
Coastal Protection by Planted Mangrove Forest during Typhoon Mangkhut


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and the Planted Mangrove Forest
2.2. Aerial Survey
2.3. Hydrodynamic Survey
3. Results and Discussions
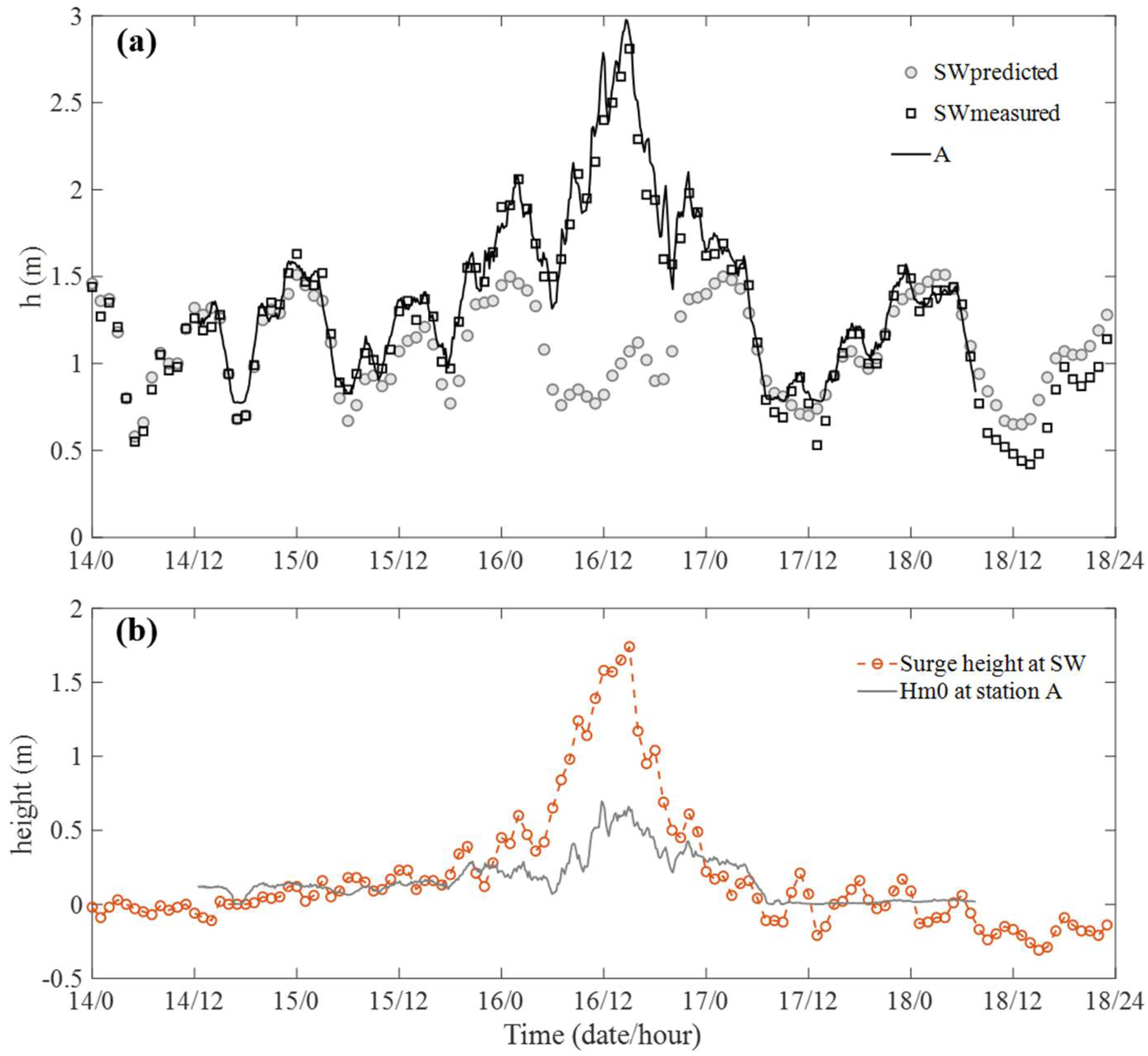
3.1. Tide Condition and Typhoon Influence
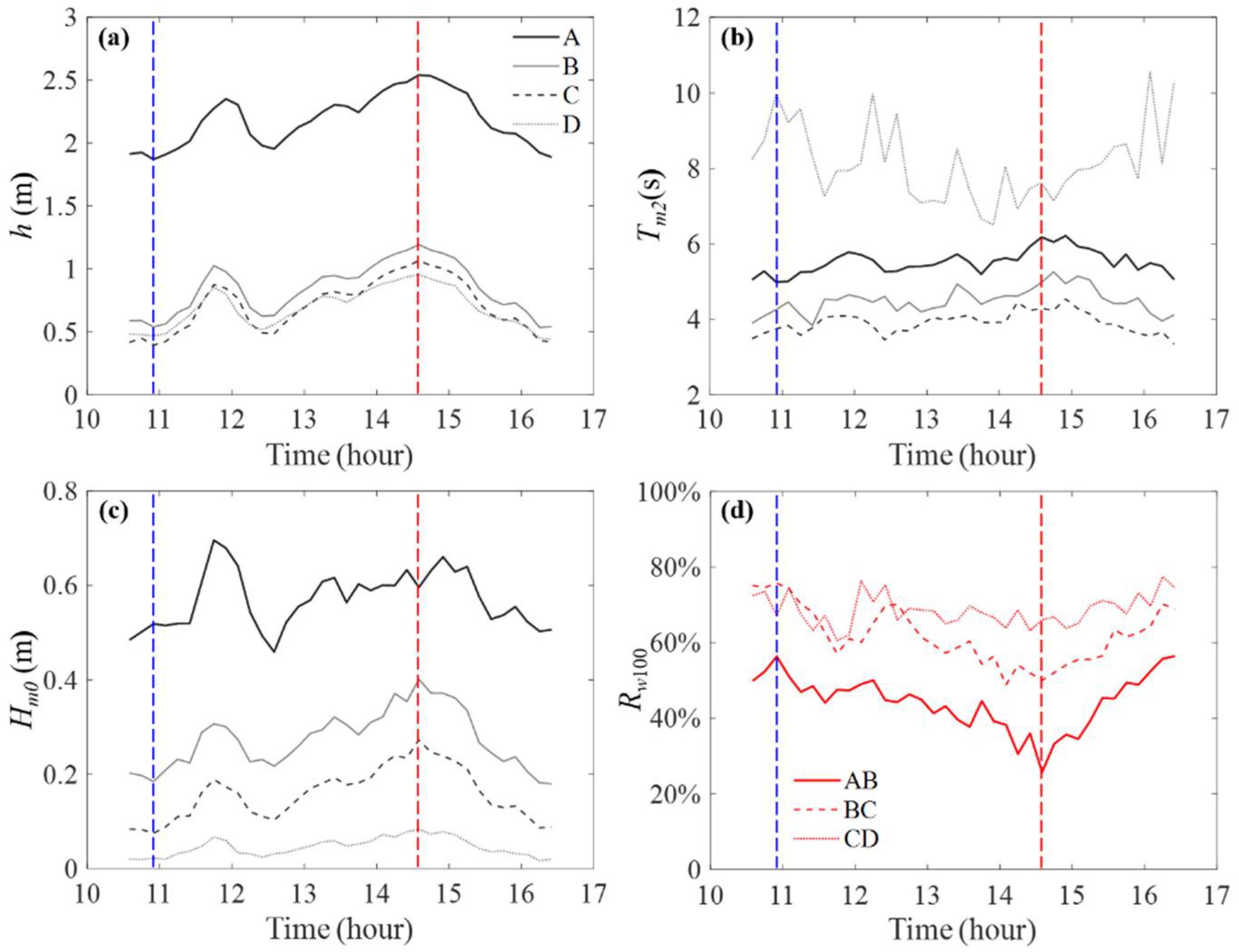
3.2. Wave Condition and Wave Attenuation
3.3. Resilience of Planted Mangrove to Intense Storm
3.4. Protection of Fish Ponds and Embankments by Mangroves during Storms
3.5. Coastal Sedimentation and Erosion Prevention within Mangroves
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitra, A. Mangrove Forests in India: Exploring Ecosystem Services; Springer International Publishing: Cham, Switzerland, 2020; ISBN 978-3-030-20594-2. [Google Scholar]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the Most Carbon-Rich Forests in the Tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Pendleton, L.; Donato, D.C.; Murray, B.C.; Crooks, S.; Jenkins, W.A.; Sifleet, S.; Craft, C.; Fourqurean, J.W.; Kauffman, J.B.; Marbà, N.; et al. Estimating Global “Blue Carbon” Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems. PLoS ONE 2012, 7, e43542. [Google Scholar] [CrossRef] [PubMed]
- Akber, M.A.; Patwary, M.M.; Islam, M.A.; Rahman, M.R. Storm Protection Service of the Sundarbans Mangrove Forest, Bangladesh. Nat. Hazards 2018, 94, 405–418. [Google Scholar] [CrossRef]
- Koh, H.; Teh, S.; Kh’ng, X.; Raja Barizan, R. Mangrove Forests: Protection against and Resilience to Coastal Disturbances. JTFS 2018, 30, 446–460. [Google Scholar] [CrossRef]
- Lee, W.K.; Tay, S.H.X.; Ooi, S.K.; Friess, D.A. Potential Short Wave Attenuation Function of Disturbed Mangroves. Estuar. Coast. Shelf Sci. 2021, 248, 106747. [Google Scholar] [CrossRef]
- Mazda, Y.; Magi, M.; Ikeda, Y.; Kurokawa, T.; Asano, T. Wave Reduction in a Mangrove Forest Dominated by Sonneratia sp. Wetl. Ecol. Manag. 2006, 14, 365–378. [Google Scholar] [CrossRef]
- Quartel, S.; Kroon, A.; Augustinus, P.G.E.F.; Van Santen, P.; Tri, N.H. Wave Attenuation in Coastal Mangroves in the Red River Delta, Vietnam. J. Asian Earth Sci. 2007, 29, 576–584. [Google Scholar] [CrossRef]
- Dasgupta, S.; Islam, M.S.; Huq, M.; Huque Khan, Z.; Hasib, M.R. Quantifying the Protective Capacity of Mangroves from Storm Surges in Coastal Bangladesh. PLoS ONE 2019, 14, e0214079. [Google Scholar] [CrossRef]
- Mullarney, J.C.; Henderson, S.M.; Reyns, J.A.H.; Norris, B.K.; Bryan, K.R. Spatially Varying Drag within a Wave-Exposed Mangrove Forest and on the Adjacent Tidal Flat. Cont. Shelf Res. 2017, 147, 102–113. [Google Scholar] [CrossRef]
- McIvor, A.; Spencer, T.; Möller, I.; Spalding, M. Storm Surge Reduction by Mangroves; Natural Coastal Protection Series: Report 2; Cambridge Coastal Research Unit Working Paper 41; University of Cambridge: Cambridge, UK, 2012. [Google Scholar]
- Badola, R.; Hussain, S.A. Valuing Ecosystem Functions: An Empirical Study on the Storm Protection Function of Bhitarkanika Mangrove Ecosystem, India. Environ. Conserv. 2005, 32, 85–92. [Google Scholar] [CrossRef]
- Mazda, Y.; Magi, M.; Kogo, M.; Hong, P.N. Mangroves as a Coastal Protection from Waves in the Tong King Delta, Vietnam. Mangroves Salt Marshes 1997, 1, 127–135. [Google Scholar] [CrossRef]
- Koh, H.L.; Teh, S.Y.; Liu, P.L.; Ismail, A.I.M.; Lee, H.L. Simulation of Andaman 2004 Tsunami for Assessing Impact on Malaysia. J. Asian Earth Sci. 2009, 36, 74–83. [Google Scholar] [CrossRef]
- Gedan, K.B.; Kirwan, M.L.; Wolanski, E.; Barbier, E.B.; Silliman, B.R. The Present and Future Role of Coastal Wetland Vegetation in Protecting Shorelines: Answering Recent Challenges to the Paradigm. Clim. Chang. 2011, 106, 7–29. [Google Scholar] [CrossRef]
- Horstman, E.M.; Dohmen-Janssen, C.M.; Bouma, T.J.; Hulscher, S.J.M.H. Tidal-Scale Flow Routing and Sedimentation in Mangrove Forests: Combining Field Data and Numerical Modelling. Geomorphology 2015, 228, 244–262. [Google Scholar] [CrossRef]
- Van Santen, P.; Augustinus, P.G.E.F.; Janssen-Stelder, B.M.; Quartel, S.; Tri, N.H. Sedimentation in an Estuarine Mangrove System. J. Asian Earth Sci. 2007, 29, 566–575. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove Forests: Resilience, Protection from Tsunamis, and Responses to Global Climate Change. Estuar. Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- McIvor, A.; Spencer, T.; Möller, I.; Spalding, M. The Response of Mangrove Soil Surface Elevation to Sea Level Rise; Natural Coastal Protection Series: Report 3; Cambridge Coastal Research Unit Working Paper 42; University of Cambridge: Cambridge, UK, 2013; p. 59. [Google Scholar]
- Duarte, C.M.; Losada, I.J.; Hendriks, I.E.; Mazarrasa, I.; Marbà, N. The Role of Coastal Plant Communities for Climate Change Mitigation and Adaptation. Nat. Clim Chang. 2013, 3, 961–968. [Google Scholar] [CrossRef]
- Morris, R.; Strain, E.M.A.; Konlechner, T.M.; Fest, B.J.; Kennedy, D.M.; Arndt, S.K.; Swearer, S.E. Developing a Nature-Based Coastal Defence Strategy for Australia. Aust. J. Civ. Eng. 2019, 17, 167–176. [Google Scholar] [CrossRef]
- Morris, R.L.; Konlechner, T.M.; Ghisalberti, M.; Swearer, S.E. From Grey to Green: Efficacy of Eco-Engineering Solutions for Nature-Based Coastal Defence. Glob. Chang. Biol. 2018, 24, 1827–1842. [Google Scholar] [CrossRef]
- Chang, C.-W.; Mori, N. Green Infrastructure for the Reduction of Coastal Disasters: A Review of the Protective Role of Coastal Forests against Tsunami, Storm Surge, and Wind Waves. Coast. Eng. J. 2021, 63, 370–385. [Google Scholar] [CrossRef]
- Spalding, M.; Mcivor, A.; Tonneijck, F.; Tol, S.; van Eijk, P. Mangroves for Coastal Defence. In Guidelines for Coastal Managers & Policy Makers; University of Cambridge: Cambridge, UK, 2014. [Google Scholar]
- McIvor, A.; Spencer, T.; Spalding, M.; Lacambra, C.; Möller, I. Mangroves, Tropical Cyclones, and Coastal Hazard Risk Reduction. In Coastal and Marine Hazards, Risks, and Disasters; Elsevier: Amsterdam, The Netherlands, 2015; pp. 403–429. ISBN 978-0-12-396483-0. [Google Scholar]
- Van Coppenolle, R. Potenties Voor Natuur Gebaseerde Mitigatie Van Overstromingsrisico’s in Kustgebieden Een Regionale Tot Globale Studie; University of Antwerp: Antwerp, Belgium, 2018. [Google Scholar]
- Yudha, R.P. Forest Structure of 26-Year-Old Planted Mangroves. J. Sylva Indones. 2021, 4, 61–69. [Google Scholar] [CrossRef]
- Bouma, T.J.; van Belzen, J.; Balke, T.; Zhu, Z.; Airoldi, L.; Blight, A.J.; Davies, A.J.; Galvan, C.; Hawkins, S.J.; Hoggart, S.P.G.; et al. Identifying Knowledge Gaps Hampering Application of Intertidal Habitats in Coastal Protection: Opportunities & Steps to Take. Coast. Eng. 2014, 87, 147–157. [Google Scholar] [CrossRef]
- Vo Luong, P.; Massel, S. Experiments on Wave Motion and Suspended Sediment Concentration at Nang Hai, Can Gio Mangrove Forest, Southern Vietnam. Oceanologia 2006, 48, 23–40. [Google Scholar]
- Horstman, E.M.; Dohmen-Janssen, C.M.; Narra, P.M.F.; van den Berg, N.J.F.; Siemerink, M.; Hulscher, S.J.M.H. Wave Attenuation in Mangroves: A Quantitative Approach to Field Observations. Coast. Eng. 2014, 94, 47–62. [Google Scholar] [CrossRef]
- Mori, N.; Chang, C.-W.; Inoue, T.; Akaji, Y.; Hinokidani, K.; Baba, S.; Takagi, M.; Mori, S.; Koike, H.; Miyauchi, M.; et al. Parameterization of Mangrove Root Structure of Rhizophora Stylosa in Coastal Hydrodynamic Model. Front. Built Environ. 2022, 7, 782219. [Google Scholar] [CrossRef]
- Yoshikai, M.; Nakamura, T.; Suwa, R.; Argamosa, R.; Okamoto, T.; Rollon, R.; Basina, R.; Primavera-Tirol, Y.H.; Blanco, A.C.; Adi, N.S.; et al. Scaling Relations and Substrate Conditions Controlling the Complexity of Rhizophora Prop Root System. Estuar. Coast. Shelf Sci. 2021, 248, 107014. [Google Scholar] [CrossRef]
- Sippo, J.Z.; Lovelock, C.E.; Santos, I.R.; Sanders, C.J.; Maher, D.T. Mangrove Mortality in a Changing Climate: An Overview. Estuar. Coast. Shelf Sci. 2018, 215, 241–249. [Google Scholar] [CrossRef]
- Spencer, T.; Möller, I. Mangrove Systems. In Treatise on Geomorphology; Elsevier: Amsterdam, The Netherlands, 2013; pp. 360–391. ISBN 978-0-08-088522-3. [Google Scholar]
- Jimenez, J.; Lugo, A.; Cintron, G. Tree Mortality in Mangrove Forests. Biotropica 1985, 17, 177. [Google Scholar] [CrossRef]
- Paling, E.I.; Kobryn, H.T.; Humphreys, G. Assessing the Extent of Mangrove Change Caused by Cyclone Vance in the Eastern Exmouth Gulf, Northwestern Australia. Estuar. Coast. Shelf Sci. 2008, 77, 603–613. [Google Scholar] [CrossRef]
- Cahoon, D.; Hensel, P.; Rybczyk, J.; McKee, K.; Proffitt, E.; Perez, B. Mass Tree Mortality Leads to Mangrove Peat Collapse at Bay Islands, Honduras after Hurricane Mitch. J. Ecol. 2003, 91, 1093–1105. [Google Scholar] [CrossRef]
- Luo, Z.; Huang, B.; Chen, X.; Tan, C.; Qiu, J.; Huang, G. Effects of Wave–Current Interaction on Storm Surge in the Pearl River Estuary: A Case Study of Super Typhoon Mangkhut. Front. Mar. Sci. 2021, 8, 692359. [Google Scholar] [CrossRef]
- Rumsey, D.J. How to Determine the Minimum Size Needed for a Statistical Sample. In Statistics for Dummies; Wiley: Hoboken, NJ, USA, 2016; ISBN 978-1-119-29352-1. [Google Scholar]
- Cao, H.; Chen, Y.; Tian, Y.; Feng, W. Field Investigation into Wave Attenuation in the Mangrove Environment of the South China Sea Coast. J. Coast. Res. 2016, 322, 1417–1427. [Google Scholar] [CrossRef]
- Maza, M.; Lara, J.L.; Losada, I.J. Predicting the Evolution of Coastal Protection Service with Mangrove Forest Age. Coast. Eng. 2021, 168, 103922. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, P.; Gong, Z.; Li, B.; Chen, X. Wave Attenuation by Spartina Alterniflora under Macro-Tidal and Storm Surge Conditions. Wetlands 2020, 40, 2151–2162. [Google Scholar] [CrossRef]
- Jadhav, R.S.; Chen, Q.; Smith, J.M. Spectral Distribution of Wave Energy Dissipation by Salt Marsh Vegetation. Coast. Eng. 2013, 77, 99–107. [Google Scholar] [CrossRef]
- Dalrymple, R.A.; Kirby, J.T.; Hwang, P.A. Wave Diffraction Due to Areas of Energy Dissipation. J. Waterw. Port Coast. Ocean Eng. 1984, 110, 67–79. [Google Scholar] [CrossRef]
- Green, M.O.; Coco, G. Review of Wave-Driven Sediment Resuspension and Transport in Estuaries. Rev. Geophys. 2014, 52, 77–117. [Google Scholar] [CrossRef]
- Krauss, K.W.; Doyle, T.W.; Doyle, T.J.; Swarzenski, C.M.; From, A.S.; Day, R.H.; Conner, W.H. Water Level Observations in Mangrove Swamps during Two Hurricanes in Florida. Wetlands 2009, 29, 142–149. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, H.; Li, Y.; Xu, H.; Shen, J.; Rhome, J.; Smith, T.J., III. The Role of Mangroves in Attenuating Storm Surges. Estuar. Coast. Shelf Sci. 2012, 102, 11–23. [Google Scholar] [CrossRef]
- Montgomery, J.M.; Bryan, K.R.; Horstman, E.M.; Mullarney, J.C. Attenuation of Tides and Surges by Mangroves: Contrasting Case Studies from New Zealand. Water 2018, 10, 1119. [Google Scholar] [CrossRef]
- Paquier, A.-E.; Haddad, J.; Lawler, S.; Ferreira, C.M. Quantification of the Attenuation of Storm Surge Components by a Coastal Wetland of the US Mid Atlantic. Estuaries Coasts 2017, 40, 930–946. [Google Scholar] [CrossRef]
- Peng, Z.; Zou, Q.; Reeve, D.; Wang, B. Parameterisation and Transformation of Wave Asymmetries over a Low-Crested Breakwater. Coast. Eng. 2009, 56, 1123–1132. [Google Scholar] [CrossRef]
- Harada, K.; Imamura, F. Effects of Coastal Forest on Tsunami Hazard Mitigation—A Preliminary Investigation. In Tsunamis; Satake, K., Ed.; Springer: Dordrecht, The Netherlands, 2005; Volume 23, pp. 279–292. [Google Scholar]
- Mendes, D.; Fortunato, A.B.; Bertin, X.; Martins, K.; Lavaud, L.; Nobre Silva, A.; Pires-Silva, A.A.; Coulombier, T.; Pinto, J.P. Importance of Infragravity Waves in a Wave-Dominated Inlet under Storm Conditions. Cont. Shelf Res. 2020, 192, 104026. [Google Scholar] [CrossRef]
- McIvor, A.; Möller, I.; Spencer, T.; Spalding, M. Reduction of Wind and Swell Waves by Mangroves; Natural Coastal Protection Series: Report 1; Cambridge Coastal Research Unit Working Paper 40; University of Cambridge: Cambridge, UK, 2012. [Google Scholar]
- Mendez, F.J.; Losada, I.J. An Empirical Model to Estimate the Propagation of Random Breaking and Nonbreaking Waves over Vegetation Fields. Coast. Eng. 2004, 51, 103–118. [Google Scholar] [CrossRef]
- Lagomasino, D.; Fatoyinbo, T.; Castañeda-Moya, E.; Cook, B.D.; Montesano, P.M.; Neigh, C.S.R.; Corp, L.A.; Ott, L.E.; Chavez, S.; Morton, D.C. Storm Surge and Ponding Explain Mangrove Dieback in Southwest Florida Following Hurricane Irma. Nat. Commun. 2021, 12, 4003. [Google Scholar] [CrossRef]
- McCoy, E.D.; Mushinsky, H.R.; Johnson, D.; Meshaka, W.E. Mangrove Damage Caused by Hurricane Andrew on the Southwestern Coast of Florida. Bull. Mar. Sci. 1996, 59, 1–8. [Google Scholar]
- Yanagisawa, H.; Koshimura, S.; Miyagi, T.; Imamura, F. Tsunami Damage Reduction Performance of a Mangrove Forest in Banda Aceh, Indonesia Inferred from Field Data and a Numerical Model. J. Geophys. Res. Ocean. 2010, 115, C06032. [Google Scholar] [CrossRef]
- Yanagisawa, H.; Koshimura, S.; Goto, K.; Miyagi, T.; Imamura, F.; Ruangrassamee, A.; Tanavud, C. The Reduction Effects of Mangrove Forest on a Tsunami Based on Field Surveys at Pakarang Cape, Thailand and Numerical Analysis. Estuar. Coast. Shelf Sci. 2009, 81, 27–37. [Google Scholar] [CrossRef]
- Hensel, P.; Proffitt, E. Hurricane Mitch: Acute Impacts on Mangrove Forest Structure and an Evaluation of Recovery Trajectories Executive Summary; USGS Open File Report 03-182. 2003. Available online: https://www.usgs.gov/publications/hurricane-mitch-acute-impacts-mangrove-forest-structure-and-evaluation-recovery (accessed on 27 July 2022).
- Hespen, R.; Hu, Z.; Peng, Y.; Borsje, B.W.; Kleinhans, M.; Ysebaert, T.; Bouma, T.J. Analysis of Coastal Storm Damage Resistance in Successional Mangrove Species. Limnol. Oceanogr. 2021, 66, 3221–3236. [Google Scholar] [CrossRef]
- Carlos, C.; Delfino, R.J.; Juanico, D.E.; David, L.; Lasco, R. Vegetation Resistance and Regeneration Potential of Rhizophora, Sonneratia and Avicennia in the Typhoon Haiyan-Affected Mangroves in the Philippines: Implications on Rehabilitation Practices. CDDJ 2015, 1, 1–8. [Google Scholar] [CrossRef]
- Primavera, J.H.; dela Cruz, M.; Montilijao, C.; Consunji, H.; dela Paz, M.; Rollon, R.N.; Maranan, K.; Samson, M.S.; Blanco, A. Preliminary Assessment of Post-Haiyan Mangrove Damage and Short-Term Recovery in Eastern Samar, Central Philippines. Mar. Pollut. Bull. 2016, 109, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Feng, L.; Hu, C.; Kramer, P. Hurricane-Induced Changes in the Everglades National Park Mangrove Forest: Landsat Observations Between 1985 and 2017. J. Geophys. Res. Biogeosci. 2018, 123, 3470–3488. [Google Scholar] [CrossRef]
- Bricker, J.D.; Takagi, H.; Mas, E.; Kure, S.; Adriano, B.; Yi, C.; Roeber, V. Spatial Variation of Damage Due to Storm Surge and Waves during Typhoon Haiyan in the Philippines. J. Jpn. Soc. Civ. Eng. Ser. B2 (Coast. Eng.) 2014, 70, I_231–I_235. [Google Scholar] [CrossRef]
- Lei, J.; Nepf, H. Blade Dynamics in Combined Waves and Current. J. Fluids Struct. 2019, 87, 137–149. [Google Scholar] [CrossRef]
- Zhang, X.; Nepf, H. Flow-Induced Reconfiguration of Aquatic Plants, Including the Impact of Leaf Sheltering. Limnol. Oceanogr. 2020, 65, 2697–2712. [Google Scholar] [CrossRef]
- Zhu, L.; Zou, Q.; Huguenard, K.; Fredriksson, D.W. Mechanisms for the Asymmetric Motion of Submerged Aquatic Vegetation in Waves: A Consistent-Mass Cable Model. J. Geophys. Res. Ocean. 2020, 125, e2019JC015517. [Google Scholar] [CrossRef]
- van Hespen, R.; Hu, Z.; Peng, Y.; Zhu, Z.; Ysebaert, T.; Bouma, T.J. Identifying Trait-based Tolerance to Sediment Dynamics during Seedling Establishment across Eight Mangrove Species. Limnol. Oceanogr. 2022, lno.12202. [Google Scholar] [CrossRef]
- Xie, D.; Schwarz, C.; Kleinhans, M.G.; Zhou, Z.; Maanen, B. Implications of Coastal Conditions and Sea-Level Rise on Mangrove Vulnerability: A Bio-Morphodynamic Modeling Study. JGR Earth Surf. 2022, 127, e2021JF006301. [Google Scholar] [CrossRef]
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.; et al. Ecological Role and Services of Tropical Mangrove Ecosystems: A Reassessment: Reassessment of Mangrove Ecosystem Services. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Laguncularia racemosa | Sonneratia apetala | ||||
|---|---|---|---|---|---|---|
| Mean ± Std | Min–Max | Samplings | Mean ± Std | Min–Max | Samplings | |
| hp (m) | 3.04 ± 0.86 | 1.4–4.8 | 14 | 4.00 ± 0.95 | 2.7–5.2 | 5 |
| Ns (per plant) | 29 ± 10 | 19–39 | 4 | 3.2 ± 1.3 | 2–5 | 5 |
| Ds (mm) | 14.6 ± 11.5 | 4–61 | 115 | 79 ± 20 | 46–131 | 16 |
| LB (cm) | 92 ± 26 | 58–128 | 16 | 195 ± 62 | 142–335 | 11 |
| DB (mm) | 11.7 ± 3.6 | 6.9–15.6 | 16 | 44 ± 10 | 31–61 | 16 |
| SB (cm) | 11.9 ± 4.5 | 7–22 | 16 | 17 ± 11 | 6–47 | 16 |
| LT (cm) | 32 ± 12 | 19–50 | 6 | 109 ± 20 | 72–132 | 8 |
| DT (mm) | 4.7 ± 2.7 | 2–8 | 6 | 21 ± 6 | 13–31 | 16 |
| ST (cm) | 7 ± 3 | 3–10 | 6 | 19 ± 9 | 4–38 | 16 |
| Nl (per twig) | 13 ± 3 | 8–18 | 6 | 169 ± 64 | 64–256 | 16 |
| Ll (cm) | 8.0 ± 1.4 | 6.4–10.6 | 16 | 7.7 ± 1.0 | 6.4–10 | 16 |
| Wl (cm) | 3.4 ± 0.4 | 2.9–4.1 | 16 | 2.3 ± 0.3 | 1.8–2.7 | 16 |
| Nr (m−2) | 144 ± 28 | 122–186 | 4 | |||
| hr (cm) | 11 ± 3 | 7–18 | 16 | |||
| Dr (mm) | 9 ± 2 | 5–12 | 16 | |||
| Gage | Brand | Measuring Range | Sampling Rate | Storage | Continuous Working Time | Operating Temperature | Sensor Height from Bottom |
|---|---|---|---|---|---|---|---|
| A | Aanderaa Seaguard WLR (Norway) | 0 to 60 Mpa | 4 Hz | 2 Gb | 6 months | −4 to 36 °C | 35 cm |
| B and C | RBRduet T.D|tide (Canada) | 0 to 20 m | 2 Hz | 2 Gb | 3 years | −5 to 35 °C | 24 cm |
| D | YPS600-J (China) | 0 to 3 m | 20 Hz | 8 Gb | 120 h | −40 to 60 °C | 2 cm |
| Station | hmax (m) | hmean (m) | Hmax (m) | ||||
|---|---|---|---|---|---|---|---|
| A | 2.54 | 2.18 | 5.6 | 1.16 | 0.57 | 0.60 | 0.52 |
| 33% | 43% | 26% | 56% | ||||
| B | 1.19 | 0.84 | 4.6 | 0.69 | 0.28 | 0.40 | 0.18 |
| 35% | 59% | 50% | 76% | ||||
| C | 1.07 | 0.71 | 4.1 | 0.55 | 0.17 | 0.27 | 0.09 |
| 77% | 67% | 66% | 67% | ||||
| D | 0.96 | 0.69 | 7.8 | 0.11 | 0.05 | 0.08 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Lin, P.; Chen, X. Coastal Protection by Planted Mangrove Forest during Typhoon Mangkhut. J. Mar. Sci. Eng. 2022, 10, 1288. https://doi.org/10.3390/jmse10091288
Zhang X, Lin P, Chen X. Coastal Protection by Planted Mangrove Forest during Typhoon Mangkhut. Journal of Marine Science and Engineering. 2022; 10(9):1288. https://doi.org/10.3390/jmse10091288
Chicago/Turabian StyleZhang, Xiaoxia, Pengzhi Lin, and Xinping Chen. 2022. "Coastal Protection by Planted Mangrove Forest during Typhoon Mangkhut" Journal of Marine Science and Engineering 10, no. 9: 1288. https://doi.org/10.3390/jmse10091288
APA StyleZhang, X., Lin, P., & Chen, X. (2022). Coastal Protection by Planted Mangrove Forest during Typhoon Mangkhut. Journal of Marine Science and Engineering, 10(9), 1288. https://doi.org/10.3390/jmse10091288