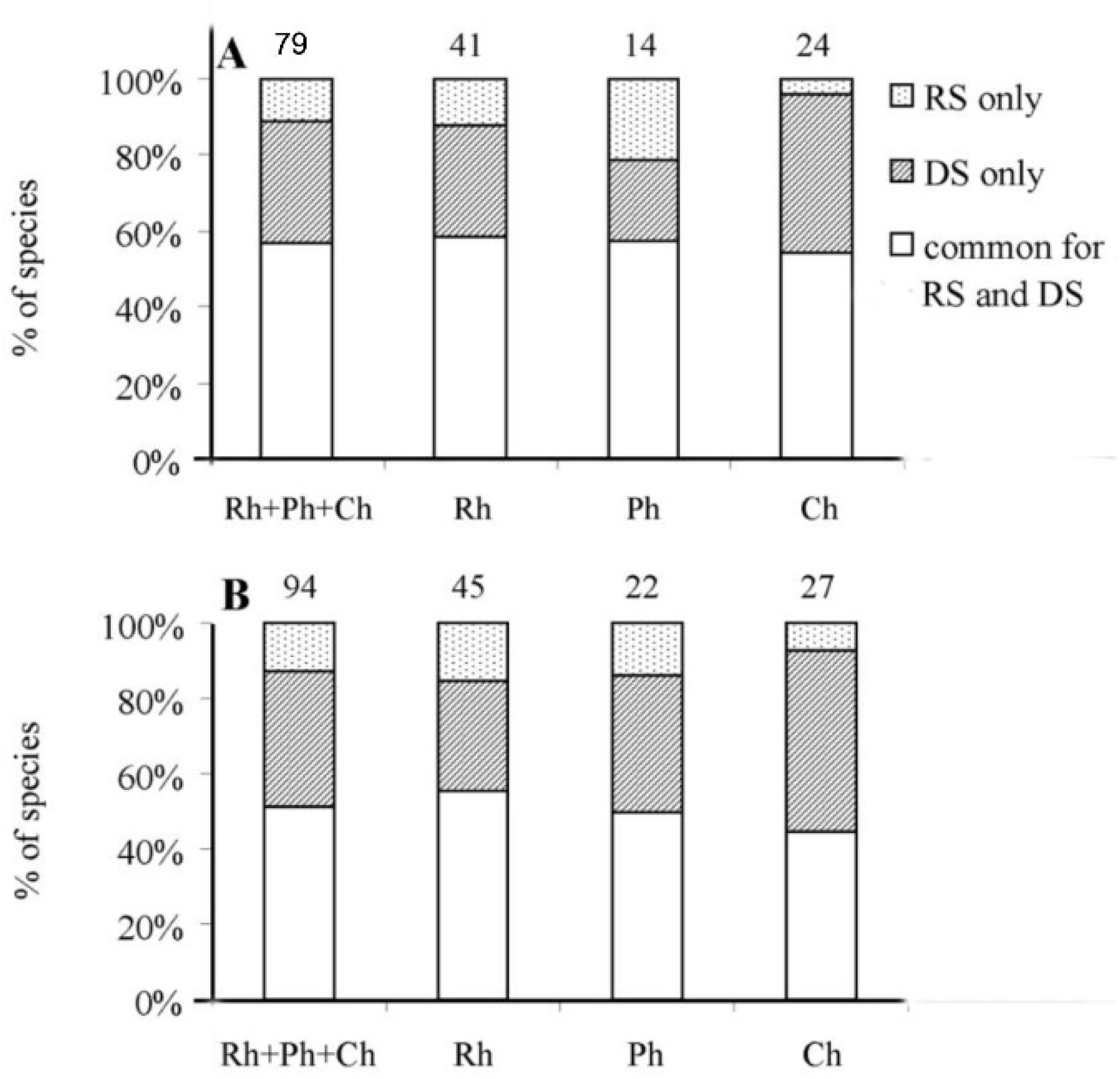
4.1. Richness and Taxonomic Composition of the Marine Flora of Hainan Island
4.2. Seasonal Changes in the Benthic Flora in Rainy and Dry Seasons
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fong, P.; Zedler, J.B. Temperature and light effects on the seasonal succession of algal communities in shallow coastal ecosystems. J. Exp. Mar. Biol. Ecol. 1993, 171, 259–272. [Google Scholar] [CrossRef]
- Kennish, R. Diet composition influences the fitness of the herbivorous crab Grapsus albolineatus. Oecologia 1996, 105, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.F.; Borum, J. Nutrient control of estuarine macroalgae: Growth strategy and the balance between nitrogen requirements and uptake. Mar. Ecol. Prog. Ser. 1996, 161, 155–163. [Google Scholar] [CrossRef]
- Su, S.W.; Chung, I.C.; Lee, T.M. Temporal dynamics of rockyshore macroalgal assemblage structures in relation to coastal construction threats in Orchard Island (Taiwan): Impacts of turbidity and nutrients on the blooms of Galaxaura oblongata and a red alga-spongesymbiose Ceratodictyon/Haliclona. Kuroshio Sci. 2009, 3, 63–80. [Google Scholar]
- Lopez Gappa, J.; Tablado, A.; Magaldi, N.H. Seasonal changes in an intertidal community affected by sewage pollution. Environ. Pollut. 1993, 82, 157–165. [Google Scholar] [CrossRef]
- Costa, O.S., Jr.; Zman, L.; Nimmo, M.; Attrill, M.J. Nitrification impacts on coral reefs from northern Bahia, Brazil. Hydrobiologia 2000, 440, 307–315. [Google Scholar] [CrossRef]
- Costa, O.S., Jr.; Attrilla, M.J.; Pedrinib, A.G.; De-Paulab, J.C. Spatial and seasonal distribution of seaweeds on coral reefs from southern Bahia, Brazil. Bot. Mar. 2002, 45, 346–355. [Google Scholar] [CrossRef]
- Mayakun, J.; Prathep, A. Seasonal variations in diversity and abundance of macroalgae at Samui Island, Surat Thani Province, Thailand Songklanakarin. J. Sci. Technol. 2005, 27, 653–663. [Google Scholar]
- Ateweberhan, M.; Bruggemann, J.H.; Breeman, A.M. Effects of extreme seasonality on community structure and functional group dynamics of coral reef algae in the southern Red Sea (Eritrea). Coral Reefs 2006, 25, 391–406. [Google Scholar] [CrossRef]
- García, C.B.; Díaz-Pulido, G. Dynamics of a macroalgal rocky intertidal community in the Colombian Caribbean. Bol. Investig. Mar. Cost. 2006, 35, 7–18. [Google Scholar] [CrossRef]
- Lirman, D.; Deangelo, G.; Serafy, J.; Hazra, A.; Hazra Smith, D.; Herlan, J.; Luo, J.; Bellmund, S.; Wang, J.; Clausing, R. Seasonal changes in the abundance and distribution of submerged aquatic vegetation in a highly managed coastal lagoon. Hydrobiologia 2008, 596, 105–120. [Google Scholar] [CrossRef]
- Thakur, M.C.; Reddy, C.R.K.; Jha, B. Seasonal variation in biomass and species composition of seaweeds stranded along Port Okha, northwest coast of India. J. Earth Syst. Sci. 2008, 117, 211–218. [Google Scholar] [CrossRef]
- Maraşlıoğlu, F.; Soylu, E.N.; Karaca, S.A. Seasonal and spatial variation of epilithic algal community in Batlama Stream (Giresun, Turkey). Hittite J. Sci. Eng. 2017, 4, 39–44. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Kiyashko, S.I.; Titlyanova, T.V.; Yakovleva, I.M.; Li, X.B.; Huang, H. Nitrogen sources to macroalgal growth in Sanya Bay (Hainan Island, China). Curr. Dev. Oceanogr. 2011, 2, 65–84. [Google Scholar]
- Titlyanov, E.A.; Titlyanova, T.V.; Huang, H.; Li, X. Seasonal changes in benthic algal communities of the upper subtidal zone in Sanya Bay (Hainan Island, China). J. Mar. Biol. Assoc. UK 2013, 94, 51–64. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Li, X.B.; Hansen, G.I.; Huang, H. Seasonal changes in the intertidal algal communities of Sanya Bay (Hainan Island, China). J. Mar. Biol. Assoc. UK 2014, 94, 879–893. [Google Scholar] [CrossRef]
- Huang, H.; Li, X.B.; Titlyanov, E.A.; Ye, C.; Titlyanova, T.V.; Guo, P.Y.; Zhang, J. Linking macroalgal δ15 N-values to nitrogen sources and effects of nutrient stress on coral condition in an upwelling region. Bot. Mar. 2013, 56, 471–480. [Google Scholar] [CrossRef]
- Li, X.B.; Titlyanov, E.A.; Zhang, J.; Titlyanova, T.V.; Zhang, G.; Huang, H. Macroalgal assemblage changes on coral reefs along a natural gradient from fish farms in southern Hainan Island Aquatic. Ecosyst. Health Manag. 2016, 19, 74–82. [Google Scholar] [CrossRef]
- Morand, P.; Briand, X. Excessive growth of macroalgae: A symptom of environmental disturbance. Bot. Mar. 1996, 39, 491–516. [Google Scholar] [CrossRef]
- Morand, P.; Merceron, M. Coastal eutrophication and excessive growth of macroalgae. Recent Res. Dev. Environ. Biol. 2004, 1, 395–449. [Google Scholar]
- Lapointe, B.E.; Barile, P.J.; Littler, M.M.; Littler, D.S.; Bedford, B.J.; Gasque, C. Macroalgal blooms on southeast Florida coral reefs: I. Nutrient stoichiometry of the invasive green alga Codium isthmocladum in the wider Caribbean indicates nutrient enrichment. Harmful Algae 2005, 4, 1092–1105. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Barile, P.J.; Littler, M.M.; Littler, D.S. Macroalgal blooms on southeast Florida coral reefs: II. Cross-shelf discrimination of nitrogen sources indicates widespread assimilation of sewage nitrogen. Harmful Algae 2005, 4, 1106–1122. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Li, X.B.; Huang, H. An inventory of marine benthic macroalgae of Hainan Island, China. Russ. J. Mar. Biol. 2018, 44, 175–184. [Google Scholar] [CrossRef]
- Furnas, M.; Mitchell, A.; Skuza, M. Shelf-scale nitrogen and phosphorus budgets for the central Great Barrier Reef (16°−19° S). In Proceedings of the 8th International Coral Reef Symposium, Balboa, Panama, 24–29 June 1997; Volume 1, pp. 809–814. [Google Scholar]
- Charpy, L.; Charpy-Roubaud, C.; Buat, P. Excess primary production, calcification and nutrient fluxes of a patch reef (Tikehau atoll, French Polynesia). Mar. Ecol. Prog. Ser. 1998, 173, 139–147. [Google Scholar] [CrossRef]
- Charpy-Roubaud, C.J.; Charpy, L. Nutrients, particulate organic matter, and planktonic and benthic production of the Tikehau Atoll (Tuamotu Archipelago French Polynesia). Atoll Res. Bull. 1994, 415–425, 1–30. [Google Scholar] [CrossRef]
- Briggs, J.C. Marine Zoogeography; McGraw-Hill: New York, NY, USA, 1974; 475p. [Google Scholar]
- Lüning, K. Seaweeds: Their Environment, Biogeography and Ecophysiology; John Wiley & Sons, Inc.: New York, NY, USA; Chichester, UK; Brisbane, Australia; Toronto, ON, Canada; Singapore, 1990; 527p. [Google Scholar]
- Li, X.B. Identification of Major Factors Influencing the Composition, Spatial and Temporal Variation of Scleractinian Coral Community in Sanya, China. Ph.D. Thesis, Chinese Academy of Sciences, Beijing, China, 2011; pp. 1–107. (In Chinese). [Google Scholar]
- Hutchings, P.A.; Wu, B.L. Coral reefs of Hainan Island, South China Sea. Mar. Pollut. Bull. 1987, 18, 25–26. [Google Scholar] [CrossRef]
- Fiege, D.; Neumann, V.; Jinhe, L. Observations on coral reefs of Hainan Island, South China Sea. Mar. Pollut. Bull. 1994, 29, 84–89. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, X.Z.; Long, X.M. A numerical study on internal tides in the northeast of the South China Sea. J. Trop. Oceanol. 1996, 14, 15–23. [Google Scholar]
- Hodgson, G.; Yau, E.P.M. Physical and biological controls of coral communities in Hong Kong. In Proceedings of the 8th International Coral Reef Symposium; Lessios, H.A., Macintyre, I.G., Eds.; Tropical Research Institute: Balboa, Panama, 1997; Volume 1, pp. 459–464. [Google Scholar]
- Zhang, G.; Que, H.; Liu, X.; Xu, H. Abalone mariculture in China. J. Shellfish Res. 2004, 23, 947–950. [Google Scholar]
- Tadashi, K.; Dai, C.F.; Park, H.-S.; Huang, H.; Ang, P.O. Status of coral reefs in East and North Asia (China, Hong Kong, Taiwan, South Korea and Japan. In Status of Coral Reefs of the World; Wilkinson, C., Ed.; Global Coral Reef Monitoring: Townsville, Australia, 2008; pp. 145–158. [Google Scholar]
- Tseng, C.K. Common Seaweeds of China; Science Press Book: Beijing, China, 1983; 316p. [Google Scholar]
- Tseng, C.K. The past, present and future of phycology in China. Hydrobiologia 2004, 512, 11–20. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Xia, B.M.; Bartsch, I. Checklist of marine benthic green algae (Chlorophyta) on Hainan, a subtropical island off the coast of China: Comparisons between the 1930s and 1990–2009 reveal environmental changes. Bot. Mar. 2011, 54, 523–535. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Belous, O.S.; Kalita, T.L. Inventory change (1990s–2010s) in the marine flora of Sanya Bay (Hainan Island, China). J. Mar. Biol. Assoc. UK 2015, 95, 461–470. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Li, X.; Huang, H. Recent (2008–2012) seaweed flora of Hainan Island, South China Sea. Mar. Biol. Res. 2015, 11, 540–550. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Xia, B.M.; Bartsch, I. Retrospective analysis of diversity and species composition of marine macroalgae of Hainan Island (China). Ocean Sci. 2016, 51, 485–506. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Li, X.B.; Huang, H. Coral Reef Marine Plants of Hainan/Island; Academic Press: Amsterdam, The Netherlands, 2016; p. 243. [Google Scholar]
- Titlyanova, T.V.; Titlyanov, E.A.; Xia, B.; Bartsch, I. New records of benthic marine green algae (Chlorophyta) for the island of Hainan, China. Nova Hedwig. 2012, 94, 441–470. [Google Scholar] [CrossRef] [PubMed]
- Titlyanova, T.V.; Titlyanov, E.A.; Kalita, T.L. Marine algal flora of Hainan Island: A comprehensive synthesis. Coast. Ecosyst. 2014, 1, 28–53. [Google Scholar]
- Titlyanov, E.A.; Titlyanova, T.V. Changes in the species composition of benthic macroalgal communities of the upper subtidal zone on a coral reef in Sanya Bay (Hainan Island, China) during 2009–2012. Russ. J. Mar. Biol. 2013, 39, 413–419. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. National University of Ireland: Galeay, Ireland, 2018. Available online: http://www.algaebase.org (accessed on 10 December 2018).
- Saunders, G.W.; Hommersand, M.H. Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. Am. J. Bot. 2004, 91, 1494–1507. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, R.T. Checklist and bibliography of the marine benthic algae from the Mariana Islands (Guam and CNMI). Univ. Guam Mar. Lab. Technol. Rep. 2003, 107, 1–49. [Google Scholar]
- Tsuda, R.T. Checklist and bibliography of the marine benthic algae within Chuuk, Pohnpei, and Kosrae States, Federated States of Micronesia. Univ. Guam Mar. Lab. Technol. Rep. 2006, 34, 1–35. [Google Scholar]
- Clarke, K.R.; Green, R.H. Statistical design and analysis for a ‘biological effects’ study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v5: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Nabivailo, Y.V.; Titlyanov, E.A. Competitive relationships in natural and artificial algal communities. Russ. J. Mar. Biol. 2006, 32, 21–31. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Location | Sampling Transects | Number of Samplings | Number of Found and Analyzed Algal Communities | Number of Samples | Number of Species |
|---|---|---|---|---|---|
| In front of the outlet, heavily polluted | 2 | 2 | 22 | 66 | 85 |
| In front of the Marine Biological Station, moderately polluted | 2 | 2 | 27 | 81 | 103 |
| Total analyzed and found | 4 | 4 | 49 | 147 | 109 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Titlyanov, E.A.; Titlyanova, T.V.; Huang, H.; Scriptsova, A.V.; Xu, H.; Li, X. Seasonal Changes in the Intertidal and Subtidal Algal Communities of Extremely and Moderately Polluted Coastal Regions of Sanya Bay (Hainan Island, China). J. Mar. Sci. Eng. 2019, 7, 93. https://doi.org/10.3390/jmse7040093
Titlyanov EA, Titlyanova TV, Huang H, Scriptsova AV, Xu H, Li X. Seasonal Changes in the Intertidal and Subtidal Algal Communities of Extremely and Moderately Polluted Coastal Regions of Sanya Bay (Hainan Island, China). Journal of Marine Science and Engineering. 2019; 7(4):93. https://doi.org/10.3390/jmse7040093
Chicago/Turabian StyleTitlyanov, Eduard A., Tamara V. Titlyanova, Hui Huang, Anna V. Scriptsova, Huili Xu, and Xiubao Li. 2019. "Seasonal Changes in the Intertidal and Subtidal Algal Communities of Extremely and Moderately Polluted Coastal Regions of Sanya Bay (Hainan Island, China)" Journal of Marine Science and Engineering 7, no. 4: 93. https://doi.org/10.3390/jmse7040093
APA StyleTitlyanov, E. A., Titlyanova, T. V., Huang, H., Scriptsova, A. V., Xu, H., & Li, X. (2019). Seasonal Changes in the Intertidal and Subtidal Algal Communities of Extremely and Moderately Polluted Coastal Regions of Sanya Bay (Hainan Island, China). Journal of Marine Science and Engineering, 7(4), 93. https://doi.org/10.3390/jmse7040093