


Sea Surface Temperature Fronts and North Atlantic Right Whale Sightings in the Western Gulf of St. Lawrence


Abstract
1. Introduction
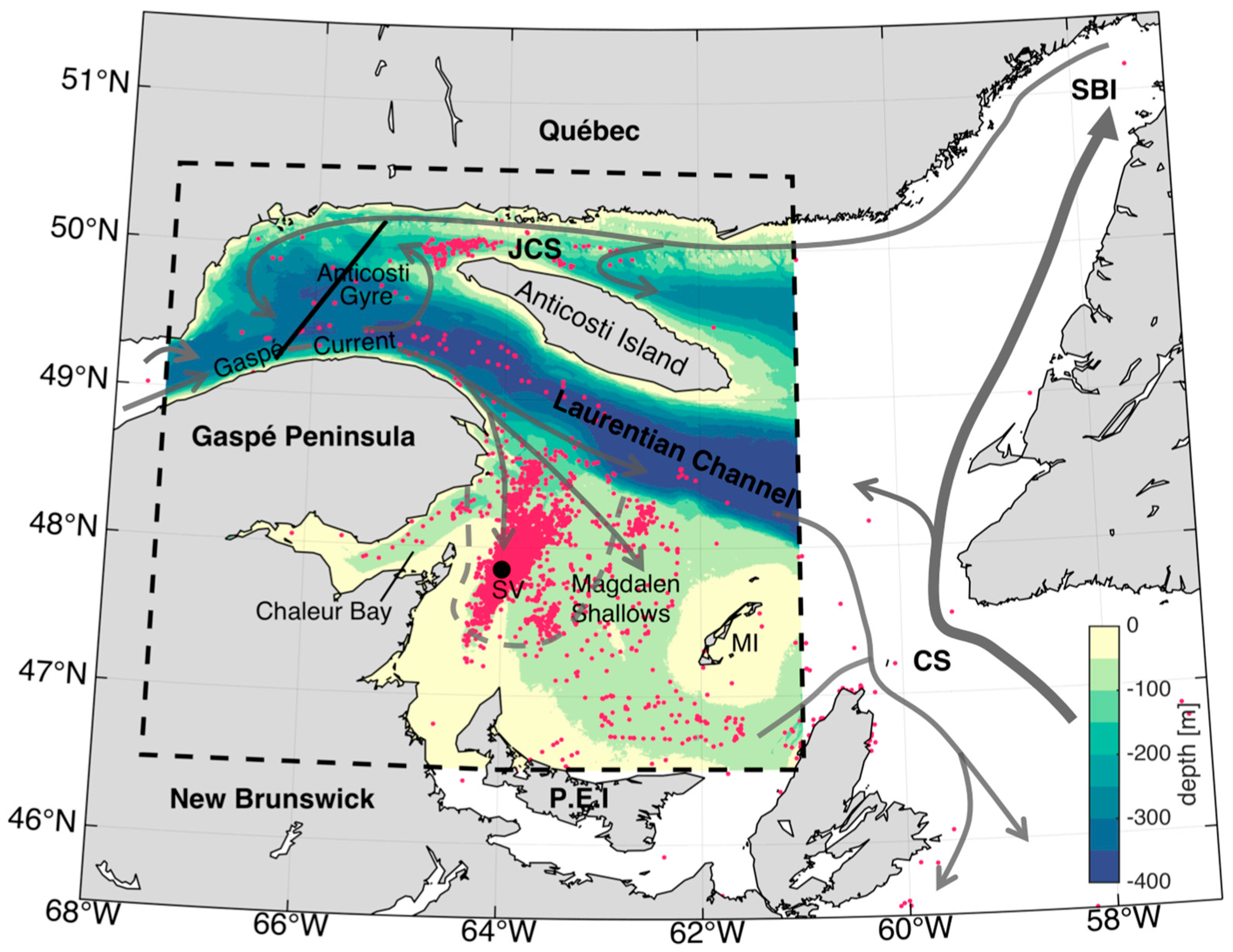
1.1. GSL Circulation
1.2. The North Atlantic Right Whale in the sGSL
2. Materials and Methods
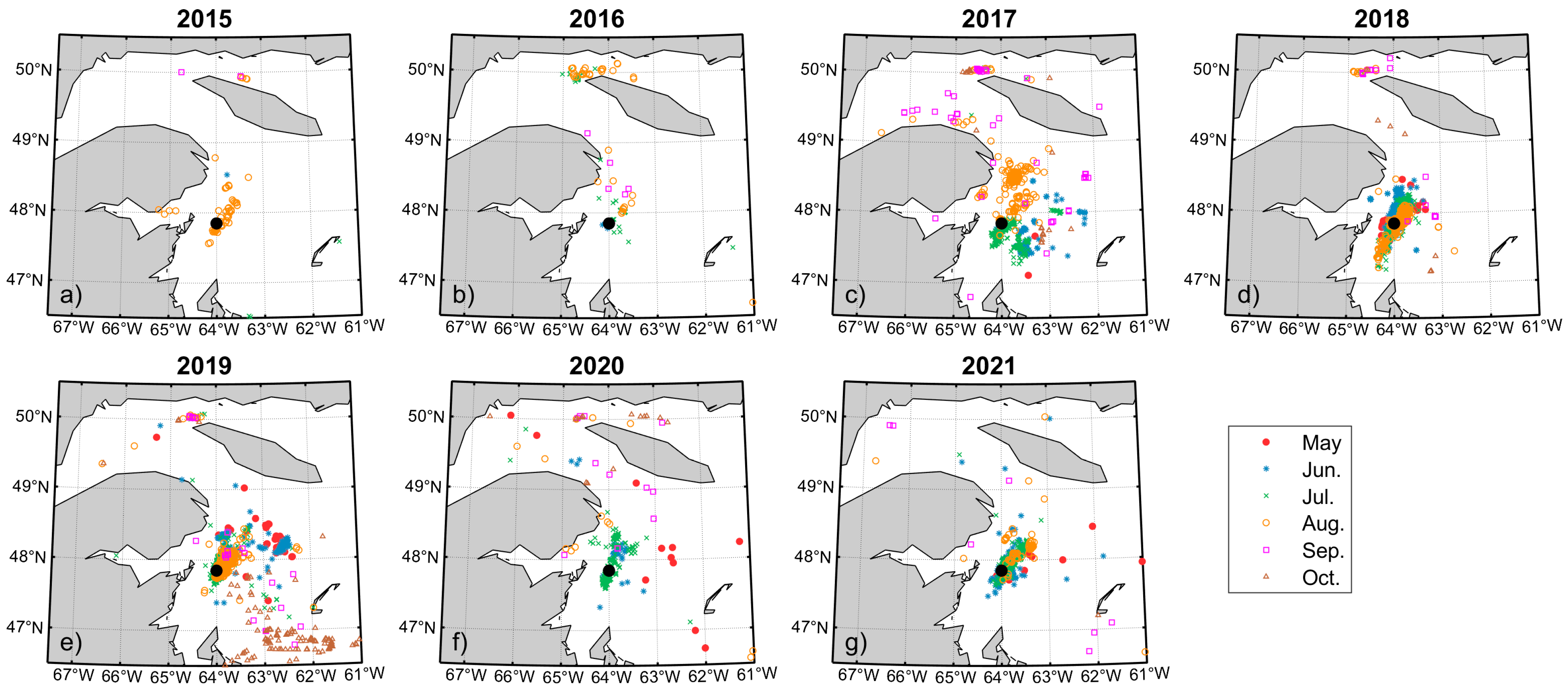
2.1. NARW Sightings
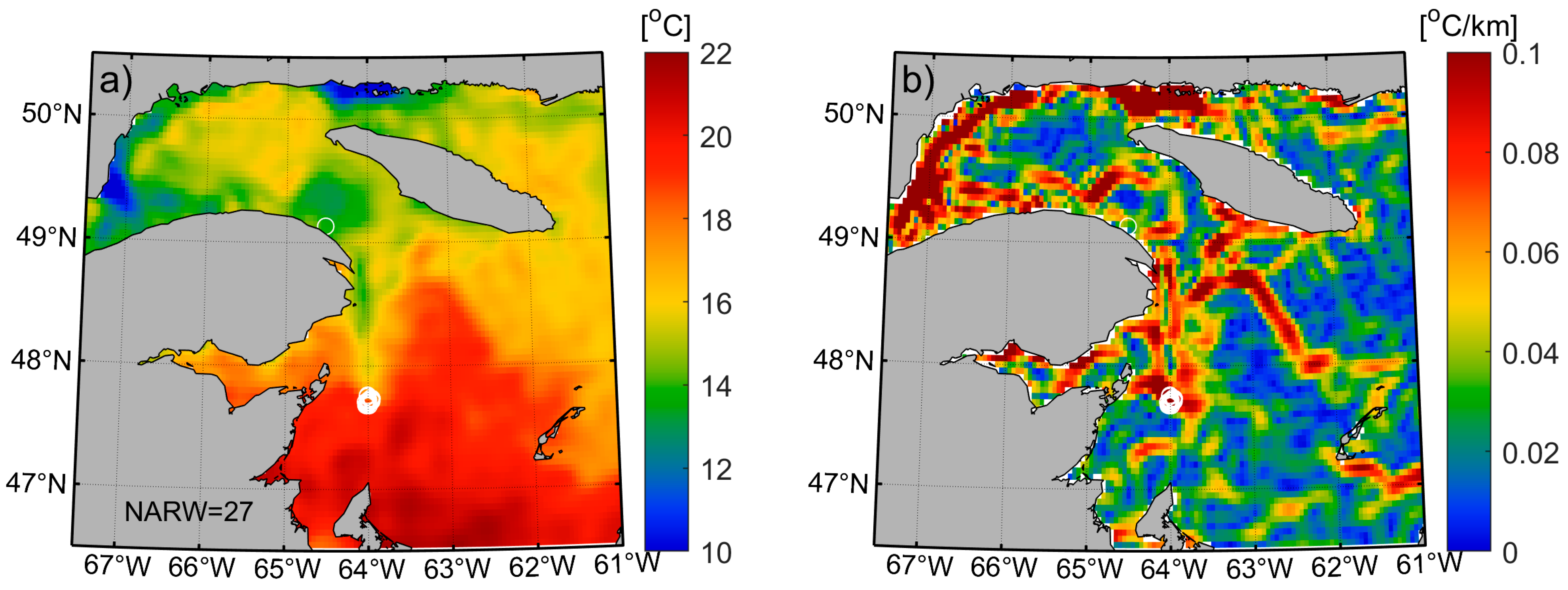
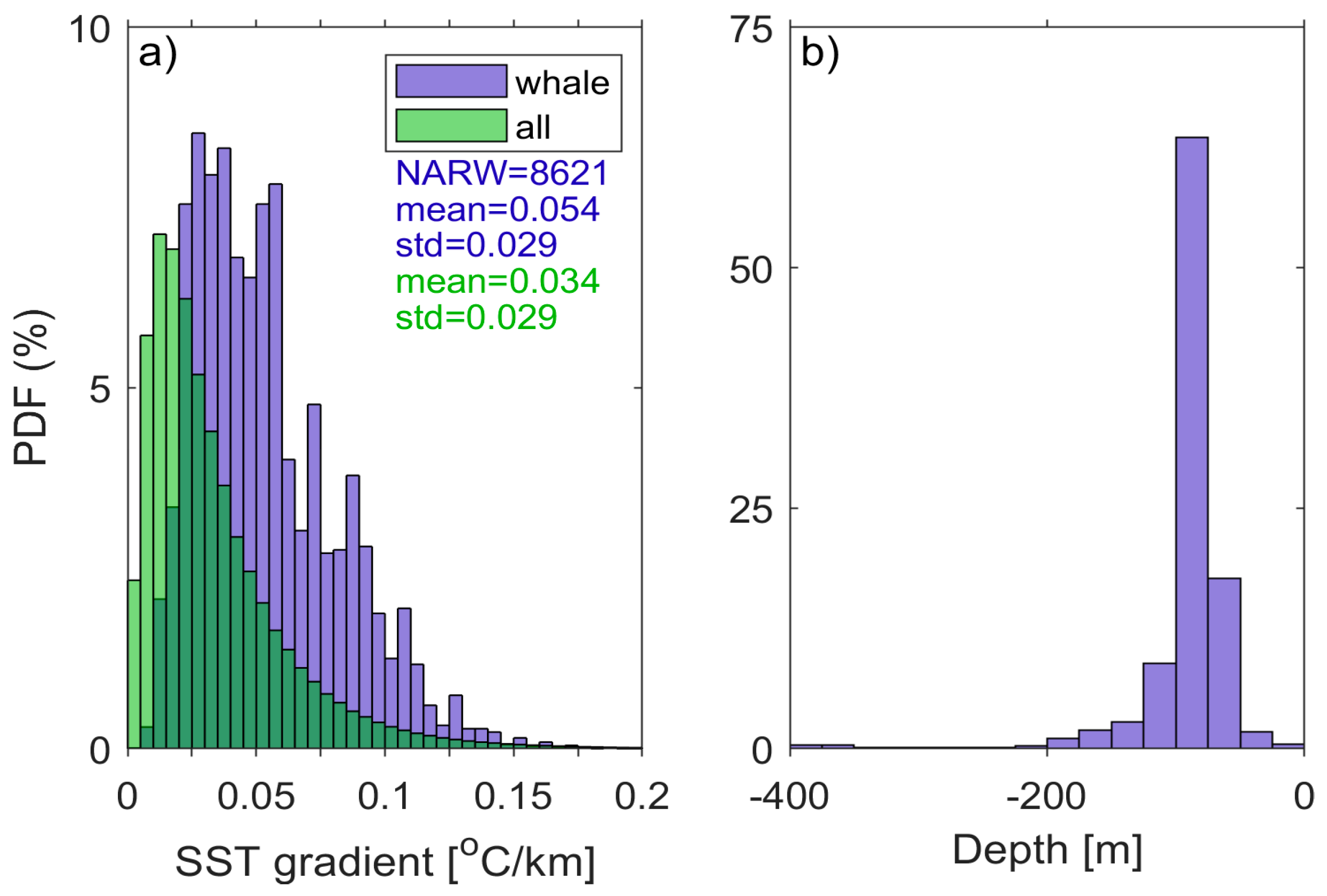
2.2. Satellite SST and Gradients
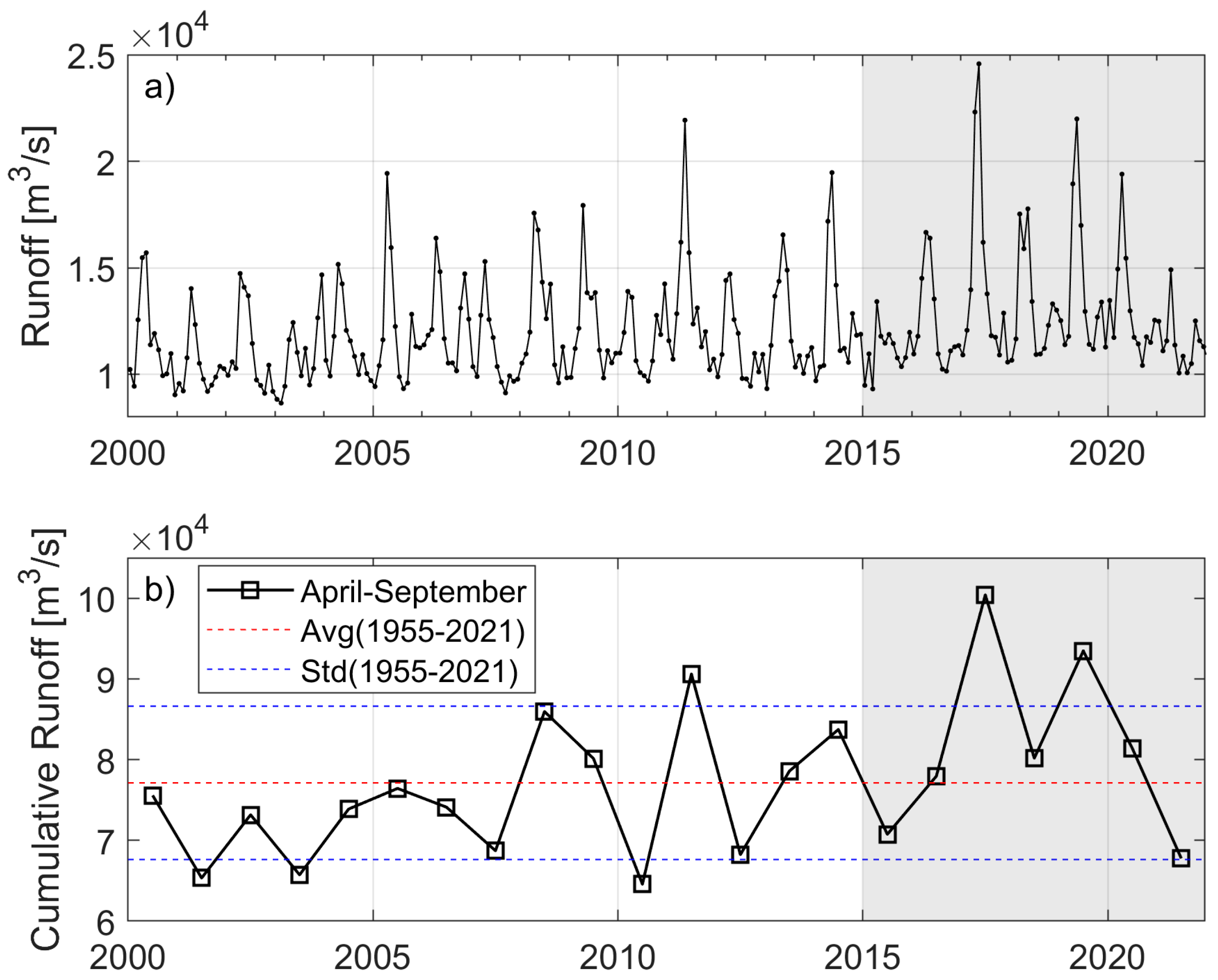
2.3. Freshwater Discharge and Bathymetry Data
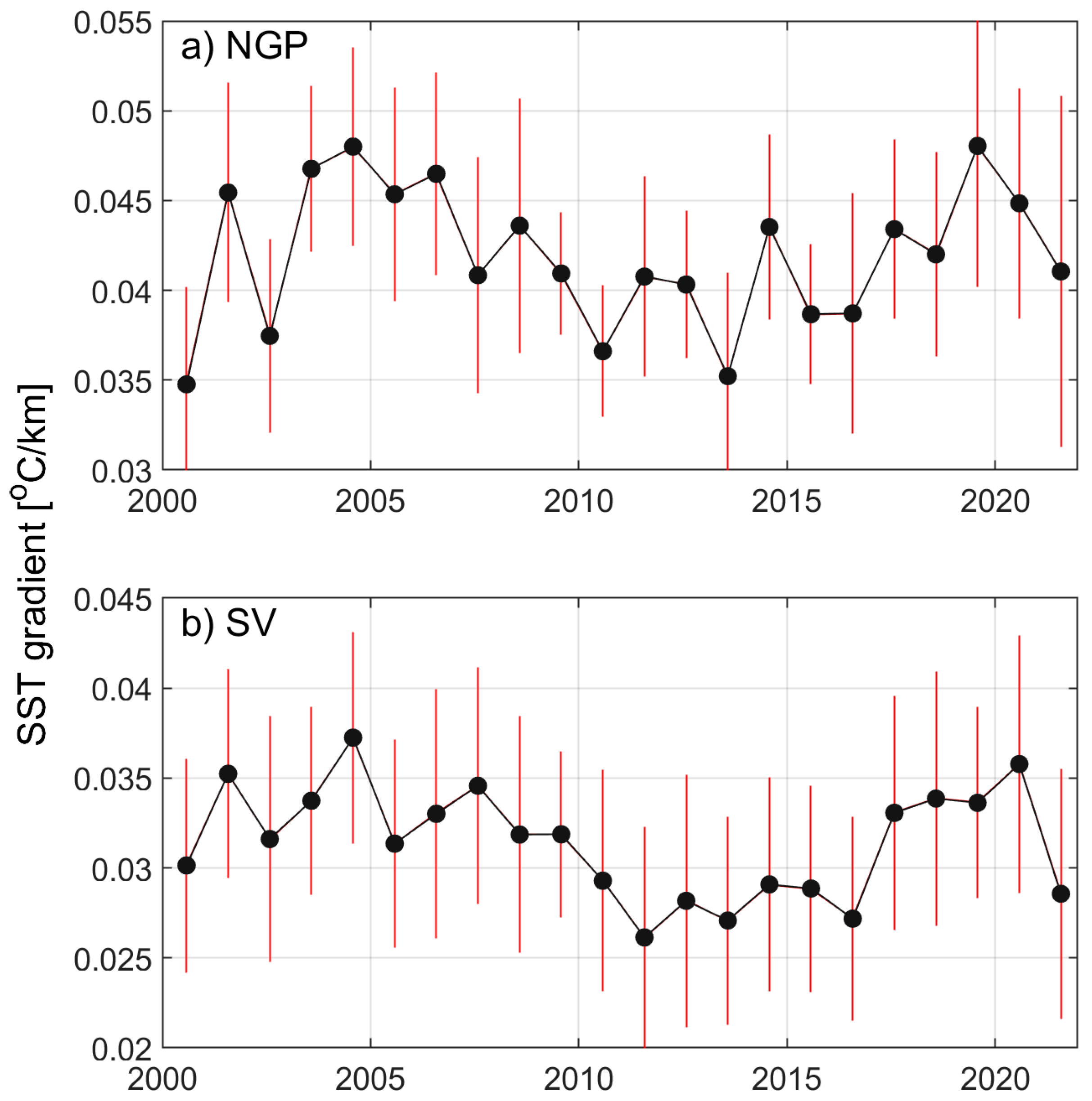
2.4. Altimetric Index of Gaspé Current Intensity
3. Results
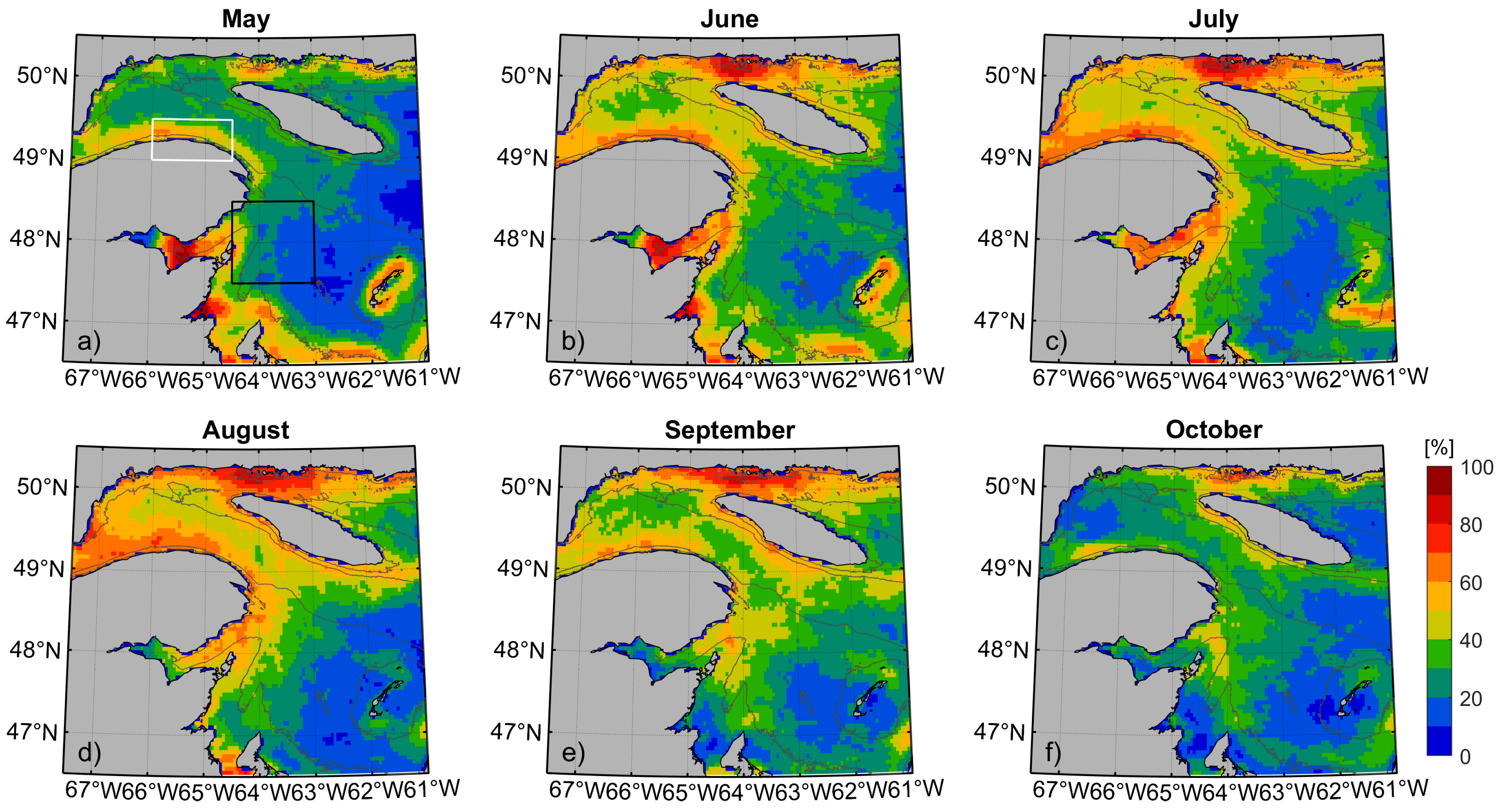
3.1. Variability of Front Occurrence Probability Patterns
3.2. Monthly Probability Pattern and Whale Sightings
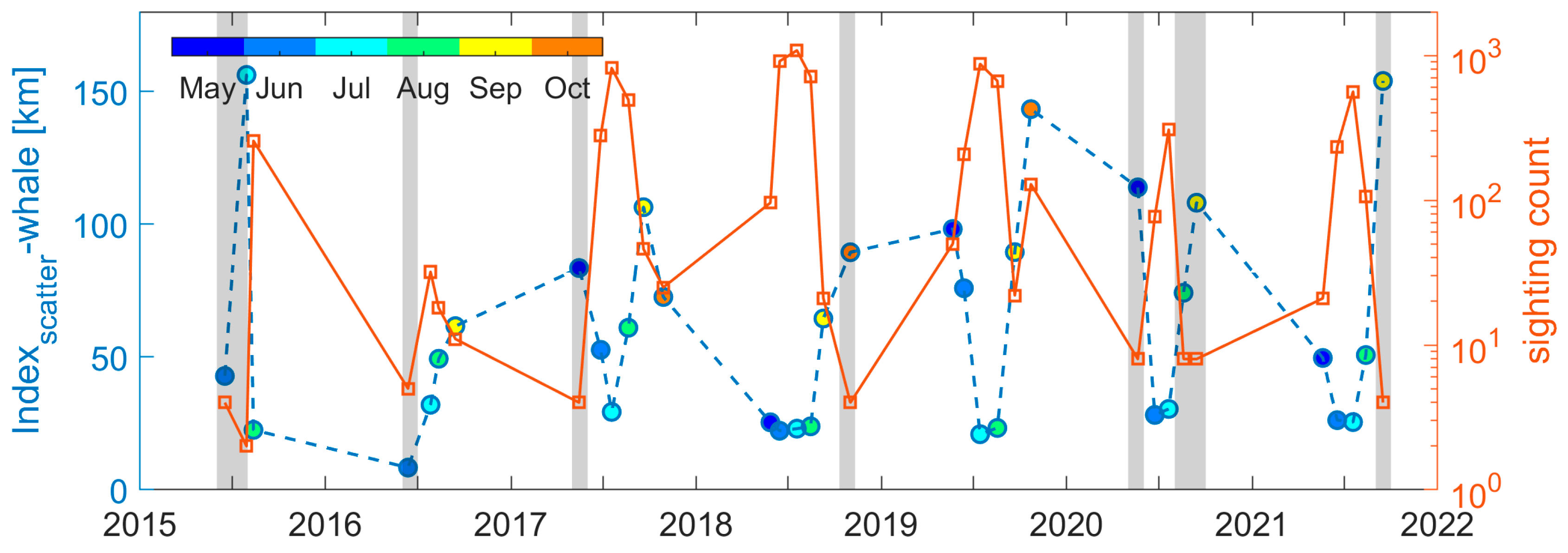
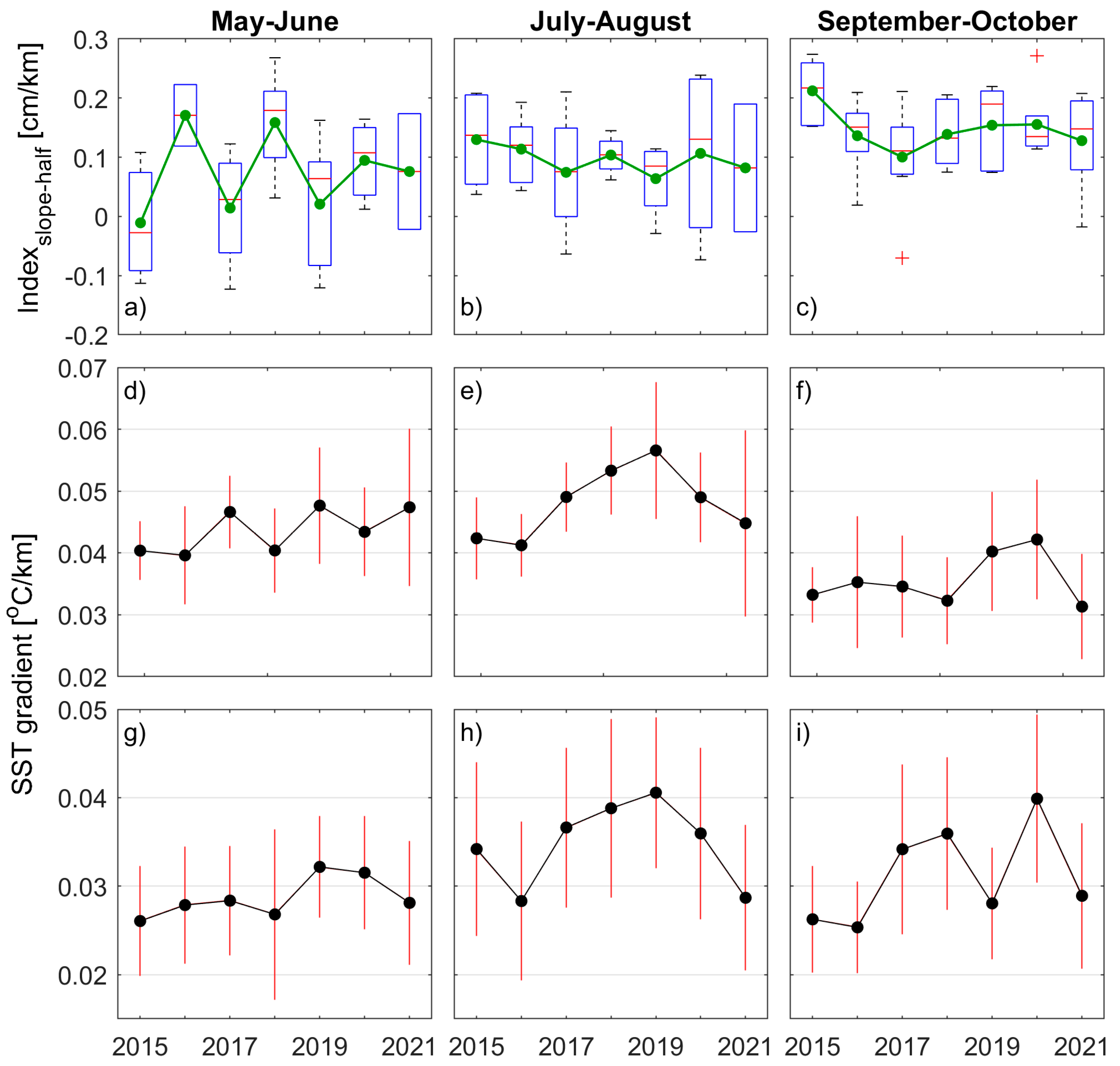
3.3. Variations in the Gaspé Current Indices
4. Discussion
4.1. Probability Patterns of Whale Occurrence
4.2. Linkage Between Indices and NARWs Foraging Habitat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daoust, P.Y.; Couture, E.L.; Wimmer, T.; Bourque, L. Incident Report: North Atlantic Right Whale Mortality Event in the Gulf of St. Lawrence. In Canadian Wildlife Health Cooperative, Marine Animal Response Society, and Fisheries and Oceans Canada; Fisheries and Oceans Canada (DFO): Ottawa, ON, Canada, 2017. [Google Scholar]
- Sorochan, K.A.; Plourde, S.; Baumgartner, M.F.; Johnson, C.L. Availability, Supply, and Aggregation of Prey (Calanus spp.) in Foraging Areas of the North Atlantic Right Whale (Eubalaena glacialis). ICES J. Mar. Sci. 2021, 78, 3498–3520. [Google Scholar] [CrossRef]
- Cyr, F.; Galbraith, P.S. A Climate Index for the Newfoundland and Labrador Shelf. Earth Syst. Sci. Data Discuss. 2020, 13, 1807–1828. [Google Scholar] [CrossRef]
- Boyce, D.G.; Tittensor, D.P.; Garilao, C.; Henson, S.; Kaschner, K.; Kesner-Reyes, K.; Pigot, A.; Reyes, R.B.; Reygondeau, G.; Schleit, K.E.; et al. A Climate Risk Index for Marine Life. Nat. Clim. Change 2022, 12, 854–862. [Google Scholar] [CrossRef]
- Cyr, F.; Lewis, K.; Bélanger, D.; Regular, P.; Clay, S.; Devred, E. Physical Controls and Ecological Implications of the Timing of the Spring Phytoplankton Bloom on the Newfoundland and Labrador Shelf. Limnol. Oceanogr. Lett. 2024, 9, 191–198. [Google Scholar] [CrossRef]
- Tao, J.; Shen, H.; Danielson, R.E.; Perrie, W. Characterizing the Variability of a Physical Driver of North Atlantic Right Whale Foraging Habitat Using Altimetric Indices. J. Mar. Sci. Eng. 2023, 11, 1760. [Google Scholar] [CrossRef]
- Danielson, R.E.; Shen, H.; Tao, J.; Perrie, W.A. Dependence of of ocean surface filaments on wind speed: An observational study of North Atlantic right whale habitat. Remote Sens. Environ. 2023, 287, 113494. [Google Scholar] [CrossRef]
- Franks, P.J.S. Sink or Swim: Accumulation of Biomass at Fronts. Mar. Ecol. Prog. Ser. 1992, 82, 1–12. [Google Scholar] [CrossRef]
- Pineda, J. Circulation and Larval Distribution in Internal Tidal Bore Fronts. Limnol. Oceanogr. 1999, 44, 1400–1414. [Google Scholar] [CrossRef]
- Epstein, A.W.; Beardsley, R.C. Flow-Induced Aggregation of Plankton at a Front: A 2-D Eulerian Model Study. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 395–418. [Google Scholar] [CrossRef]
- Prairie, J.C.; Sutherland, K.R.; Nickols, K.J.; Kaltenberg, A.M. Biophysical Interactions in the Plankton: A Cross-Scale Review. Limnol. Oceanogr. Fluids Environ. 2012, 2, 121–145. [Google Scholar] [CrossRef]
- Belkin, I.M. Remote Sensing of Ocean Fronts in Marine Ecology and Fisheries. Remote Sens. 2021, 13, 883. [Google Scholar] [CrossRef]
- Owen, R.W. Fronts and Eddies in the Sea: Mechanisms, Interactions and Biological Effects. In Analysis of Marine Ecosystems; Academic Press: New York, NY, USA, 1981; pp. 197–233. [Google Scholar]
- Yoder, J.A.; Ackleson, S.G.; Barber, R.T.; Flament, P.; Balch, W.M. A Line in the Sea. Nature 1994, 371, 689–692. [Google Scholar] [CrossRef]
- Genin, A.; Jaffe, J.S.; Reef, R.; Richter, C.; Franks, P.J. Swimming against the Flow: A Mechanism of Zooplankton Aggregation. Science 2005, 308, 860–862. [Google Scholar] [CrossRef] [PubMed]
- Belkin, I.M.; Cornillon, P.C.; Sherman, K. Fronts in Large Marine Ecosystems. Prog. Oceanogr. 2009, 81, 223–236. [Google Scholar] [CrossRef]
- Cyr, F.; Larouche, P. Thermal fronts atlas of Canadian coastal waters. Atmosphere-Ocean 2015, 53, 212–236. [Google Scholar] [CrossRef]
- Olson, D.B.; Backus, R.H. The Concentrating of Organisms at Fronts: A Cold-Water Fish and a Warm-Core Gulf Stream Ring. J. Mar. Res. 1985, 43, 113–137. [Google Scholar] [CrossRef]
- Chambault, P.; Roquet, F.; Benhamou, S.; Baudena, A.; Pauthenet, E.; Thoisy, B.; Bonola, M.; Dos Reis, V.; Crasson, R.; Brucker, M.; et al. The Gulf Stream Frontal System: A Key Oceanographic Feature in the Habitat Selection of the Leatherback Turtle? Deep Sea Res. Part I Oceanogr. Res. Pap. 2017, 123, 35–47. [Google Scholar] [CrossRef]
- Ryan, J.P.; Benoit-Bird, K.J.; Oestreich, W.K.; Leary, P.; Smith, K.B.; Waluk, C.M.; Cade, D.E. Oceanic Giants Dance to Atmospheric Rhythms: Ephemeral Wind-driven Resource Tracking by Blue Whales. Ecol. Lett. 2022, 25, 2435–2447. [Google Scholar] [CrossRef]
- Hofmann, E.E.; Powell, T.M. Environmental Variability Effects on Marine Fisheries: Four Case Histories. Ecol. Appl. 1998, 8, S23–S32. [Google Scholar] [CrossRef]
- Brennan, C.E.; Maps, F.; Gentleman, W.C.; Lavoie, D.; Chassé, J.; Plourde, S.; Johnson, C.L. Ocean Circulation Changes Drive Shifts in Calanus Abundance in North Atlantic Right Whale Foraging Habitat: A Model Comparison of Cool and Warm Year Scenarios. Prog. Oceanogr. 2021, 197, 102629. [Google Scholar] [CrossRef]
- Abo-Taleb, H. Importance of Plankton to Fish Community. Biol. Res. Aquat. Sci. 2019, 83, 83–89. [Google Scholar]
- Marshall, S.M.; Orr, A.P. The Biology of a Marine Copepod: Calanus Finmarchicus (Gunnerus); Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Wishner, K.; Durbin, E.; Durbin, A.; Macaulay, M.; Winn, H.; Kenney, R. Copepod Patches and Right Whales in the Great South Channel off New England. Bull. Mar. Sci. 1988, 43, 825–844. [Google Scholar]
- Doniol-Valcroze, T.; Berteaux, D.; Larouche, P.; Sears, R. Influence of Thermal Fronts on Habitat Selection by Four Rorqual Whale Species in the Gulf of St. Lawrence. Mar. Ecol. Prog. Ser. 2007, 335, 207–216. [Google Scholar] [CrossRef]
- Hamazaki, T. Spatiotemporal prediction models of cetacean habitats in the mid-western North Atlantic Ocean (from Cape Hatteras, North Carolina, USA to Nova Scotia, Canada. Mar. Mammal Sci. 2002, 18, 920–939. [Google Scholar] [CrossRef]
- Baumgartner, M.F.; Cole, T.V.; Clapham, P.J.; Mate, B.R. North Atlantic Right Whale Habitat in the Lower Bay of Fundy and on the SW Scotian Shelf during 1999–2001. Mar. Ecol. Prog. Ser. 2003, 264, 137–154. [Google Scholar] [CrossRef]
- Baumgartner, M.F.; Mate, B.R. Summer and Fall Habitat of North Atlantic Right Whales (Eubalaena glacialis) Inferred from Satellite Telemetry. Can. J. Fish. Aquat. Sci. 2005, 62, 527–543. [Google Scholar] [CrossRef]
- Castelao, R.M.; Wang, Y. Wind-driven Variability in Sea Surface Temperature Front Distribution in the California Current System. J. Geophys. Res. Oceans 2014, 119, 1861–1875. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, R.; Yu, Y.; Ji, F. Variability in the sea surface temperature gradient and its impacts on chlorophyll-a concentration in the Kuroshio extension. Remote Sens. 2021, 13, 888. [Google Scholar] [CrossRef]
- Saldías, G.S.; Hernández, W.; Lara, C.; Muñoz, R.; Rojas, C.; Vásquez, S.; Pérez-Santos, I.; Soto-Mardones, L. Seasonal Variability of SST Fronts in the Inner Sea of Chiloé and Its Adjacent Coastal Ocean, Northern Patagonia. Remote Sens. 2021, 13, 181. [Google Scholar] [CrossRef]
- Gaskin, D.E. Updated Status of the Right Whale, Eubalaena glacialis, in Canada. Can. Field-Naturalist. 1987, 101, 295–309. [Google Scholar] [CrossRef]
- Etnoyer, P.; Canny, D.; Mate, B.R.; Morgan, L.E.; Ortega-Ortiz, J.G.; Nichols, W.J. Sea-Surface Temperature Gradients across Blue Whale and Sea Turtle Foraging Trajectories off the Baja California Peninsula, Mexico. Deep Sea Res. Part II Top. Stud. Oceanogr. 2006, 53, 340–358. [Google Scholar] [CrossRef]
- Runge, J.A.; Castonguay, M.; Lafontaine, Y.; Ringuette, M.; Beaulieu, J.L. Covariation in Climate, Zooplankton, Biomass and Mackerel Recruitment in the Southern Gulf of St. Lawrence. Fish. Oceanogr. 1999, 8, 139–149. [Google Scholar] [CrossRef]
- Koutitonsky, V.G.; Bugden, G.L. The Physical Oceanography of the Gulf of St. Lawrence: A Review with Emphasis on the Synoptic Variability of the Motion, p.57-90. The Gulf of St. Lawrence: Small ocean or big estuary? Can. Spec. Publ. Fish. Aquat. Sci. 1991, 113, 57–90. [Google Scholar]
- Han, G. Sea Level and Surface Current Variability in the Gulf of St Lawrence from Satellite Altimetry. Int. J. Remote Sens. 2004, 25, 5069–5088. [Google Scholar] [CrossRef]
- Benoit, J.; El-Sabh, M.I.; Tang, C.L. Structure and Seasonal Characteristics of the Gaspé Current. J. Geophys. Res. Oceans 1985, 90, 3225–3236. [Google Scholar] [CrossRef]
- El-Sabh, M.I. Surface Circulation Pattern in the Gulf of St. Lawrence. J. Fish. Board. Can. 1976, 33, 124–138. [Google Scholar] [CrossRef]
- Sheng, J. Dynamics of a Buoyancy-Driven Coastal Jet: The Gaspé Current. J. Phys. Oceanogr. 2001, 31, 3146–3162. [Google Scholar] [CrossRef]
- Gan, J.; Ingram, R.G.; Greatbatch, R.J. On the Unsteady Separation/Intrusion of the Gaspé Current and Variability in Baie Des Chaleurs: Modeling Studies. J. Geophys. Res. 1997, 102, 15567–15581. [Google Scholar] [CrossRef]
- Lavoie, D.; Chassé, J.; Simard, Y.; Lambert, N.; Galbraith, P.S.; Roy, N.; Brickman, D. Large-Scale Atmospheric and Oceanic Control on Krill Transport into the St. Lawrence Estuary Evidenced with Three-Dimensional Numerical Modelling. Atmosphere-Ocean 2016, 54, 299–325. [Google Scholar] [CrossRef]
- DFO. Updated Information on the Distribution of North Atlantic Right Whale in Canadian Waters. In DFO Canadian Science Advisory Secretariat Science Advisory Report; DFO: Ottawa, ON, Canada, 2020. [Google Scholar]
- Simard, Y.; Roy, N.; Giard, S.; Aulanier, F. North Atlantic Right Whale Shift to the Gulf of St. Lawrence in 2015, Revealed by Long-Term Acoustics. Endanger. Species Res. 2019, 40, 271–284. [Google Scholar] [CrossRef]
- Lehoux, C.; Plourde, S.; Lesage, V. Significance of Dominant Zooplankton Species to the North Atlantic Right Whale Potential Foraging Habitats in the Gulf of St. Lawrence: A Bio-Energetic Approach; Government of Canada Publications: Ottawa, ON, Canada, 2020; p. 44.
- Gavrilchuk, K.; Lesage, V.; Fortune, S.; Trites, A.W.; Plourde, S. A Mechanistic Approach to Predicting Suitable Foraging Habitat for Reproductively Mature North Atlantic Right Whales in the Gulf of St. Lawrence; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2020; Volume 34.
- Blais, M.; Galbraith, P.S.; Plourde, S.; Devine, L.; Lehoux, C. Chemical and Biological Oceanographic Conditions in the Estuary and Gulf of St. Lawrence During 2019; DFO Canadian Science Advisory Secretariat Science Advisory Report; DFO: Ottawa, ON, Canada, 2021; p. 66.
- Maps, F.; Zakardjian, B.A.; Plourde, S.; Saucier, F.J. Modeling the Interactions between the Seasonal and Diel Vertical Migration Behaviours of Calanus Finmarchicus and the Circulation in the Gulf of St. Lawrence (Canada). J. Mar. Syst. 2011, 88, 183–202. [Google Scholar] [CrossRef]
- Benkort, D.; Lavoie, D.; Plourde, S.; Dufresne, C.; Maps, F. Arctic and Nordic Krill Circuits of Production Revealed by the Interactions between Their Physiology, Swimming Behaviour and Circulation. Prog. Oceanogr. 2020, 182, 102270. [Google Scholar] [CrossRef]
- Brennan, C.E.; Maps, F.; Gentleman, W.C.; Plourde, S.; Lavoie, D.; Chassé, J.; Lehoux, C.; Krumhansl, K.A.; Johnson, C.L. How Transport Shapes Copepod Distributions in Relation to Whale Feeding Habitat: Demonstration of a New Modelling Framework. Prog. Oceanogr. 2019, 171, 1–21. [Google Scholar] [CrossRef]
- Plourde, S.; Runge, J.A. Reproduction of the Planktonic Copepod Calanus Finmarchicus in the Lower St. Lawrence Estuary: Relation to the Cycle of Phytoplankton Production and Evidence for a Calanus Pump. Mar. Ecol. Prog. Ser. 1993, 102, 217–227. [Google Scholar] [CrossRef]
- Plourde, S.; Joly, P.; Runge, J.A.; Zakardjian, B.; Dodson, J.J. Life Cycle of Calanus Finmarchicus in the Lower St. Lawrence Estuary: The Imprint of Circulation and Late Timing of the Spring Phytoplankton Bloom. Can. J. Fish. Aquat. 2001, 58, 647–658. [Google Scholar] [CrossRef]
- Mayo, C.A.; Marx, M.K. Surface Foraging Behaviour of the North Atlantic Right Whale, Eubalaena glacialis, and Associated Zooplankton Characteristics. Can. J. Zool. 1990, 68, 2214–2220. [Google Scholar] [CrossRef]
- Plourde, S.; Lehoux, C.; Johnson, C.L.; Perrin, G.; Lesage, V. North Atlantic Right Whale (Eubalaena glacialis) and Its Food: (I) a Spatial Climatology of Calanus Biomass and Potential Foraging Habitats in Canadian Waters. J. Plankton 2019, 41, 667–685. [Google Scholar] [CrossRef]
- Baumgartner, M.F.; Wenzel, F.W.; Lysiak, N.S.J.; Patrician, M.R. North Atlantic Right Whale Foraging Ecology and Its Role in Human-Caused Mortality. Mar. Ecol. Prog. Ser. 2017, 581, 165–181. [Google Scholar] [CrossRef]
- Kenney, R.D. The North Atlantic Right Whale Consortium Database: A Guide for Users and Contributors; North Atlantic Right Whale Consortium: Halifax, NS, Canada, 2019. [Google Scholar]
- NARWC. Identification and Sightings Data of the North Atlantic Right Whale Consortium as of January 2021 (Online Archive at New England Aquarium; NARWC: Halifax, NS, Canada, 2021. [Google Scholar]
- DFO. Whale Sightings Database, Team Whale, Fisheries and Oceans; DFO: Ottawa, ON, Canada, 2022.
- DFO. Review of North Atlantic Right Whale Occurrence and Risk of Entanglements in Fishing Gear and Vessel Strikes in Canadian Waters; DFO Canadian Science Advisory Secretariat Science Advisory Report; DFO: Ottawa, ON, Canada, 2019.
- Donlon, C.J.; Martin, M.; Stark, J.; Roberts-Jones, J.; Fiedler, E.; Wimmer, W. The Operational Sea Surface Temperature and Sea Ice Analysis (OSTIA) System. Remote Sens. Environ. 2012, 116, 140–158. [Google Scholar] [CrossRef]
- Good, S.; Fiedler, E.; Mao, C.; Martin, M.J.; Maycock, A.; Reid, R.; Roberts-Jones, J.; Searle, T.; Waters, J.; While, J.; et al. The Current Configuration of the OSTIA System for Operational Production of Foundation Sea Surface Temperature and Ice Concentration Analyses. Remote Sens. 2020, 12, 720. [Google Scholar] [CrossRef]
- Waite, A.M.; Raes, E.; Beckley, L.E.; Thompson, P.A.; Griffin, D.; Saunders, M.; Säwström, C.; O’Rorke, R.; Wang, M.; Landrum, J.P.; et al. Production and Ecosystem Structure in Cold-Core vs. Warm-Core Eddies: Implications for the Zooplankton Isoscape and Rock Lobster Larvae. Limnol. Ocean. 2019, 64, 2405–2423. [Google Scholar] [CrossRef]
- Wang, Y.; Castelao, R.M.; Yuan, Y. Seasonal Variability of Alongshore Winds and Sea Surface Temperature Fronts in Eastern Boundary Current Systems. J. Geophys. Res. Ocean. 2015, 120, 2385–2400. [Google Scholar] [CrossRef]
- Bourgault, D.; Koutitonsky, V.G. Real-time Monitoring of the Freshwater Discharge at the Head of the St. Lawrence Estuary. Atmosphere-Ocean 1999, 37, 203–220. [Google Scholar] [CrossRef]
- Galbraith, P.S.; Chassé, J.; Shaw, J.-L.; Dumas, J.; Caverhill, C.; Lefaivre, D.; Lafleur, C. Physical Oceanographic Conditions in the Gulf of St. Lawrence During 2020; DFO Canadian Science Advisory Secretariat Science Advisory Report 2021/045. Iv +; DFO: Ottawa, ON, Canada, 2021; p. 81.
- Fu, L.L.; Chelton, D.B. Large-Scale Ocean Circulation. In International Geophysics; Academic Press: Cambridge, MA, USA, 2001; Volume 69, p. 133. [Google Scholar]
- Bonjean, F.; Lagerloef, G.S. Diagnostic Model and Analysis of the Surface Currents in the Tropical Pacific Ocean. J. Phys. Oceanogr. 2002, 32, 2938–2954. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, L.; Li, Z. Geostrophic Current Estimation Using Altimetric Cross-Track Method in Northwest Pacific. IOP Conf. Ser. Earth Environ. Sci. 2014, 17, 012105. [Google Scholar] [CrossRef]
- Perrie, W.; Shen, H.; Tao, J.; Danielson, R.E. A Physical Index Towards Real Time Monitoring of Potential Whale Habitats from Space. Can. Tech. Rep. Fish. Aquat. Sci. 2023, 3533, 35. [Google Scholar]
- Van Sebille, E.; Aliani, S.; Law, K.L.; Maximenko, N.; Alsina, J.M.; Bagaev, A.; Bergmann, M.; Chapron, B.; Chubarenko, I.; Cózar, A.; et al. The Physical Oceanography of the Transport of Floating Marine Debris. Environ. Res. Lett. 2020, 15, 023003. [Google Scholar] [CrossRef]
- Cohen, S.; Bush, E.; Zhang, X.; Gillett, N.; Bonsal, B.; Derksen, C.; Flato, G.; Greenan, B.; Watson, E. Changes in Canada’s Regions in a National and Global Context. In Canada’s Changing Climate Report; Bush, E., Lemmen, D.S., Eds.; Government of Canada: Ottawa, ON, Canada, 2019; pp. 424–443. [Google Scholar]
- Stone, G.S.; Kraus, S.D.; Prescott, J.H.; Hazard, K.W. Significant Aggregations of the Endangered Right Whale Eubalaena glacialis, on the Continental Shelf of Nova Scotia. Can. Field-Nat. 1988, 102, 471–474. [Google Scholar] [CrossRef]
- Murison, L.D.; Gaskin, G.E. The Distribution of Right Whales and Zooplankton in the Bay of Fundy, Canada. Can. J. Zool. 1989, 67, 1411–1420. [Google Scholar] [CrossRef]
- Sameoto, D.D.; Herman, A.W. Life Cycle and Distribution of Calanus Finmarchicus in Deep Basins on the Nova Scotia Shelf and Seasonal Changes in Calanus spp. Mar. Ecol. Prog. Ser. 1990, 66, 225–237. [Google Scholar] [CrossRef]
- Miller, C.B.; Cowles, T.J.; Wiebe, P.H.; Copley, N.J.; Grigg, H. Phenology in Calanus Finmarchicus; Hypotheses about Control Mechanisms. Mar. Ecol. Prog. Ser. 1991, 72, 79–91. [Google Scholar] [CrossRef]
- Woodley, T.H.; Gaskin, D.E. Environmental Characteristics of North Atlantic Right and Fin Whale Habitat in the Lower Bay of Fundy, Canada. Can. J. Zool. 1996, 74, 75–84. [Google Scholar] [CrossRef]
- Côté, B.; El-Sabh, M.; Durantaye, R. Biological and Physical Characteristics of a Frontal Region Associated with the Arrival of Spring Freshwater Discharge in the Southwestern Gulf of St. Lawrence. In The Role of Freshwater Outflow in Coastal Marine Ecosystems; Springer: Berlin/Heidelberg, Germany, 1986; pp. 261–269. [Google Scholar]
- Baumgartner, M.F.; Mate, B.R. Summertime Foraging Ecology of North Atlantic Right Whales. Mar. Ecol. Prog. Ser. 2003, 264, 123–135. [Google Scholar] [CrossRef]
- Maps, F.; Plourde, S.; McQuinn, I.H.; St.-Onge-Drouin, S.; Lavoie, D.; Chassé, J.; Lesage, V. Linking Acoustics and Finite-Time Lyapunov Exponents Reveals Areas and Mechanisms of Krill Aggregation within the Gulf of St. Lawrence, eastern Canada. Limnol. Oceanogr. 2015, 60, 1965–1975. [Google Scholar] [CrossRef]
- Franklin, K.; Cole, T.; Cholewiak, D.; Duley, P.; Crowe, L.; Hamilton, P.; Knowlton, A.; Taggart, C.; Johnson, H. Using Sonobuoys and Visual Surveys to Characterize North Atlantic Right Whale (Eubalaena glacialis) Calling Behavior in the Gulf of St. Lawrence. Endanger. Species Res. 2022, 49, 159–174. [Google Scholar] [CrossRef]
- Parks, S.E.; Hamilton, P.K.; Kraus, S.D.; Tyack, P.L. The Gunshot Sound Produced by Male North Atlantic Right Whales (Eubalaena glacialis) and Its Potential Function in Reproductive Advertisement. Mar. Mammal Sci. 2005, 21, 458–475. [Google Scholar] [CrossRef]
- Matthews, L.P.; McCordic, J.A.; Parks, S.E. Remote Acoustic Monitoring of North Atlantic Right Whales (Eubalaena glacialis) Reveals Seasonal and Diel Variations in Acoustic Behavior. PLoS ONE 2014, 9, 91367. [Google Scholar] [CrossRef]
- Jaquet, N. How Spatial and Temporal Scales Influence Understanding of Sperm Whale Distribution: A Review. Mammal Rev. 1996, 26, 51–65. [Google Scholar] [CrossRef]
- Barlow, D.R.; Klinck, H.; Ponirakis, D.; Garvey, C.; Torres, L.G. Temporal and Spatial Lags between Wind, Coastal Upwelling, and Blue Whale Occurrence. Sci. Rep. 2021, 11, 6915. [Google Scholar] [CrossRef]
- Ciani, D.; Sabatini, M.; Buongiorno Nardelli, B.; Lopez Dekker, P.; Rommen, B.; Wethey, D.S.; Yang, C.; Liberti, G.L. Sea Surface Temperature Gradients Estimation Using Top-of-Atmosphere Observations from the ESA Earth Explorer 10 Harmony Mission: Preliminary Studies. Remote Sens. 2023, 15, 1163. [Google Scholar] [CrossRef]
- Sorochan, K.A.; Brennan, C.E.; Plourde, S.; Johnson, C.L. Spatial Variation and Transport of Abundant Copepod Taxa in the Southern Gulf of St. Lawrence in autumn. J. Plankton Res. 2021, 43, 908–926. [Google Scholar] [CrossRef]
- Ohashi, K.; Sheng, J. Influence of St. Lawrence River Discharge on the Circulation and Hydrography in Canadian Atlantic Waters. Cont. Shelf Res. 2013, 58, 32–49. [Google Scholar] [CrossRef]
- Meyer-Gutbrod, E.L.; Greene, C.H.; Davies, K.T.; Johns, D.G. Ocean Regime Shift Is Driving Collapse of the North Atlantic Right Whale Population. Oceanography 2021, 34, 22–31. [Google Scholar] [CrossRef]
- Blais, M.; Galbraith, P.S.; Plourde, S.; Devine, L.; Lehoux, C. Chemical and Biological Oceanographic Conditions in the Estuary and Gulf of St. Lawrence During 2023; DFO Canadian Science Advisory Secretariat Science Advisory Research Document 385; DFO: Ottawa, ON, Canada, 2024; p. 66.
- Edelmann, D.; Móri, T.F.; Székely, G.J. On Relationships between the Pearson and the Distance Correlation Coefficients. Stat. Prob. Lett. 2021, 169, 1–6. [Google Scholar] [CrossRef]
- Fisher, R.A. On the Mathematical Foundations of Theoretical Statistics. Proc. R. Soc. Lond. A 1922, 222, 309–368. [Google Scholar] [CrossRef]
- Lenth, R.V. Least-Squares Means: The R Package Lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Redfern, J.V.; Ferguson, M.C.; Becker, E.A.; Hyrenbach, K.D.; Good, C.; Barlow, J.; Kaschner, K.; Baumgartner, M.F.; Forney, K.A.; Ballance, L.T.; et al. Techniques for Cetacean–Habitat Modeling. Mar. Ecol. Prog. Ser. 2006, 310, 271–295. [Google Scholar] [CrossRef]
- St-Pierre, A.P.; Koll-Egyed, T.; Harvey, V.; Lawson, J.W.; Sauvé, C.; Ollier, A.; Goulet, P.J.; Hammill, M.O.; Gosselin, J.-F. Distribution of North Atlantic Right Whales, Eubalaena glacialis, in Eastern Canada from Line-Transect Surveys from 2017 to 2022, Canadian Science Advisory Secretariat (CSAS) Research Document 2024/059. 2024. Available online: https://publications.gc.ca/collections/collection_2024/mpo-dfo/fs70-5/Fs70-5-2024-059-eng.pdf (accessed on 3 April 2025).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month Year | May | Jun. | Jul. | Aug. | Sep. | Oct. | All Month |
|---|---|---|---|---|---|---|---|
| 2015 | 0 | 4 | 3 | 264 | 2 | 0 | 273 |
| 2016 | 0 | 5 | 69 | 70 | 12 | 0 | 156 |
| 2017 | 4 | 280 | 828 | 513 | 141 | 45 | 1811 |
| 2018 | 96 | 912 | 1090 | 725 | 39 | 22 | 2884 |
| 2019 | 52 | 210 | 891 | 685 | 28 | 178 | 2044 |
| 2020 | 13 | 82 | 318 | 17 | 32 | 31 | 493 |
| 2021 | 24 | 246 | 564 | 117 | 8 | 1 | 960 |
| All Year | 189 | 1739 | 3763 | 2391 | 262 | 277 | 8621 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, J.; Shen, H.; Danielson, R.E.; Perrie, W. Sea Surface Temperature Fronts and North Atlantic Right Whale Sightings in the Western Gulf of St. Lawrence. J. Mar. Sci. Eng. 2025, 13, 1280. https://doi.org/10.3390/jmse13071280
Tao J, Shen H, Danielson RE, Perrie W. Sea Surface Temperature Fronts and North Atlantic Right Whale Sightings in the Western Gulf of St. Lawrence. Journal of Marine Science and Engineering. 2025; 13(7):1280. https://doi.org/10.3390/jmse13071280
Chicago/Turabian StyleTao, Jing, Hui Shen, Richard E. Danielson, and William Perrie. 2025. "Sea Surface Temperature Fronts and North Atlantic Right Whale Sightings in the Western Gulf of St. Lawrence" Journal of Marine Science and Engineering 13, no. 7: 1280. https://doi.org/10.3390/jmse13071280
APA StyleTao, J., Shen, H., Danielson, R. E., & Perrie, W. (2025). Sea Surface Temperature Fronts and North Atlantic Right Whale Sightings in the Western Gulf of St. Lawrence. Journal of Marine Science and Engineering, 13(7), 1280. https://doi.org/10.3390/jmse13071280