Abstract
The Devonian–Carboniferous transition was marked by a series of perturbations in the geological and biological evolution. The palaeontological data from Transcaucasia allowed the bryozoan diversity dynamics on the northern Gondwanan margin (southern periphery of the Palaeo-Tethys Ocean) to be documented at this transition. Taxonomic ranges of 43 species, 26 genera, 19 families, and 4 orders were analysed to reveal changes in the total diversity, the number of appearances, the number of disappearances, and the turnover rates per substages. It is established that the bryozoan diversity was rather high in the beginning and the end of the Famennian, as well as in the Late Tournaisian. It declined significantly in the Middle–Late Famennian and the Early Tournaisian due to the combination of the high number of disappearances and the low number of appearances. The turnovers remained strong, and they peaked in the mid-Famennian. These regionally documented diversity changes match the patterns recorded globally and in Southern Siberia. Hypothetically, the Middle–Late Famennian and Early Tournaisian crises established in Transcaucasia were related to the global events (anoxia and mass extinctions), a series of which weakened the bryozoans’ resistivity to negative external influences.
1. Introduction
The Devonian–Carboniferous transition (~370–350 Ma) was one of the most critical intervals in the Palaeozoic geological and biological evolution. It was marked by anoxia [1,2,3], glacial advance [4,5], sea-level drop [6], and mass extinction [7,8,9]. However, many aspects of these events and their spatio-temporal appearance are yet to be fully understood, and new investigations bring “surprises” [10,11]. In order to document better the biotic events at the Devonian–Carboniferous transition, attention should be paid to the patterns of the diversity dynamics of many fossil groups and in many regions. Of special interest are organisms, preliminary investigations of which revealed uncertain reactions to the mid-Palaeozoic global crises, and regions, where diversity changes of these organisms were not studied earlier. In regard to these needs, investigation of Famennian–Tournaisian bryozoans from Transcaucasia seems to be urgent.
The previous analyses of diversity dynamics of bryozoans revealed their resistivity to the mid-Palaeozoic mass extinctions [8,12,13], which itself is important to consider when these biotic crises are addressed (notably, bryozoans were stressed significantly by the later catastrophes [14]). However, it remains unclear whether this resistivity is a global phenomenon. The Devonian–Carboniferous bryozoans have been studied in Transcaucasia for more than three decades [15,16,17]. A significant amount of the accumulated information would serve ideally for measuring various patterns of the bryozoan diversity dynamics. This regional record represents the northern Gondwanan margin where the knowledge of middle Palaeozoic bryozoans remains limited [18,19,20]. “The Paleobiology Database” [21] demonstrates the outstanding scarcity of the information about Devonian and Carboniferous bryozoans from the African-Arabian margin of mid-Palaeozoic Gondwana, which means the present knowledge of these organisms is seriously biased (the data from Transcaucasia are also absent in this source).
The objective of the present paper is to reconstruct the bryozoan diversity dynamics in Transcaucasia at the Devonian–Carboniferous transition. This is necessary to extend the general knowledge of bryozoan evolution at this transition with the information from the northern Gondwanan margin.
2. Materials and Methods
2.1. Study Area
The study area is located in the southern part of Transcaucasia (Figure 1). The latter has three meanings, namely administrative (Georgia, Armenia, and Azerbaijan taken together), geographical (Lesser Caucasus mountains and adjacent Rioni and Kura depressions), and geological (Lesser Caucasus as “exotic” terrane). The study area corresponds to the noted terrane (see the works [22,23] for more tectonic considerations).
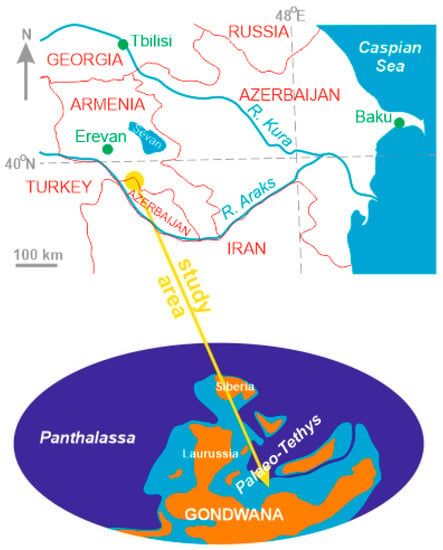
Figure 1.
Present and Famennian location of the study area (created by the authors; the palaeogeographical reconstruction follows the contours shown by [24]).
The local stratigraphical framework was characterized by Grechishnikova and Levitskii [25], Mamedov [26,27], and Rzhonsnitskaya and Mamedov [20]. The transitional Devonian–Carboniferous deposits are represented in a dozen of closely related sections and smaller outcrops, which form together the full succession of the Famennian–Tournaisian strata. These are intercalating coarse and fine siliciclastics and limestones, with different proportions in different intervals (Figure 2). The total thickness reaches 900 m. Although formations have been introduced, the deposits are usually subdivided by biozones and substages (also in individual sections—for instance, see [25]), and thus, formations are not used in the present study. These strata bear abundant fossils, including marine macroinvertebrates. Brachiopods and conodonts are especially useful to establish the age with precision and to undertake accurate zonal correlations [22,25,26,27,28]. These deposits accumulated in shelf environments, with significant flux of siliciclastic matter and under slightly changed sea level [25].
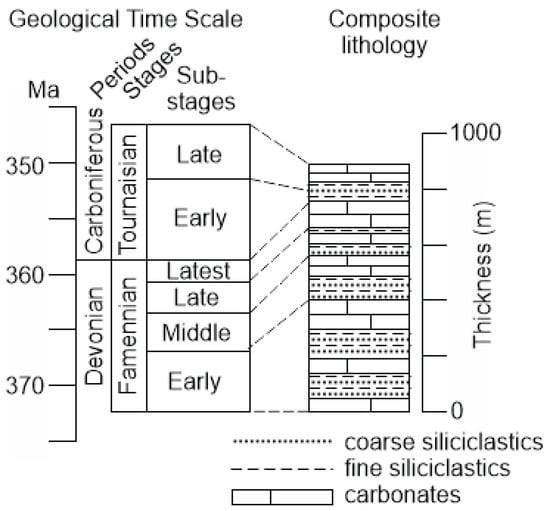
Figure 2.
Stratigraphical framework of the Devonian–Carboniferous transition in the study area (created by the authors; the geological time scale follows [29], and the regional stratigraphy is compiled from [25,26,27]).
The palaeogeographical position of the study area can be derived from present interpretations of the pre-Jurassic history of the Lesser Caucasus terrane. Taking into account its relative position to the Greater Caucasus, which is thought to be a Galatian terrane [30,31], and the trajectories of the Middle Eastern blocks depicted on modern plate tectonic reconstructions [24,32,33,34,35,36,37], it appears that the Lesser Caucasus was a part of the northern (African-Arabian) Gondwanan margin until the Cimmerian terranes were detached from it in the late Palaeozoic together with the opening of the Neo-Tethys to reach the Laurasian margin in the early Mesozoic. If so, marine sedimentation in the study area can be attributed to the southern periphery of the Palaeo-Tethys Ocean, where there were also epeiric seas ingressed in Gondwana. The plate tectonic reconstructions noted above imply that the study area lied in the tropical latitudes and, most probably, in the Southern Hemisphere (Figure 1).
2.2. Palaeontological Information
The material for the present analysis was prepared in three steps. First, the sources devoted to the Famennian–Tournaisian bryozoans of the study area were collected. These included the works by Goryunova [15,38,39], Gorjunova and Lavrentjeva [40], Grechishnikova and Levitskii [25], Lavrentjeva [16,41], and Tolokonnikova [17,42]. These works summarize the outcomes of the previous research and make the considered palaeontological record suitable to the quantitative analysis. Importantly, the palaeontological data were collected in the “Soviet traditions”, which means sampling was massive and aimed initially at representing the record of the entire area, not particular localities. Such an approach makes these data very suitable for diversity analysis.
Second, the stratigraphic ranges of all bryozoan taxa were compiled from the sources indicated above to compose a database for further quantitative analysis. For this purpose, the regional stratigraphical framework [22,25,26,27,28] was brought in correspondence to the modern version of the geological time scale [29,43] and, particularly, the stratigraphy of the Devonian–Carboniferous transition [1,44,45,46,47,48,49]. The presence of bryozoan taxa in the four substages of the Famennian and the two substages of the Tournaisian was established. Indeed, the previously published data are heterogeneous, and some generalizations were required to avoid inconsistencies on the below-substage level. One can question whether the employed palaeontological data represent each substage adequately. The related uncertainties are generally unavoidable in palaeobiological studies because in only rare cases fossils of the same kind can be found in all layers of the studied stratigraphical interval. Nonetheless, it should be said that palaeontological collecting in the study area was massive, and thus, the bryozoan representation of each stage seems to be adequate. If bryozoans were not found at some levels, this means they did not exist, and thus, the diversity of the related substage was relatively small because there were environments unfavourable for bryozoans. At least, the sampling of the examined Famennian–Tournaisian succession during the previous studies, from which we compile the information, was massive and rather homogeneous.
Third, the preliminary information was reviewed and systematized to update the suprageneric taxonomy, to avoid synonyms, and to justify the stratigraphic ranges of species. This way, the final version of the database is prepared (Supplementary Materials). It registers the stratigraphic ranges of 43 bryozoan species, 26 genera, 19 families, and 4 orders in the study area.
2.3. Diversity Dynamics Analysis
The present analysis pays attention to two aspects of the bryozoan diversity dynamics at the Devonian–Carboniferous transition, namely principal patterns and turnover rates. The principal patterns include total diversity (total number of taxa), number of appearances, and number of disappearances, as done in previous investigations (for instance, [12]). The three principal patterns are measured for each substage (six time slices correspond to four Famennian and two Tournaisian substages) and four taxonomic levels of bryozoans, namely species, genera, families, and orders. Importantly, the number of appearances cannot be measured for the first time slice due to the absence of information from the preceding slice, and the number of disappearances cannot be measured for the last time slice due to the absence of information from the following slice.
Notably, appearances and disappearances should be distinguished from originations and extinctions because the former can be only regional (for instance, a given taxon could originate earlier in other regions) and/or temporary (for instance, a given taxon could disappear due to anoxia and then re-appear). Attention to the numbers of appearances and disappearances is reasonable when local palaeontological records are considered.
The turnover rates are measured by similarity indices using the approach tested previously [50]. The R index is the well-known Jaccard’s similarity [51]. It is calculated for each two successive pairs of assemblages of bryozoan species, genera, families, and orders. The RST index is more complex because it reflects similarity of assemblages of higher-ranked taxa taking into account their diversity, i.e., the number of the related lower-ranked taxa. Two indices, namely the Czekanowski’s quantified coefficient (C) [52] and the Gower’ index (G) [53] can be employed. As they reflect essentially the same pattern, but on the basis of different mathematical procedures, they are mutually important, and thus, RST (C) and RST (G) are calculated for each of two successive pairs of assemblages of bryozoan orders taking into account their species diversity. All three indices (R, RST (C), and RST (G)) change from 0 to 1, where 0 indicates total turnover, and 1 indicates absence of any turnover. R reflects simple turnover rate, and RST (C) and RST (G) reflect turnovers in the taxonomic structure of diversity. Additionally, it appears reasonable to measure RST (C) and RST (G) for not only successive, but all possible pairs of bryozoan assemblages, which would permit us to understand whether a given turnover had relatively long-term effects. It cannot be excluded that the oldest and youngest assemblages demonstrate high similarity despite of strong turnovers in-between, which means some orders “re-gained” their importance for the species diversity after a certain pause.
3. Results
3.1. Principal Patterns
The diversity of bryozoans in the study area was measured for six geological time slices (Table 1), and this information allows for judgments of the principal patterns of its dynamics. The total diversity of species, genera, families, and orders changed rather similarly across the Devonian–Carboniferous transition (Figure 3). After the relatively high diversity in the beginning of the Famennian, a pronounced decline occurred in the midst of this stage. Diversification near its end was followed by small diversity drop (not registered for families) in the Early Tournaisian, after which a strong diversification happened (the latter did not involve orders).

Table 1.
Taxonomic diversity of the studied bryozoans.
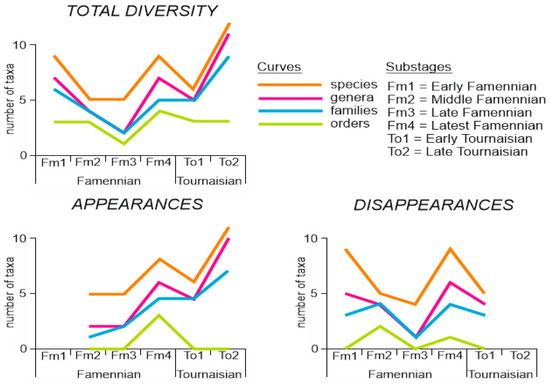
Figure 3.
Principal patterns of the taxonomic diversity dynamics of the studied bryozoans (created by the authors, see Table 1 for numbers).
The numbers of appearances rose synchronously in the Latest Famennian (Figure 3). They dropped slightly in the Early Tournaisian and rose significantly in the second part of this stage (this did not happen with orders). The numbers of disappearances were high in the Early and Latest Famennian, but low in the Late Famennian (Figure 3). The comparison of the principal patterns implies that the mid-Famennian total diversity decline occurred due to the high number of disappearances in the Early Famennian and the lower number of appearances in the Middle and Late Famennian. The Early Tournaisian total diversity drop was driven more by the high number of disappearances in the Latest Famennian.
3.2. Turnover Rates
The R, RST (C), and RST (G) indices are calculated for five successive pairs of assemblages corresponding to the substages, as well as to the non-successive assemblages (Table 2). The turnovers established with R were especially strong at the Middle–Late Famennian transition (Figure 4). The assemblages changed totally (except for orders). Another strengthening of turnovers occurred at the Latest Famennian–Early Tournaisian transition, but it was less pronounced than previously for genera, families, and orders. One should also note the relatively high rate of turnovers throughout the entire considered time interval.
Table 2.
R and RST indices for the studied bryozoan assemblages.
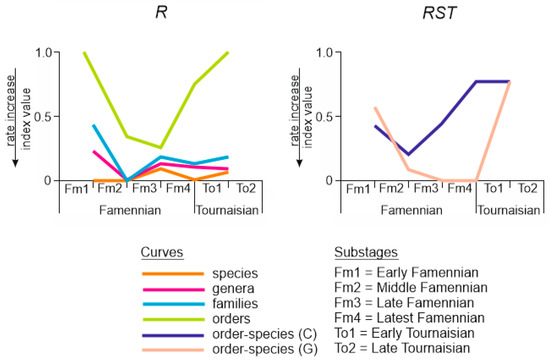
Figure 4.
Rates of changes among the assemblages of the studied bryozoans (created by the authors, see Table 2 for numbers).
The turnovers in the order–species structure of bryozoan assemblages are revealed with less certainty (Figure 4). Both RST (C) and RST (G) indicate significant turnover at the Middle–Late Famennian transition, after which the intensity of turnovers either decreased gradually (RST (C)) or remained high (RST (G)) (Figure 4). Both indices indicate moderate-to-low turnovers within the Tournaisian. This pattern resembles what was registered by R (Figure 4). Consideration of non-successive pairs of assemblages indicates high similarity of the Early Famennian and Late Tournaisian assemblages measured by RST (C) and RST (G) (Table 2). This means limited effects of the Middle Famennian–Early Tournaisian turnovers on the order–species structure of the bryozoan diversity at the Devonian–Carboniferous transition.
4. Discussion
4.1. Global and Regional Records
Globally, the total species diversity of bryozoans changed when measured per stages: Horowitz and Pachut [54] noted 153 species in the Famennian and 405 species in the Tournaisian. “The Paleobiology Database” [21] indicates 160 genera in the Famennian and 154 genera in the Tournaisian, which means the almost comparable total generic diversity. The data from Transcaucasia (Supplementary Materials) implies the presence of 26 species and 15 genera in the Famennian and 17 species and 15 genera in the Tournaisian, which indicates on the impoverished species diversity and the unchanged generic diversity. The global and regional generic diversity changes at the Devonian–Carboniferous transition seem to be the same. If the estimates of the global species diversity by Horowitz and Pachut [54] remain correct, striking difference between the global and regional diversity changes should be explained anyhow (for instance, by the stronger regional biotic crises). However, these estimates are, most probably, outdated to certain degree because of accumulation of more palaeontological information in the beginning of the 21st century. If so, we should focus on the only significant similarity of the global and regional generic diversity changes (see above).
A detailed analysis of the Late Devonian bryozoan diversity dynamics was carried out in Southern Siberia [12]. The outcomes from this analysis indicate the same principal patterns as registered in Transcaucasia (Figure 3). Particularly, the total number of species, genera, and families was relatively high in both regions in the beginning and the end of the Famennian, and the mid-Famennian diversity decline was sharp. Such a similarity is somewhat surprising in regard to the big distance between Southern Siberia and Transcaucasia in the Late Devonian (Figure 1). The latter was located on the southern periphery of the Palaeo-Tethys, whereas the former was very far from the northern periphery of this ocean. If so, the similarity of the Famennian diversity dynamics of bryozoans between these regions reveals some general, global mechanisms discussed below.
4.2. Influence of Biotic Crises
The results of the present analysis allow for the recognition of two crises in the development of bryozoans in Transcaucasia at the Devonian–Carboniferous transition. The first encompasses the Middle–Late Famennian, and the second took place in the Early Tournaisian. These regional crises can be brought in correspondence to the global events at this transition. Their nomenclature was summarized by Becker et al. [1]. In addition to the “purely” Famennian events, it is necessary to consider the Fasnian/Famennian mass extinction and the related anoxia known as the Lower and Upper Kellwasser events [55,56,57,58,59]. A comparison of the two regional bryozoan crises and the global events reveals their complex temporal relationship (Figure 5). The bryozoan diversity was rather high in the Early Famennian. The subsequent global events in the Early Famennian were followed by the regional bryozoan crisis, and the latter could have been facilitated by the other global events in the Late Famennian. Similarly, the Hangenberg event and the related mass extinction [2,7,8,9,58] were followed by the bryozoan crisis in the beginning of the Tournaisian.
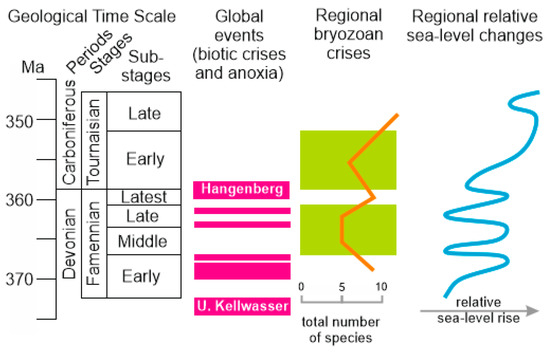
Figure 5.
Correlation of the global and regional events (created by the authors; the geological time scale follows [29], the global events are shown according to [1], the regional bryozoan events are interpreted in this study (see text for more explanations), and the regional relative sea-level changes follow the reconstruction [25] and taking into account the information from [27]). All shown patterns are justified to substages.
Generally, it appears that there is some causal relationship between the global events and regional crises, and the above-mentioned similarity of the records of Transcaucasia and South Siberia also proves the idea of some general mechanisms. However, in this case, it is notable that the Frasnian/Famennian mass extinction did not stress bryozoans so strongly [8,12,13]. It is possible to hypothesize that the only series of several, closely happened events would stress bryozoan assemblages so as to make them vulnerable to subsequent negative influences (Figure 5). This hypothesis deserves further verification with data from different regional records.
It should be noted that the bryozoan diversity dynamics can be calculated only per entire substages. If so, one would wonder whether the established patterns may be influenced by the different duration of these time units. The present version of the geological time scale [29,43] implies that the Early Famennian was longer than the other Famennian substages (Figure 2). However, the latter were more or less comparable in duration, although the diversity differed significantly (Table 1). Moreover, the inequity of the Tournaisian substages cannot explain their differences in diversity (the shorter substage was characterized by the higher number of taxa). If so, the factor of the substage duration does not seem to be significant.
It is also necessary to pay attention to the regional palaeoenvironmental factors, namely relative sea-level changes and oxygen depletion. Sea level fluctuated in the Famennian, and trend to deepening established in the Tournaisian (Figure 5). The comparison of these fluctuations with the total diversity changes (Figure 3) indicates the absence of any clear correspondence. Moreover, it is very notable that the total generic diversity is equal in the Famennian and the Tournaisian (when calculated per stages—see above), although the sea was generally deeper in the latter. However, one may wonder whether the decline of species diversity from 26 species in the Famennian and 17 species in the Tournaisian can be linked to the noted deepening. As for anoxia, geochemical investigations are yet to be conducted in the study area. However, grey-to-black shales are noted among the Famennian deposits [25,27]. Their presence permits us to hypothesize episodes of oxygen depletion, which generally correspond to the multiple Famennian anoxic events [1]. Indeed, geochemical investigations coupled with high precision dating of potential oxygen depletion beds are necessary in the study area for their accurate correlations. However, if even the global anoxia did not leave signatures in the study area, bryozoans could be stressed by the global events. They evolved in the open sea connected to the world’s water masses (Figure 1), and thus, the global-scale collapses of fossil communities linked to anoxia and mass extinctions were able to stress organisms even where the palaeoenvironmental triggers of the stress did not “work”.
4.3. Morphological Notes on Cryptostomata
Representatives of the Order Cryptostomata played important role in the bryozoan assemblages in Transcaucasia at the Devonian–Carboniferous transition; several genera and species appeared in the study area in the Famennian and the Tournaisian (Supplementary Materials). Preparation of the material for the purposes of the present study has permitted to note some interesting morphological peculiarities of the originated taxa. Mentioning these peculiarities together with diversity dynamics is reasonable to demonstrate that the changes considered in the present study associated with some peculiarities of the development of bryozoans.
New cryptostomate bryozoans were characterized by tiny little, two layer-symmetrical or branching colonies 1–2 mm in size, which are spiral in plan. Evidently, such morphological adaptations as bundle of axial zooids (Pseudoascopora) and mesotheca (Geranopora, Mysticella, and Taeniodictya) facilitated strength and stability of the colonies growing up. The colonies allowed cryptostomate bryozoans to inhabit small places in densely populated palaeoecosystems. In order to decrease competition with bryozoans of other orders and other benthic organisms, cryptostomate bryozoans implemented the strategy of increase in the efficiency of feeding via highly regular location of apertures by diagonally crossed rows in all originated genera and development of monticules in Taeniodictya.
Apparently, bryozoans possessed a specific form of camera (chamber with an arched roof) of autozooids and actinotostyles for prohibiting their colonies from predators. A knee-curved camera of autozooids with lower and/or upper hemisepta developed at the edge of the inflection protected soft body of polypide from any invasion. Many originated genera (except for Taeniodictya) demonstrate development of actinotostyles with different size and frequency in the exozone of colonies. These often bounded each autozooid in significant amount for creation of a kind of barrier (“stockade”). All new taxa have a developed network of microstyles linking structural elements of colonies for better feeding and gas exchange. For scaring away predators, regulating water flows, and swamping with sand, colonies beard crests and ribs (Geranopora, Mysticella, and Taeniodictya) separating rows of apertures in local lows.
5. Conclusions
The present study of the bryozoan diversity dynamics in Transcaucasia at the Devonian–Carboniferous transition allows making three conclusions. First, the regional bryozoan assemblages experienced significant changes in the total diversity, the number of appearances, and the number of disappearances during the Famennian–Tournaisian, and the turnover rate remained chiefly high. Second, there were two crises in the regional development of bryozoans, namely the Middle–Late Famennian and Early Tournaisian crises. Third, a series of global events could decrease the resistivity of bryozoans to palaeoenvironmental stress, and thus, the established regional crises can be related hypothetically to widespread anoxic events and mass extinctions.
This paper stresses the importance of regional palaeontological records for deciphering mechanisms of critical transitions in the Earth’s history. Further investigations of bryozoan diversity dynamics are necessary in different regions (not only remote from one another, but also located in different palaeoenvironmental conditions) in order to understand the influence on the global perturbations at the Devonian–Carboniferous transition on this fossil group. More investigations in the other parts of northern Gondwanan margin can improve the knowledge of mid-Palaeozoic bryozoans from this palaeogeographical domain.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jmse10070959/s1, Supplement: Taxonomic ranges of the Famennian–Tournaisian bryozoans from the study area; see text for sources, numbers: 1—presence, 0—absence.
Author Contributions
Conceptualization, Z.A.T. and D.A.R.; methodology, D.A.R.; data curation, Z.A.T. and D.A.R.; formal analysis, Z.A.T. and D.A.R.; writing, Z.A.T. and D.A.R. All authors have read and agreed to the published version of the manuscript.
Funding
The work by Z.A.T. is supported by RSF (project 22-27-00030).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Becker, R.T.; Königshof, P.; Brett, C.E. Devonian climate, sea level and evolutionary events: An introduction. Geol. Soc. Lond. Spec. Publ. 2016, 423, 1–10. [Google Scholar] [CrossRef]
- Caplan, M.L.; Bustin, R.M. Devonian-Carboniferous Hangenberg mass extinction event, widespread organic-rich mudrock and anoxia: Causes and consequences. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1999, 148, 187–207. [Google Scholar] [CrossRef]
- Zhang, F.; Dahl, T.W.; Lenton, T.M.; Luo, G.; She, S.; Algeo, T.J.; Planavsky, N.; Liu, J.; Cui, Y.; Qie, W.; et al. Extensive marine anoxia associated with the Late Devonian Hangenberg Crisis. Earth Planet. Sci. Lett. 2020, 533, 115976. [Google Scholar] [CrossRef]
- Scotese, C.R.; Song, H.; Mills, B.J.W.; van der Meer, D.G. Phanerozoic paleotemperatures: The earth’s changing climate during the last 540 million years. Earth-Sci. Rev. 2021, 215, 103503. [Google Scholar] [CrossRef]
- Streel, M.; Caputo, M.V.; Loboziak, S.; Melo, J.H.G. Late Frasnian-Famennian climates based on palynomorph analyses and the question of the Late Devonian glaciations. Earth-Sci. Rev. 2000, 52, 121–173. [Google Scholar] [CrossRef]
- Haq, B.U.; Schutter, S.R. A chronology of Paleozoic sea-level changes. Science 2008, 322, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Assemat, A.; Girard, C.; Joachimski, M.M.; Adnet, S. Vertebrate diversity reveals perturbations in faunal communities prior to the Hangenberg event in the Montagne Noire (France). Bull. Geosci. 2022, 97, 109–122. [Google Scholar] [CrossRef]
- Kaiser, S.I.; Aretz, M.; Becker, R.T. The global Hangenberg Crisis (Devonian–Carboniferous transition): Review of a first-order mass extinction. Geol. Soc. Lond. Spec. Publ. 2016, 423, 387–437. [Google Scholar] [CrossRef] [Green Version]
- Racki, G. A volcanic scenario for the Frasnian–Famennian major biotic crisis and other Late Devonian global changes: More answers than questions? Glob. Planet. Chang. 2020, 189, 103174. [Google Scholar] [CrossRef]
- Li, S.; Wignall, P.B.; Poulton, S.W.; Hedhli, M.; Grasby, S.E. Carbonate shutdown, phosphogenesis and the variable style of marine anoxia in the late Famennian (Late Devonian) in western Laurentia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 589, 110835. [Google Scholar] [CrossRef]
- Young, A.; Flament, N.; Williams, S.E.; Merdith, A.; Cao, X.; Müller, R.D. Long-term Phanerozoic sea level change from solid Earth processes. Earth Planet. Sci. Lett. 2022, 584, 117451. [Google Scholar] [CrossRef]
- Gutak, J.M.; Tolokonnikova, Z.A.; Ruban, D.A. Bryozoan diversity in Southern Siberia at the Devonian-Carboniferous transition: New data confirm a resistivity to two mass extinctions. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 264, 93–99. [Google Scholar] [CrossRef]
- Tolokonnikova, Z.A.; Ernst, A. Richness of Famennian-Tournaisian (Late Devonian-Eraly Carboniferous) bryozoans in shallow areas of Palaeotethys and Palaeoasian oceans. Palaeobiodiversity Palaeoenvironments 2021, 101, 885–906. [Google Scholar] [CrossRef]
- Powers, C.M.; Bottjer, D.J. The effects of mid-Phanerozoic environmental stress on bryozoan diversity, paleoecology, and paleogeography. Glob. Planet. Chang. 2009, 65, 146–154. [Google Scholar] [CrossRef]
- Gorjunova, R.V. New cryptostomide bryozoans from the Upper Devonian of Transcaucasia and some aspects of the evolution of the order Cryptostomida. Paleontol. J. 2007, 41, 600–613. [Google Scholar] [CrossRef]
- Lavrentjeva, V.D. Upper Devonian Bryozoans of Transcaucasia. Izvestiya vuzov. Geologiya i Razvedka 1985, 8, 12–18. (In Russian) [Google Scholar]
- Tolokonnikova, Z.A. New data on the Tournaisian bryozoans (Lower Carboniferous) from Azerbaijan and Armenia. Paleontol. J. 2016, 50, 388–395. [Google Scholar] [CrossRef]
- Ernst, A.; Kora, M.; El-Desouky, H.; Herbig, H.-G.; Wyse Jackson, P.N. Stenolaemate bryozoans from the Carboniferous of Egypt. J. Afr. Earth Sci. 2020, 165, 103811. [Google Scholar] [CrossRef]
- Sakagami, S.; Sciunnach, D.; Garzanti, E. Late Paleozoic and Triassic bryozoans from the Tethys Himalaya (N India, Nepal and S Tibet). Facies 2006, 52, 279–298. [Google Scholar] [CrossRef]
- Webb, G.E. Quantitative analysis and paleoecology of earliest Mississippian microbial reefs, Gudman Formation, Queensland, Australia: Not just post-disaster phenomena. J. Sediment. Res. 2005, 75, 877–896. [Google Scholar] [CrossRef]
- The Paleobiology Database. Available online: https://paleobiodb.org (accessed on 23 May 2022).
- Aristov, V.A. Conodonts of the Devonian–Lower Carboniferous of Eurasia: Assemblages, zonation, correlation of polyfacial deposits. Trudy GIN RAN 1994, 484, 1–192. (In Russian) [Google Scholar]
- Rustamov, M.I. Plate tectonics of Kur depression within Meso-Tethys geodynamic evolution. ANAS Trans. Earth Sci. 2018, 2, 57–71. (In Russian) [Google Scholar] [CrossRef]
- Kocsis, Á.T.; Scotese, C.R. Mapping paleocoastlines and continental flooding during the Phanerozoic. Earth-Sci. Rev. 2021, 213, 103463. [Google Scholar] [CrossRef]
- Grechishnikova, I.A.; Levitskii, E.S. The Famennian–Lower Carboniferous reference section Geran-Kalasi (Nakhichevan Autonomous Region, Azerbaijan). Stratigr. Geol. Correl. 2011, 19, 21–43. [Google Scholar] [CrossRef]
- Mamedov, A.B. Zonal subdivision of the Tournaisian Stage of the Lower Carboniferous of the Nakhichevan ASSR. Trudy Akademii nauk Azerbaidzhanskoy SSR 1980, 12, 53–57. (In Russian) [Google Scholar]
- Mamedov, A.B. The Famennian stage of the southern Transcaucasia. Stratigr. Geol. Correl. 2002, 10, 57–68. [Google Scholar]
- Rzhonsnitskaya, M.A.; Mamedov, A.B. Devonian stage boundaries in the southern Transcaucasus. CFS Cour. Forsch. Senckenberg 2000, 225, 329–333. [Google Scholar]
- International Commission on Stratigraphy (ICS). International Chronostratigraphic Chart. 2022. Available online: https://stratigraphy.org/chart (accessed on 24 May 2022).
- Ruban, D.A. The Greater Caucasus—A Galatian or Hanseatic Terrane? Comment on “The formation of Pangea” by G.M. Stampfli, C. Hochard, C. Vérard, C. Wilhem and J. von Raumer [Tectonophysics 593 (2013) 1–19]. Tectonophysics 2013, 608, 1442–1444. [Google Scholar] [CrossRef]
- Ruban, D.A.; Al-Husseini, M.I.; Iwasaki, Y. Review of Middle East Paleozoic plate tectonics. GeoArabia 2007, 12, 35–56. [Google Scholar] [CrossRef]
- Cao, W.; Zahirovic, S.; Flament, N.; Williams, S.; Golonka, J.; Müller, R.D. Improving global paleogeography since the late Paleozoic using paleobiology. Biogeosciences 2017, 14, 5425–5439. [Google Scholar] [CrossRef] [Green Version]
- Domeier, M.; Torsvik, T.H. Plate tectonics in the late Paleozoic. Geosci. Front. 2014, 5, 303–350. [Google Scholar] [CrossRef] [Green Version]
- Matthews, K.J.; Maloney, K.T.; Zahirovic, S.; Willims, S.E.; Seton, M.; Müller, R.D. Global plate boundary evolution and kinematics since the late Paleozoic. Glob. Planet. Chang. 2016, 146, 226–250. [Google Scholar] [CrossRef]
- Metcalfe, I. Multiple Tethyan ocean basins and orogenic belts in Asia. Gondwana Res. 2021, 100, 87–130. [Google Scholar] [CrossRef]
- Van Hinsbergen, D.J.J.; Torsvik, T.H.; Schmid, S.M.; Matenco, L.C.; Maffione, M.; Vissers, R.L.M.; Gürer, D.; Spakman, W. Orogenic architecture of the Mediterranean region and kinematic reconstruction of its tectonic evolution since the Triassic. Gondwana Res. 2020, 81, 79–229. [Google Scholar] [CrossRef]
- Williams, S.; Wright, N.M.; Cannon, J.; Flament, N.; Müller, R.D. Reconstructing seafloor age distributions in lost ocean basins. Geosci. Front. 2021, 12, 769–780. [Google Scholar] [CrossRef]
- Goryunova, R.V. Morphology, System and Phylogeny of Bryozoans (Order Rhabdomesida); Nauka: Moscow, Russia, 1985; p. 151. (In Russian) [Google Scholar]
- Gorjunova, R.V. Family Coelotubuliporidae fam. nov. and morphological parallelism in the evolution of bryozoans. Paleontol. J. 2011, 45, 510–624. [Google Scholar] [CrossRef]
- Gorjunova, R.V.; Lavrentjeva, V.D. New bryozoans from the Devonian-Carboniferous boundary beds of Transcaucasia. Paleontol. J. 2007, 41, 146–155. [Google Scholar] [CrossRef]
- Lavrentjeva, V.D. New bryozoan species of the family Atactotoechidae from the Devonian of Transcaucasia. Paleontol. J. 2001, 35, 152–156. [Google Scholar]
- Tolokonnikova, Z.A. New Famennian bryozoans (Upper Devonian) from Azerbaijan and Armenia. Paleontol. J. 2016, 50, 245–254. [Google Scholar] [CrossRef]
- Gradstein, F.M.; Ogg, J.G.; Schmitz, M.; Ogg, G. (Eds.) Geologic Time Scale 2020; Elsevier: Amsterdam, The Netherlands, 2020; p. 1390. [Google Scholar]
- Aretz, M.; Corradini, C.; Denayer, J. The Devonian-Carboniferous Boundary around the globe: A complement. Palaeobiodiversity Palaeoenvironments 2021, 101, 633–662. [Google Scholar] [CrossRef]
- Devuyst, F.-X.; Hance, L.; Hou, H.; Wu, X.; Tian, S.; Coen, M.; Sevastopulo, G. A proposed Global Stratotype Section and Point for the base of the Viséan Stage (Carboniferous): The Pengchong section, Guangxi, Sputh China. Episodes 2003, 26, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, C.O.; Schmitz, M.D.; Over, D.J.; Trayler, R.B.; Davydov, V.I. Recalibrating the Devonian time scale: A new method for integrating radioisotopic and astrochronologic ages in a Bayesian framework. Geol. Soc. Am. Bull. 2021, 134, 1931–1948. [Google Scholar] [CrossRef]
- Klapper, G.; Feist, R.; Becker, R.T.; House, M.R. Definition of the Frasnian/Famennian Stage boundary. Episodes 1993, 16, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Paproth, E.; Feist, R.; Flajs, G. Decision on the Devonian-Carboniferous boundary stratotype. Episodes 1991, 14, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Streel, M.; Brice, D.; Degardin, J.-M.; Derycke, C.; Dressen, R.; Groessens, E.; Hance, L.; Legarnd-Blain, M.; Lethiers, F.; Loboziak, S.; et al. Proposal for a Struntian Substage and a subdivision of the Famennian Stage into four Substages. Subcomission Devonian Stratigr. Newsl. 1998, 15, 47–49. [Google Scholar]
- Ruban, D.A.; Radulović, B.V.; Radulović, V.J. Diversity dynamics of Early Cretaceous brachiopods in the tectonic units of Serbia: Regional versus global patterns. Proc. Geol. Assoc. 2016, 127, 691–698. [Google Scholar] [CrossRef]
- Jaccard, P. Etude comparative de la distribution florale dans une portion des Alpes et du Jura. Bull. Société Vaud. Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- Sepkoski, J.J., Jr. Quantified Coefficients of Association of Measurement of Similarity. Math. Geol. 1974, 6, 135–152. [Google Scholar] [CrossRef]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–871. [Google Scholar] [CrossRef]
- Horowitz, A.S.; Pachut, J.F. The fossil record of bryozoan species diversity. In Proceedings of the 11th International Bryozoology Association Conference; Smithsonian Tropical Research Institute: Calzada de Amador, Panama, 2000; pp. 245–248. [Google Scholar]
- Bond, D.; Wignall, P.B.; Racki, G. Extent and duration of marine anoxia during the Frasnian-Famennian (Late Devonian) mass extinction in Poland, Germany, Austria and France. Geol. Mag. 2004, 141, 173–193. [Google Scholar] [CrossRef] [Green Version]
- Buggisch, W. The global Frasnian-Famennian »Kellwasser Event«. Geol. Rundsch. 1991, 80, 49–72. [Google Scholar] [CrossRef]
- Cohen, P.A.; Junium, C.K.; King Phillips, E.; Uveges, B.T. Carbon cycle dynamics and ecology revealed by the carbon isotopic composition of single organic microfossils during the Late Devonian Biotic Crisis. Geobiology 2022, 20, 346–362. [Google Scholar] [CrossRef] [PubMed]
- Racki, G.; Wignall, P.B. Devonian global changes—Recent advances and challenges in different domains. Glob. Planet. Chang. 2020, 191, 103200. [Google Scholar] [CrossRef]
- Sandberg, C.A.; Morrow, J.R.; Ziegler, W. Late Devonian sea-level changes, catastrophic events, and mass extinctions. Spec. Pap. Geol. Soc. Am. 2002, 356, 473–487. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).