Abstract
Teleost Immunoglobulin M (IgM) is a crucial molecule in the adaptive immune response in teleost fish. Therefore, specific antibodies against IgM are essential tools for studying immune responses and evaluating vaccine efficacy across various teleost species. Producing antibodies using synthetic peptide antigens is a convenient approach when the IgM sequence is known. However, anti-peptide antibodies often fail to recognize natural antigens in either their denatured or native form, limiting their utility for antigen detection. In the present study, we identified a highly antigenic amino acid sequence stretch between the CH3 and CH4 domains of the IgM heavy chain. Using medaka fish (Oryzias latipes) as a model, we generated anti-peptide antibodies and tested their specificity and reactivity against native serum IgM. Based on in silico analysis, we hypothesize that this antigenic site is also applicable to other teleost species. Using this antibody, we successfully assessed systemic and mucosal antibody response following medaka exposure to bovine serum albumin-coated microplastics via immersion.
1. Introduction
With the global expansion of aquaculture, the diversity of farmed fish species has increased, leading to a growing need for species-specific disease control measures. Analyzing antibody production response is particularly critical for vaccine development and monitoring pathogen infections. Therefore, the development of primary antibodies that can specifically recognize the immunoglobulins of each fish species is essential for the development of reliable immunoassays [1,2]. However, preparing anti-immunoglobulin antibodies for each fish species is a labor-intensive process, often requiring extensive immunogen purification. With the availability of whole genome sequences for many teleost species, the amino acid sequences of immunoglobulins can now be easily retrieved from public databases [3]. Antipeptide antibodies derived from these sequences offer a promising and efficient method of generating species-specific primary antibodies. However, these antibodies frequently face challenges such as low specificity, limited reactivity, and poor recognition of native antigenic proteins [4,5,6].
Medaka (Oryzias latipes), a small laboratory fish, has been widely recognized as a useful model organism in fields such as genetics, embryology, environmental toxicology, and physiology [7,8]. In recent years, medaka has also been increasingly used as a model for studying immune responses. However, immunological tools, such as anti-immunoglobulin antibodies essential for analyzing antibody production, remain underdeveloped in this species [9,10]. We are particularly focused on investigating the immunomodulatory role of microplastics on medaka’s antibody production, as microplastics have emerged as a significant environmental pollutant impacting fish physiology [11].
In this study, we developed an anti-medaka IgM antibody as an anti-peptide antibody to serve as a fundamental tool for detecting antibody production in medaka. Its specificity and reactivity were analyzed, resulting in a primary antibody with excellent specificity and reactivity. This was achieved using a peptide antigen corresponding to the linker region between the CH3 and CH4 domains of the medaka IgM heavy chain. Furthermore, we also successfully analyzed the effect of microplastic exposure on the antibody production response in medaka through immunoassay.
To extend the methodology of producing highly specific and reactive anti-IgM antibodies using peptide antigens to other fish species, we analyzed the three-dimensional structure of the corresponding region in representative teleost species. The results suggest that this region is suitable as an antigen for generating anti-peptide across various fish species.
2. Materials and Methods
2.1. Fish
The Japanese medaka (48.1 ± 1.8 mg) (Oryzias latipes; orange–red strain) was provided by the Laboratory of Marine Environmental Science, Kyushu University (Fukuoka, Japan). Medaka were fed with Artemia nauplii and were maintained at a temperature of 25 ± 1 °C in artificial seawater (at a salinity of 0.01%) with an aerating filter system under a photoperiod condition of a 14:10 h light–dark cycle.
2.2. Peptide Sequence and Polyclonal Antiserum Production
The full-length amino acid sequence for the medaka IgM heavy (H)-chain (XP_023813427.1) was retrieved from the NCBI database (accessed on 1 April 2020). The peptide sequence between the CH3 and CH4 regions (IQYKRQNSGPAQR), which is likely at the surface protein and is not an alpha helix or beta sheet, was selected based on the prediction from the Bepipred Linear Epitope Prediction Tool 2.0. Multiple sequence alignment with six other teleost species, Cyprinus carpio (QDP16243.1), Danio rerio (AAT67447.1), Elops saurus (AAA49238.1), Siniperca chuatsi (AAQ14862.1), Ictalurus punctatus (AAA79003.1), and Gadus morhua (A46538), was performed using the ClustalW software (Ver. 1.83). The 3D structure of the IgM heavy chain of the teleost species was predicted using Alphafold 2. The antigenic peptide was synthesized with an N-terminal addition of Cys residue and conjugated with the carrier protein keyhole limpet hemocyanin (KLH). The KLH-peptide conjugate was injected into a rabbit (four doses at 14-day intervals) and the antiserum was collected 49 days after the initial dose.
2.3. Purification of Antiserum
Anti-medaka IgM was purified from the antiserum using affinity chromatography on an immobilized antigen peptide column, which was prepared by conjugating the Cys-containing peptide with Epoxy-Toyoperl (Tosoh Co., Tokyo, Japan) following the manufacturer’s instruction. The peptide Toyopearl was packed in a PD-10 column (Cytiva, Tokyo, Japan). To the column (1 mL) equilibrated with PBS (pH 7.4), 2 mL of antiserum was applied. After washing the column with PBS, bound antibodies were eluted with 0.1 M glycine-HCl (pH 2.5) into tubes containing 1/10 volume of 1 M Tris-HCl (pH 9.0). The antibody-rich fractions of the eluate were pooled, concentrated using an Amicon filter (10,000 MW cut-off, Merck, Tokyo, Japan), and stored at −80 °C in the presence of 0.02% sodium azide.
2.4. Size Exclusion Chromatography of Medaka Serum
Medaka serum (20 µL) was applied to a Superdex 200 Increase column (1 × 30 cm, Cytiva, Tokyo, Japan) equilibrated with 50 mM sodium phosphate buffer (pH 7.4) containing 0.15 M NaCl. Fractions were collected at a flow rate of 0.5 mL/min and subjected to immunoassays using an anti-medaka IgM antibody. The fractions rich in IgM were pooled and checked for purity using Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
2.5. SDS-PAGE and Western Blot
The sample was run on 10% polyacrylamide gels under reducing conditions, followed by staining with Coomassie Brilliant Blue. For western blotting, the separated proteins were transferred to a PVDF membrane at 60 V for 60 min in 0.1 M Tris-0.192 M glycine-5% methanol-0.02% SDS. The membrane was blocked in 5% (w/v) skimmed milk for 1 h at room temperature and then incubated with anti-medaka IgM (10 µg/mL) for 1 h. After thorough washing with 0.5% Tween 20-PBS (TPBS), the membrane was further treated with 1/5000-diluted HRP-conjugated anti-rabbit IgG (Medical & Biological Laboratories Co., Ltd., Tokyo, Japan) for 1 h, washed with TPBS, and developed using a TMB substrate (EzWest Blue, ATTO, Tokyo, Japan).
2.6. Dot Blot Analysis
Dot blot analysis was performed with the IgM fractions and the purified anti-medaka IgM. Briefly, 10 µL of the sample was dotted onto a nitrocellulose membrane. The membrane was blocked in 0.5% skimmed milk in PBS and treated with anti-medaka IgM (10 µg/mL), the secondary antibody, and HRP-substrate, essentially as described above.
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
The sample was diluted in 50 mM sodium bicarbonate buffer (pH 9.6). A total of 100 µL of this solution was added to a 96-well microtiter plate and incubated overnight at 4 °C. The coated plate was blocked with 0.5% BSA-0.2% gelatin-0.1% thimerosal-PBS (pH 7.4) and incubated at room temperature for 1 h. The plate was further treated with 10 µg/mL anti-medaka IgM, and then with a secondary antibody, as above. The plate was then developed using TMB microwell peroxidase substrate (KPL, Sera care, Milford, MA, USA). The absorbance at 450 nm was measured using a Multiskan FC Microplate Spectrophotometer (ThermoFisher Scientific, Waltham, MA, USA). All measurements were performed in triplicates.
2.8. Adsorption of BSA to Microplastics
Bovine serum albumin (100 µg/mL, Sigma, Tokyo, Japan) was incubated with 1 mg of 1 µm polystyrene spherical microplastics (MP, Thermo Fisher Scientific, G0100) in an acetate buffer containing 0.03 M sodium acetate and 0.02 M acetic acid adjusted to pH 4.5 at 4 °C overnight. The particles were washed in 0.1 M sodium phosphate buffer (pH 7.4) by centrifuging at 10,000 rpm for 10 min and resuspended at 0.1 M PBS. The amount of BSA adsorbed on the microplastics was estimated by measuring BSA concentration in the supernatant using a Micro BCA protein assay kit (Pierce, Tokyo, Japan). The particles were run on SDS-PAGE to further confirm the adsorption of BSA on the microplastics.
2.9. Exposure Experiments
Medaka fish were divided into five groups (10 fish/group). Three groups were exposed to MP (1 mg), Microplastics-BSA (MP-BSA) (1 mg), and BSA (50 µg), respectively. Fish were transferred to a fresh dose of antigen every 14 days. The positive control group fish received intraperitoneal injection of BSA (20 µg) and the negative control group was kept with no exposure or injection. Serum and gut mucus were collected every 14 days from three fish of each group. The protein concentration of the gut mucus sample was measured using the BCA protein Assay Kit (Pierce). ELISA using anti-medaka IgM was performed to monitor anti-BSA medaka IgM level in the serum and mucus.
2.10. Induction of Body Transparency and Fluorescent Microscopy
Medaka fish exposed to MP, MP-BSA, or BSA for 24 h underwent transparency treatment following a modified version of the method reported by Liu et al. [12]. Briefly, the fish were fixed in 10% formalin for 24 h and washed with several changes of water over the next 24 h. They were subsequently immersed in several changes of 50% ethanol for 24 h at 4 °C. The fish were then incubated in a solution containing 30 mL saturated borax, 70 mL water, and 0.3 g of trypsin (pH 8.0) at 40 °C with gentle rotation (50 rpm) for 48 h. After washing with water, the fish were sequentially transferred through mixtures of 0.5%-KOH and glycerol in ratios of 3:1, 1:1, and 1:3, incubating in each solution for 24 h. The transparent fish were stored in glycerol at 4 °C and observed using a fluorescent microscope (Model BZ-X810, KEYENCE, Osaka, Japan) equipped with a GFP filter (λem = 525/50 nm and λex = 470/40 nm).
2.11. Statistical Analysis
The quantitative data were expressed as mean ± standard deviation. Statistical significance is calculated based on a one-way ANOVA with post hoc Tukey HSD using GraphPad Ver. 10.4.0 software (Dotmatics, Boston, MA, USA).
3. Results
3.1. Specificity and Reactivity of Anti-Medaka IgM Anti-Peptide Antibody
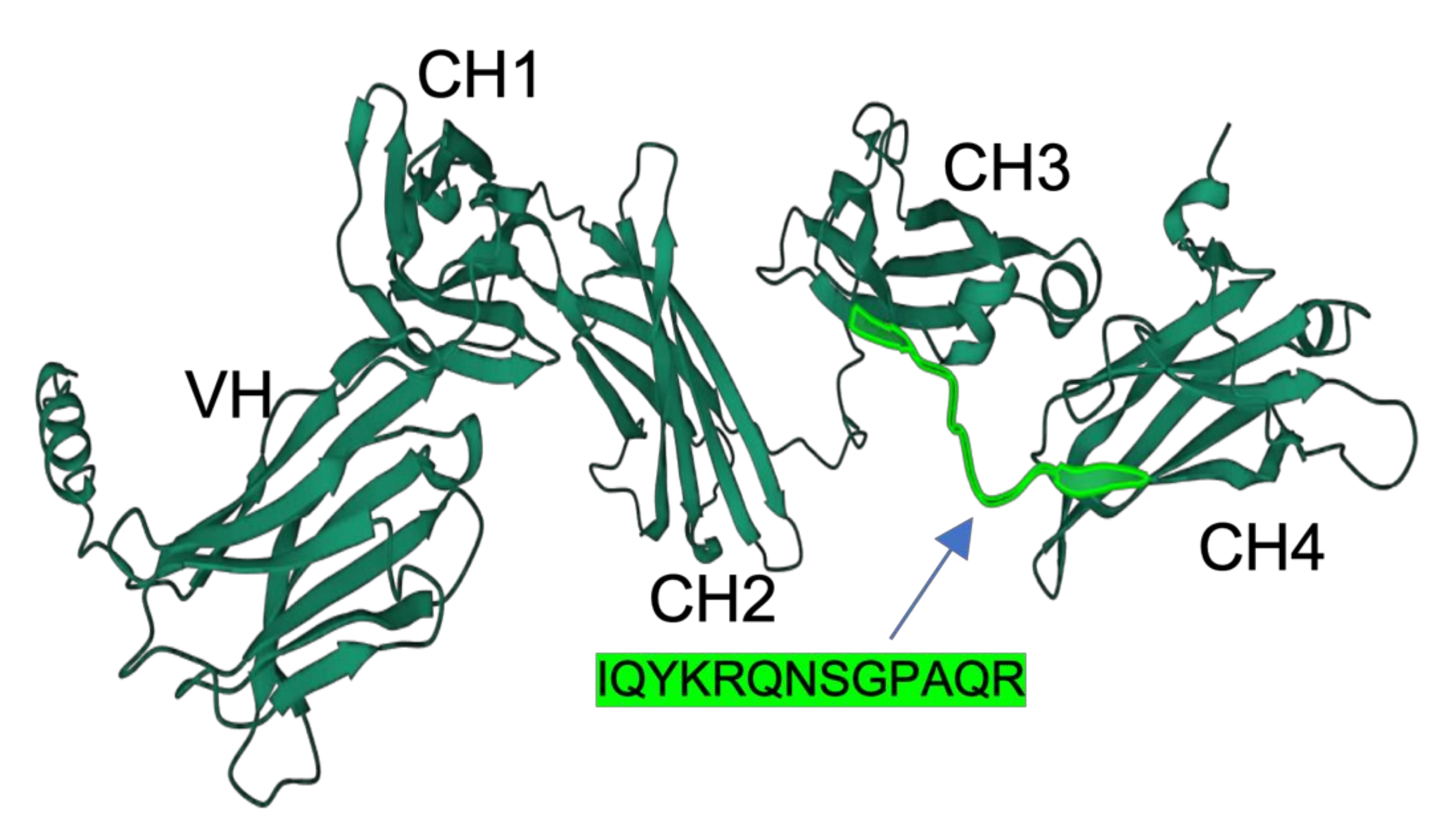
The full-length primary structure of IgM heavy-chain of medaka was retrieved from NCBI (XP_023813428.1). The EST sequence of CH1, CH2, CH3, and CH4 of medaka IgM had previously been reported by Magadan-Mompo et al. [2]. In the present study, we selected the amino acid sequence IQYKRQNSGPAQR as an antigenic peptide based on predictions from the Bepipred Linear Epitope Prediction Tool 2.0 [13]. A similar approach was employed by Velazquez et al., who developed a monoclonal antibody against the IgM heavy-chain Oreochromis niloticus using an 11-residue peptide identified through epitope prediction tools and BLAST searches. Their selected peptide was hydrophilic, solvent-accessible, and flexible [4]. The peptide used in our study shared similar characteristics, with Bepipred prediction indicating high hydrophilicity, antigenicity, and surface accessibility. Additionally, structural modeling performed using AlphaFold 2 revealed that the peptide sequence resides within loop regions of the CH3-CH4 domains (Figure 1). AlphaFold, a novel machine-learning tool, accurately predicts protein structure with atomic-level precision [14]. Loop regions are particularly important because they exhibit greater solvent accessibility compared to alpha helices or beta sheets, increasing their chance of interacting with biomolecules [15].
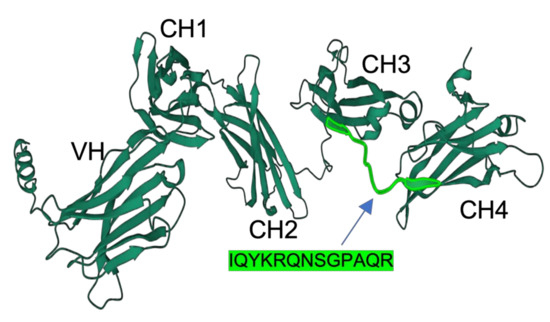
Figure 1.
Three-dimensional structure of medaka IgM heavy chains predicted by Alphafold2 software (Ver. 2.0). Domain names (VH, CH1, CH2, CH3, and CH4) are inserted. The position and sequence of the antigenic peptide are highlighted in light green.
Antipeptide antibodies are relatively easy to produce, but they often face challenges with specificity and reactivity against native protein antigens. While many studies have used purified IgG from rabbit antiserum containing antipeptide antibodies, this study employed affinity purification using immobilized antigen peptides to enhance antibody specificity and titer. To achieve this, a column was prepared by immobilizing the antigen peptide on Epoxy-Toyopearl resin via the SH group of a cysteine residue that was artificially added to the peptide’s N-terminus.
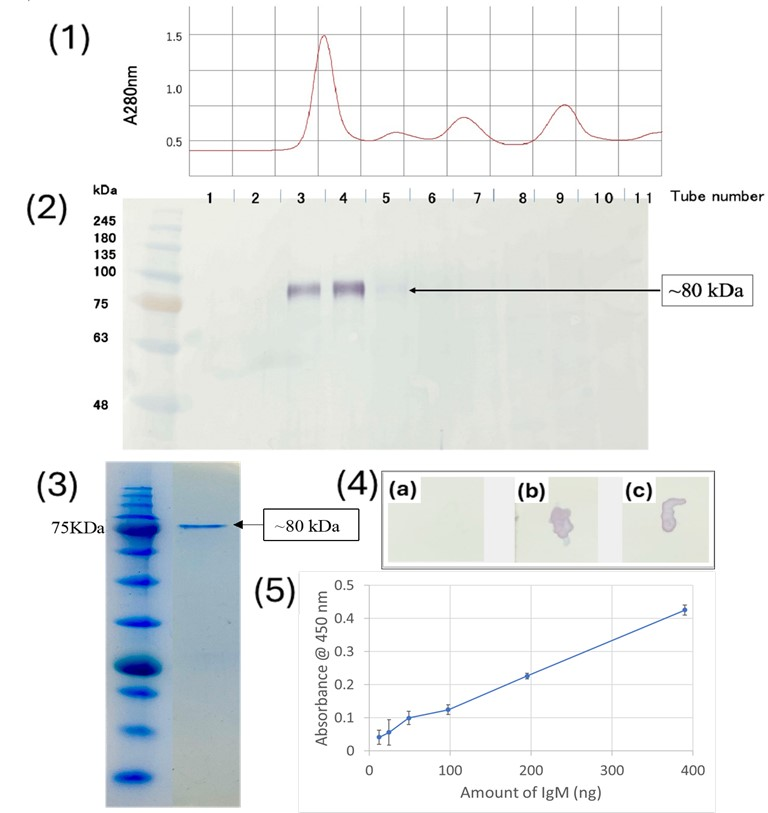
To test the specificity of the anti-medaka IgM, western blot analysis was performed using medaka serum proteins fractionated by size exclusion chromatography as antigens. As shown in Figure 2(1,2), the anti-medaka IgM detected an 80 kDa single band in the fractions near the void volume of the Superdex 200 column, indicating the 80 kDa band represents the H-chain of medaka IgM since the molecular mass of teleost IgM has previously been reported to be approximately 75 kDa [16,17]. Medaka IgM purified by the size-exclusion chromatography was analyzed by SDS-PAGE, showing the 80 kDa band of H-chain, while a light-chain band with a predicted molecular mass of ~25 kDa was not visible, probably due to the relatively low protein concentration (Figure 2(3)). It is noteworthy that the dot blot performed against native antigens, the purified IgM, and medaka serum showed an intense signal of the anti-peptide antibody binding (Figure 2(4)). The sensitivity of the anti-medaka IgM antibody to detect medaka IgM by ELISA was also tested by using a 96-well plate coated with a serially diluted purified IgM. As shown in Figure 2(5), the anti-medaka IgM antibody gave a significant signal of medaka IgM as low as 12.2 ng protein. This sensitivity is comparable to that reported for ELISA using the anti-tilapia IgM antibody (~20 ng/mL) [4].
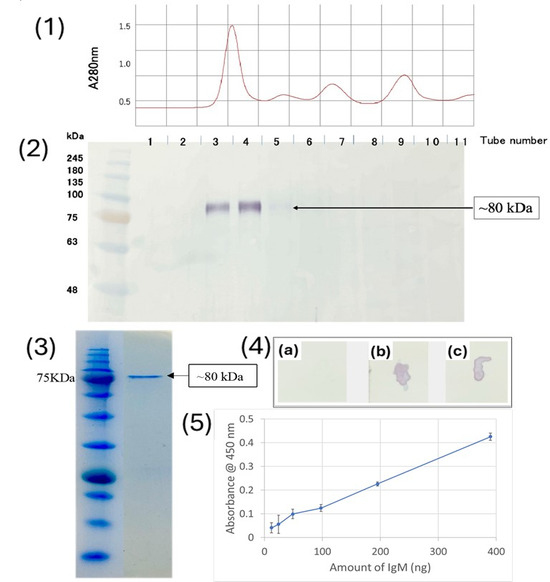
Figure 2.
(1) Chromatogram showing the purification of medaka IgM; (2) western blot to check the affinity of anti-medaka IgM antibody against the serum fractions; (3) SDS PAGE showing the purified medaka IgM; (4) Dotblot analysis to check the affinity of anti-medaka IgM antibody against (a) PBS, (b) purified medaka IgM, (c) medaka serum; (5) ELISA to check the sensitivity of anti-medaka IgM.
3.2. Prediction of Antigenic Sequences in Another Teleost IgM
Based on the strong reactivity of the anti-medaka IgM antibody raised against the linker region between the CH3 and CH4 domains, we hypothesized that this region may serve as an effective antigen for generating versatile anti-peptide antibodies. Such antibodies could potentially recognize both native and denatured IgM proteins in various teleost species.
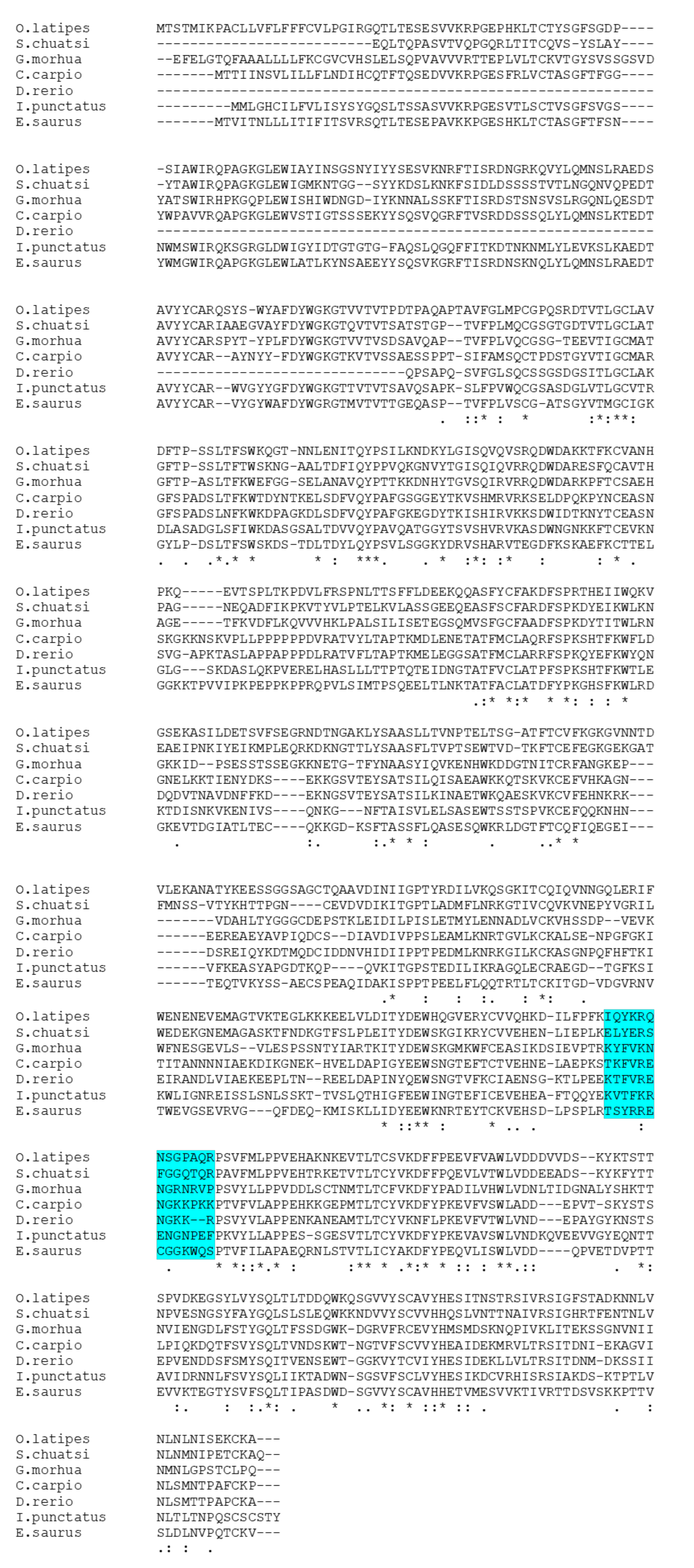
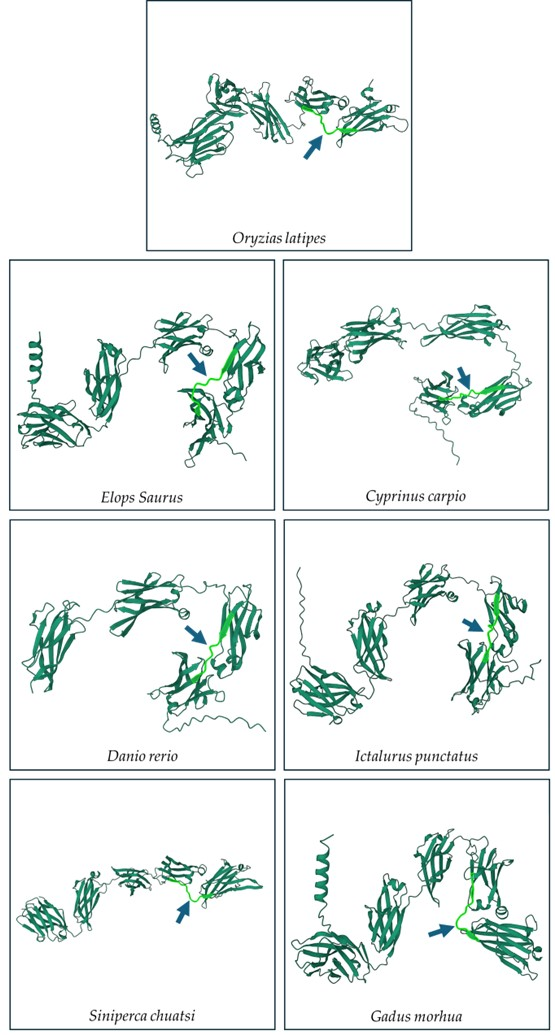
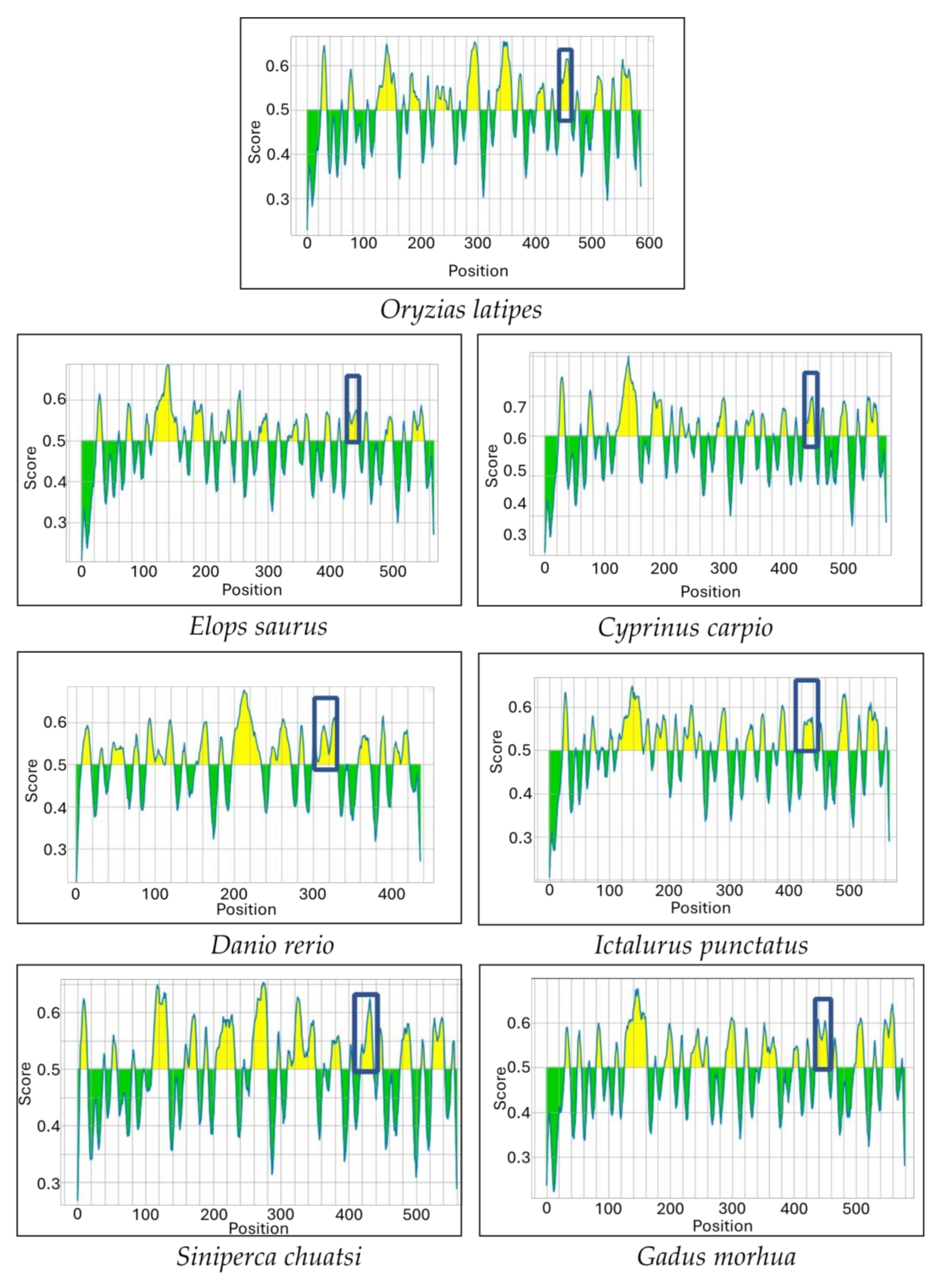
Multiple sequence alignments of the IgM heavy-chain sequences from several teleost species revealed significant divergence in the CH3-CH4 linker sequence (Figure 3). However, AlphaFold2 predicts that the linker region is a loop exposed on the molecular surface in all the species (Figure 4). Bepipred analysis further indicated that the peptide sequence is hydrophilic, antigenic, solvent accessible, and flexible. Collectively, these findings suggest that the CH3-CH4 linker region is a highly promising epitope for developing anti-IgM heavy-chain-specific anti-peptide antibodies applicable to a broad range of teleost species (Figure 5).
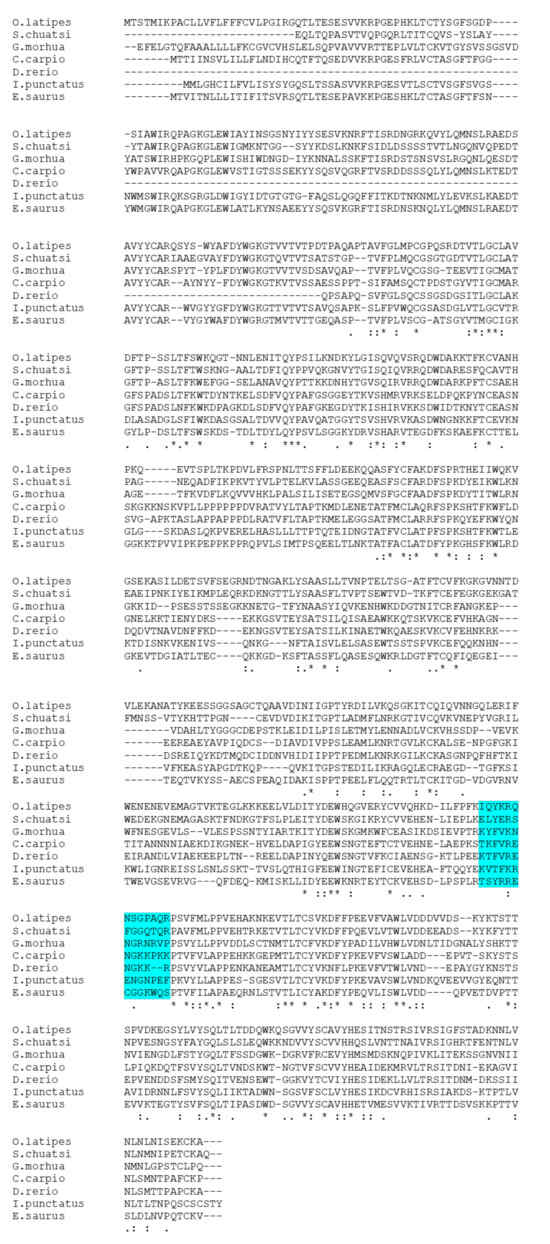
Figure 3.
Multiple sequence analysis of IgM between seven different teleost species. The highlighted area shows amino acid sequences aligned with six teleost fish species with the medaka peptide chosen for raising anti-medaka IgM antibodies. “*” indicates the residues are identical in all sequences in the alignment, “:” indicates that conserved substitutions have been observed, and “.” indicates that semi-conserved substitutions have been observed. Accession number of the sequences: Siniperca chuatsi (AAQ14862.1), Gadus morhua (A46538), Cyprinus carpio (QDP16243.1), Danio rerio (AAT67447.1), Ictalurus punctatus (AAA79003.1), and Elops saurus (AAA49238.1).
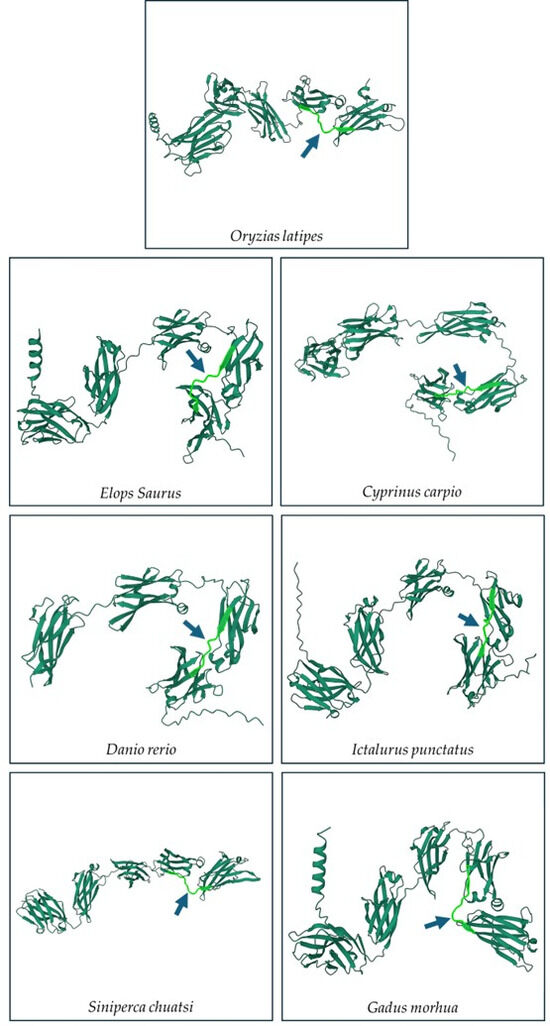
Figure 4.
Secondary structure prediction of IgM sequence of seven different teleost species using Alphafold 2. The light-green part with arrow indicates the region between the CH3 and CH4 domains.
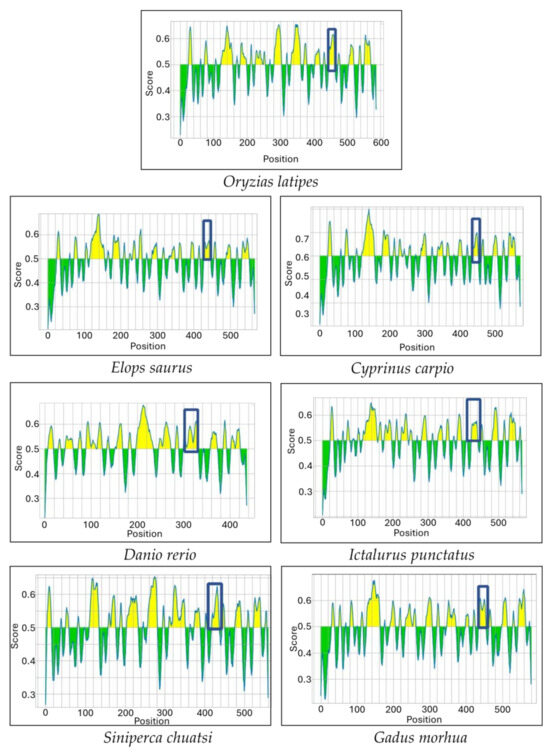
Figure 5.
Bepipred analysis shows the potential epitopes (Yellow colored area) in the IgM sequence of seven different teleost species. The boxed region corresponds to the linker between the CH3 and CH4 domains.
3.3. Application of Anti-Medaka IgM Anti-Peptide Antibody for Evaluation of Adjuvant Effect in Bath Immunization
3.3.1. Preparation of BSA-Coated Microplastic Beads
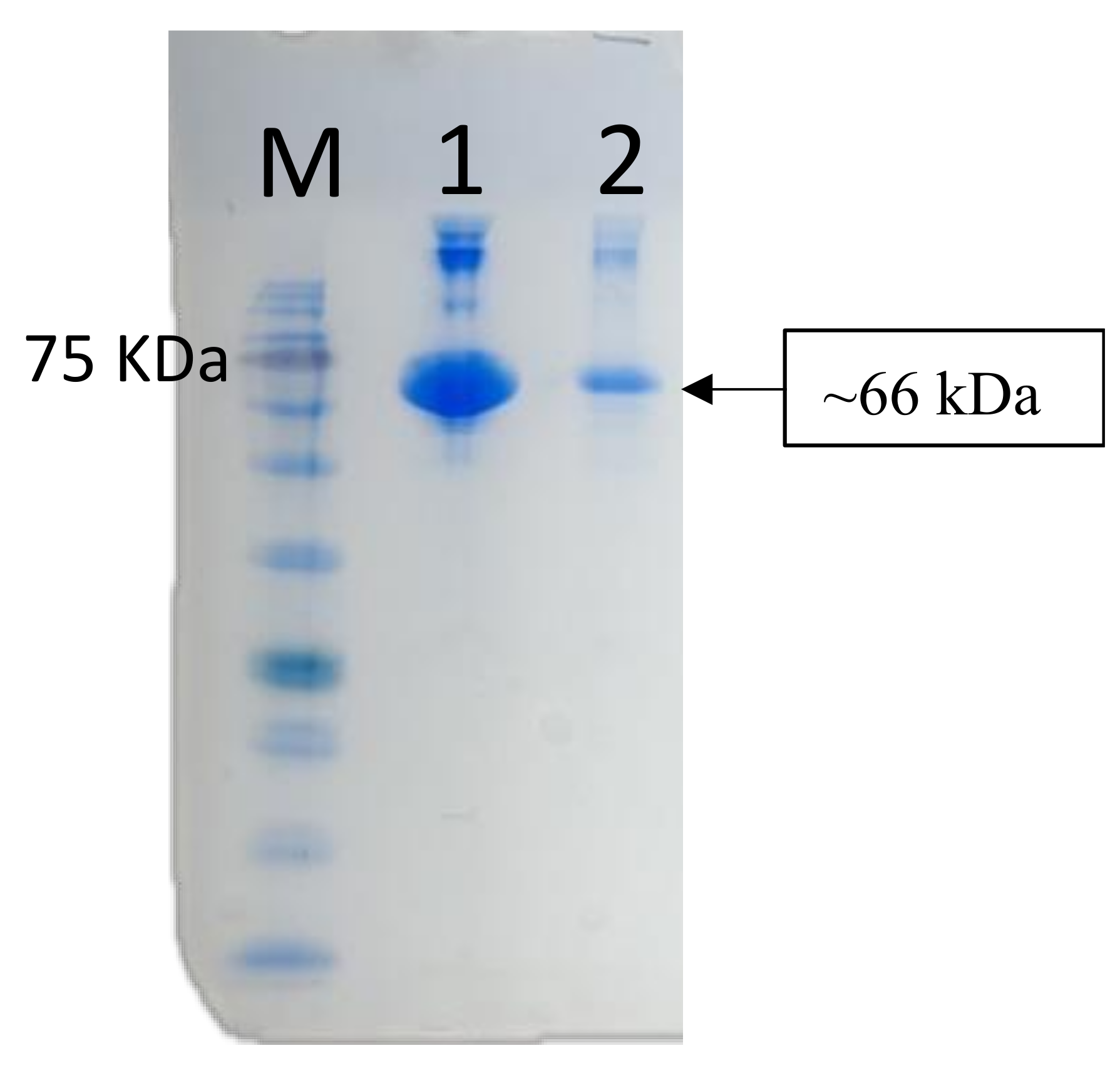
The anti-medaka IgM heavy-chain antibody was utilized to detect antibody response in medaka serum and gut mucus for evaluation of the adjuvant effect of microplastic beads in bath immunization. BSA was used as a model antigen adsorbed on the fluorescent polystyrene microparticles (MP). Actual BSA adsorption was confirmed by eluting the MP-bound BSA with SDS-sample buffer and following SDS-PAGE. As shown in Figure 6, a 66 kDa band of BSA was detected by SDS-PAGE. The amount of BSA adsorbed onto the MP was estimated to be 48 ± 6 µg/mg MP by measuring the protein content in the eluate.
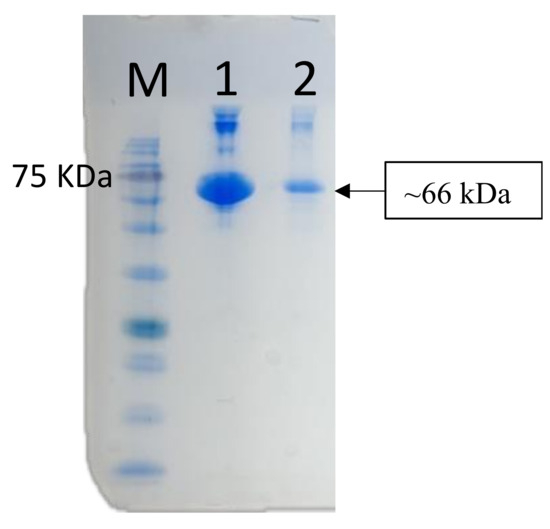
Figure 6.
SDS PAGE to confirm the adsorption of BSA onto the 1 µm microplastics. Lane M, marker proteins; Lane 1, BSA alone (positive control); Lane 2, eluate from MP.
3.3.2. Tracing MP in the Medaka Body After Bath Exposure
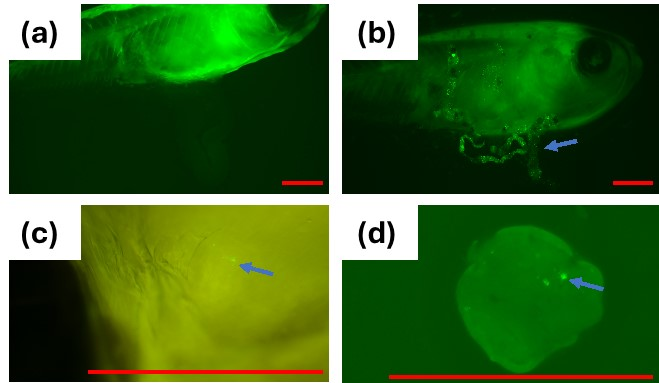
Medaka were exposed to the MP suspension (1 mg/mL) for 24 h. The distribution of MP in the fish was examined by fluorescent microscopy after making the body transparent. The fluorescent MP was detected in the intestine, gills, and liver of the medaka (Figure 7). Similar uptake of MP into various organs, including the liver, muscle, gills, and gonads, after oral administration has been reported by several studies [18,19,20]. Liu et al. exposed medaka with microplastics with a diameter from 2 to 200 µm and found that 2–20 µm beads were ingested into the gills, head, and gastrointestinal tract 24 h after the initial exposure [12]. Although the exact mechanism of the bead ingestion is unclear, phagocytic ingestion by intestine leukocytes may play a role, especially for the uptake of smaller-sized MPs, such as the 1 µm MPs used in the present study.
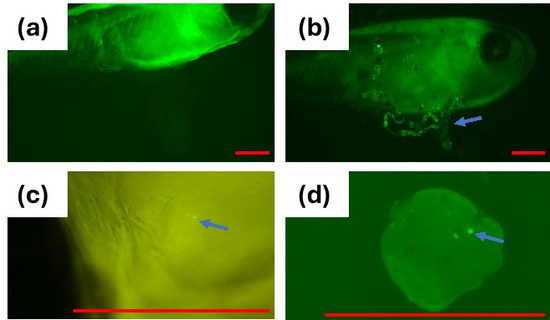
Figure 7.
Fluorescence microscopy of medaka after exposing fish to fluorescent polystyrene 1 µm MP; (a) control fish (2×); (b) MP in the gastrointestinal tract of fish (2×); (c) MP in the gills of the fish (8×); (d) MP in the liver (8×); Arrow shows MPs; Red Bar = 1 mm.
3.3.3. Bath Immunization Test for Antibody Response in Serum and Gut Mucus
The medaka were immunized by immersion in BSA solution or in BSA-coated MP suspension. Intraperitoneal injection of BSA served as a positive control of immunization. During the immunization period, no mortality was observed in all test groups. In addition, there were no differences in the weight gains among the experimental groups (Table 1).

Table 1.
Body weight of fish at 0th and 70th days (Mean ± SD).
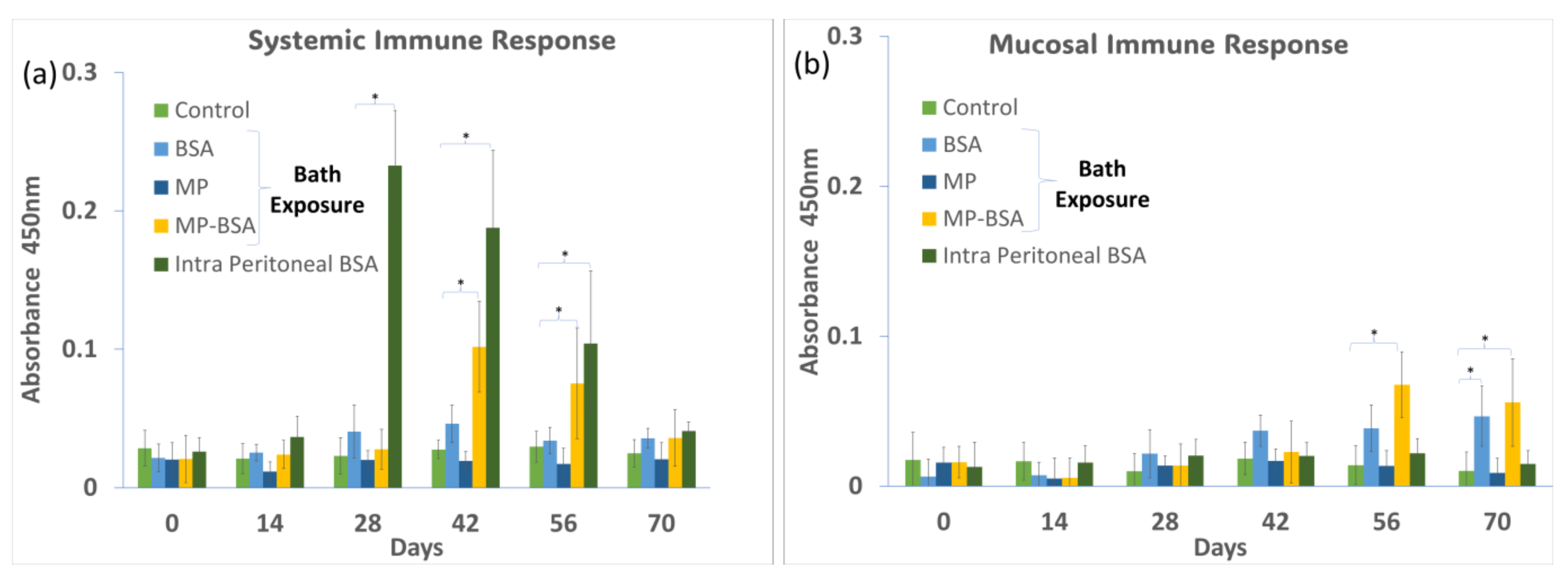
Anti-BSA antibody levels in the serum and gut mucus were measured at 2-week intervals during 70 days of the immunization protocol. The systemic immune response (represented by serum antibody levels) showed the fastest and highest increase by intraperitoneal injection (positive control), followed by the bath exposure group of BSA-coated MP (Figure 8a). On the other hand, the bath exposure in the BSA solution gave no significant increase in antibody levels compared to that of the negative control group. The local immune response (represented by antibody levels in mucus) was significantly increased by BSA-coated MP and BSA alone in the bath exposure, showing the highest values in the BSA-coated MP group (Figure 8b). The intraperitoneal injection of BSA did not increase the IgM levels in the mucus. Previous studies have shown that immersion vaccination in fish can stimulate both systemic and mucosal immune responses, while vaccination by injection induces only a systemic response [21].
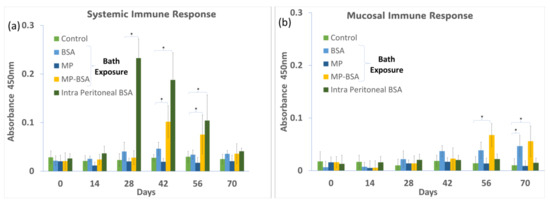
Figure 8.
(a) Systemic immune response when medaka was exposed to BSA, MP, and MP-BSA via bath exposure and intraperitoneal injection of BSA; (b) mucosal immune response when medaka was exposed to BSA, MP, and MP-BSA via bath exposure and intraperitoneal injection of BSA. “*” indicates a significant increase compared to control p < 0.05.
In teleost, it is known that intact proteins can be partially absorbed from the intestinal tract and transferred into the bloodstream, inducing a systemic immune response [22,23]. In the present study, slightly higher serum antibody levels were detected in the BSA-alone bath exposure group than in the negative control group, possibly due to the direct absorption of the antigenic protein. On the other hand, the BSA-MP group induced a significant antibody production response, suggesting that MPs stimulated the immune response more effectively than BSA alone by being taken up by phagocytes from the intestinal tract. Thus, the present results suggest MPs can play an adjuvant role in oral and immersion vaccines.
The mechanisms of the adjuvant effect of microplastic remain unclear, the microplastic may play the role of vector for antigen proteins by protecting the adsorbed proteins from proteolytic digestion in the digestive tracts and by enhancing the uptake of antigen protein by phagocytes. The teleost intestine is equipped with gut-associated lymphoid tissues, which harbor lymphocytes, macrophages, and other antigen-presenting cells [24]. MP has the ability to nonspecifically adsorb a variety of proteins [25]. Thus, proteins adsorbed on the surface of microplastics taken up from the intestine by oral exposure to microplastics may trigger an antibody-producing response.
4. Conclusions
This study successfully developed a highly reactive and specific anti-peptide antibody against medaka IgM by utilizing a peptide antigen corresponding to a linker region between the CH3 and CH4 domains of the IgM heavy chain. The antibody was found to have an excellent affinity for both denatured and native IgM. This study proposes that the region is promising for generating anti-peptide antibodies for other teleost species as well. Using the anti-medaka IgM antibody as a tool, we evaluated the adjuvant effect of microplastic beads in bath immunization against BSA, a model antigen protein, in medaka. The results suggested that MP can enhance both the systemic and mucosal antibody responses in bath immunization.
Author Contributions
Conceptualization, M.N., Y.O., T.S., T.N. and A.K.; methodology, A.K., I.M. and H.P.; validation, A.K., T.S., T.N. and M.N.; Laboratory works, A.K., I.M., H.P. and T.N.; data curation, T.N., T.S. and M.N.; writing—original draft preparation, A.K.; writing—review and editing, M.N., T.S. and A.K.; supervision, M.N., T.S. and T.N.; project administration, M.N.; funding acquisition, Y.O. and M.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded and supported by JSPS KAKENHI, grant numbers JP19H03050, JP23K23699, and JP22H03760.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors are grateful to Yuki Takai for technical assistance in the immunization experiment of the medaka.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| IgM | Immunoglobulin M |
| KLH | Keyhole Limpet hemocyanin |
| PBS | Phosphate buffered saline |
| TPBS | 0.5% Tween 20-Phosphate buffered saline |
| BSA | Bovine Serum Albumin |
| MP | Microplastics |
References
- Solem, S.T.; Stenvik, J. Antibody repertoire development in teleosts—A review with emphasis on salmonids and Gadus morhua L. Dev. Comp. Immunol. 2006, 30, 57–76. [Google Scholar] [PubMed]
- Jones, E.M.; Oliver, L.P.; Ma, J.; Leeuwis, R.H.J.; Myrsell, V.; Arkoosh, M.R.; Dietrich, J.P.; Schuster, C.M.; Hawkyard, M.; Gamperl, A.K.; et al. Production of a monoclonal antibody specific to sablefish (Anoplopoma fimbria) IgM and its application in ELISA, western blotting, and immunofluorescent staining. Fish. Shellfish Immunol. 2022, 130, 479–489. [Google Scholar] [PubMed]
- Volff, J.N. Genome evolution and biodiversity in teleost fish. Heredity 2005, 94, 280–294. [Google Scholar]
- Velázquez, J.; Rodríguez, A.; Aragón, H.; Haidar, A.; González, M.; Valdés, R.; Garay, H.E.; Abreu, D.D.; Ramos, Y.; Cabrales, A.; et al. Monoclonal antibody against Nile tilapia (Oreochromis niloticus) IgM heavy chain: A valuable tool for detection and quantification of IgM and IgM+ cells. Fish. Shellfish Immunol. 2021, 110, 44–54. [Google Scholar]
- Trier, N.H.; Hansen, P.R.; Houen, G. Production and characterization of peptide antibodies. Methods 2012, 56, 136–144. [Google Scholar] [PubMed]
- Jirapongpairoj, W.; Hirono, I.; Kondo, H. Development and evaluation of polyclonal antisera for detection of the IgM heavy chain of multiple fish species. J. Immunol. Methods 2017, 449, 71–75. [Google Scholar]
- Ozato, K.; Wakamatsu, Y. Developmental Genetics of Medaka: (medaka/mutants/transgenic/chimeras/pluripotent cell line). Dev. Growth Differ. 1994, 36, 437–443. [Google Scholar]
- Chen, X.; Li, L.; Cheng, J.; Chan, L.L.; Wang, D.Z.; Wang, K.J.; Baker, M.E.; Hardiman, G.; Schlenk, D.; Cheng, S.H. Molecular staging of marine medaka: A model organism for marine ecotoxicity study. Mar. Pollut. Bull. 2011, 63, 309–317. [Google Scholar]
- Sakaguchi, H.; Sato, Y.; Matsumoto, R.; Gomikawa, J.; Yoshida, N.; Suzuki, T.; Matsuda, M.; Iwanami, N. Maturation of the medaka immune system depends on reciprocal interactions between the microbiota and the intestinal tract. Front. Immunol. 2023, 14, 1259519. [Google Scholar]
- Jun, B.; Ling, C.; Jia-He, X.; Ke-Jian, W.; Doris, W.T.A. The marine medaka Oryzias melastigma—A potential marine fish model for innate immune study. Mar. Pollut. Bull. 2011, 63, 267–276. [Google Scholar]
- Alberghini, L.; Truant, A.; Santonicola, S.; Colavita, G.; Giaccone, V. Microplastics in Fish and Fishery Products and Risks for Human Health: A Review. Int. J. Environ. Res. Public Health 2022, 20, 789. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qiu, X.; Xu, X.; Takai, Y.; Ogawa, H.; Shimasaki, Y.; Oshima, Y. Uptake and depuration kinetics of microplastics with different polymer types and particle sizes in Japanese medaka (Oryzias latipes). Ecotoxicol. Environ. Saf. 2021, 212, 112007. [Google Scholar] [PubMed]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, 24–29. [Google Scholar]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar]
- Choi, Y.; Agarwal, S.; Deane, C.M. How long is a piece of loop? PeerJ 2013, 1, e1. [Google Scholar]
- Elcombe, B.M.; Chang, R.J.; Taves, C.J.; Winkelhake, J.L. Evolution of antibody structure and effector functions: Comparative hemolytic activities of monomeric and tetrameric IgM from rainbow trout, Salmo gairdnerii. Comp. Biochem. Physiol. B 1985, 80, 697–706. [Google Scholar]
- Zhang, Y.A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 9, 827–835. [Google Scholar]
- Xiao, K.; Song, L.; Li, Y.; Li, C.; Zhang, S. Dietary intake of microplastics impairs digestive performance, induces hepatic dysfunction, and shortens lifespan in the annual fish Nothobranchius guentheri. Biogerontology 2023, 24, 207–223. [Google Scholar]
- Qiang, L.; Cheng, J. Exposure to polystyrene microplastics impairs gonads of zebrafish (Danio rerio). Chemosphere 2021, 263, 128161. [Google Scholar]
- Buwono, N.R.; Risjani, Y.; Soegianto, A. Oxidative stress responses of microplastic-contaminated Gambusia affinis obtained from the Brantas River in East Java, Indonesia. Chemosphere 2022, 293, 133543. [Google Scholar]
- Wei, G.K.; Da, C.Q.; Qing, J.M.; Zhao, R.D.; Yang, Z.L.; Tao, S.A.; Zhen, X. Mucosal immune responses and protective efficacy in yellow catfish after immersion vaccination with bivalent inactivated Aeromonas veronii and Edwardsiella ictaluri vaccine. Water Biol. Secur. 2022, 1, 100032. [Google Scholar]
- Marie, F.S.; Jean-Marie, V. Intestinal absorption of protein in teleost fish. Comp. Biochem. Physiol. A 1992, 103, 771–781. [Google Scholar]
- Mutoloki, S.; Munang’andu, H.M.; Evensen, Ø. Oral Vaccination of Fish—Antigen Preparations, Uptake, and Immune Induction. Front. Immunol. 2015, 6, 519. [Google Scholar] [CrossRef]
- Salinas, I. The Mucosal Immune System of Teleost Fish. Biology 2015, 4, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Dawson, A.L.; Bose, U.; Ni, D. Unravelling protein corona formation on pristine and leached microplastics. Micropl. Nanopl. 2024, 4, 9. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).