
Numerical Study on the Hydrodynamics of Manta Rays Exiting Water

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Physical Model and Kinematics Model
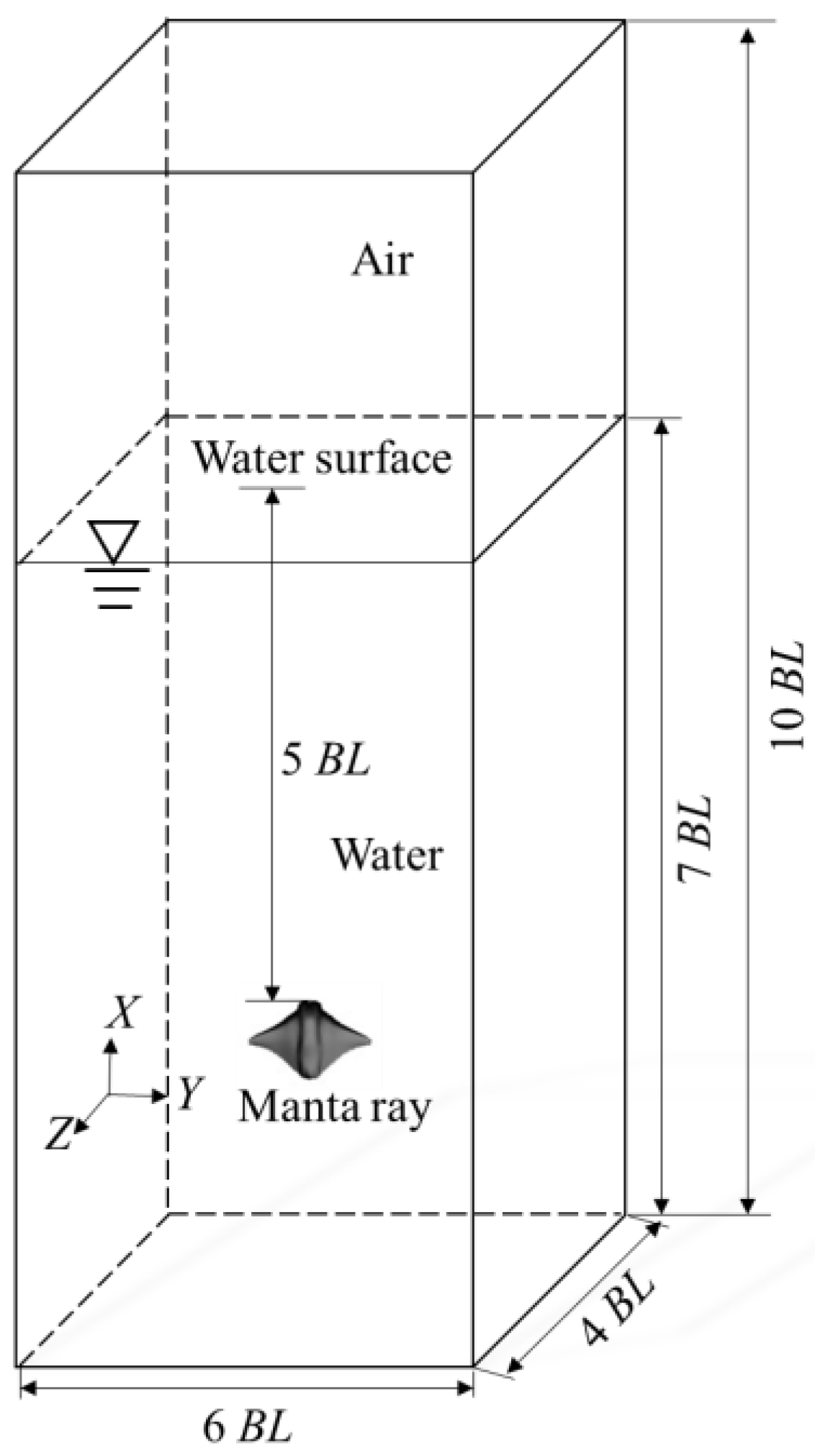
2.1. Physical Model
2.2. Kinematics Model
3. Numerical Method
3.1. Governing Equations of Fluids
3.2. Dynamic Equations of the Manta Ray
3.3. Numerical Method and Computational Grid
3.4. Sensitivity Study and Validation Test
4. Results and Discussion
4.1. Time History of Displacement, Velocity and Force of the Manta Ray
4.2. Transient Variation in Flow Field
4.3. Effect of Kinematic Parameters on Porpoising Performance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosenberger, L.J. Pectoral fin locomotion in batoid fishes: Undulation versus oscillation. J. Exp. Biol. 2001, 204, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.D.; Compagno, L.J.; Bennett, M.B. Redescription of the genus manta with resurrection of manta alfredi (Krefft, 1868) (Chondrichthyes; Myliobatoidei; Mobulidae). Zootaxa 2009, 2301, 1–28. [Google Scholar] [CrossRef]
- Fontanella, J.E.; Fish, F.E.; Barchi, E.I.; Campbell-Malone, R.; Nichols, R.H.; DiNenno, N.K.; Beneski, J.T. Two-and three-dimensional geometries of batoids in relation to locomotor mode. J. Exp. Mar. Biol. Ecol. 2013, 446, 273–281. [Google Scholar] [CrossRef]
- Heine, C.E. Mechanics of Flapping Fin Locomotion in the Cownose Ray, Rhinoptera Bonasus (Elasmobranchii: Myliobatidae). Ph.D. Thesis, Duke University, Durham, NC, USA, 1992. [Google Scholar]
- Blevins, E.L.; Lauder, G.V. Rajiform locomotion: Three-dimensional kinematics of the pectoral fin surface during swimming in the freshwater stingray Potamotrygon orbignyi. J. Exp. Biol. 2012, 215, 3231–3241. [Google Scholar] [CrossRef]
- Russo, R.S.; Blemker, S.S.; Fish, F.E.; Bart-Smith, H. Biomechanical model of batoid (skates and rays) pectoral fins predicts the influence of skeletal structure on fin kinematics: Implications for bio-inspired design. Bioinspir. Biomim. 2015, 10, 046002. [Google Scholar] [CrossRef]
- Fish, F.E.; Schreiber, C.M.; Moored, K.W.; Liu, G.; Dong, H.; Bart-Smith, H. Hydrodynamic performance of aquatic flapping: Efficiency of underwater flight in the manta. Aerospace 2016, 3, 20. [Google Scholar] [CrossRef]
- Fish, F.E.; Dong, H.; Zhu, J.J.; Bart-Smith, H. Kinematics and hydrodynamics of mobuliform swimming: Oscillatory winged propulsion by large pelagic batoids. Mar. Technol. Soc. J. 2017, 51, 35–47. [Google Scholar] [CrossRef]
- Zhang, D.; Huang, Q.G.; Pan, G.; Yang, L.M.; Huang, W.X. Vortex dynamics and hydrodynamic performance enhancement mechanism in batoid fish oscillatory swimming. J. Fluid Mech. 2021, 930, A28. [Google Scholar] [CrossRef]
- Liu, G.; Ren, Y.; Zhu, J.; Bart-Smith, H.; Dong, H. Thrust producing mechanisms in ray-inspired underwater vehicle propulsion. Theor. Appl. Mech. Lett. 2015, 5, 54–57. [Google Scholar] [CrossRef]
- Thekkethil, N.; Sharma, A.; Agrawal, A. Three-dimensional biological hydrodynamics study on various types of batoid fishlike locomotion. Phys. Rev. Fluids 2020, 5, 023101. [Google Scholar] [CrossRef]
- Huang, Q.G.; Zhang, D.; Pan, G. Computational model construction and analysis of the hydrodynamics of a rhinoptera javanica. IEEE Access 2020, 8, 30410–30420. [Google Scholar] [CrossRef]
- Menzer; Gong, Y.; Fish, F.E.; Dong, H. Bio-inspired propulsion: Towards understanding the role of pectoral fin kinematics in manta-like swimming. Biomimetics 2022, 7, 45. [Google Scholar] [CrossRef]
- Bianchi, G.; Cinquemani, S.; Schito, P.; Resta, F. A numerical model for the analysis of the locomotion of a cownose ray. J. Fluids Eng. 2022, 144, 031203. [Google Scholar] [CrossRef]
- Xing, C.; Cao, Y.H.; Cao, Y.; Pan, G.; Huang, Q.G. Asymmetrical oscillating morphology hydrodynamic performance of a novel bionic pectoral fin. J. Mar. Sci. Eng. 2022, 10, 289. [Google Scholar] [CrossRef]
- Manduca, G.; Santaera, G.; Miraglia, M.; Van Vuuren, G.J.; Dario, P.; Stefanini, C.; Romano, D. A bioinspired control strategy ensures maneuverability and adaptability for dynamic environments in an underactuated robotic fish. J. Intell. Robot. Syst. 2024, 110, 69. [Google Scholar] [CrossRef]
- Mo, Y.; Su, W.; Hong, Z.; Li, Y.; Zhong, Y. Finite-time line-of-sight guidance-based path-following control for a wire-driven robot fish. Biomimetics 2024, 9, 556. [Google Scholar] [CrossRef]
- Miao, G. Hydrodynamic forces and dynamic responses of circular cylinders in wave zones. In Marine Hydrodynamics; NTH: Trondheim, Norway, 1989. [Google Scholar]
- Liju, P.Y.; Machane, R.; Cartellier, A. Surge effect during the water exit of an axis-symmetric body traveling normal to a plane interface: Experiments and BEM simulation. Exp. Fluids 2001, 31, 241–248. [Google Scholar] [CrossRef]
- Colicchio, G.; Greco, M.; Miozzi, M.; Lugni, C. Experimental and numerical investigation of the water-entry and water-exit of a circular cylinder. In Proceedings of the 24th International Workshop on Water Waves and Floating Bodies, Zelenogorsk, Russia, 19–22 April 2009. [Google Scholar]
- Moshari, S.; Nikseresht, A.H. Numerical analysis of two and three dimensional buoyancy driven water-exit of a circular cylinder. Int. J. Nav. Arch. Ocean Eng. 2014, 6, 219–235. [Google Scholar] [CrossRef]
- Ye, Q.; He, Y. Perturbation solution to the nonlinear problem of oblique water exit of an axisymmetric body with a large exit-angle. Appl. Math. Mech. 1991, 12, 327–338. [Google Scholar] [CrossRef]
- Zhang, J.; Hong, F.W.; Xu, F.; Wang, L.P.; Zhao, F. Experimental research of transient flow field near free surface due to body exiting from water. J. Ship Mech. 2002, 6, 45–50. [Google Scholar]
- Xia, D.; Yin, Q.; Li, Z.; Chen, W.; Shi, Y.; Dou, J. Numerical study on the hydrodynamics of porpoising behavior in dolphins. Ocean Eng. 2021, 229, 108985. [Google Scholar] [CrossRef]
- Hou, T.G.; Yang, X.B.; Wang, T.M.; Liang, J.H.; Li, S.W.; Fan, Y.B. Locomotor transition: How squid jet from water to air. Bioinspir. Biomim. 2020, 15, 036014. [Google Scholar] [CrossRef] [PubMed]
- Menter, F.R. Two-equation eddy-viscosity transport turbulence model for engineering applications. AIAA J. 1994, 32, 1598–1605. [Google Scholar] [CrossRef]
- Wang, Z.J. A conservative overlapped (chimera) grid algorithm for multiple moving body flows. In Proceedings of the 34th Aerospace Sciences Meeting and Exhibit, Reno, NV, USA, 15–18 January 1996; p. 823. [Google Scholar]
- Wei, C.; Hu, Q.; Shi, X.; Zeng, Y. A comparison for hydrodynamic performance of undulating fin propulsion on numerical self-propulsion and tethered models. Ocean Eng. 2022, 265, 112471. [Google Scholar] [CrossRef]















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, D.-H.; Zhang, M.-H.; Wu, X.-Y.; Pei, Y.; Liu, X.-J.; Xing, C.; Cao, Y.; Cao, Y.-H.; Pan, G. Numerical Study on the Hydrodynamics of Manta Rays Exiting Water. J. Mar. Sci. Eng. 2024, 12, 2125. https://doi.org/10.3390/jmse12122125
Zhou D-H, Zhang M-H, Wu X-Y, Pei Y, Liu X-J, Xing C, Cao Y, Cao Y-H, Pan G. Numerical Study on the Hydrodynamics of Manta Rays Exiting Water. Journal of Marine Science and Engineering. 2024; 12(12):2125. https://doi.org/10.3390/jmse12122125
Chicago/Turabian StyleZhou, Dong-Hui, Min-Hui Zhang, Xiao-Yang Wu, Yu Pei, Xue-Jing Liu, Cheng Xing, Yong Cao, Yong-Hui Cao, and Guang Pan. 2024. "Numerical Study on the Hydrodynamics of Manta Rays Exiting Water" Journal of Marine Science and Engineering 12, no. 12: 2125. https://doi.org/10.3390/jmse12122125
APA StyleZhou, D.-H., Zhang, M.-H., Wu, X.-Y., Pei, Y., Liu, X.-J., Xing, C., Cao, Y., Cao, Y.-H., & Pan, G. (2024). Numerical Study on the Hydrodynamics of Manta Rays Exiting Water. Journal of Marine Science and Engineering, 12(12), 2125. https://doi.org/10.3390/jmse12122125