

A Hybrid Design for the Functional Assay of LvLRRm (Protein Containing LRR Domain) of the White Leg Shrimp, Litopenaeus vannamei



Abstract
:1. Introduction
2. Materials and Methods
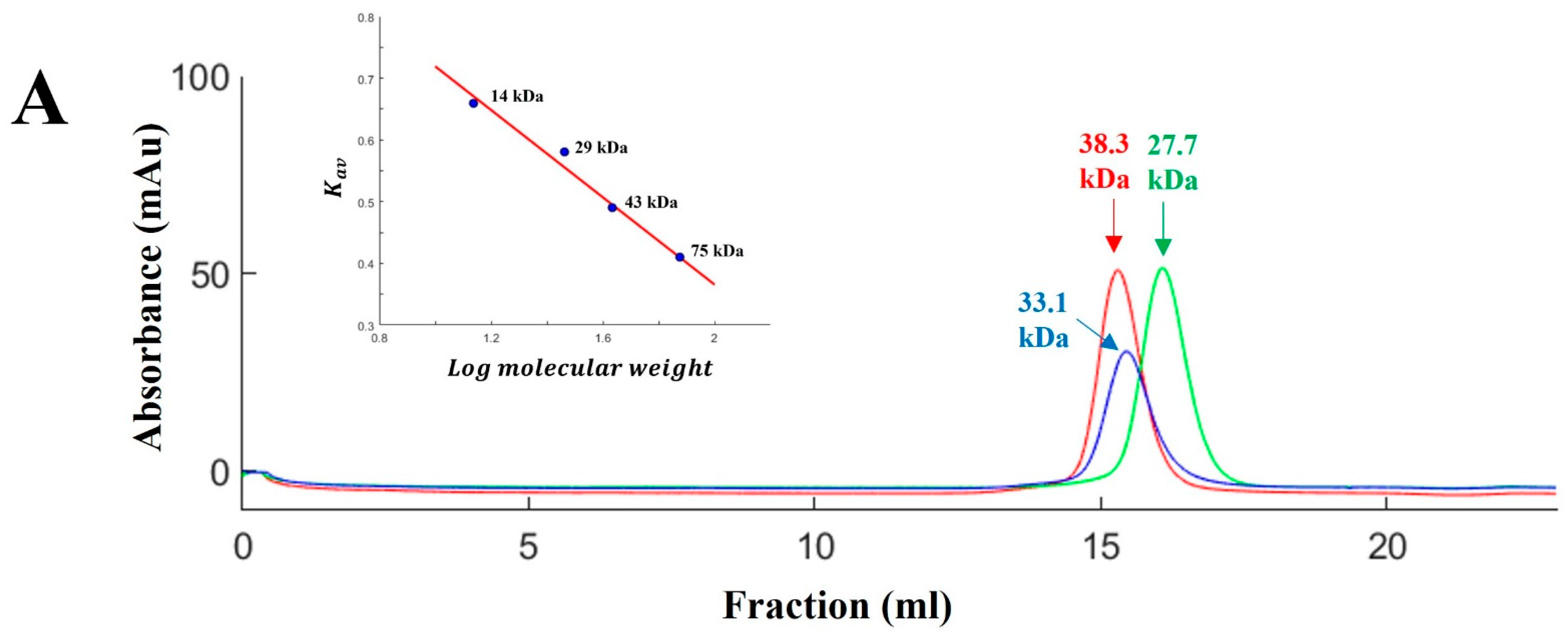
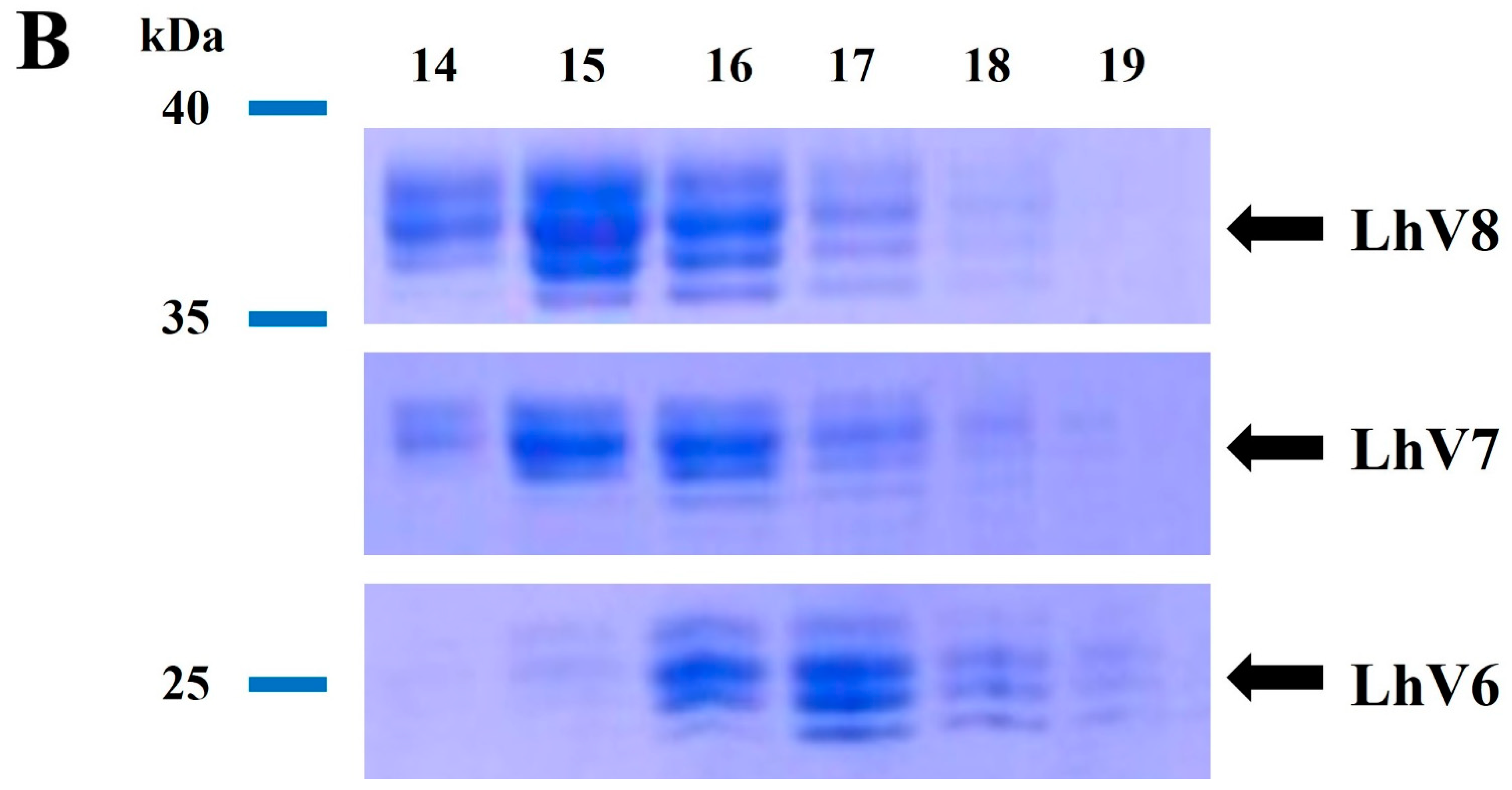
2.1. Cell Culture, Recombinant Protein Expression, and Purification
2.2. SPR (Surface Plasmon Resonance) Experiments by Single-Cycle Kinetics
3. Results
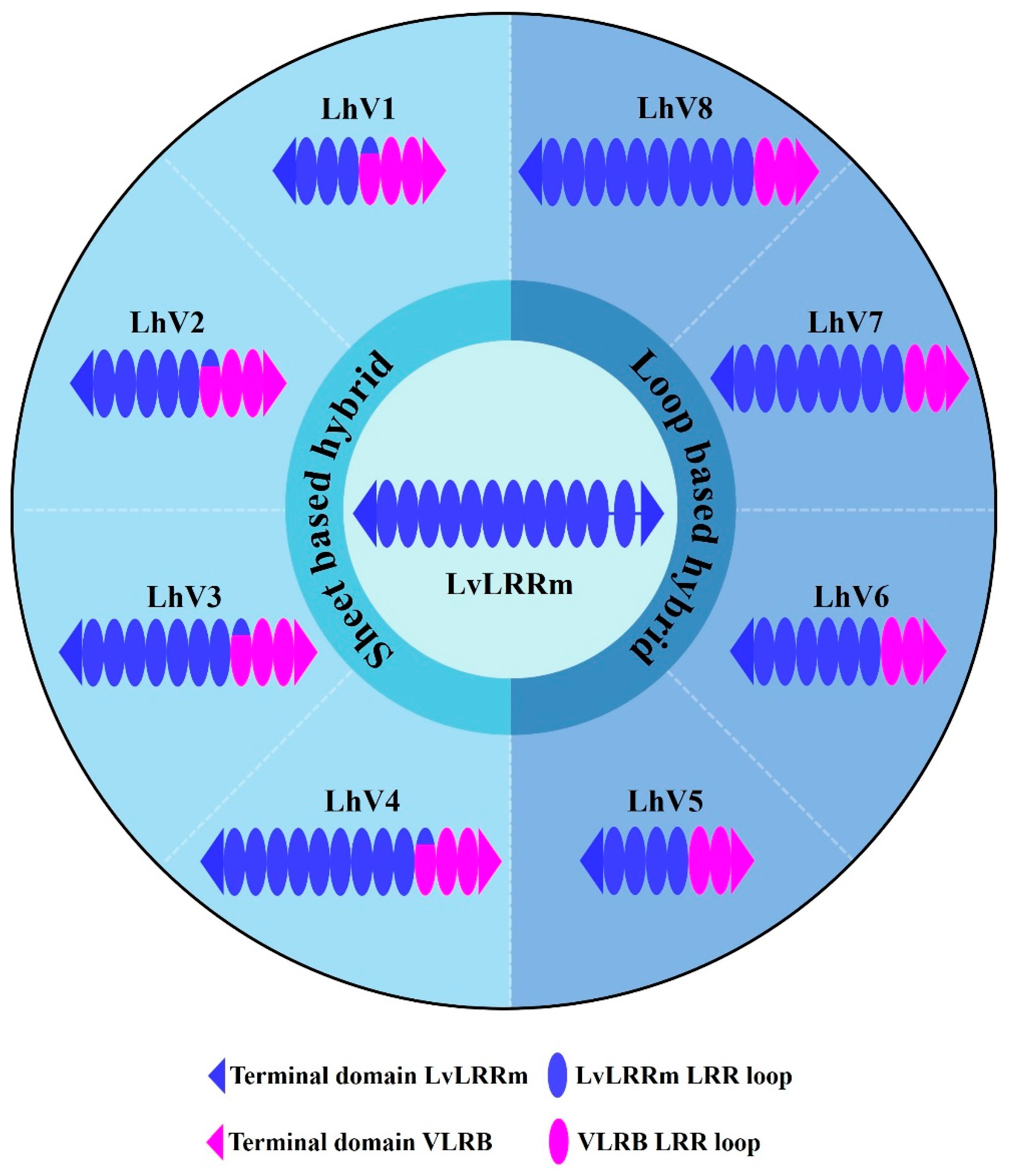
3.1. Design of LvLRRm Hybrid Proteins, LhV Proteins
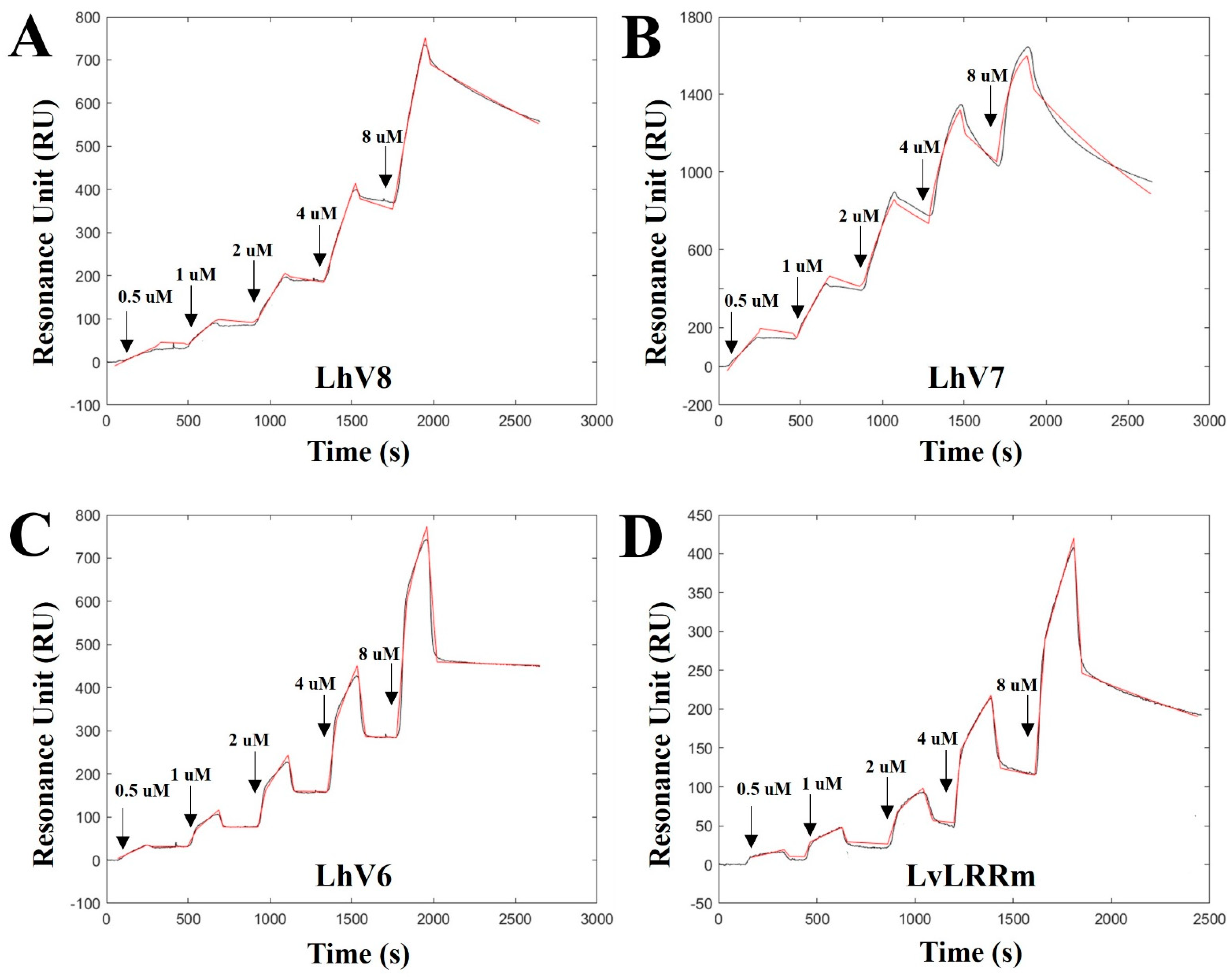
3.2. Binding Kinetics Validation of LvLRRm and LhV6–8 to Stflagellin
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-León, P.; Luna-González, A.; Escamilla-Montes, R.; del Carmen Flores-Miranda, M.; Fierro-Coronado, J.A.; Álvarez-Ruiz, P.; Diarte-Plata, G. Isolation and characterization of infectious Vibrio parahaemolyticus, the causative agent of AHPND, from the whiteleg shrimp (Litopenaeus vannamei). Lat. Am. J. Aquat. Res. 2016, 44, 470–479. [Google Scholar] [CrossRef]
- Lomelí-Ortega, C.O.; Martínez-Díaz, S.F. Phage therapy against Vibrio parahaemolyticus infection in the whiteleg shrimp (Litopenaeus vannamei) larvae. Aquaculture 2014, 434, 208–211. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Xu, K.; Zhang, X.; Sun, H.; Fan, L.; Yan, M. White spot syndrome virus (WSSV) infection impacts intestinal microbiota composition and function in Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 84, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, L.; Alpuche, J.; Maldonado, G.; Agundis, C.; Pereyra-Morales, A.; Zenteno, E. Review: Immunity mechanisms in crustaceans. Innate Immun. 2009, 15, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Soderhall, I. Crustacean hematopoiesis and the astakine cytokines. Blood 2011, 117, 6417–6424. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.T.; Kong, T.; Zhang, M.; Li, S. Pattern recognition receptors and their roles on the innate immune system of mud crab (Scylla paramamosain). Dev. Comp. Immunol. 2020, 102, 103469. [Google Scholar] [CrossRef]
- Zhang, H.; Li, S.; Wang, F.; Xiang, J.; Li, F. Identification and functional study of an LRR domain containing membrane protein in Litopenaeus vannamei. Dev. Comp. Immunol. 2020, 109, 103713. [Google Scholar] [CrossRef] [PubMed]
- Habib, Y.J.; Zhang, Z. The involvement of crustaceans toll-like receptors in pathogen recognition. Fish Shellfish Immunol. 2020, 102, 169–176. [Google Scholar] [CrossRef]
- Park, H.; Huxley-Jones, J.; Boot-Handford, R.P.; Bishop, P.N.; Attwood, T.K.; Bella, J. LRRCE: A leucine-rich repeat cysteine capping motif unique to the chordate lineage. BMC Genom. 2008, 9, 599. [Google Scholar] [CrossRef]
- Wen, D.; Wildes, C.P.; Silvian, L.; Walus, L.; Mi, S.; Lee, D.H.; Meier, W.; Pepinsky, R.B. Disulfide structure of the leucine-rich repeat C-terminal cap and C-terminal stalk region of Nogo-66 receptor. Biochemistry 2005, 44, 16491–16501. [Google Scholar] [CrossRef]
- Kajava, A.V.; Kobe, B. Assessment of the ability to model proteins with leucine-rich repeats in light of the latest structural information. Protein Sci. 2002, 11, 1082–1090. [Google Scholar] [CrossRef]
- Wang, X.W.; Gao, J.; Xu, Y.H.; Xu, J.D.; Fan, Z.X.; Zhao, X.F.; Wang, J.X. Novel Pattern Recognition Receptor Protects Shrimp by Preventing Bacterial Colonization and Promoting Phagocytosis. J. Immunol. 2017, 198, 3045–3057. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.C.; Xu, J.Y.; Liu, H.P. Cellular entry of white spot syndrome virus and antiviral immunity mediated by cellular receptors in crustaceans. Fish Shellfish Immunol. 2019, 93, 580–588. [Google Scholar] [CrossRef]
- Jin, M.S.; Lee, J.O. Application of hybrid LRR technique to protein crystallization. BMB Rep. 2008, 41, 353–357. [Google Scholar] [CrossRef]
- Park, B.S.; Song, D.H.; Kim, H.M.; Choi, B.S.; Lee, H.; Lee, J.O. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar] [CrossRef]
- Jin, M.S.; Kim, S.E.; Heo, J.Y.; Lee, M.E.; Kim, H.M.; Paik, S.G.; Lee, H.; Lee, J.O. Crystal structure of the TLR1-TLR2 heterodimer induced by binding of a tri-acylated lipopeptide. Cell 2007, 130, 1071–1082. [Google Scholar] [CrossRef]
- Speltz, E.B.; Brown, R.S.H.; Hajare, H.S.; Schlieker, C.; Regan, L. A designed repeat protein as an affinity capture reagent. Biochem. Soc. Trans. 2015, 43, 874–880. [Google Scholar] [CrossRef]
- Kim, H.M.; Park, B.S.; Kim, J.I.; Kim, S.E.; Lee, J.; Oh, S.C.; Enkhbayar, P.; Matsushima, N.; Lee, H.; Yoo, O.J.; et al. Crystal structure of the TLR4-MD-2 complex with bound endotoxin antagonist Eritoran. Cell 2007, 130, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Oh, S.C.; Lim, K.J.; Kasamatsu, J.; Heo, J.Y.; Park, B.S.; Lee, H.; Yoo, O.J.; Kasahara, M.; Lee, J.O. Structural diversity of the hagfish variable lymphocyte receptors. J. Biol. Chem. 2007, 282, 6726–6732. [Google Scholar] [CrossRef]
- Pancer, Z.; Saha, N.R.; Kasamatsu, J.; Suzuki, T.; Amemiya, C.T.; Kasahara, M.; Cooper, M.D. Variable lymphocyte receptors in hagfish. Proc. Natl. Acad. Sci. USA 2005, 102, 9224–9229. [Google Scholar] [CrossRef]
- Guo, P.; Gartland, L.; Li, J.X.; Hirano, M.; Alder, M.; Herrin, B.; Sides, J.; Kasahara, M.; Cooper, M. Characterization of adaptive immune receptors in hagfish. J. Immunol. 2011, 186, 170.13. [Google Scholar] [CrossRef]
- Boehm, T.; Hirano, M.; Holland, S.J.; Das, S.; Schorpp, M.; Cooper, M.D. Evolution of Alternative Adaptive Immune Systems in Vertebrates. Annu. Rev. Immunol. 2018, 36, 19–42. [Google Scholar] [CrossRef]
- Usharauli, D. Chronic infection and the origin of adaptive immune system. Med. Hypotheses 2010, 75, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Hauton, C. The scope of the crustacean immune system for disease control. J. Invertebr. Pathol. 2012, 110, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, E.P.; Beis, K.; Cameron, A.D.; Iwata, S. Overcoming the challenges of membrane protein crystallography. Curr. Opin. Struct. Biol. 2008, 18, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Errasti-Murugarren, E.; Bartoccioni, P.; Palacin, M. Membrane Protein Stabilization Strategies for Structural and Functional Studies. Membranes 2021, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Nurnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate immunity in plants and animals: Striking similarities and obvious differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef]
- Coscia, M.R.; Giacomelli, S.; Oreste, U. Toll-like receptors: An overview from invertebrates to vertebrates. ISJ Invertebr. Surviv. J. 2011, 8, 210–226. [Google Scholar]
- Bella, J.; Hindle, K.L.; McEwan, P.A.; Lovell, S.C. The leucine-rich repeat structure. Cell. Mol. Life Sci. 2008, 65, 2307–2333. [Google Scholar] [CrossRef]
- Shen, X.; Wang, Y.; Hu, J.; Bao, Z.; Wang, M. Molecular characterization of an LRR-only protein gene in Pacific white shrimp Litopenaeus vannamei: Sequence feature, expression pattern, and protein activity. Fish Shellfish Immunol. 2022, 129, 199–206. [Google Scholar] [CrossRef]
- Yoon, S.I.; Kurnasov, O.; Natarajan, V.; Hong, M.; Gudkov, A.V.; Osterman, A.L.; Wilson, I.A. Structural basis of TLR5-flagellin recognition and signaling. Science 2012, 335, 859–864. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Jacchieri, S.G.; Torquato, R.; Brentani, R.R. Structural study of binding of flagellin by Toll-like receptor 5. J. Bacteriol. 2003, 185, 4243–4247. [Google Scholar] [CrossRef]
- Buchanan, S.G.; Gay, N.J. Structural and functional diversity in the leucine-rich repeat family of proteins. Prog. Biophys. Mol. Biol. 1996, 65, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Mizel, S.B.; West, A.P.; Hantgan, R.R. Identification of a sequence in human toll-like receptor 5 required for the binding of Gram-negative flagellin. J. Biol. Chem. 2003, 278, 23624–23629. [Google Scholar] [CrossRef] [PubMed]
- Song, W.S.; Jeon, Y.J.; Namgung, B.; Hong, M.; Yoon, S.I. A conserved TLR5 binding and activation hot spot on flagellin. Sci. Rep. 2017, 7, 40878. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Shen, B.W.; Parmeggiani, F.; Huang, P.S.; Stoddard, B.L.; Baker, D. Control of repeat-protein curvature by computational protein design. Nat. Struct. Mol. Biol. 2015, 22, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Bi, C.; Li, Z.; Podariu, M.; Hage, D.S. Analytical methods for kinetic studies of biological interactions: A review. J. Pharm. Biomed. Anal. 2015, 113, 163–180. [Google Scholar] [CrossRef]
- Karlsson, R.; Katsamba, P.S.; Nordin, H.; Pol, E.; Myszka, D.G. Analyzing a kinetic titration series using affinity biosensors. Anal. Biochem. 2006, 349, 136–147. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct | Primer (5′ → 3′) | Gene | |
|---|---|---|---|
| LhV1 | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | Polymerized by Hybrid LRR technique |
| Hybrid1_F | GTACTCTCACTCAACCAGCTGAAG | ||
| Hybrid1_R | CTTCAGCTGGTTGAGTGAGAGTAC | ||
| VLRB.61_R | ACTACGCGGCCGCTAGTAGGGCAGATGATACTTCGGACGGG | ||
| LhV2 | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | Polymerized by Hybrid LRR technique |
| Hybrid2_F | GTGCTTCATTTGAACCAGCTGAAG | ||
| Hybrid2_R | CTTCAGCTGGTTCAAATGAAGCAC | ||
| VLRB.61_R | ACTACGCGGCCGCTAGTAGGGCAGATGATACTTCGGACGGG | ||
| LhV3 | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | Polymerized by Hybrid LRR technique |
| Hybrid3_F | CTCGACCTTTCGAACCAGCTGAAG | ||
| Hybrid3_R | CTTCAGCTGGTTCGAAAGGTCGAG | ||
| VLRB.61_R | ACTACGCGGCCGCTAGTAGGGCAGATGATACTTCGGACGGG | ||
| LhV4 | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | Polymerized by Hybrid LRR technique |
| Hybrid4_F | GTAAGTGGCTGCAACCAGCTGAAG | ||
| Hybrid4_R | CTTCAGCTGGTTGCAGCCACTTAC | ||
| VLRB.61_R | ACTACGCGGCCGCTAGTAGGGCAGATGATACTTCGGACGGG | ||
| LhV5 | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | Polymerized by Hybrid LRR technique |
| Hybrid5_F | CAGCTGAAACAGCTCAAAGAGCTCGCCCTGGACACC | ||
| Hybrid5_R | GAGCTCTTTGAGCTGTTTCAGCTGATAAAGACACC | ||
| VLRB.61_R | ACTACGCGGCCGCTAGTAGGGCAGATGATACTTCGGACGGG | ||
| LhV6 | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | Polymerized by Hybrid LRR technique |
| Hybrid6_F | CTGGTACCCGATCTCAAAGAGCTCGCCCTGGACACC | ||
| Hybrid6_R | GAGCTCTTTGAGATCGGGTACCAGTGCAAGATTCTG | ||
| VLRB.61_R | ACTACGCGGCCGCTAGTAGGGCAGATGATACTTCGGACGGG | ||
| LhV7 | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | Polymerized by Hybrid LRR technique |
| Hybrid7_F | GGACTGTCTCTTCTCAAAGAGCTCGCCCTGGACACC | ||
| Hybrid7_R | GAGCTCTTTGAGAAGAGACAGTCCTTGGAGACTGTT | ||
| VLRB.61_R | ACTACGCGGCCGCTAGTAGGGCAGATGATACTTCGGACGGG | ||
| LhV8 | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | Polymerized by Hybrid LRR technique |
| Hybrid8_F | GAGAATCGTAGACTCAAAGAGCTCGCCCTGGACACC | ||
| Hybrid8_R | GAGCTCTTTGAGTCTACGATTCTCTTTGAAGGCATC | ||
| VLRB_R | ACTACGCGGCCGCTAGTAGGGCAGATGATACTTCGGACGGG | ||
| LvLRRm | LvLRRm_F | ATCGAGGATCCTCTTTGTCCCAACAATTGT | MT180094 |
| LvLRRm_R | ATTCGGCGGCCGCTACCGCAAGCGAGATCCGACAG | ||
| EbVLRB.61 | VLRB.61_F | ACTACGGATCCATGATGGGTCCGGTCTTGGCTGCAT | 2Z62_A |
| VLRB.61_R | ACTACGCGGCCGCTATTTGGAGAGACGCAATCTGAGGCCGC |
| Construct Number | Amino Acid Sequences | Soluble Expression | |
|---|---|---|---|
| LvLRRm | VLRB.61 | ||
| LhV1 | 51 205 | 236 303 | |
| VSL•••KLIVLSL | NQLKSVP•••CPT | No | |
| LxxLxLxxNx | LxxLxLxxNxLx | ||
| LhV2 | 51 253 | 236 303 | |
| VSL•••NLRVLHL | NQLKSVP•••CPT | No | |
| LxxLxLxxNx | LxxLxLxxNxLx | ||
| LhV3 | 51 302 | 236 303 | |
| VSL•••LKDLDLS | NQLKSVP•••CPT | No | |
| LxxLxLxxNx | LxxLxLxxNxLx | ||
| LhV4 | 51 377 | 236 303 | |
| VSL•••SLFVSGC | NQLKSVP•••CPT | No | |
| LxxLxLxxNx | LxxLxLxxNxLx | ||
| LhV5 | 96 224 | 229 303 | |
| LCP•••FIQLKQL | KELALVT•••CPT | No | |
| LxxLxLxxNx | LxxLxLxxNxLx | ||
| LhV6 | 96 272 | 229 303 | |
| LCP•••LALVPDL | KELALVT•••CPT | Yes | |
| LxxLxLxxNx | LxxLxLxxNxLx | ||
| LhV7 | 96 321 | 229 303 | |
| LCP•••LQGLSLL | KELALVT•••CPT | Yes | |
| LxxLxLxxNx | LxxLxLxxNxLx | ||
| LhV8 | 96 369 | 229 303 | |
| LCP•••FKENRRL | KELALVT•••CPT | Yes | |
| LxxLxLxxNx | LxxLxLxxNxLx | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, J.; Lee, W.; Kim, T.; Park, B. A Hybrid Design for the Functional Assay of LvLRRm (Protein Containing LRR Domain) of the White Leg Shrimp, Litopenaeus vannamei. J. Mar. Sci. Eng. 2023, 11, 2257. https://doi.org/10.3390/jmse11122257
Cho J, Lee W, Kim T, Park B. A Hybrid Design for the Functional Assay of LvLRRm (Protein Containing LRR Domain) of the White Leg Shrimp, Litopenaeus vannamei. Journal of Marine Science and Engineering. 2023; 11(12):2257. https://doi.org/10.3390/jmse11122257
Chicago/Turabian StyleCho, Jaemin, Wooyong Lee, Taewon Kim, and Beomseok Park. 2023. "A Hybrid Design for the Functional Assay of LvLRRm (Protein Containing LRR Domain) of the White Leg Shrimp, Litopenaeus vannamei" Journal of Marine Science and Engineering 11, no. 12: 2257. https://doi.org/10.3390/jmse11122257
APA StyleCho, J., Lee, W., Kim, T., & Park, B. (2023). A Hybrid Design for the Functional Assay of LvLRRm (Protein Containing LRR Domain) of the White Leg Shrimp, Litopenaeus vannamei. Journal of Marine Science and Engineering, 11(12), 2257. https://doi.org/10.3390/jmse11122257