
Sessile Biofouling on Electrolytic Carbonated Structures: Stages of Colonization and Succession


 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling
2.3. Data Treatment
3. Results
3.1. Taxonomic Study
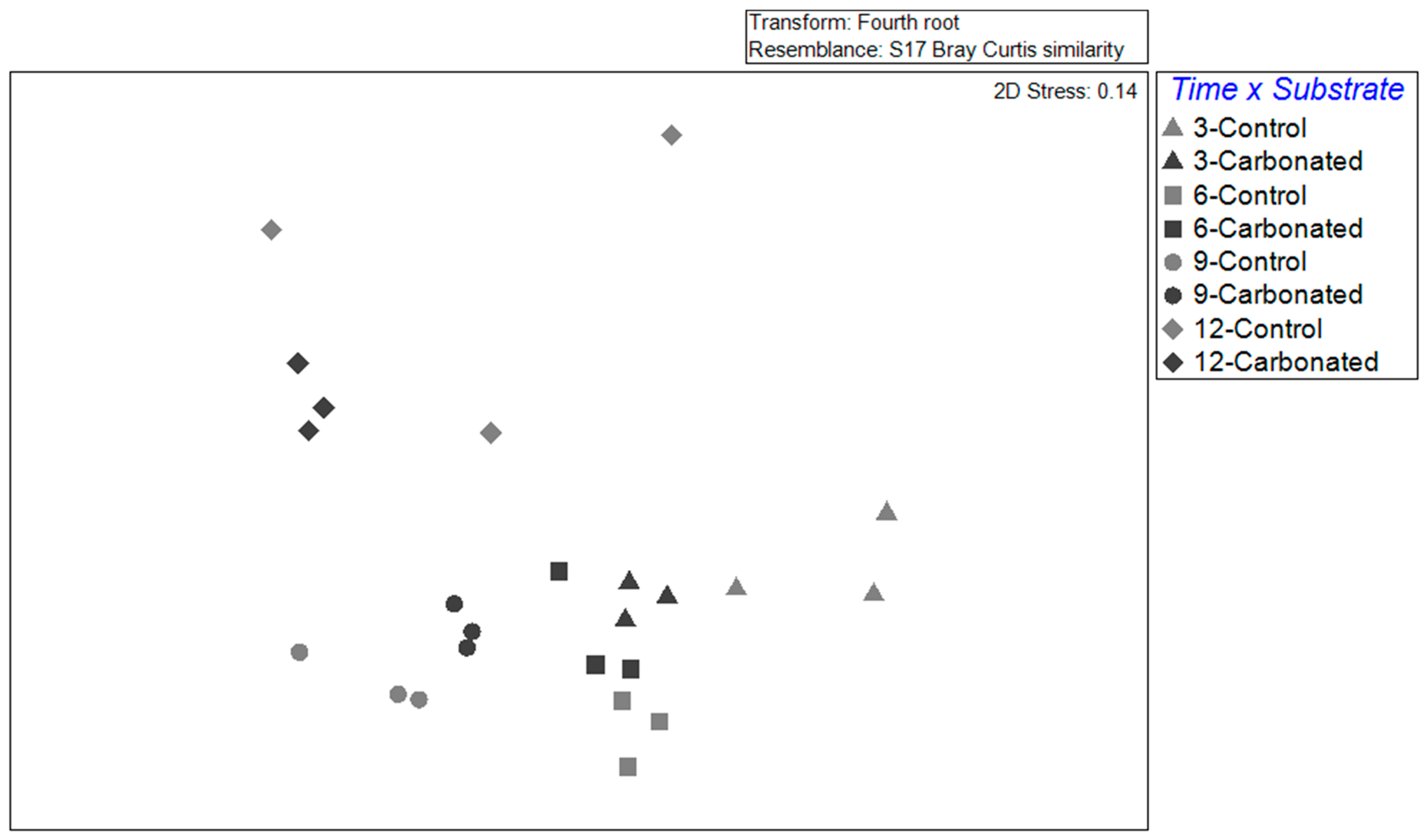
3.2. Biofouling Assemblages
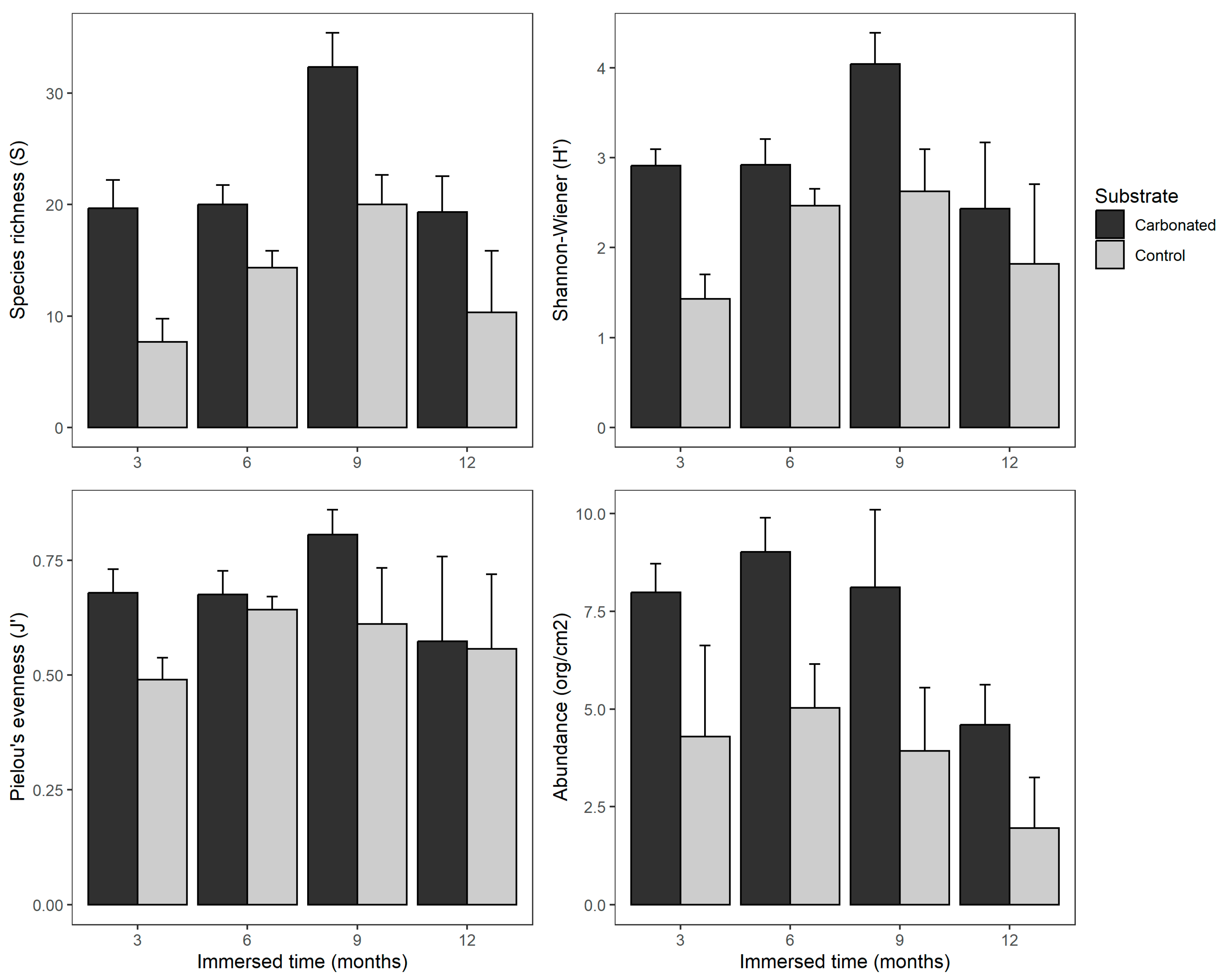
3.3. Biofouling Diversity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenkins, S.R.; Martins, G.M. Succession on Hard Substrata. In Biofouling; Simone, D., Jeremy, C.T., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2010; pp. 60–72. [Google Scholar]
- Anderson, M.J.; Underwood, A.J. Effects of Substratum on the Recruitment and Development of an Intertidal Estuarine Fouling Assemblage. J. Exp. Mar. Biol. Ecol. 1994, 184, 217–236. [Google Scholar] [CrossRef]
- Brown, K.M.; Swearingen, D.C. Effects of Seasonality, Length of Immersion, Locality and Predation on an Intertidal Fouling Assemblage in the Northern Gulf of Mexico. J. Exp. Mar. Biol. Ecol. 1998, 225, 107–121. [Google Scholar] [CrossRef]
- Petraitis, P.S.; Dudgeon, S.R. Experimental Evidence for the Origin of Alternative Communities on Rocky Intertidal Shores. Oikos 1999, 84, 239. [Google Scholar] [CrossRef]
- Lin, H.-J.; Shao, K.-T. The Development of Subtidal Fouling Assemblages on Artificial Structures in Keelung Harbor, Northern Taiwan. Zool. Stud. 2002, 41, 170–182. [Google Scholar]
- Lezzi, M.; Giangrande, A. Seasonal and Bathymetric Effects on Macrofouling Invertebrates’ Primary Succession in a Mediterraenan Non-Indigenous Species Hotspot Area. Mediterr. Mar. Sci. 2018, 19, 572–588. [Google Scholar] [CrossRef]
- Muthukrishnan, T.; Hassenrück, C.; Al Fahdi, D.; Jose, L.; Al Senafi, F.; Mahmoud, H.; Abed, R.M.M. Monthly Succession of Biofouling Communities and Corresponding Inter-Taxa Associations in the North- and South-West of the Arabian Gulf. Front. Mar. Sci. 2022, 8, 787879. [Google Scholar] [CrossRef]
- Bulleri, F. Role of Recruitment in Causing Differences between Intertidal Assemblages on Seawalls and Rocky Shores. Mar. Ecol. Prog. Ser. 2005, 287, 53–65. [Google Scholar] [CrossRef]
- Pierri, C.; Longo, C.; Giangrande, A. Variability of Fouling Communities in the Mar Piccolo of Taranto (Northern Ionian Sea, Mediterranean Sea). J. Mar. Biol. Assoc. 2010, 90, 159–167. [Google Scholar] [CrossRef]
- Megina, C.; González-Duarte, M.M.; López-González, P.J.; Piraino, S. Harbours as Marine Habitats: Hydroid Assemblages on Sea-Walls Compared with Natural Habitats. Mar. Biol. 2013, 160, 371–381. [Google Scholar] [CrossRef]
- Sutherland, J.P.; Karlson, R.H. Development and Stability of the Fouling Community at Beaufort, North Carolina. Ecol. Monogr. 1977, 47, 425–446. [Google Scholar] [CrossRef]
- Benedetti-Cecchi, L. Predicting Direct and Indirect Interactions during Succession in a Mid-Littoral Rocky Shore Assemblage. Ecol. Monogr. 2000, 70, 45–72. [Google Scholar] [CrossRef]
- Lu, L.; Wu, R.S.S. Seasonal Effects on Recolonization of Macrobenthos in Defaunated Sediment: A Series of Field Experiments. J. Exp. Mar. Biol. Ecol. 2007, 351, 199–210. [Google Scholar] [CrossRef]
- Cifuentes, M.; Krueger, I.; Dumont, C.P.; Lenz, M.; Thiel, M. Does Primary Colonization or Community Structure Determine the Succession of Fouling Communities? J. Exp. Mar. Biol. Ecol. 2010, 395, 10–20. [Google Scholar] [CrossRef]
- Lezzi, M.; Del Pasqua, M.; Pierri, C.; Giangrande, A. Seasonal Non-Indigenous Species Succession in a Marine Macrofouling Invertebrate Community. Biol. Invasions 2018, 20, 937–961. [Google Scholar] [CrossRef]
- Vaz-Pinto, F.; Torrontegi, O.; Prestes, A.C.L.; Álvaro, N.V.; Neto, A.I.; Martins, G.M. Invasion Success and Development of Benthic Assemblages: Effect of Timing, Duration of Submersion and Substrate Type. Mar. Environ. Res. 2014, 94, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Sempere-Valverde, J.; Ostalé-Valriberas, E.; Farfán, G.M.; Espinosa, F. Substratum Type Affects Recruitment and Development of Marine Assemblages over Artificial Substrata: A Case Study in the Alboran Sea. Estuar. Coast. Shelf Sci. 2018, 204, 56–65. [Google Scholar] [CrossRef]
- Firth, L.B.; White, F.J.; Schofield, M.; Hanley, M.E.; Burrows, M.T.; Thompson, R.C.; Skov, M.W.; Evans, A.J.; Moore, P.J.; Hawkins, S.J. Facing the Future: The Importance of Substratum Features for Ecological Engineering of Artificial Habitats in the Rocky Intertidal. Mar. Freshw. Res. 2016, 67, 131. [Google Scholar] [CrossRef]
- Brown, C.J. Epifaunal Colonization of the Loch Linnhe Artificial Reef: Influence of Substratum on Epifaunal Assemblage Structure. Biofouling 2005, 21, 73–85. [Google Scholar] [CrossRef]
- Fainburg, L.A.; Trassens, M.E.; Bastida, J.; Farenga, M.O.; Isla, F.I.; Bastida, R.O. Nearshore Benthic Communities and Bioengineers from the Macrotidal San Jorge Gulf: Patagonia, Argentina. Thalass. Int. J. Mar. Sci. 2012, 28, 45–56. [Google Scholar]
- Chase, A.L.; Dijkstra, J.A.; Harris, L.G. The Influence of Substrate Material on Ascidian Larval Settlement. Mar. Pollut. Bull. 2016, 106, 35–42. [Google Scholar] [CrossRef]
- Megina, C.; González-Duarte, M.M.; López-González, P.J. Biofouling the Journal of Bioadhesion and Biofilm Research Benthic Assemblages, Biodiversity and Invasiveness in Marinas and Commercial Harbours: An Investigation Using a Bioindicator Group. Biofouling 2016, 32, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Albano, M.J.; Obenat, S.M. Fouling Assemblages of Native, Non-Indigenous and Cryptogenic Species on Artificial Structures, Depths and Temporal Variation. J. Sea Res. 2019, 144, 1–15. [Google Scholar] [CrossRef]
- Bulleri, F.; Chapman, M.G.; Underwood, A.J. Intertidal Assemblages on Seawalls and Vertical Rocky Shores in Sydney Harbour, Australia. Austral. Ecol. 2005, 30, 655–667. [Google Scholar] [CrossRef]
- De Meester, L.; Vanoverbeke, J.; Kilsdonk, L.J.; Urban, M.C. Evolving Perspectives on Monopolization and Priority Effects. Trends Ecol. Evol. 2016, 31, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Ushiama, S.; Smith, J.A.; Suthers, I.M.; Lowry, M.; Johnston, E.L. The Effects of Substratum Material and Surface Orientation on the Developing Epibenthic Community on a Designed Artificial Reef. Biofouling 2016, 32, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Schuhmacher, H.; Schillak, L. Integrated Electrochemical and Biogenic Deposition of Hard Material—A Nature-like Colonization Substrate. Bull. Mar. Sci. 1994, 55, 672–679. [Google Scholar]
- Beisner, B.E.; Haydon, D.T.; Cuddington, K. Alternative Stable States in Ecology. Front. Ecol. Environ. 2003, 1, 343–396. [Google Scholar] [CrossRef]
- Walls, A.; Edwards, M.; Firth, L.; Johnson, M. Successional Changes of Epibiont Fouling Communities of the Cultivated Kelp Alaria Esculenta: Predictability and Influences. Aquac. Environ. Interact. 2017, 9, 57–71. [Google Scholar] [CrossRef]
- Obaza, A.K.; Williams, J.P. Spatial and Temporal Dynamics of the Overwater Structure Fouling Community in Southern California. Mar. Freshw. Res. 2018, 69, 1771–1783. [Google Scholar] [CrossRef]
- Siboni, N.; Martinez, S.; Abelson, A.; Sivan, A.; Kushmaro, A. Conditioning Film and Initial Biofilm Formation on Electrochemical CaCO3 Deposition on a Metallic Net in the Marine Environment. Biofouling 2009, 25, 675–683. [Google Scholar] [CrossRef]
- McManus, R.S.; Archibald, N.; Comber, S.; Knights, A.M.; Thompson, R.C.; Firth, L.B. Partial Replacement of Cement for Waste Aggregates in Concrete Coastal and Marine Infrastructure: A Foundation for Ecological Enhancement? Ecol. Eng. 2018, 120, 655–667. [Google Scholar] [CrossRef]
- Perkol-Finkel, S.; Hadary, T.; Rella, A.; Shirazi, R.; Sella, I. Seascape Architecture—Incorporating Ecological Considerations in Design of Coastal and Marine Infrastructure. Ecol. Eng. 2018, 120, 645–654. [Google Scholar] [CrossRef]
- Cox, G.C. Anticorrosive and Antifouling Coating and Method of Application. U.S. Patent No. 2,200,469, 14 May 1940. [Google Scholar]
- Eickhoff, A.J.; Shaw, W.E. Principles of Protecting Metals with Organic Coatings. Corrosion 1948, 4, 463–475. [Google Scholar] [CrossRef]
- Hilbertz, W.H. Electrodeposition of Minerals in Sea Water: Experiments and Applications. IEEE J. Ocean. Eng. 1979, 4, 94–113. [Google Scholar] [CrossRef]
- Hartt, W.H.; Culberson, C.H.; Smith, S.W. Calcareous Deposits on Metal Surfaces in Seawater—A Critical Review. Corrosion 1984, 40, 609–618. [Google Scholar] [CrossRef]
- Bakti, L.A.A.; Virgota, A.; Damayanti, L.P.A.; Radiman, T.H.U.; Retnowulan, A.; Sabil, A.; Robbe, D. Biorock Reef Restoration in Gili Trawangan, North Lombok, Indonesia. In Innovative Methods of Marine Ecosystem Restoration; Goreau, T.J., Trench, R.K., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 59–80. [Google Scholar]
- Fitri, D.; Rachman, M.A. Gorgonian Soft Corals Have Higher Growth and Survival in Electrical Fields. In Innovative Methods of Marine Ecosystem Restoration; Goreau, T.J., Trench, R.K., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 105–111. [Google Scholar]
- Goreau, T.J. Marine Electrolysis for Building Materials and Environmental Restoration. In Electrolysis; Linkov, V., Kleperis, J., Eds.; IntechOpen: London, UK, 2012; pp. 273–290. [Google Scholar]
- Goreau, T.J.; Hilbertz, W. Reef Restoration Using Seawater Electrolysis in Jamaica. In Innovative Methods of Marine Ecosystem Restoration; Goreau, T.J., Trench, R.K., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 35–45. [Google Scholar]
- Carmona-Rodríguez, A.; Antón, C.; Climent, M.Á.; Garcés, P.; Montiel, V.; Ramos-Esplá, A.A. Early Colonization of Sessile Megabenthos on Electrolytic Carbonated Structures (Alicante’s Harbor, Western Mediterranean). Sci. Total Environ. 2023, 900, 165796. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 1–15. [Google Scholar] [CrossRef]
- Clarke, K.; Gorley, R.; Somerfield, P.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; Primer-E Ltd.: Albany, New Zealand, 2014. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; Primer-E: Albany, New Zealand, 2008; pp. 1–224. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Boero, F. The Ecology of Marine Hydroids and Effects of Environmental Factors: A Review. Mar. Ecol. 1984, 5, 93–118. [Google Scholar] [CrossRef]
- Dean, T.A.; Hurd, L.E. Development in an Estuarine Fouling Community: The Influence of Early Colonists on Later Arrivals. Oecologia 1980, 46, 295–301. [Google Scholar] [CrossRef]
- Dean, T.A. Structural Aspects of Sessile Invertebrates as Organizing Forces in an Estuarine Fouling Community. J. Exp. Mar. Biol. Ecol. 1981, 53, 163–180. [Google Scholar] [CrossRef]
- Dijkstra, J.; Sherman, H.; Harris, L.G. The Role of Colonial Ascidians in Altering Biodiversity in Marine Fouling Communities. J. Exp. Mar. Biol. Ecol. 2007, 342, 169–171. [Google Scholar] [CrossRef]
- Aldred, N.; Clare, A.S. Mini-Review: Impact and Dynamics of Surface Fouling by Solitary and Compound Ascidians. Biofouling 2014, 30, 259–270. [Google Scholar] [CrossRef]
- Lara-Romero, C.; de la Cruz, M.; Escribano-Ávila, G.; García-Fernández, A.; Iriondo, J.M. What Causes Conspecific Plant Aggregation? Disentangling the Role of Dispersal, Habitat Heterogeneity and Plant–Plant Interactions. Oikos 2016, 125, 1304–1313. [Google Scholar] [CrossRef]
- Schimanski, K.B.; Goldstien, S.J.; Hopkins, G.A.; Atalah, J.; Floerl, O. Life History Stage and Vessel Voyage Profile Can Influence Shipping-Mediated Propagule Pressure of Non-Indigenous Biofouling Species. Biol. Invasions 2017, 19, 2089–2099. [Google Scholar] [CrossRef]
- Eashwar, M.; Ananth, V.; Paulraj, S.; Subramanian, G. Biofouling Studies Relating to Cathodic Protection of Some Metals in Seawater. Bull. Electrochem. 1985, 1, 19–21. [Google Scholar]
- Eashwar, M.; Subramanian, G.; Chandrasekaran, P.; Manickam, S.T.; Maruthamuthu, S.; Balakrishnan, K. The Interrelation of Cathodic Protection and Marine Macrofouling. Biofouling 1995, 8, 303–312. [Google Scholar] [CrossRef]
- Ramos-Esplá, A.A. Ascidias Litorales del Mediterráneo Ibérico: Faunística, Ecología y Biogeografía. Ph.D. Thesis, Universidad de Barcelona: Barcelona, Spain, 1988. [Google Scholar]
- Bae, S.; Ubagan, M.D.; Shin, S.; Kim, D.G. Comparison of Recruitment Patterns of Sessile Marine Invertebrates According to Substrate Characteristics. Int. J. Environ. Res. Public. Health 2022, 19, 1083. [Google Scholar] [CrossRef]
- Brümmer, F.; Nickel, M.; Sidri, M. Porifera (Esponjas). In El Mar Mediterráneo II/1: Guía Sistemática y de Identificación; Hofrichter, R., Ed.; Ediciones Omega, S.A.: Barcelona, Spain, 2005; pp. 302–383. [Google Scholar]
- Neves, C.S.; Rocha, R.M.; Pitombo, F.B.; Roper, J.J. Use of Artificial Substrata by Introduced and Cryptogenic Marine Species in Paranaguá Bay, Southern Brazil. Biofouling 2007, 23, 319–330. [Google Scholar] [CrossRef]
- Vieira, E.A.; Dias, G.M.; Flores, A.A.V. Adding Early-Stage Engineering Species Affects Advanced-Stage Organization of Shallow-Water Fouling Assemblages. Hydrobiologia 2018, 818, 211–222. [Google Scholar] [CrossRef]
- Oricchio, F.T.; Marques, A.C.; Hajdu, E.; Pitombo, F.B.; Azevedo, F.; Passos, F.D.; Vieira, L.M.; Stampar, S.N.; Rocha, R.M.; Dias, G.M. Exotic Species Dominate Marinas between the Two Most Populated Regions in the Southwestern Atlantic Ocean. Mar. Pollut. Bull. 2019, 146, 884–892. [Google Scholar] [CrossRef]
- Kuklinski, P.; Berge, J.; McFadden, L.; Dmoch, K.; Zajaczkowski, M.; Nygård, H.; Piwosz, K.; Tatarek, A. Seasonality of Occurrence and Recruitment of Arctic Marine Benthic Invertebrate Larvae in Relation to Environmental Variables. Polar Biol. 2013, 36, 549–560. [Google Scholar] [CrossRef]
- Lord, J.P. Temperature, Space Availability, and Species Assemblages Impact Competition in Global Fouling Communities. Biol. Invasions 2017, 19, 43–55. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | DF | MS | Pseudo-F | p-Value (MC) | Post Hoc Test |
|---|---|---|---|---|---|
| Substrate = S | 1 | 6513.1 | 9.2472 | *** | |
| Immersed time = T | 3 | 7517 | 10.673 | *** | |
| S × T | 3 | 1215.9 | 1.7263 | * | 3 and 9: C ≠ H 6 and 12: C ≈ H (p < 0.1) |
| Residual | 16 | 704.33 | |||
| Total | 23 |
| Source of Variation | DF | MS | F | p-Value | SNK | |
|---|---|---|---|---|---|---|
| S | Substrate = S | 1 | 570.4 | 62.507 | *** | C > I |
| Immersed time = T | 3 | 192.4 | 21.082 | *** | 9 > 3 = 6 = 12 | |
| S × T | 3 | 14.5 | 1.588 | 0.231 | ||
| Residuals | 16 | 9.1 | ||||
| H | Substrate | 1 | 5.894 | 24.935 | *** | C > I |
| Immersed time | 3 | 1.912 | 8.088 | ** | 9 > 3 = 6 = 12 | |
| S × T | 3 | 0.424 | 1.794 | 0.188788 | ||
| Residuals | 16 | 0.236 | ||||
| J | Substrate | 1 | 0.07071 | 6.512 | * | C > I |
| Immersed time | 3 | 0.02659 | 2.449 | 0.1012 | ||
| S × T | 3 | 0.01403 | 1.292 | 0.3113 | ||
| Residuals | 16 | 0.01086 | ||||
| N | Substrate | 1 | 78.94 | 36.596 | *** | C > I |
| Immersed time | 3 | 15.78 | 7.314 | ** | 12 < 3 = 6 = 9 | |
| S × T | 3 | 0.7 | 0.326 | 0.80647 | ||
| Residuals | 16 | 2.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carmona-Rodríguez, A.; Antón, C.; Climent, M.-Á.; Garcés, P.; Montiel, V.; Ramos-Esplá, A.A. Sessile Biofouling on Electrolytic Carbonated Structures: Stages of Colonization and Succession. J. Mar. Sci. Eng. 2024, 12, 443. https://doi.org/10.3390/jmse12030443
Carmona-Rodríguez A, Antón C, Climent M-Á, Garcés P, Montiel V, Ramos-Esplá AA. Sessile Biofouling on Electrolytic Carbonated Structures: Stages of Colonization and Succession. Journal of Marine Science and Engineering. 2024; 12(3):443. https://doi.org/10.3390/jmse12030443
Chicago/Turabian StyleCarmona-Rodríguez, Alejandro, Carlos Antón, Miguel-Ángel Climent, Pedro Garcés, Vicente Montiel, and Alfonso A. Ramos-Esplá. 2024. "Sessile Biofouling on Electrolytic Carbonated Structures: Stages of Colonization and Succession" Journal of Marine Science and Engineering 12, no. 3: 443. https://doi.org/10.3390/jmse12030443
APA StyleCarmona-Rodríguez, A., Antón, C., Climent, M.-Á., Garcés, P., Montiel, V., & Ramos-Esplá, A. A. (2024). Sessile Biofouling on Electrolytic Carbonated Structures: Stages of Colonization and Succession. Journal of Marine Science and Engineering, 12(3), 443. https://doi.org/10.3390/jmse12030443