
Jellyfish and Ctenophores Around Gotland in the Baltic Sea—Local Data Contributing to Global Assessments




Abstract
1. Introduction
2. Materials and Methods
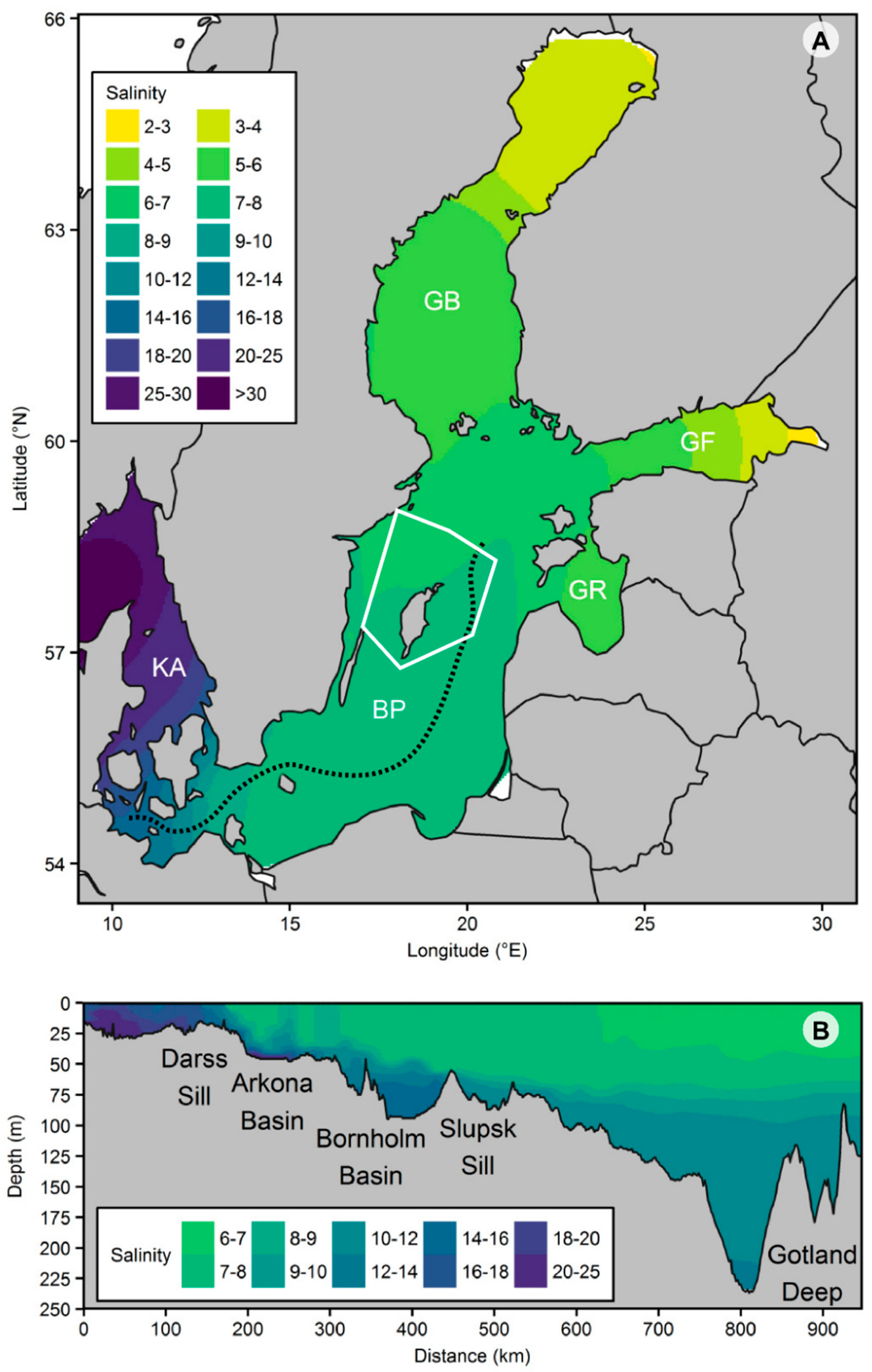
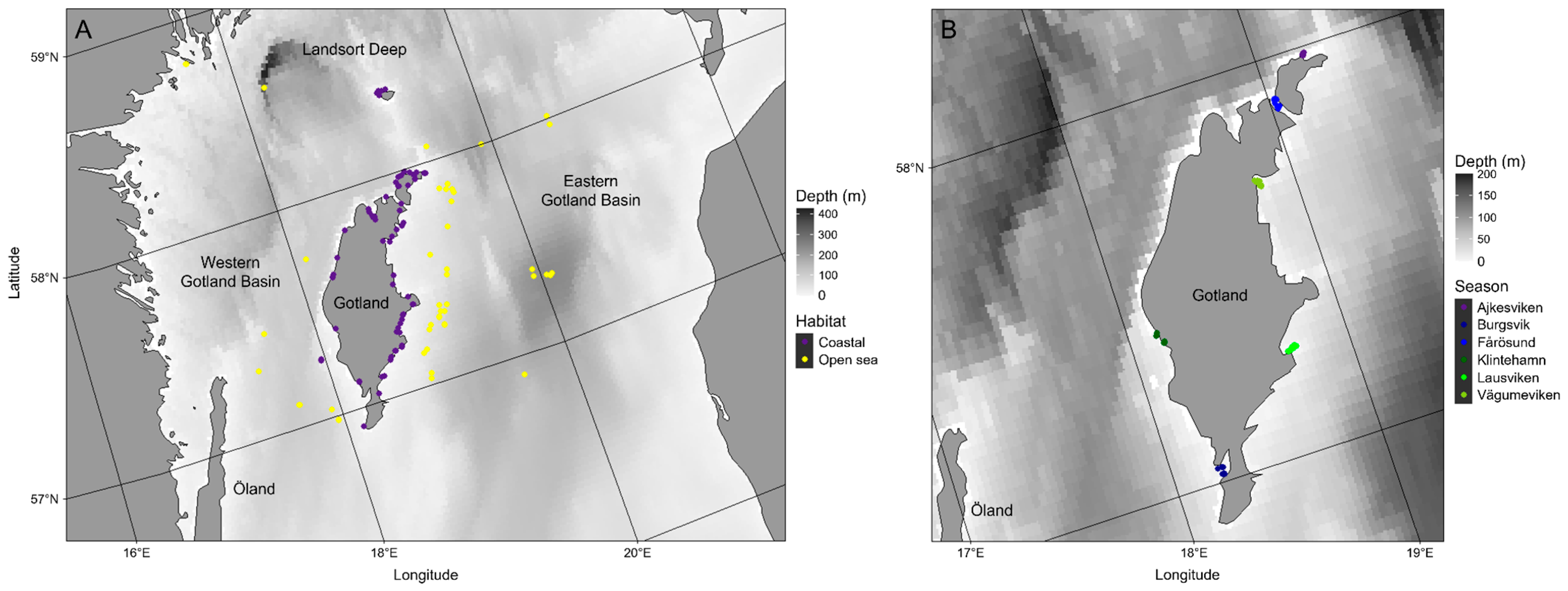
2.1. Study Area
2.2. Literature and Open-Source Data Mining
2.3. Opportunistic Jellyfish Abundance Estimates in 2022
2.4. Data Visualisation
3. Results
3.1. Species Richness (Literature and Open-Access Data)
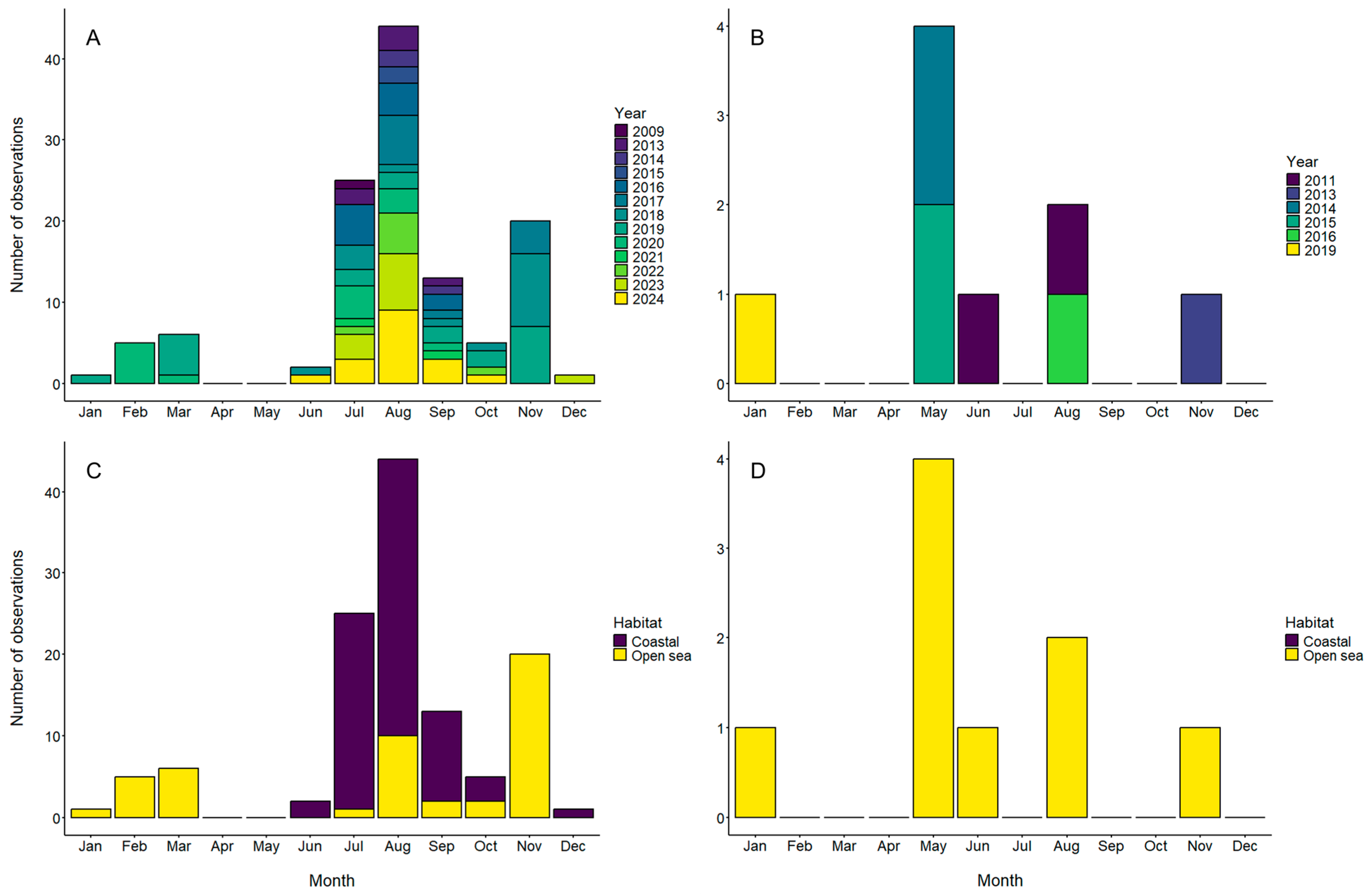
3.2. Phenology of Aurelia aurita and Mertensia ovum in Gotlandic Waters
3.3. Radio and Newspaper Mentioning of Jellyfish Around Gotland
3.4. Abundance Estimation Based on Camera Transects
4. Discussion
4.1. Gelatinous Zooplankton Species Richness in the Baltic Proper
4.2. Phenology of Aurelia aurita in the Central Baltic Sea
4.3. Summer Aurelia aurita Abundance in Gotlandic Coastal Waters
4.4. Long-Term Trends of Gelatinous Zooplankton in the Baltic Proper
4.5. New Ways for Gelatinous Zooplankton Research Around Gotland and FAIR Data
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inácio, M.; Karnauskaitė, D.; Baltranaitė, E.; Kalinauskas, M.; Bogdzevič, K.; Gomes, E.; Pereira, P. Ecosystem services of the Baltic Sea: An assessment and mapping perspective. Geogr. Sustain. 2020, 1, 256–265. [Google Scholar] [CrossRef]
- Rosciszewski-Dodgson, M.J.; Cirella, G.T. Environmental drivers affecting the status of top commercial fish stocks in the Baltic Sea: Review. Front. Mar. Sci. 2024, 11, 1399707. [Google Scholar] [CrossRef]
- Gustafsson, B.G.; Westman, P. On the causes for salinity variations in the Baltic Sea during the last 8500 years. Paleoceanogr. Paleoclimatol. 2002, 17, 12/1–12/14. [Google Scholar] [CrossRef]
- Remane, A. Die Brackwasserfauna. Zool. Anz. 1934, 36, 34–74. [Google Scholar]
- Kinne, O. (Ed.) Salinity: 3. Animals: 1. Invertebrates. In Marine Ecology: A Comprehensive, Integrated Treatise on Life in Oceans and Coastal Waters: 1. Environmental Factors: 2; Wiley: London, UK, 1971; pp. 821–995. [Google Scholar]
- Khlebovich, V.V. Critical Salinity of Biological Processes; Leningrad: Nauka, Russia, 2010; p. 235. [Google Scholar]
- Telesh, I.; Schubert, H.; Skarlato, S. Life in the salinity gradient: Discovering mechanisms behind a new biodiversity pattern. Estuar. Coast. Shelf. Sci. 2013, 135, 317–327. [Google Scholar] [CrossRef]
- Ojaveer, H.; Jaanus, A.; MacKenzie, B.R.; Martin, G.; Olenin, S.; Radziejewska, T.; Telesh, I.; Zettler, M.L.; Zaiko, A. Status of biodiversity in the Baltic Sea. PLoS ONE 2010, 5, e12467. [Google Scholar] [CrossRef]
- Jaspers, C.; Bezio, N.; Hinrichsen, H.-H. Diversity and physiological tolerance of native and invasive jellyfish/ctenophores along the extreme salinity gradient of the Baltic Sea. Diversity 2021, 13, 57. [Google Scholar] [CrossRef]
- Gershwin, L.-A. Jellyfish–A Natural History; The University of Chicago Press: Chicago, IL, USA, 2016; p. 224. [Google Scholar]
- Chi, X.; Dierking, J.; Hoving, H.-J.T.; Lüskow, F.; Denda, A.; Christiansen, B.; Sommer, U.; Hansen, T.; Javidpour, J. Tackling the jelly web: New insights into the trophic ecology of gelatinous zooplankton in oceanic food webs of the eastern tropical Atlantic assessed by stable isotope analysis. Limnol. Ocean. 2021, 66, 289–305. [Google Scholar] [CrossRef]
- Bossier, S.; Palacz, A.P.; Nielsen, J.R.; Christensen, A.; Hoff, A.; Maar, M.; Gislason, H.; Bastardie, F.; Gorton, R.; Fulton, E.A. The Baltic Sea Atlantis: An integrated end-to-end modelling framework evaluating ecosystem-wide effects of human-induced pressures. PLoS ONE 2018, 13, e0199168. [Google Scholar] [CrossRef]
- Brodeur, R.D.; Auth, T.D.; Phillips, A.J. Major shifts in pelagic micronekton and macrozooplankton community structure in an upwelling ecosystem related to an unprecedented marine heatwave. Front. Mar. Sci. 2019, 6, 212. [Google Scholar] [CrossRef]
- Tinta, T.; Klun, K.; Herndl, G.J. The importance of jellyfish–microbe interactions for biogeochemical cycles in the ocean. Limnol. Ocean. 2021, 66, 2011–2032. [Google Scholar] [CrossRef]
- Madin, L.P.; Harbison, G.R. Gelatinous zooplankton. In Encyclopedia of Ocean Sciences; Steele, J., Thorpe, S., Turekian, K., Eds.; Academic Press: London, UK, 2019; Volume 2, pp. 1120–1130. [Google Scholar]
- Harbison, G.R.; Madin, L.P.; Swanberg, N.R. On the natural history and distribution of oceanic ctenophores. Deep. Sea Res. 1978, 25, 233–256. [Google Scholar] [CrossRef]
- Graham, W.M.; Gelcich, S.; Robinson, K.L.; Duarte, C.M.; Brotz, L.; Purcell, J.E.; Madin, L.P.; Mianzan, H.; Sutherland, K.R.; Uye, S.; et al. Linking human well-being and jellyfish: Ecosystem services, impacts, and societal responses. Front. Ecol. Environ. 2014, 12, 515–523. [Google Scholar] [CrossRef]
- Sigurdsson, G.M.; Lüskow, F.; Gislason, A.; Svavarsson, J. Detached tentacles of lion’s mane jellyfish Cyanea capillata can injure aquaculture fish. Aquacult. Env. Interact. 2024, 16, 263–266. [Google Scholar] [CrossRef]
- Labuce, A.; Batare, L.; Ikauniece, A. Spatiotemporal dataset on moon jellyfish Aurelia aurita incidental observations in the Gulf of Riga and Eastern Gotland Basin, Baltic Sea. Data Brief. 2024, 56, 110880. [Google Scholar] [CrossRef]
- Brotz, L.; Cheung, W.W.L.; Kleisner, K.; Pakhomov, E.A.; Pauly, D. Increasing jellyfish populations: Trends in Large Marine Ecosystems. Hydrobiologia 2012, 690, 3–20. [Google Scholar] [CrossRef]
- Sommer, F.; Stibor, H.; Sommer, U.; Velimirov, B. Grazing by mesozooplankton from Kiel Bight, Baltic Sea, on different sized algae and natural seston size fractions. Mar. Ecol. Prog. Ser. 2000, 199, 43–53. [Google Scholar] [CrossRef]
- Stoltenberg, I.; Dierking, J.; Müller-Navarra, D.C.; Javidpour, J. Review of jellyfish trophic interactions in the Baltic Sea. Mar. Biol. Res. 2021, 17, 311–326. [Google Scholar] [CrossRef]
- Javidpour, J.; Molinero, J.C.; Peschutter, J.; Sommer, U. Seasonal changes and population dynamics of the ctenophore Mnemiopsis leidyi after its first year of invasion in the Kiel Fjord, Western Baltic Sea. Biol. Inv. 2009, 11, 873–882. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Goldstein, J.; Lundgreen, K.; Lüskow, F. Jellyfish and ctenophores in the environmentally degraded Limfjorden (Denmark) during 2014—Species composition, population densities and predation impact. Fish Aquac. J. 2015, 6, 1000137. [Google Scholar] [CrossRef]
- Lüskow, F.; Riisgård, H.U. Population predation impact of jellyfish (Aurelia aurita) controls the maximum umbrella size and somatic degrowth in temperate Danish waters (Kertinge Nor and Mariager Fjord). Vie et Milieu 2016, 66, 233–243. [Google Scholar]
- Majaneva, S.; Setälä, O.; Gorokhova, E.; Lehtiniemi, M. Feeding of the Arctic ctenophore Mertensia ovum in the Baltic Sea: Evidence of the use of microbial prey. J. Plank. Res. 2014, 36, 91–103. [Google Scholar] [CrossRef]
- Jaspers, C.; Huwer, B.; Antajan, E.; Hosia, A.; Hinrichsen, H.; Biastoch, A.; Angel, D.; Asmus, R.; Augustin, C.; Bagheri, S.; et al. Ocean current connectivity propelling the secondary spread of a marine invasive comb jelly across western Eurasia. Glob. Ecol. Biogeogr. 2018, 27, 814–827. [Google Scholar] [CrossRef]
- Cant, J.; Jones, O.R.; Ellingsen, I.; Laverick, J.H.; Majaneva, S.; Dierking, J.; Aberle, N.; Javidpour, J. Coupling hydrodynamic drifting simulations and seasonal demographics to forecast the occurrence of jellyfish blooms. bioRxiv 2025. bioRxiv:584451. [Google Scholar]
- Janas, U.; Witek, Z. The occurrence of medusae in the southern Baltic and their importance in the ecosystem, with special emphasis on Aurelia aurita. Oceanologia 1993, 34, 69–84. [Google Scholar]
- Karasiova, E.M.; Semenova, A.S.; Zezera, A.S.; Karpushevski, I.V. The quantitative distribution of scyphozoan medusae Aurelia aurita and crustacean plankton in the Baltic Sea during autumn season. Tr. AtlantNIRO 2019, 3, 12–25. [Google Scholar]
- Lischka, M. Abundanz, Verteilung und Nahrungsökologie von Scyphomedusen in der Zentralen Ostsee. Diploma Thesis, Christian Albrechts University, Kiel, Germany, 1999; 91p. [Google Scholar]
- Olenycz, M. Gelatinous zooplankton—A potential threat to the ecosystem of the Puck Bay (the southern Baltic Sea, Poland). Bull. Mar. Inst. Gdańsk 2015, 30, 78–85. [Google Scholar]
- Brulińska, D.; Olenycz, M.; Ziółkowska, M.; Mudrak-Cegiołka, S.; Wołowicz, M. Moon jellyfish, Aurelia aurita, in the Gulf of Gdańsk: Threatening predator or not? Boreal Environ. Res. 2016, 21, 528–540. [Google Scholar]
- Polunina, J.J.; Kondrashov, A.A.; Utkina, J.V. Distribution of invader Mnemiopsis leidyi (Ctenophora) in early stages of development in the southeastern Baltic Sea in autumn 2021. Russ. J. Biol. Inv. 2023, 14, 222–228. [Google Scholar] [CrossRef]
- Jaspers, C.; Hauss, H.; Hinrichsen, H.H. Jellyfish and comb jellies in the Baltic Sea: Depth resolved distribution pattern along the salinity gradient of the Baltic Sea during September 2020. Data Brief 2025, 60, 111511. [Google Scholar] [CrossRef]
- Barz, K.; Hirche, H.J. Seasonal development of scyphozoan medusae and the predatory impact of Aurelia aurita on the zooplankton community in the Bornholm Basin (Central Baltic Sea). Mar. Biol. 2005, 147, 465–476. [Google Scholar] [CrossRef]
- Karasiova, E.M.; Karpushevski, I.V.; Zezera, A.S. Scyphozoan medusae: The seasonal and spatial distribution along transects through the Baltic Sea. In Proceedings of the ICES Ann Sci Conference, A Coruña, Spain, 2014. A 17. [Google Scholar]
- Storr-Paulsen, M.; Huwer, B. Changes in distribution and lengths of Mnemiopsis leidyi in the central Baltic Sea between fall and spring. Aquat. Inv. 2008, 3, 429–434. [Google Scholar] [CrossRef]
- Huwer, B.; Storr-Paulsen, M.; Riisgård, H.U.; Haslob, H. Abundance, horizontal and vertical distribution of the invasive ctenophore Mnemiopsis leidyi in the central Baltic Sea, November 2007. Aquat. Inv. 2008, 3, 113–124. [Google Scholar] [CrossRef]
- Haraldsson, M.; Jaspers, C.; Tiselius, P.; Aksnes, D.L.; Andersen, T.; Titelman, J. Environmental constraints of the invasive Mnemiopsis leidyi in Scandinavian waters. Limnol. Ocean. 2013, 58, 37–48. [Google Scholar] [CrossRef]
- Jaspers, C.; Haraldsson, M.; Lombard, F.; Bolte, S.; Kiørboe, T. Seasonal dynamics of early life stages of invasive and native ctenophores give clues to invasion and bloom potential in the Baltic Sea. J. Plank. Res. 2013, 35, 582–594. [Google Scholar] [CrossRef]
- Oesterwind, D.; Bartolino, V.; Behrens, J.W.; Erlandsson, M.; Florin, A.-B.; Henseler, C.; Jakubowska-Lehrmann, M.; Jaspers, C.; Lehtiniemi, M.; Naddafi, R.; et al. Disentangling the potential effects of four non-indigenous species on commercially and recreationally used fish stocks in the Baltic Sea—A review. Biol. Inv. 2025, 27, 76. [Google Scholar] [CrossRef]
- Gorokhova, E.; Lehtiniemi, M.; Viitasalo-Frösen, S.; Haddock, S.H.D. Molecular evidence for the occurrence of ctenophore Mertensia ovum in the northern Baltic Sea and implications for the status of the Mnemiopsis leidyi invasion. Limnol. Ocean. 2009, 54, 2025–2033. [Google Scholar] [CrossRef]
- Lehtiniemi, M.; Gorokhova, E.; Bolte, S.; Haslob, H.; Huwer, B.; Katajisto, T.; Lennuk, L.; Majaneva, S.; Põllumäe, A.; Schaber, M.; et al. Distribution and reproduction of the Arctic ctenophore Mertensia ovum in the Baltic Sea. Mar. Ecol. Prog. Ser. 2013, 491, 111–124. [Google Scholar] [CrossRef]
- Schneider, G. Role of advection in the distribution and abundance of Pleurobrachia pileus in Kiel Bight. Mar. Ecol. Prog. Ser. 1987, 41, 99–102. [Google Scholar] [CrossRef]
- Lüskow, F. Importance of environmental monitoring: Long-term record of jellyfish (Aurelia aurita) biomass in a shallow semi-enclosed cove (Kertinge Nor, Denmark). Reg. Stud. Mar. Sci. 2020, 34, 100998. [Google Scholar] [CrossRef]
- Nolan, G.; Cusack, C.; Fitzhenry, D.; McGovern, E.; Cronin, M.; O’Donnell, G.; O’Dowd, L.; Clarke, M.; Reid, D.G.; Clarke, D.; et al. Baseline Study of Essential Ocean Variable Monitoring in Irish Waters; Current Measurement Programmes & Data Quality; Marine Institute: Galway, Ireland, 2021; 63p. [Google Scholar]
- Wieczorek, G.; Hagen, E.; Umlauf, L. Eastern Gotland Basin case study of thermal variability in the wake of deep water intrusions. J. Mar. Syst. 2008, 74, S65–S79. [Google Scholar] [CrossRef]
- Liblik, T.; Naumann, M.; Alenius, P.; Hansson, M.; Lips, U.; Nausch, G.; Tuomi, L.; Wesslander, K.; Laanemets, J.; Viktorsson, L. Propagation of impact of the recent major Baltic inflows from the Eastern Gotland Basin to the Gulf of Finland. Front. Mar. Sci. 2018, 5, 222. [Google Scholar] [CrossRef]
- Laine, A.O.; Sandler, H.; Andersin, A.-B.; Stigzelius, J. Long-term changes of macrozoobenthos in the Eastern Gotland Basin and the Gulf of Finland (Baltic Sea) in relation to the hydrographical regime. J. Sea Res. 1997, 38, 135–159. [Google Scholar] [CrossRef]
- Kanarik, H.; Tuomi, L.; Björkqvist, J.-V.; Kärnä, T. Improving Baltic Sea wave forecasts using modelled surface currents. Ocean. Dyn. 2021, 71, 635–653. [Google Scholar] [CrossRef]
- Müller, J.D. Ocean Acidification in the Baltic Sea. Involved Processes, Metrology of pH in Brackish Waters, and Calcification Under Fluctu-ating Conditions. Ph.D. Thesis, University of Rostock, Rostock, Germany, 2018; p. 52. [Google Scholar]
- Wihen, A. Identification and Population Genetic Analysis of Jellyfish Blooms in Trondheimsfjord. Master’s Thesis, Norwegian University of Science and Technology, Trondheim, Norway, 2021; p. 55. [Google Scholar]
- OBIS Ocean Biodiversity Information System (2025). Available online: https://obis.org (accessed on 6 March 2025).
- ICES Historical Plankton Dataset (1901–1912); ICES: Copenhagen, Denmark, 2010.
- Fish Trawl Survey: ICES Baltic International Trawl Survey for Commercial Fish Species; ICES Database of Trawl Surveys: Copenhagen, Denmark, 2010.
- SHARK; Stockholm University; University of Gothenburg; Linnaeus University; Swedish Agency for Marine and Water Management; Swedish Environmental Protection Agency; Swedish Meteorological and Hydrological Institute. National Epibenthos monitoring in Sweden Since 1992; EMODnet Product Catalogue: Online, 2017. [Google Scholar]
- SHARK; Swedish County Administration Boards; Swedish Municipalities; Swedish Coalitions of Water Conservation; Swedish Meteorological and Hydrological Institute. Regional Monitoring and Monitoring Projects of Epibenthos in Sweden Since 1994; EMODnet Product Catalogue: Online, 2017. [Google Scholar]
- Finnish Environment Institute SYKE. Finnish Baltic Sea Zooplankton Monitoring. 2018. Available online: https://obis.org/dataset/531637ff-203d-447a-8595-519080144a1d (accessed on 15 February 2025).
- Estonian Marine Institute; University of Tartu. Zooplankton biomass and abundance in Estonian territorial waters 1994–2016. 2019. Available online: https://emodnet.ec.europa.eu/geonetwork/srv/api/records/6d617269-6e65-696e-666f-000000006022 (accessed on 15 February 2025).
- ICES Eggs and Larvae Database: Copenhagen, Denmark. Available online: https://www.ices.dk/data/data-portals/Pages/Eggs-and-larvae.aspx (accessed on 15 February 2025).
- Lindberg, A.; Andersson, M.; Nordlund, L.M.; Olsson, I. Provfisken i ålgräsängar runt Gotland 2022; Report to the County Administrative Board; Länsstyrelsen i Gotlands län: Visby, Sweden, 2023; p. 54. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2024; Available online: http://www.R-project.org/ (accessed on 15 February 2025).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, version 3.5.2.; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 15 February 2025).
- Vihtakari, M. ggOceanMaps: Plot data on Oceanographic Maps Using ‘ggplot2’, R package version 2.1.1; Comprehensive R Archive Network: Vienna, Austria, 2023. [Google Scholar]
- Dunnington, D. ggspatial: Spatial Data Framework for ggplot2; 2023. Available online: https://paleolimbot.github.io/ggspatial/ (accessed on 15 February 2025).
- Fridolfsson, E.; Bunse, C.; Lindehoff, E.; Farnelid, H.; Pontiller, B.; Bergström, K.; Pinhassi, J.; Legrand, C.; Hylander, S. Multiyear analysis uncovers coordinated seasonality in stocks and composition of the planktonic food web in the Baltic Sea proper. Sci. Rep. 2023, 13, 11865. [Google Scholar] [CrossRef]
- Leppäkoski, E.; Gollasch, S.; Gruszka, P.; Ojaveer, H.; Olenin, S.; Panov, V. The Baltic—A sea of invaders. Can. J. Fish. Aquat. Sci. 2002, 59, 1175–1188. [Google Scholar] [CrossRef]
- von Numers, M. The first record of Maeotias marginata (Modeer, 1791) (Cnidaria, Hydrozoa) from Finland, northern Baltic Sea. BioInv. Rec. 2013, 2, 39–42. [Google Scholar] [CrossRef]
- Jaspers, C.; Huwer, B.; Weiland-Bräuer, N.; Clemmesen, C. First record of the non-indigenous jellyfish Blackfordia virginica (Mayer, 1910) in the Baltic Sea. Helgol. Mar. Res. 2018, 72, 13. [Google Scholar] [CrossRef]
- Serandour, B.; Leroy, B.; Blenckner, T.; Mittermayer, F.; Clemmesen, C.; Cruz, J.; Nowaczyk, A.; Winder, M.Q. Assessing the invasion risk of the cnidaria Blackfordia virginica Mayer, 1910: A threat to the Baltic Sea ecosystem? Biol. Inv. 2025, 27, 106. [Google Scholar] [CrossRef]
- Lundberg, S.; Svensson, J.-E.; Petrusek, A. Craspedacusta invasions in Sweden. Verh. Intern. Ver. Limnol 2005, 29, 899–902. [Google Scholar] [CrossRef]
- Lüskow, F.; Lehtiniemi, M.; Väinölä, R.; von Numers, M.; Pakhomov, E.A. Spreading of the non-indigenous freshwater jellyfish Craspedacusta sowerbii in Finland. Hydrobiologia 2025. [Google Scholar]
- Dutz, J.; Kremp, A.; Zettler, M.L. Biological assessment of the Baltic Sea 2020. Mar. Sci. Rep. Warn. 2020, 120, 82. [Google Scholar]
- Dutz, J.; Zettler, M.L.; Kremp, A.; Kube, S. Biological assessment of the Baltic Sea 2023. Mar Sci Rep Warn. 2025, 129, 79. [Google Scholar]
- Compagnone, F.; Varricchione, M.; Stanisci, A.; Matteucci, G.; Carranza, M.L. Exploring the contribution of a generalist citizen science project for alien species detection and monitoring in coastal Areas. A case study on the Adriatic of central Italy. Diversity 2024, 16, 746. [Google Scholar] [CrossRef]
- Gröndahl, F. A comparative ecological study on the scyphozoans Aurelia aurita, Cyanea capillata and C. lamarckii in the Gullmar Fjord, western Sweden, 1982 to 1986. Mar. Biol. 1988, 97, 541–550. [Google Scholar] [CrossRef]
- Thill, H. Beiträge zur Kenntnis der Aurelia aurita (L.). Zeitschr. Wiss. Zool. 1937, 150, 51–96. [Google Scholar]
- Stoltenberg, I.; Mittermayer, F.; Clemmensen, C.; Dierking, J.; Javidpour, J. Predation on Baltic Sea yolk-sac herring larvae (Clupea harengus) by the invasive ctenophore Mnemiopsis leidyi. Fish. Res. 2024, 273, 106973. [Google Scholar] [CrossRef]
- Miyake, H.; Terazaki, M.; Kakinuma, Y. On the polyps of the common jellyfish Aurelia aurita in Kagoshima Bay. J. Ocean. 2002, 58, 451–459. [Google Scholar] [CrossRef]
- Di Camillo, C.G.; Betti, F.; Bo, M.; Martinelli, M.; Puce, S.; Bavestrello, G. Contribution to the understanding of seasonal cycle of Aurelia aurita (Cnidaria: Scyphozoa) scyphopolyps in the northern Adriatic Sea. J. Mar. Biol. Assoc. UK 2010, 90, 1105–1110. [Google Scholar] [CrossRef]
- Toyokawa, M.; Aoki, K.; Yamada, S.; Yasuda, A.; Murata, Y.; Kikuchi, T. Distribution of ephyrae and polyps of jellyfish Aurelia aurita (Linnaeus 1758) sensu lato in Mikawa Bay. Jpn. J. Ocean. 2011, 67, 209–218. [Google Scholar] [CrossRef]
- Shibata, H.; Miyake, H.; Goto, T.; Adachi, A.; Toshino, S. Wild polyps of the blooming jellyfish Aurelia limbata (Brandt, 1838) (Cnidaria: Scyphozoa) found on deep-sea debris off Sanriku, Japan. Plank. Benthos. Res. 2015, 10, 133–140. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Jaspers, C.; Serre, S.; Lundgreen, K. Occurrence, inter-annual variability and zooplankton-predation impact of the invasive ctenophore Mnemiopsis leidyi and the native jellyfish Aurelia aurita in Limfjorden (Denmark) in 2010 and 2011. BioInv. Rec. 2012, 1, 145–159. [Google Scholar] [CrossRef]
- Janßen, H.; Augustin, C.B.; Hinrichsen, H.H.; Kube, S. Impact of secondary hard substrate on the distribution and abundance of Aurelia aurita in the western Baltic Sea. Mar. Poll. Bull. 2013, 75, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.L.; Ruzicka, J.J.; Decker, M.B.; Brodeur, R.D.; Hernandez, F.J.; Quiñones, J.; Acha, E.M.; Uye, S.; Mianzan, H.; Graham, W.M. Jellyfish, forage fish, and the world’s major fisheries. Oceanogr 2013, 27, 104–115. [Google Scholar] [CrossRef]
- Schnedler-Meyer, N.A.; Mariani, P.; Kiørboe, T. The global susceptibility of coastal forage fish to competition by large jellyfish. Proc. R. Soc. B. 2016, 283, 20161931. [Google Scholar] [CrossRef]
- Holland, M.M.; Artigas, L.F.; Atkinson, A.; Best, M.; Bresnan, E.; Devlin, M.; Eerkes-Medrano, D.; Johansen, M.; Johns, D.G.; Machairopoulou, M.; et al. Mind the gap—The need to integrate novel plankton methods alongside ongoing long-term monitoring. Ocean. Coast. Manag. 2025, 262, 107542. [Google Scholar] [CrossRef]
- Bashiri, B.; Barzandeh, A.; Männik, A.; Raudsepp, U. Variability of marine heatwaves’ characteristics and assessment of their potential drivers in the Baltic Sea over the last 42 years. Sci. Rep. 2024, 14, 22419. [Google Scholar] [CrossRef]
- Lindenthal, A.; Hinrichs, C.; Jandt-Scheelke, S.; Kruschke, T.; Lagemaa, P.; Lee, E.M.v.d.; Morrison, H.E.; Panteleit, T.R.; Raudsepp, U. Baltic Sea surface temperature analysis 2022: A study of marine heatwaves and overall high seasonal temperatures. State Planet 2024, 4, 1–16. [Google Scholar] [CrossRef]
- Quilestino-Olario, R.; Concolis, B.M.M.; Atup, D.P.D.; Cortes, A.; Yñiguez, A.T.; Edullantes, B. Co-occurrence of a marine heatwave and a reported tomato jellyfish (Crambione mastigophora Maas, 1903) bloom in March 2020 at El Nido, Palawan, Philippines. Plank. Benthos. Res. 2023, 18, 93–105. [Google Scholar] [CrossRef]
- Capidos, C.G.G.; Boco, S.R.; Morandini, A.C. Blooms of the dragon thimble jellyfish, Linuche draco, sometimes co-occur with marine heatwaves. Mar. Biodiv. 2024, 54, 38. [Google Scholar] [CrossRef]
- Hoving, H.J.T.; Neitzel, P.; Hauss, H.; Christiansen, S.; Kiko, R.; Robison, B.H.; Silva, P.; Körtzinger, A. In situ observations show vertical community structure of pelagic fauna in the eastern tropical North Atlantic off Cape Verde. Sci. Rep. 2020, 10, 21798. [Google Scholar] [CrossRef] [PubMed]
- Aubert, A.; Antajan, E.; Lynam, C.; Pitois, S.; Pliru, A.; Vaz, S.; Thibault, D. No more reason for ignoring gelatinous zooplankton in ecosystem assessment and marine management: Concrete cost-effective methodology during routine fishery trawl surveys. Mar. Pol. 2018, 89, 100–108. [Google Scholar] [CrossRef]
- Ratnarajah, L.; Abu-Alhaija, R.; Atkinson, A.; Batten, S.; Bax, N.J.; Bernard, K.S.; Canonico, G.; Cornils, A.; Everett, J.D.; Grigoratou, M.; et al. Monitoring and modelling marine zooplankton in a changing climate. Nat. Comm. 2023, 14, 564. [Google Scholar] [CrossRef] [PubMed]
- Titocci, J.; Pata, P.R.; Durazzano, T.; Ayata, S.-D.; Clerc, C.; Cornils, A.; Duffy, P.; Greer, A.T.; Halsband, C.; Heneghan, R.F.; et al. Pathways for converting zooplankton traits to ecological insights are paved with findable, accessible, interoperable, and reusable (FAIR) data practices. ICES J. Mar. Sci. 2025, 82, fsaf017. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Literature | OBIS | SBDI | iNaturalist® |
|---|---|---|---|---|
| Aurelia aurita (Linnaeus, 1758) | × | × (36) | × (63) | × (23) |
| Cyanea capillata (Linnaeus, 1758) | × | |||
| Mertensia ovum (Fabricius, 1780) | × | × (8) | × (1) | |
| Mnemiopsis leidyi A. Agassiz, 1865 * | × (11) | × (1) | ||
| Pleurobrachia pileus (O. F. Müller, 1776) * | × (5) |
| Site | Station | Date | Time (Local) | Latitude | Longitude | Bottom Depth (m) | Surface Temperature (°C) | Bottom Temperature (°C) | ΔTemperature Surface/Bottom (°C) | Wind Direction (°) | Wind Speed (m s−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ajkesviken | A1 | 18-Aug-22 | 19:18 | 57.9856 | 19.2295 | 6.0 | 21.2 | 20.6 | 0.7 | 30.0 | 1.5 |
| A2 | 17-Aug-22 | 18:11 | 57.9923 | 19.2402 | 3.0 | 20.7 | 21.2 | 0.4 | 160.0 | 4.0 | |
| A3 | 17-Aug-22 | 18:23 | 57.9914 | 19.2346 | 3.0 | 21.0 | 21.4 | 0.4 | 160.0 | 4.0 | |
| A4 | 17-Aug-22 | 18:33 | 57.9904 | 19.2293 | 2.3 | 21.0 | 21.4 | 0.4 | 160.0 | 4.0 | |
| A5 | 18-Aug-22 | 18:50 | 57.9863 | 19.2289 | 6.0 | 21.2 | 21.3 | 0.0 | 30.0 | 1.5 | |
| A6 | 17-Aug-22 | 19:02 | 57.9871 | 19.2273 | 5.2 | 20.3 | 20.8 | 0.5 | 160.0 | 4.5 | |
| A7 | 17-Aug-22 | 18:46 | 57.9891 | 19.2318 | 6.1 | 20.7 | 20.7 | 0.1 | 160.0 | 4.5 | |
| A8 | 18-Aug-22 | 18:10 | 57.9894 | 19.2306 | 4.8 | 20.9 | 20.8 | 0.1 | 30.0 | 1.0 | |
| A9 | 18-Aug-22 | 18:36 | 57.9883 | 19.2291 | 5.3 | 21.0 | 21.4 | 0.4 | 30.0 | 1.5 | |
| A10 | 18-Aug-22 | 18:26 | 57.9887 | 19.2319 | 6.0 | 21.0 | 20.9 | 0.1 | 30.0 | 1.5 | |
| Burgsvik | B1 | 14-Aug-22 | 9:59 | 57.0363 | 18.2330 | 3.4 | 18.2 | 12.7 | 5.5 | 75.0 | 5.0 |
| B2 | 14-Aug-22 | 9:50 | 57.0352 | 18.2284 | 3.0 | 18.2 | 12.3 | 5.9 | 75.0 | 5.0 | |
| B3 | 14-Aug-22 | 9:24 | 57.0515 | 18.2130 | 4.2 | 20.8 | 21.0 | 0.2 | 75.0 | 5.0 | |
| B4 | 14-Aug-22 | 9:09 | 57.0529 | 18.2313 | 2.7 | 22.1 | 21.8 | 0.3 | 75.0 | 5.0 | |
| B5 | 14-Aug-22 | 8:54 | 57.0515 | 18.2399 | 2.7 | 21.6 | 21.4 | 0.2 | 75.0 | 5.0 | |
| B6 | 15-Aug-22 | 19:07 | 57.0344 | 18.2364 | 2.9 | 19.3 | 18.7 | 0.6 | 82.5 | 4.0 | |
| B7 | 15-Aug-22 | 19:30 | 57.0358 | 18.2290 | 4.0 | 20.2 | 18.2 | 2.1 | 85.0 | 4.5 | |
| B8 | 15-Aug-22 | 19:53 | 57.0349 | 18.2255 | 4.3 | 20.7 | 18.7 | 2.0 | 85.0 | 4.5 | |
| B9 | 15-Aug-22 | 19:20 | 57.0359 | 18.2341 | 3.2 | 19.1 | 18.3 | 0.8 | 82.5 | 4.5 | |
| B10 | 15-Aug-22 | 19:43 | 57.0354 | 18.2270 | 4.8 | 20.5 | 18.4 | 2.1 | 85.0 | 4.5 | |
| Fårösund | F1 | 12-Sep-22 | 20:08 | 57.8928 | 19.0234 | 2.8 | 16.0 | 15.8 | 0.1 | 270.0 | 3.0 |
| F2 | 12-Sep-22 | 20:49 | 57.8780 | 19.0329 | 2.7 | 15.9 | 15.9 | 0.0 | 270.0 | 4.0 | |
| F3 | 25-Aug-22 | 18:49 | 57.9032 | 19.0365 | 2.1 | 20.5 | 20.5 | 0.0 | 61.5 | 3.0 | |
| F4 | 12-Sep-22 | 20:37 | 57.8800 | 19.0317 | 3.2 | 15.9 | 15.7 | 0.2 | 270.0 | 3.5 | |
| F5 | 25-Aug-22 | 18:57 | 57.9011 | 19.0375 | 4.5 | 20.3 | 20.5 | 0.3 | 61.5 | 4.0 | |
| F6 | 25-Aug-22 | 19:16 | 57.9006 | 19.0272 | 5.9 | 20.3 | 20.5 | 0.3 | 61.5 | 5.0 | |
| F7 | 12-Sep-22 | 20:27 | 57.8818 | 19.0303 | 2.9 | 15.7 | 15.7 | 0.0 | 270.0 | 3.5 | |
| F8 | 12-Sep-22 | 19:45 | 57.8828 | 19.0433 | 6.2 | 15.8 | 15.6 | 0.2 | 270.0 | 3.0 | |
| F9 | 25-Aug-22 | 19:28 | 57.9040 | 19.0280 | 5.5 | 20.5 | 20.5 | 0.0 | 61.5 | 5.5 | |
| F10 | 25-Aug-22 | 19:05 | 57.9012 | 19.0363 | 3.9 | 20.3 | 20.5 | 0.2 | 61.5 | 4.0 | |
| Klintehamn | K1 | 23-Aug-22 | 19:46 | 57.4016 | 18.1565 | 4.1 | 19.4 | 18.1 | 1.4 | 360.0 | 3.5 |
| K2 | 23-Aug-22 | 19:36 | 57.4022 | 18.1569 | 4.2 | 19.5 | 18.3 | 1.3 | 360.0 | 3.0 | |
| K3 | 16-Aug-22 | 19:40 | 57.4307 | 18.1387 | 3.0 | 21.7 | 21.8 | 0.1 | 180.0 | 5.0 | |
| K4 | 24-Aug-22 | 9:14 | 57.4021 | 18.1588 | 3.9 | 15.4 | 13.2 | 2.2 | 210.0 | 4.0 | |
| K5 | 23-Aug-22 | 19:23 | 57.4044 | 18.1615 | 3.0 | 19.4 | 19.0 | 0.4 | 360.0 | 3.0 | |
| K6 | 24-Aug-22 | 19:03 | 57.4020 | 18.1573 | 4.2 | 15.4 | 13.2 | 2.2 | 210.0 | 3.5 | |
| K7 | 16-Aug-22 | 19:31 | 57.4310 | 18.1367 | 4.2 | 21.6 | 21.5 | 0.1 | 180.0 | 5.0 | |
| K8 | 22-Aug-22 | 20:07 | 57.4255 | 18.1340 | 5.0 | 21.4 | 20.1 | 1.3 | 187.5 | 2.0 | |
| K9 | 22-Aug-22 | 19:52 | 57.4251 | 18.1325 | 2.5 | 21.2 | 20.5 | 0.6 | 187.5 | 1.5 | |
| K10 | 24-Aug-22 | 18:55 | 57.4024 | 18.1559 | 3.7 | 15.5 | 12.9 | 2.6 | 30.0 | 3.5 | |
| Lausviken | L1 | 14-Sep-22 | 19:14 | 57.2807 | 18.7012 | 2.8 | 15.9 | 15.9 | 0.0 | 90.0 | 6.0 |
| L2 | 16-Sep-22 | 19:17 | 57.2863 | 18.7459 | 2.8 | 14.2 | 14.0 | 0.1 | 310.0 | 5.5 | |
| L3 | 14-Sep-22 | 20:07 | 57.2862 | 18.7248 | 4.8 | 16.2 | 16.2 | 0.0 | 90.0 | 6.5 | |
| L4 | 16-Sep-22 | 19:49 | 57.2892 | 18.7389 | 2.5 | 13.9 | 13.5 | 0.5 | 310.0 | 5.5 | |
| L5 | 14-Sep-22 | 19:56 | 57.2836 | 18.7304 | 4.6 | 16.0 | 16.0 | 0.0 | 90.0 | 6.5 | |
| L6 | 16-Sep-22 | 18:57 | 57.2879 | 18.7279 | 4.8 | 14.3 | 11.5 | 2.8 | 310.0 | 5.5 | |
| L7 | 14-Sep-22 | 19:38 | 57.2801 | 18.7114 | 3.0 | 15.9 | 15.9 | 0.0 | 90.0 | 7.0 | |
| L8 | 16-Sep-22 | 19:28 | 57.2880 | 18.7427 | 4.2 | 13.8 | 13.8 | 0.1 | 310.0 | 5.5 | |
| L9 | 16-Sep-22 | 19:39 | 57.2897 | 18.7404 | 2.9 | 13.7 | 13.5 | 0.2 | 310.0 | 6.0 | |
| L10 | 14-Sep-22 | 19:26 | 57.2794 | 18.7036 | 2.4 | 16.0 | 15.9 | 0.1 | 90.0 | 6.0 | |
| Vägumeviken | V1 | 6-Sep-22 | 20:40 | 57.7200 | 18.8238 | 2.9 | 17.1 | 17.1 | 0.0 | 285.0 | 6.5 |
| V2 | 7-Sep-22 | 21:02 | 57.7180 | 18.8155 | 5.4 | 16.8 | 16.8 | 0.0 | 50.0 | 5.5 | |
| V3 | 6-Sep-22 | 20:26 | 57.7162 | 18.8332 | 2.8 | 16.4 | 16.4 | 0.0 | 285.0 | 6.5 | |
| V4 | 7-Sep-22 | 20:28 | 57.7116 | 18.8331 | 2.8 | 15.9 | 15.9 | 0.0 | 50.0 | 3.5 | |
| V5 | 6-Sep-22 | 20:53 | 57.7223 | 18.8120 | 4.3 | 16.9 | 16.9 | 0.0 | 285.0 | 5.0 | |
| V6 | 7-Sep-22 | 20:42 | 57.7169 | 18.8235 | 4.8 | 16.7 | 16.6 | 0.0 | 50.0 | 4.5 | |
| V7 | 7-Sep-22 | 20:12 | 57.7058 | 18.8364 | 4.2 | 16.6 | 16.5 | 0.1 | 50.0 | 5.5 | |
| V8 | 6-Sep-22 | 20:13 | 57.7130 | 18.8288 | 4.2 | 17.0 | 16.9 | 0.1 | 285.0 | 6.5 | |
| V9 | 7-Sep-22 | 19:58 | 57.7059 | 18.8286 | 6.9 | 16.9 | 16.9 | 0.0 | 45.0 | 5.0 | |
| V10 | 6-Sep-22 | 19:53 | 57.7026 | 18.8344 | 6.4 | 17.1 | 17.0 | 0.1 | 285.0 | 5.0 | |
| Mean | 4.0 | 18.3 | 17.7 | 0.7 | 155.6 | 4.4 | |||||
| SD | 1.2 | 2.5 | 2.9 | 1.2 | 106.3 | 1.4 | |||||
| Min | 8:54 | 2.1 | 13.7 | 11.5 | 0.0 | 30.0 | 1.0 | ||||
| Max | 21:02 | 6.9 | 22.1 | 21.8 | 5.9 | 360.0 | 7.0 |
| Site | Station | Recording Length (min) | Recording Length (s) | Orientation | Estimated Volume (m3) | No. Jellyfish | Abundance (ind. m−3) |
|---|---|---|---|---|---|---|---|
| Ajkesviken | A1 | 3:53 | 233 | F | 146.85 | 0 | 0.0 |
| A2 | 3:00 | 180 | F | 146.85 | 11 | 0.1 | |
| A3 | 3:54 | 234 | F | 146.85 | 1 | 0.0 | |
| A4 | 3:28 | 208 | F | 146.85 | 8 | 0.1 | |
| A5 | 3:46 | 226 | F | 146.85 | 0 | 0.0 | |
| A6 | 3:40 | 220 | F | 146.85 | 90 | 0.6 | |
| A7 | 3:35 | 215 | F | 146.85 | 6 | 0.0 | |
| A8 | 3:45 | 225 | F | 146.85 | 2 | 0.0 | |
| A9 | 3:29 | 209 | F | 146.85 | 0 | 0.0 | |
| A10 | 3:56 | 236 | F | 146.85 | 4 | 0.0 | |
| Burgsvik | B1 | 2:28 | 148 | D | 55.55 | 84 | 1.5 |
| B2 | 2:45 | 165 | D | 55.55 | 46 | 0.8 | |
| B3 | 2:44 | 164 | D | 55.55 | 125 | 2.3 | |
| B4 | 2:44 | 164 | D | 55.55 | 29 | 0.5 | |
| B5 | 2:38 | 158 | D | 55.55 | 2 | 0.0 | |
| B6 | 5:17 | 317 | D | 55.55 | 204 | 3.7 | |
| B7 | 5:47 | 347 | D | 55.55 | 31 | 0.6 | |
| B8 | 6:57 | 417 | D | 55.55 | 16 | 0.3 | |
| B9 | 4:25 | 265 | D | 55.55 | 32 | 0.6 | |
| B10 | 5:55 | 355 | D | 55.55 | 22 | 0.4 | |
| Fårösund | F1 | 3:40 | 220 | F | 146.85 | 102 | 0.7 |
| F2 | 3:21 | 201 | F | 146.85 | 46 | 0.3 | |
| F3 | 2:00 | 120 | F | 146.85 | 35 | 0.2 | |
| F4 | 2:20 | 140 | F | 146.85 | 153 | 1.0 | |
| F5 | 2:02 | 122 | F | 146.85 | 34 | 0.2 | |
| F6 | 4:11 | 251 | F | 146.85 | 2 | 0.0 | |
| F7 | 3:30 | 210 | F | 146.85 | 183 | 1.2 | |
| F8 | 6:02 | 362 | F | 146.85 | 126 | 0.9 | |
| F9 | 5:15 | 315 | F | 146.85 | 0 | 0.0 | |
| F10 | 4:27 | 267 | F | 146.85 | 7 | 0.0 | |
| Klintehamn | K1 | 4:07 | 247 | F | 146.85 | 366 | 2.5 |
| K2 | 3:50 | 230 | F | 146.85 | 208 | 1.4 | |
| K3 | 2:37 | 157 | D | 55.55 | 9 | 0.2 | |
| K4 | 2:47 | 167 | F | 146.85 | 638 | 4.3 | |
| K5 | 4:31 | 271 | F | 146.85 | 286 | 1.9 | |
| K6 | 4:49 | 289 | F | 146.85 | 108 | 0.7 | |
| K7 | 2:55 | 175 | D | 55.55 | 5 | 0.1 | |
| K8 | 5:38 | 338 | F | 146.85 | 106 | 0.7 | |
| K9 | 5:56 | 356 | F | 146.85 | 19 | 0.1 | |
| K10 | 3:26 | 206 | F | 146.85 | 203 | 1.4 | |
| Lausviken | L1 | 4:00 | 240 | F | 146.85 | 15 | 0.1 |
| L2 | 4:15 | 255 | F | 146.85 | 0 | 0.0 | |
| L3 | 3:19 | 199 | F | 146.85 | 0 | 0.0 | |
| L4 | 2:59 | 179 | F | 146.85 | 4 | 0.0 | |
| L5 | 3:04 | 184 | F | 146.85 | 0 | 0.0 | |
| L6 | 4:41 | 281 | F | 146.85 | 19 | 0.1 | |
| L7 | 3:48 | 228 | F | 146.85 | 5 | 0.0 | |
| L8 | 3:49 | 229 | F | 146.85 | 0 | 0.0 | |
| L9 | 3:18 | 198 | F | 146.85 | 3 | 0.0 | |
| L10 | 3:33 | 213 | F | 146.85 | 3 | 0.0 | |
| Vägumeviken | V1 | 3:57 | 237 | F | 146.85 | 145 | 1.0 |
| V2 | 4:57 | 297 | F | 146.85 | 12 | 0.1 | |
| V3 | 3:54 | 234 | F | 146.85 | 2 | 0.0 | |
| V4 | 4:48 | 288 | F | 146.85 | 2 | 0.0 | |
| V5 | 4:45 | 285 | F | 146.85 | 135 | 0.9 | |
| V6 | 5:35 | 335 | F | 146.85 | 86 | 0.6 | |
| V7 | 4:34 | 274 | F | 146.85 | 143 | 1.0 | |
| V8 | 3:17 | 197 | F | 146.85 | 4 | 0.0 | |
| V9 | 3:38 | 218 | F | 146.85 | 101 | 0.7 | |
| V10 | 3:34 | 214 | F | 146.85 | 4 | 0.0 | |
| Mean | 235 | 0.6 | |||||
| SD | 64 | 0.9 | |||||
| Min | 120 | 0.0 | |||||
| Max | 417 | 4.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lüskow, F.; Neitzel, P.; Lawrence, E.R.; Nordlund, L.M. Jellyfish and Ctenophores Around Gotland in the Baltic Sea—Local Data Contributing to Global Assessments. J. Mar. Sci. Eng. 2025, 13, 852. https://doi.org/10.3390/jmse13050852
Lüskow F, Neitzel P, Lawrence ER, Nordlund LM. Jellyfish and Ctenophores Around Gotland in the Baltic Sea—Local Data Contributing to Global Assessments. Journal of Marine Science and Engineering. 2025; 13(5):852. https://doi.org/10.3390/jmse13050852
Chicago/Turabian StyleLüskow, Florian, Philipp Neitzel, Elizabeth R. Lawrence, and Lina M. Nordlund. 2025. "Jellyfish and Ctenophores Around Gotland in the Baltic Sea—Local Data Contributing to Global Assessments" Journal of Marine Science and Engineering 13, no. 5: 852. https://doi.org/10.3390/jmse13050852
APA StyleLüskow, F., Neitzel, P., Lawrence, E. R., & Nordlund, L. M. (2025). Jellyfish and Ctenophores Around Gotland in the Baltic Sea—Local Data Contributing to Global Assessments. Journal of Marine Science and Engineering, 13(5), 852. https://doi.org/10.3390/jmse13050852