
The Sub-Ice Algal Communities of the Barents Sea Pack Ice: Temporal and Spatial Distribution of Biomass and Species

Abstract
1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Analyses
3. Results
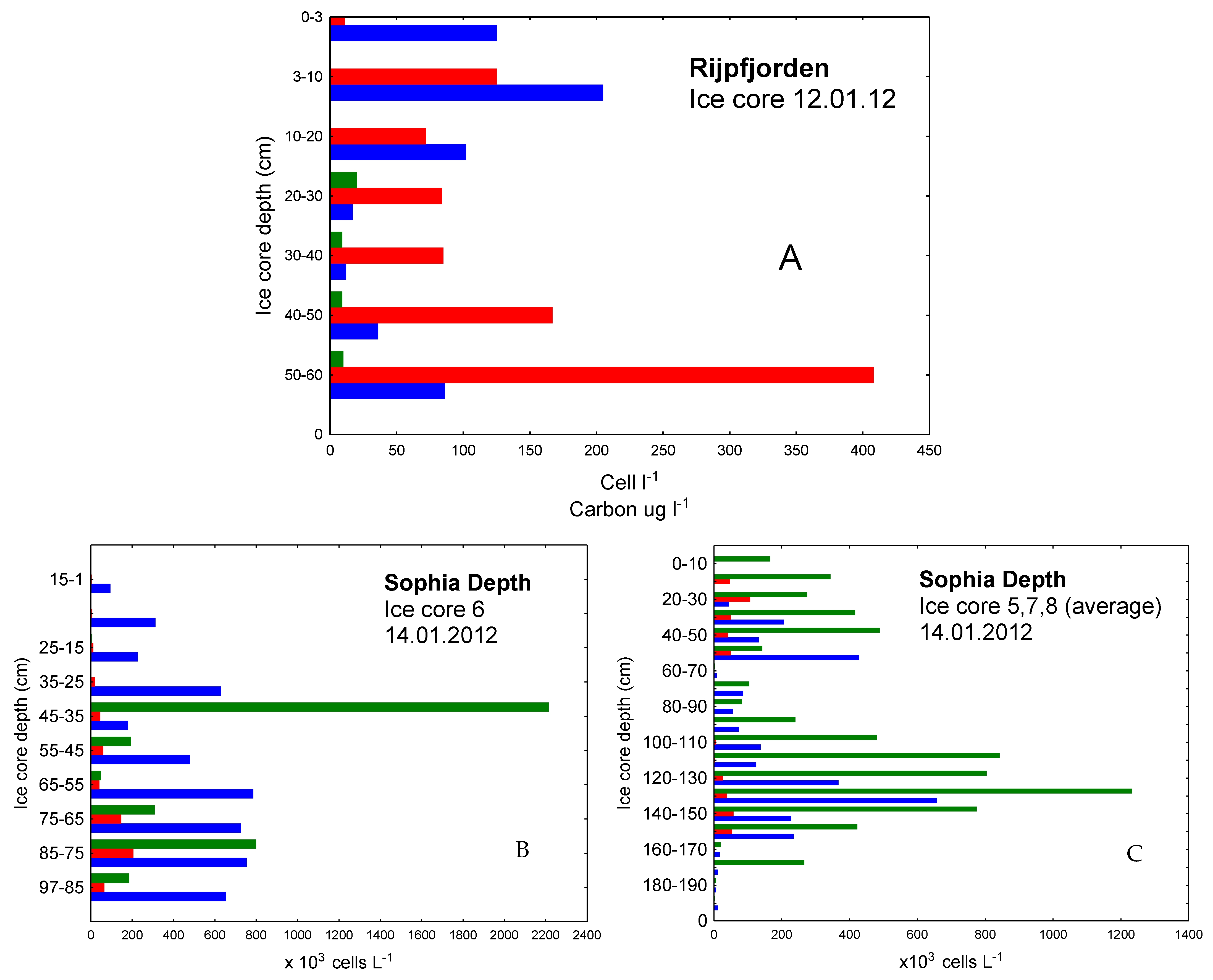
3.1. The Winter Season
3.2. The Spring Season
3.3. The Summer and Autumn Season
3.4. Statistics
4. Discussion
4.1. Biomass Distribution and Biodiversity in Relation to Environmental Factors
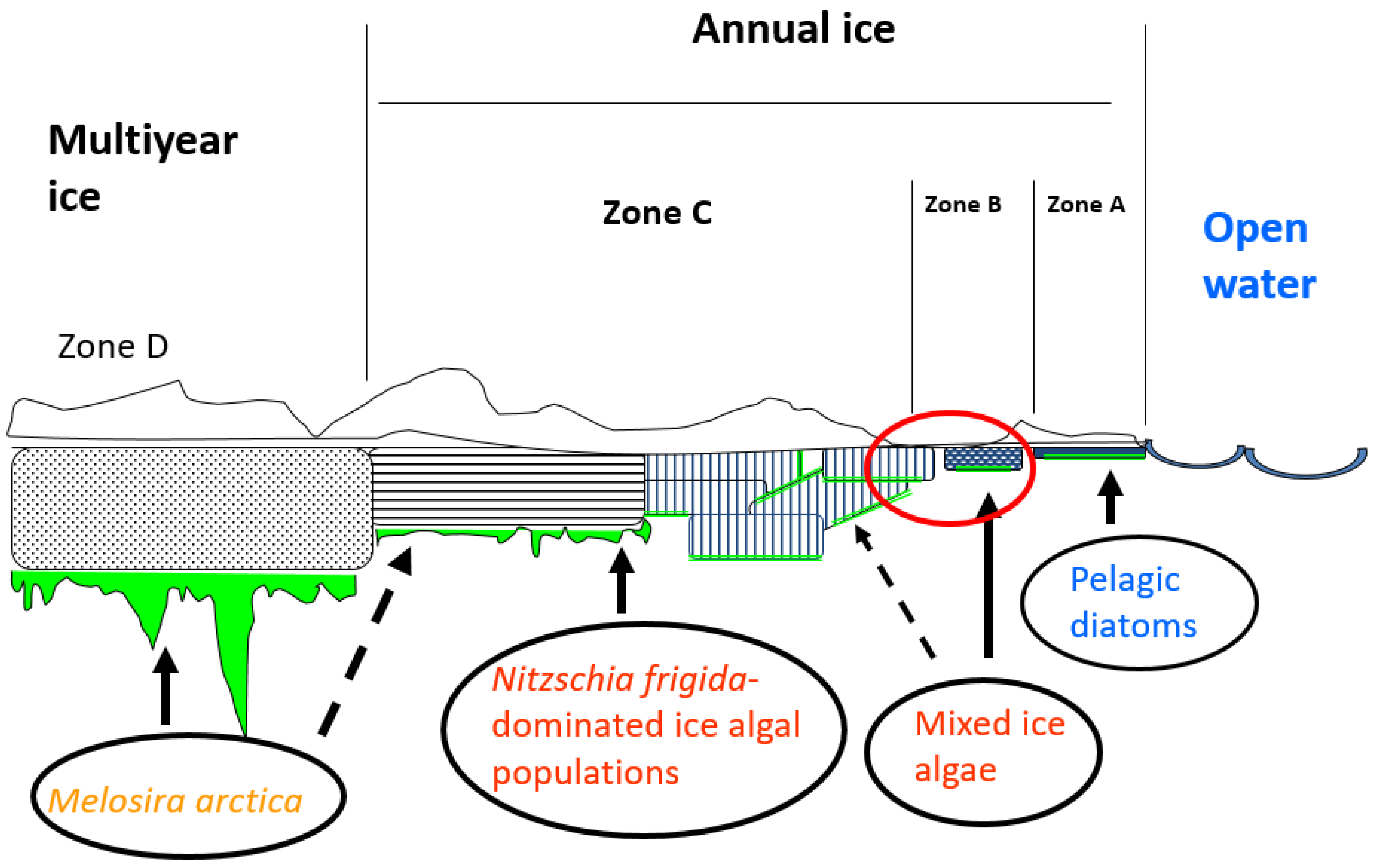
4.2. Vertical Distribution
4.3. Horisontal Distribution
4.4. Seasonal Occurrence
4.5. The Fate of the Ice Algal Blooms
4.6. Initiation of Ice Algal Blooms
4.7. The Future of Barents Sea Ice Algae
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poulin, M.; Daugbjerg, N.; Gradinger, R.; Ilyash, L.; Ratkova, T.; von Quillfeldt, C. The pan-Arctic biodiversity of marine pelagic and sea-ice unicellular eukaryotes: A first attempt assessment. Mar. Biodiv. 2011, 41, 13–28. [Google Scholar] [CrossRef]
- Bluhm, B.A.; Hop, H.; Melnikov, I.A.; Poulin, M.; Vihtakari, M.; Collins, E.; Gradinger, R.; Juul-Pedersen, T.; von Quillfeldt, C. State of the Arctic Marine Biodiversity Report; Sea-Ice Biota; Conservation of Arctic Flora and Fauna International Secretariat: Akureyri, Iceland, 2017; pp. 33–62. ISBN 978-9935-431-63-9. [Google Scholar]
- Leu, E.; Mundy, C.J.; Assmy, P.; Campbell, K.; Gabrielsen, T.; Gosselin, M.; Juul-Pedersen, T.; Gradinger, R. Arctic spring awakening—Steering principles behind phenology of vernal ice algal blooms. Progr. Ocean 2015, 139, 151–170. [Google Scholar] [CrossRef]
- Gradinger, R.; Friedrich, C.; Spindler, M. Abundance, biomass and composition of the sea ice biota of the Greenland Sea pack ice. Deep. Sea Res. Part II Top. Stud. Oceanogr. 1999, 46, 1457–1472. [Google Scholar] [CrossRef]
- Gradinger, R.; Spindler, M.; Weissenberger, J. On the structure and development of Arctic pack ice communities in Fram Strait: A multivariate approach. Polar Biol. 1992, 12, 727–733. [Google Scholar] [CrossRef]
- Gradinger, R.; Zhang, Q. Vertical distribution of bacteria in Arctic sea ice from the Barents and Laptev Seas. Polar Biol. 1997, 17, 448–454. [Google Scholar] [CrossRef]
- Gosselin, M.; Levasseur Wheeler, P.A.; Horner, R.A.; Booth, B. New measurements of phytoplankton and ice algae production in the Arctic Ocean. Deep-Sea Res. II 1997, 44, 1623–1644. [Google Scholar] [CrossRef]
- Gradinger, R. Vertical fine structure of the biomass and composition of algal communities in Arctic pack ice. Mar. Biol. 1999, 133, 745–754. [Google Scholar] [CrossRef]
- Ratkova, T.N.; Wassmann, P. Sea ice algae in the White and Barents seas: Composition and origin. Polar Res. 2005, 24, 95–110. [Google Scholar] [CrossRef]
- Druzhkov, N.V.; Druzchkova, E.I.; Kuznetzov, L.L. The sea-ice algal community of seasonal pack ice in the southwestern Kara Sea in late winter. Polar Biol. 2001, 24, 70–72. [Google Scholar] [CrossRef]
- Okolodkov, Y.B. Cryopelagic Flora of the Chukchi, East Siberian and Laptev Seas; National Institute of Polar Research Symposium of Polar Biology: Tokyo, Japan, 1992; pp. 28–43. [Google Scholar]
- Syvertsen, E.E. Ice algae in the Barents Sea: Types of assemblages, origin, fate and role in the ice-edge phytoplankton bloom. Polar Res. 1991, 10, 277–288. [Google Scholar] [CrossRef]
- Hegseth, E.N. Sub-ice algal assemblages of the Barents Sea: Species composition, chemical composition, and growth rates. Polar Biol. 1992, 12, 485–496. [Google Scholar] [CrossRef]
- Hegseth, E.N. Primary production in the northern Barents Sea. Polar Res. 1998, 17, 113–123. [Google Scholar] [CrossRef]
- McMinn, A.; Hegseth, E.N. Sea ice primary productivity in the northern Barets Sea, spring 2004. Polar Biol. 2007, 30, 289–294. [Google Scholar] [CrossRef]
- Fernández-Méndez, M.; Katlein, C.; Rabe, B.; Nicolaus, M.; Peeken, I.; Bakker, K.; Flores, H.; Boetius, A. Photosynthetic production in the central Arctic Ocean during the record sea-ice minimum in 2012. Biogeosciences 2015, 12, 3525–3549. [Google Scholar] [CrossRef]
- von Quillfeldt, C.H. (Ed.) Miljøverdier og Sårbarhet i Iskantsonen; Brief Report No. 047; Norwegian Polar Institute: Tromsø, Norway, 2018; 262p. (In Norwegian) [Google Scholar]
- Vinje, T.; Kvambekk, A.S. Barents Sea drift ice characteristics. Polar Res. 1991, 10, 59–68. [Google Scholar] [CrossRef]
- Rodrigues, J. The rapid decline of the sea ice in the Russian Arctic. Cold Reg. Sci. Technol. 2008, 54, 124–142. [Google Scholar] [CrossRef]
- Rodrigues, J. The increase in the length of the ice-free season in the Arctic. Cold Reg. Sci. Technol. 2009, 59, 78–101. [Google Scholar] [CrossRef]
- Smedsrud, L.H.; Esau, I.; Ingvaldsen, R.B.; Eldevik, T.; Haugan, P.M.; Li, C.; Lien, V.S.; Olsen, A.; Omar, A.M.; Otterå, O.H.; et al. The role of the Barents Sea in the Arctic climate system. Rev. Geophys. 2013, 51, 415–449. [Google Scholar] [CrossRef]
- Onarheim, I.H.; Eldevik, T.; Smedsrud, L.H.; Stroeve, J.C. Seasonal and Regional Manifestation of Arctic Sea Ice Loss. J. Clim. 2018, 31, 4917–4932. [Google Scholar] [CrossRef]
- Hansen, E.; Gerland, S.; Granskog, M.A.; Pavlova, O.; Renner, A.H.H.; Haapala, J.; Løyning, T.B.; Tschudi, M. Thinning of Arctic sea ice observed in Fram Strait: 1990–2011. J. Geophys. Res. Oceans 2013, 118, 5202–5221. [Google Scholar] [CrossRef]
- Hansen, E.; Ekeberg, O.; Gerland, S.; Pavlova, O.; Spreen, G.; Tschudi, M. Variability in categories of Arctic sea ice in Fram Strait. J. Geophys. Res. Oceans 2014, 119, 7175–7189. [Google Scholar] [CrossRef]
- Hansen, E.; Gerland, S.; Høyland, K.V.; Pavlova, O.; Spreen, G. Time variability in the annual cycle of sea ice thickness in the T ranspolar D rift. J. Geophys. Res. Oceans 2015, 120, 8135–8150. [Google Scholar] [CrossRef]
- Spreen, G.; de Steur, L.; Divine, D.; Gerland, S.; Hansen, E.; Kwok, R. Arctic sea ice volume export through Fram Strait from 1992 to 2014. J. Geophys. Res. Ocean. 2020, 125, e2019JC016039. [Google Scholar] [CrossRef]
- Lind, S.; Ingvaldsen, R.B.; Furevik, T. Arctic warming hotspot in the northern Barents Sea linked to declining sea-ice import. Nat. Clim. Chang. 2018, 8, 634–639. [Google Scholar] [CrossRef]
- Pavlova, O.; Pavlov, V.; Gerland, S. The impact of winds and sea surface temperatures on the Barents Sea ice extent, a statistical approach. J. Mar. Syst. 2014, 130, 248–255. [Google Scholar] [CrossRef]
- Onarheim, I.H.; Årthun, M. Toward an ice-free Barents Sea, Geophys. Res. Lett. 2017, 44, 8387–8395. [Google Scholar] [CrossRef]
- Stroeve, J.; Notz, D. Changing state of Arctic sea ice across all seasons. Environ. Res. Lett. 2018, 13, 103001. [Google Scholar] [CrossRef]
- Stiansen, J.E.; Filin, A.A. Joint PINRO/IMR Report on the State of the Barents Sea Ecosystem in 2007, with Expected Situation and Considerations for Management; IMR-PINRO Joint Report Series 1; IMR: Bergen, Norway, 2008; 185p. [Google Scholar]
- Gerland, S.; Goodwin, H. Region; Greenland and Barents Sea. In September 2009 Regional Sea Ice Outlook: July Report; ARCUS: Fairbanks, AK, USA, 2009. [Google Scholar]
- Gerland, S.; Renner, A.H.H.; Godtliebsen, F.; Divine, D.; Løyning, T.B. Decrease of sea ice thickness at Hopen, Barents Sea, during 1966–2007. Geophys. Res. Lett. 2008, 35, L06501. [Google Scholar] [CrossRef]
- Gerland, S.; Tronstad, S.; Pavlova, O.; Ingvaldsen, R. Isutbredelse i Barentshavet. In Forvaltningsplan Barentshavet—Rapport Fra Overvåkingsgruppen 2009; Sunnanå, K., Fosheim, M., van der Meeren, G.I., Eds.; Fisken Hav1b, Institute of Marine Research: Bergen, Norway; pp. 25–26. (In Norwegian)
- Kwok, R. Arctic sea ice thickness, volume, and multiyear ice coverage: Losses and coupled variability (1958–2018). Environ. Res. Lett. 2018, 13, 105005. [Google Scholar] [CrossRef]
- Wadhams, P. The seasonal ice zone. In The Geophysics of Sea Ice; Plenum Press: New York, NY, USA, 1986; pp. 825–991. [Google Scholar]
- Zhang, J.; Schweiger, A.; Steele, M.; Stern, H. Sea ice floe size distribution in the marginal ice zone. Theory and numerical experiments. J. Geophys. Res. 2015, 120, 3484–3498. [Google Scholar] [CrossRef]
- Vinje, T. Drift, Composition, Morphology and Distribution of the Sea Ice Fields in the Barents Sea; Norsk Polarinst Skrifter 179C; Norsk Polarinstitutt: Oslo, Norway, 1985; pp. 1–26. [Google Scholar]
- Lönne, O.J. A diver-operated electric suction sampler for sympagic (= under-ice) invertebrates. Polar Res. 1988, 6, 135–136. [Google Scholar] [CrossRef]
- Strickland, J.H.D.; Parsons, T.R. A practical handbook of sea water analysis. Bull. Fish. Res. Booard. Can. 1972, 167, 1–311. [Google Scholar]
- Grasshoff, K. Methods of Sea Water Analyses; Chemie: Weinheim, NY, USA, 1976; 317p. [Google Scholar]
- Holm-Hansen, O.; Riemann, B. Chlorophyll a determination: Improvements in methodology. Oikos 1978, 30, 438–447. [Google Scholar] [CrossRef]
- Simonsen, R. The diatom plankton of the Indian Ocean expedition of RV “Meteor” 1964–1965. Meteor Forchiungsergeb. 1974, 19, 1–66. [Google Scholar]
- Cleve, P.T. Diatoms, collected during the expedition of the Vega. In Vegaexpeditionens Vetenskapliga Iakttagelser 3. F. and G. Beijers Forlag; Sagwan Press: Stockholm, Sweden, 1883; pp. 455–517. [Google Scholar]
- Cleve, P.T. Plankton Collected by the Swedish Expedition to Spitsbergen in 1898. K. Sven. Vetensk. Akad. Handl. 1899, 32, 1–51. [Google Scholar]
- Østrup, E. Marine Diatoméer fra Østgrønland. Meddel. Grønl. 1895, 18, 395–476. [Google Scholar]
- Gran, H.H. Diatomaceae from ice floes and plankton of the Arctic Ocean. In The Norwegian North Polar Expedition 1893–1896. Scientific Results; Nansen, F., Ed.; Fridtjof Nansen Fund for the Advancement of Science, Longman, Green and Cp.: London, UK, 1904; Volume 4, pp. 3–74. [Google Scholar]
- Lebour, M.V. The Planktonic Diatoms of Northern Seas; Koenigstein: Koeltz, Germany, 1930; 244p. [Google Scholar]
- Poulin, M.; Cardinal, A. Sea ice diatoms from Manitounuk Sound, south eastern Hudson Bay (Quebec, Canada). I. Family Naviculaceae. Can. J. Bot. 1982, 60, 1263–1277. [Google Scholar] [CrossRef]
- Poulin, M.; Cardinal, A. Sea ice diatoms from Manitounuk Sound, south eastern Hudson Bay (Quebec, Canada). II. Naviculaceae, genus Navicula. Can. J. Bot. 1982, 60, 1263–1277. [Google Scholar] [CrossRef]
- Poulin, M.; Cardinal, A. Sea ice diatoms from Manitounuk Sound, south eastern Hudson Bay (Quebec, Canada). III. Cymbellaceae, Entomoneidaceae, Gomphonematacea and Nitzschiaceae. Can. J. Bot. 1983, 61, 107–118. [Google Scholar] [CrossRef]
- Medlin, L.K.; Hasle, G.R. Some Nitzschia and related diatom species from fast-ice samples in the Arctic and Antarctic. Polar Biol. 1990, 10, 451–479. [Google Scholar] [CrossRef]
- Medlin, L.K.; Priddle, J. Polar Marine Diatoms; British Antarctic Survey: Cambridge, UK, 1990; 214p. [Google Scholar]
- Medlin, L.K.; Round, F.E. Taxonomic studies of marine gomphonemoid diatoms. Diatom Res. 1986, 1, 205–225. [Google Scholar] [CrossRef]
- Makarova, I.V. Diatoms of the Seas of the USSR: The Genus Thalassiosira Cl; Nauka: Leningrad, Russia, 1988; 117p. (In Russian) [Google Scholar]
- Hasle, G.R.; Medlin, L.K.; Syvertsen, E.E. Synedropsis gen. nov., a new genus of araphid diatoms associated with sea ice. Phycologia 1994, 33, 248–270. [Google Scholar] [CrossRef]
- Hasle, G.R.; Syvertsen, E.E.; von Quillfeldt, C.H. Fossula arctica gen. nov. spec. nov., a marine arctic araphid diatom. Diatom. Res. 1996, 11, 261–272. [Google Scholar] [CrossRef]
- Hasle, G.R.; Syvertsen, E.E. Marine diatoms. In Identifying Marine Diatoms and Dinoflagellates; Thomas, C.R., Ed.; Academic Press Incorporation: San Diego, CA, USA, 1997; pp. 5–385. [Google Scholar]
- Steidinger, K.A.; Tangen, K. Dinoflagellates. In Identifying Marine Phytoplankton; Tomas, Ed.; Acad. Press Inc.: San Diego, CA, USA, 1997; pp. 387–584. [Google Scholar]
- Montressor, M.; Lovejoy, C.; Orsini, L.; Procaccani, G.; Roy, S. Bipolar distribution of the cyst-forming dinoflagellates Polarella glacialis. Polar Biol. 2003, 26, 186–194. [Google Scholar] [CrossRef]
- McMinn, A.; Scott, F.J. Dinoflagellates. In Antarctic Marine Protists; Scott, F.J., Marchant, H.J., Eds.; ABRS, Canberrs and AAD: Hobart, Australia, 2005; pp. 202–250. [Google Scholar]
- Dytham, C. Choosing and Using Statistics; Blackwell Science: Hoboken, NJ, USA, 2001; 218p. [Google Scholar]
- Welch, H.E.; Bergmann, M.A. Seasonal development of ice algae and its prediction from environmental factors near Resolute, N.W.T., Canada. Can. J. Fish. Aquat. Sci. 1989, 46, 1793–1804. [Google Scholar] [CrossRef]
- Horner, R.A.; Schrader, G.C. Relative contribution of ice algae, phytoplankton, and benthic microalgae to primary production in nearshore regions of the Beaufort Sea. Arctic 1982, 35, 485–503. [Google Scholar] [CrossRef]
- Hsiao, S.I.C. Quantitative composition, distribution, community structure and standing stock of sea ice microalgae in the Canadian Arctic. Arctic 1980, 33, 768–793. [Google Scholar] [CrossRef]
- Hsiao, S.I.C. Dynamics of ice algae and phytoplankton in Frobisher Bay. Polar Biol. 1992, 12, 645–651. [Google Scholar] [CrossRef]
- Gradinger, R.; Spindler, M.; Henschel, D. Development of Arctic sea-ice organisms under graded snow cover. Polar Res. 1991, 10, 295–307. [Google Scholar] [CrossRef]
- McRoy, C.P.; Goering, J.J. The influence of ice on the primary productivity of the Bering Sea. In Oceanography of the Bering Sea; Hood, D.W., Kelley, E.J., Eds.; Institute of Maine Science, Occasional Publication, No. 2.; University of Alaska: Fairbanks, AK, USA, 1974; pp. 403–421. [Google Scholar]
- Booth, J.A. The epontic algal community of the ice edge zone and its significance to the Davis Strait ecosystem. Arctic 1984, 37, 234–243. [Google Scholar] [CrossRef]
- Tomczak, M.; Godfrey, J.S. Regional Oceanography: An Introduction; Pergamon: Oxford, UK, 1994; 422p. [Google Scholar]
- Codispoti, L.A.; Owens, T.G. Nutrient transport through Lancaster Sound in relation to the Arctic Ocean’s reactive silicate budget and the outflow of Bering Strait waters. Limnol. Oceanogr. 1975, 20, 115–119. [Google Scholar] [CrossRef]
- Ambrose, W.G.; Quillfeldt CH von Clough, L.M.; Tilney, P.R.; Tucker, W. The Sub-ice algal community in the Chukchi Sea: Large and small scale patters of abundance based on images from a remotely operated vehicle. Polar Biol. 2005, 28, 784–795. [Google Scholar] [CrossRef]
- Tremblay, C.; Runge, J.A.; Legendre, L. Grazing and sedimentation of ice algae during and immediately after a bloom at the ice-water interface. Mar. Ecol. Prog. Ser. 1989, 56, 291–300. [Google Scholar] [CrossRef]
- Assmy, P.; Fernandez-Mendez, M.; Duarte, P.; Meyer, A.; Randelhoff, A.; Mundy, C.J.; Olsen, L.M.; Kauko, H.M.; Bailey, A.; Chierici, M.; et al. Leads in Arctic pack ice enable early phytoplankton blooms below snow-covered sea ice. Sci. Rep. 2017, 7, 40850. [Google Scholar] [CrossRef]
- Arrigo, K.B.; Perovich, D.K.; Pickart, R.S.; Brown, Z.W.; van Dijken, G.L.; Lowry, K.E.; Mills, M.M.; Palmer, M.A.; Balch, W.M.; Bahr, F.; et al. Massive phytoplankton blooms under Arctic sea ice. Science 2012, 336, 1408. [Google Scholar] [CrossRef]
- Ardyna, M.; Mundy, C.J.; Mayot, N.; Matthes, L.C.; Oziel, L.; Horvat, C.; Leu, E.; Assmy, P.; Hill, V.; Matrai, P.A.; et al. Under-Ice Phytoplankton Blooms: Shedding Light on the “Invisible” Part of Arctic Primary Production. Front. Mar. Sci. 2020, 7, 608032. [Google Scholar] [CrossRef]
- Cross, W.E. Under-ice biota at the Pond Inlet ice edge and in adjacent fast ice areas during spring. Arctic 1982, 35, 13–27. [Google Scholar] [CrossRef]
- Johnsen, G.; Hegseth, E.N. Photoadaptation of sea-ice microalgae in the Barents Sea. Polar Biol. 1991, 11, 179–184. [Google Scholar] [CrossRef]
- Van Leeuwe, M.A.; Tedesco, L.; Arrigo, K.R.; Assmy, P.; Campbell, K.; Meiners, K.M.; Rintala, J.-M.; Selz, V.; Thomas, D.N.; Stefels, J.; et al. Micro-algal community structure and primary production in Arctic and Antarctic sea ice: A synthesis. Elem. Sci. Anth. 2018, 6, 4. [Google Scholar] [CrossRef]
- Okolodkov, Y.B. Algae in the annual sea ice at Hooker Island, Franz Josef Land, in August 1991. Polish. Pol. Res. 1993, 14, 25–32. [Google Scholar]
- von Quillfeldt, C.H. Distribution of diatoms in the Northeast Water Polynya, Greenland. J. Mar. Syst. 1997, 10, 211–240. [Google Scholar] [CrossRef]
- von Quillfeldt, C.H.; Ambrose, W.G.; Clough, L.M. High number of diatom species in first year ice from the Chukchi Sea. Polar Biol. 2003, 26, 806–818. [Google Scholar] [CrossRef]
- Hancke, K.; Lund-Hansen, L.C.; Lamare, M.L.; Pedersen, S.H.; King, M.D.; Andersen, P.; Sorrell, B.H. Extreme low light requirement for algae growth underneath sea ice: A case study from Station Nord, NE Greenland. J. Geophys. Res. 2018, 123, 985–1000. [Google Scholar] [CrossRef]
- von Quillfeldt, C.H. Common diatom species in arctic spring blooms: Their distribution and abundance. Mar. Bot. 2000, 43, 499–516. [Google Scholar] [CrossRef]
- Hegseth, E.N.; von Quillfeldt, C.H. Low phytoplankton biomass and ice algal blooms in the Weddell Sea during the ice-filled summer of 1997. Ant. Sci. 2002, 14, 231–243. [Google Scholar] [CrossRef]
- McMinn, A.; Hegseth, E.N. Quantum yield and photosynthetic parameters of marine microalgae from the southern Arctic Ocean, Svalbard. J. Mar. Biol. Ass. UK 2004, 84, 865–871. [Google Scholar] [CrossRef]
- Fernández-Méndez, M.; Olsen, L.M.; Kauko, H.M.; Meyer, A.; Rösel, A.; Merkouriadi, I.; Mundy, C.J.; Ehn, J.K.; Johansson, A.M.; Wafner, P.M.; et al. Algal hot spots in a changing Arctic Ocean: Sea-ice ridges and the snow-ice interface. Front. Mar. Sci. 2018, 5, 75. [Google Scholar] [CrossRef]
- Cota, G.F.; Smith, R.E.H. Ecology of bottom ice algae: III. Comparative physiology. J. Mar. Syst. 1991, 2, 297–315. [Google Scholar] [CrossRef]
- Horner, R.; Ackley, S.F.; Dickmann, G.S.; Gulliksen, B.; Hoshiai, T.; Legendre, L.; Melnikov, I.A.; Reeburgh, W.S.; Spindler, M.; Sullivan, C.S. Ecology of sea ice biota. I. Habitat, terminology and methodology. Polar Biol. 1992, 12, 417–427. [Google Scholar] [CrossRef]
- Buck, K.R.; Bolt, P.A.; Bentham, W.N.; Garrison, D.L. A dinoflagellate cyst from Antarctic sea ice. J. Phycol. 1992, 28, 15–18. [Google Scholar] [CrossRef]
- Stoecker, D.K.; Buck, K.R.; Putt, M. Changes in the sea-ice brine community during the spring-summer transition, McMurdo Sound, Antarctica. I. Photosynthetic protists. Mar. Ecol. Prog. Ser. 1992, 84, 265–278. [Google Scholar] [CrossRef]
- Montressor, M.; Procaccini, G.; Stoecker, D.K. Polarella glacialis gen. nov. sp. nov. (Dinophyceae): Suessiaceae are still alive! J. Phycol. 1999, 35, 186–197. [Google Scholar] [CrossRef]
- Thomson, P.G.; McMinn, A.; Kiessling, I.; Watson, M.; Goldsworthy, P.M. Composition and succession of dinoflagellates and chrysophytes in the upper fast ice of Davis Station, East Antarctica. Polar Biol. 2006, 29, 337–345. [Google Scholar] [CrossRef]
- Meunier, A. Microplankton des Mers de Barents et de Kara. Duc d’Orleans, Campagne Arctique de 1907; Imprimerie Scientifique Charles Bulens: Bruxelles, Belgium, 1910; 355p. [Google Scholar]
- Ikävalko, J.; Gradinger, R. Flagellates and heliozoans in the Greenland Sea ice studies alive using light microscopy. Polar Biol. 1997, 17, 473–481. [Google Scholar]
- Rysgaard, S.; Kühl, M.; Glud, R.N.; Hansen, J.W. Biomass, production and horizontal patchiness of sea ice algae in a high-Arctic fjord (Young Sound, NE Greenland). Mar. Ecol. Prog. Ser. 2001, 223, 15–26. [Google Scholar] [CrossRef]
- Granskog, M.A.; Kaartokallio, H.; Kuosa, H.; Thomas, D.N.; Ehn, J.; Sonninen, E. Scales of horizontal patchiness in chlorophyll a, chemical and physical properties of landfast sea ice in the Gulf of Finland (Baltic Sea). Polar Biol. 2004, 28, 276–283. [Google Scholar] [CrossRef]
- Werner, I.; Lindemann, F. Video observations of the underside of arctic sea ice-features and morphology on medium and small scales. Polar Res. 1997, 16, 27–36. [Google Scholar] [CrossRef]
- Mundy, C.J.; Barber, D.G.; Michel, C. Variability of snow and ice thermal, physical and optical properties pertinent to sea ice algae biomass during spring. J. Mar. Sys. 2005, 58, 107–120. [Google Scholar] [CrossRef]
- Lund-Hansen, L.C.; Hawes, I.; Sorrell, B.K.; Nielsen, M.H. Removal of snow cover inhibits spring growth og Arctic ice algae through physiological and behavioural effects. Polar Biol. 2013, 156, 234–244. [Google Scholar]
- von Quillfeldt, C.H. Ice Algae and Phytoplankton in North Norwegian and Arctic Waters: Species Composition, Succession and Distribution. Ph.D. Thesis, University of Tromsö, Tromsö, Norway, 1996. [Google Scholar]
- von Quillfeldt, C.H. The diatom Fragilariopsis cylindrus and its potential as an indicator species for cold water rather than for sea ice. Vie. Milieu. 2004, 54, 137–143. [Google Scholar]
- Maykut, G.A.; Grenfell, T.C. The spectral distribution of light beneath first-year sea ice in the Arctic Ocean. Limnol. Oceanogr. 1975, 20, 554–563. [Google Scholar] [CrossRef]
- Usachev, P.I. Microflora of the polar ice. Tr. Inst. Okeanol. AN SSSR (Proc. Inst. Oceanol. USSR Acad. Sci.) 1949, 3, 216–259. (In Russian) [Google Scholar]
- Melnikov, I.A. The Arctic Sea Ice Ecosystem; Gordon and Breach Science Publ.: London, UK, 1997. [Google Scholar]
- Melnikov, I.A.; Bondarchuk, L.L. Ecology of the mass accumulations of colonial diatom algae under drifting Arctic ice. Oceanology 1987, 27, 233–236. [Google Scholar]
- Hellum, C. Phytoplankton and ice algae. Species composition. Ber. Polarforsch. 1994, 142, 67–72. [Google Scholar]
- Boetius, A.; Albrecht, S.; Bakker, K.; Bienhold, C.; Felden, J.; Fernández-Méndez, M.; Hendricks, S.; Katlein, C.; Lalande, C.; Krumpen, T.; et al. Export of algal biomass from the melting Arctic sea ice. Science 2013, 339, 1430–1432. [Google Scholar] [CrossRef] [PubMed]
- Hop, H.; Vihtakari, M.; Bluhm, B.A.; Assmy, P.; Poulin, M.; Gradinger, R.; Peeken, I.; von Quillfeldt, C.H.; Olsen, L.M.; Zhitina, L.; et al. Changes in sea-ice protist diversity with declining sea ice in the Arctic Ocean from the 1980s to 2010s. Front. Mar. Sci. 2020, 7, 243. [Google Scholar] [CrossRef]
- McConville, M.J. Chemical composition and biochemistry of sea ice microalgae. In Sea Ice Biota; Horner, R.A., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1985; pp. 105–129. [Google Scholar]
- Krembs, C.; Engel, A. Abundance and variability of microorganisms and transparent exopolymer particles across the ice-water interface of melting first-year sea ice in the Laptev Sea (Arctic). Mar. Biol. 2001, 138, 173–185. [Google Scholar] [CrossRef]
- Krembs, C.; Eicken, H.; Junge, K.; Deming, J.W. High concentrations of exopolymer substances in Arctic winter ice: Implications for the polar ocean carbon cycle and cryoprotection of diatoms. Deep-Sea Res. I 2002, 49, 2163–2181. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Hop, H.; Budgell, P.; Hegseth, E.N.; Korsnes, R.; Loyning, T.; Oerbaek, J.B.; Kawamura, T.; Shirasawa, K. Physical and ecological processes in the marginal ice zone of the northern Barents Sea during summer melt period. J. Mar. Syst. 2000, 27, 131–159. [Google Scholar] [CrossRef]
- Hegseth, E.N.; Sundfjord, A. Intrusion and blooming of Atlantic phytoplankton species in the high Arctic. J. Mar. Syst. 2008, 74, 108–119. [Google Scholar] [CrossRef]
- Assmy, P.; Ehn, J.K.; Fernandez-Mendez, M.; Hop, H.; Sundfjord, A.; Bluhm, K.; Daase, M.; Engel, A.; Granskog, M.A.; Hudson, S.R.; et al. Floating ice-algal aggregates below melting Arctic sea ice. PLoS ONE 2013, 8, e76599. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; McRoy, C.P.; Joo, H.M.; Gradinger, R.; Cui, H.; Yun, M.S.; Chung, K.H.; Kang, S.-H.; Kang, C.-K.; Choy, E.J.; et al. Holes in progressively thinning Arctic sea ice lead to new ice algae habitat. Oceanography 2011, 24, 302–308. [Google Scholar] [CrossRef]
- Werner, I. Grazing of Arctic under-ice amphipods on sea-ice algae. Mar. Ecol. Prog. Ser. 1997, 160, 93–99. [Google Scholar] [CrossRef][Green Version]
- Søreide, J.E.; Hop, H.; Falk-Petersen, S.; Hegseth, E.N.; Carroll, M.L. Seasonal food web structures and sympagic-pelagic coupling in the European Arctic revealed by stable isotopes and a two-source food web model. Prog. Oceanogr. 2006, 71, 59–87. [Google Scholar] [CrossRef]
- Scott, C.L.; Falk-Petersen, S.; Gulliksen, B.; Lønne, O.J.; Sargent, J.R. Lipid indicators of the diet of the sympagic amphipod Gammarus wilkitzkii in the Marginal Ice Zone and in open waters of Svalbard (Arctic). Polar Biol. 2001, 24, 572–576. [Google Scholar] [CrossRef]
- Kohlbach, D.; Graeve, M.; Lange, B.A.; David, C.; Peeken, I.; Flores, H. The importance of ice algae-produced carbon in the central Arctic Ocean ecosystem: Food web relationships revealed by lipid and stable isotope analyses. Limnol. Oceanogr. 2016, 61, 2027–2044. [Google Scholar] [CrossRef]
- Brown, T.A.; Assmy, P.; Hop, H.; Wold, A.; Belt, S.T. Transfer of ice algae carbon to ice-associated amphipods in the high-Arctic pack ice environment. J. Plankton Res. 2017, 39, 664–674. [Google Scholar] [CrossRef]
- Nozais, C.; Gosselin, M.; Michel, C.; Tita, G. Abundance, biomass, composition and grazing impact of the sea-ice meiofauna in the North Water, northern Baffin Bay. Mar. Ecol. Prog. Ser. 2001, 217, 235–250. [Google Scholar] [CrossRef]
- Werner, I.; Arbizu, P.M. The sub-ice fauna of the Laptev Sea and the adjacent Arctic Ocean in summer 1995. Polar Biol. 1999, 21, 71–79. [Google Scholar] [CrossRef]
- Søreide, J.E.; Falk-Petersen, S.; Hegseth, E.N.; Hop, H.; Carroll, M.L.; Hobson, K.A.; Blachowiak-Samolyk, K. Seasonal feeding strategies of Calanus in the high-Arctic Svalbard region. Deep-Sea Res. II 2008, 55, 2225–2244. [Google Scholar] [CrossRef]
- Runge, J.A.; Ingram, R.G. Underice grazing by planktonic calanoid copepods in relation to a bloom of ice microalgae in Southeastern Hudson Bay. Limnol. Oceanogr. 1988, 33, 280–286. [Google Scholar] [CrossRef]
- Runge, J.A.; Ingram, R.G. Under-ice feeding and diel migration by the planktonic copepods Calanus glacialis and Pseudocalanus minutes in relation to the ice algal production cycle in southeastern Hudson Bay, Canada. Mar. Biol. 1991, 8, 217–225. [Google Scholar] [CrossRef]
- Werner, I.; Hirche, H.-J. Observations on Calanus glacialis eggs under the spring sea-ice in the Barents Se. Polar Biol. 2001, 24, 296–298. [Google Scholar] [CrossRef]
- Clasby, R.C.; Alexander, V.; Horner, R. Primary productivity of sea-ice algae. In Assessment of the Arctic Marine Environment: Selected Topics; Hood, D.W., Burrell, D.C., Eds.; Institute of Marine Science: Fairbanks, AK, USA, 1976; pp. 289–304. [Google Scholar]
- Bradstreet, M.S.W.; Cross, W.E. Trophic relationships at high Arctic ice edges. Arctic 1982, 35, 1–12. [Google Scholar] [CrossRef]
- Kern, J.C.; Carey, O.G.J. The faunal assemblage inhabiting seasonal sea-ice in the nearshore Arctic Ocean with emphasis on copepods. Mar. Ecol. Prog. Ser. 1983, 10, 159–167. [Google Scholar] [CrossRef]
- Gulliksen, B.; Lønne, O.J. Distribution, abundance, and ecological importance of marine sympagic fauna in the Arctic. Rapp. P-v. Reun. Cons. Int. Explor. Mer. 1991, 188, 133–138. [Google Scholar]
- Poltermann, M. Biology and ecology of cryopelagic amphipods from Arctic sea ice. Ber. Polarforsch. 1997, 225, 1–170. [Google Scholar]
- Poltermann, M. Arctic sea ice as feeding ground for amphipods—food resources and strategies. Polar Biol. 2001, 24, 89–96. [Google Scholar] [CrossRef]
- Weissenberger, J. Observations of sea ice organisms in the field using an endoscope. Ber. Polarforsch. 1994, 149, 64–68. [Google Scholar]
- Macdonald, R.W.; Solomon, S.M.; Cranston, R.E.; Welch, H.E.; Yunker, M.B.; Gobeil, C. A sediment and organic carbon budget for the Canadian Beaufort Shelf. Mar. Geol. 1998, 144, 255–273. [Google Scholar] [CrossRef]
- Tamelander, T.; Reigstad, M.; Hop, H.; Ratkova, T. Ice algal assemblages and vertical export of organic matter from sea ice in the Barents Sea and Nansen Basin (Arctic Ocean). Polar Biol. 2009, 32, 1261–1273. [Google Scholar] [CrossRef]
- Juul-Pedersen, T.; Michel, C.; Gosselin, M.; Seuthe, L. Seasonal changes in the sinking export of particulate material under first-year sea ice on the Mackenzie Shelf (Western Canadian Arctic). Mar. Ecol. Progr. Ser. 2008, 353, 13–25. [Google Scholar] [CrossRef]
- Fortier, M.; Fortier, L.; Michel, C.; Legendre, L. Climatic and biological forcing of the vertical flux of biogenic particles under seasonal Arctic sea ice. Mar. Ecol. Prog. Ser. 2002, 225, 1–16. [Google Scholar] [CrossRef]
- Lovvorn, J.R.; Cooper, L.W.; Brooks, M.L.; De Reyck, C.C.; Bump, J.K.; Grebmeier, J.M. Organic matter pathways to zooplankton and benthos under pack ice in late winter and open water in late summer in the north-central Bering Sea. Mar. Ecol. Prog. Ser. 2005, 291, 135–150. [Google Scholar] [CrossRef]
- Michel, C.; Ingram, R.G.; Harris, L.R. Variability in oceanographic and ecological processes in the Canadian Arctic Archipelago. Progr. Oceanogr. 2006, 71, 379–401. [Google Scholar] [CrossRef]
- Carroll, M.L.; Carroll, J. The Arctic Seas. In Biogeochemistry of Marine Systems; Black, K., Shimmield, G., Eds.; Blackwell Publ. Ltd.: Oxford, UK, 2003; pp. 127–156. [Google Scholar]
- McMahon, K.; Ambrose, W.G.; Johnson, B.J.; Sun, M.-Y.; Lopez, G.R.; Clough, L.M.; Carroll, M.L. Benthic community response to the ice algae and phytoplankton in Ny-Ålesund, Svalbard. Mar. Ecol. Prog. Ser. 2006, 110, 14. [Google Scholar] [CrossRef]
- Sun, M.-Y.; Carroll, M.L.; Ambrose, W.G.; Clough, L.M.; Zou, L.; Lopez, G.R. Rapid consumption of phytoplankton and ice algae by Arctic soft-sediment benthic communities: Evidence using natural and 13C-labeled food materials. J. Mar. Syst. 2007, 65, 561–588. [Google Scholar] [CrossRef]
- Moran, S.B.; Lomas, M.W.; Kelly, R.P.; Gradinger, R.; Iken, K.; Mathis, J.T. Seasonal succession of net primary productivity, particulate organic carbon export, and autotrophic community composition in the eastern Bering Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2012, 65–70, 84–97. [Google Scholar] [CrossRef]
- Rybakova, E.; Kremenetskaia, A.; Vedenin, A.; Boetius, A.; Gebruk, A. Deep-sea megabenthos communities of the Eurasian Central Arctic are influenced by ice-cover and sea-ice algal falls. PLoS ONE 2019, 14, 0211009. [Google Scholar] [CrossRef]
- Renaud, P.E.; Riedel, A.; Michel, C.; Morata, N.; Gosselin, M.; Juul-Pedersen, T.; Chiuchiolo, A. Seasonal variation in benthic community oxygen demand: A response to an ice algal bloom in the Beaufort Sea, Canadian Arctic? J. Mar. Syst. 2007, 67, 1–12. [Google Scholar] [CrossRef]
- Anisimova, N.A.; Jørgensen, L.L.; Lyubin, P.A.; Manushin, I.E. Mapping and Monitoring of Benthos in the Barents Sea and Svalbard Waters: Results from the Joint Russian—Norwegian Benthic Programme 2006–2008; IMR-PINRO Joint Report Series 1–2010; Institute of Marine Research: Bergen, Norway, 2010; 114p, ISSN 1502-8828. [Google Scholar]
- Søreide, J.E.; Carroll, M.L.; Hop, H.; Ambrose Jr, W.G.; Hegseth, E.N.; Falk-Petersen, S. Sympagic-pelagic-benthic coupling in Arctic and Atlantic waters around Svalbard revealed by stable isotopic and fatty acid tracers. Mar. Biol. Res. 2013, 9, 831–850. [Google Scholar] [CrossRef]
- Brown, T.A.; Hegseth, E.N.; Belt, S.T. Biomarker signatures of ice algae and phytoplankton in marine sediments provide evidence for a contributory food source for the mid-winter ecosystem in Rijpfjorden, Svalbard. Polar Biol. 2013, 38, 37–50. [Google Scholar] [CrossRef]
- Brown, T.; Belt, S. Identification of the sea ice diatom biomarker IP25 in Arctic benthic macrofauna: Direct evidence for a sea ice diatom diet in Arctic heterotrophs. Polar Biol. 2012, 35, 131–137. [Google Scholar] [CrossRef]
- Fryxell, G.A. Planktonic marine diatom winter stages: Antarctic alternatives to resting spores. Proc. II Int Diatom Symp. Calif. Acad.Sci. 1990, 17, 437–448. [Google Scholar]
- Horner, R.A. Taxonimy of sea ice microalgae. In Sea Ice Biota; Horner, R.A., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1985; pp. 147–157. [Google Scholar]
- Hegseth, E.N.; Svendsen, H.; von Quillfeldt, C.H. Phytoplankton in fjords and coastal waters of northern Norway: Environmental conditions and dynamics of the spring bloom. In Ecology of Fjords and Coastal Waters; Skjoldal, H.R., Hopkins, C., Erikstad, K.E., Leinaas, H.P., Eds.; Elsevier Science BV: Amsterdam, The Netherlands, 1995; pp. 45–72. [Google Scholar]
- Wiktor, J. Early spring microplankton development under fast ice covered fjords of Svalbard, Arctic. Oceanologia 1999, 41, 51–72. [Google Scholar]
- Eilertsen, H.C.; Sandberg, S.; Töllefsen, H. Photoperiodic control of diatom spore growth: A theory to explain the onset of phytoplankton blooms. Mar. Ecol. Prog. Ser. 1995, 116, 303–307. [Google Scholar] [CrossRef]
- Backhaus, J.; Wehde, H.; Hegseth, E.; Kämpf, J. Phyto-convection:the role of oceanic convection in primary production. Mar. Ecol. Prog. Ser. 1999, 189, 77–92. [Google Scholar] [CrossRef]
- Backhaus, J.O.; Fohrmann, H.; Kämpf, J.; Rubino, A. Formation and export of water masses produced in Arctic shelf polynyas—process studies of oceanic convection. ICES J. Mar. Sci. 1997, 54, 366–382. [Google Scholar] [CrossRef]
- Kvernvik, A.C.; Rokitta, S.D.; Leu, E.; Harms, L.; Gabrielsen, T.M.; Rost, B.; Hoppe, C.J.M. Higher sensitivity towards light stress and ocean acidification in an Arctic sea-ice-associated diatom compared to a pelagic diatom. New Phytol. 2020, 226, 1708–1724. [Google Scholar] [CrossRef]
- McMinn, A.; Hegseth, E.N. Early Spring pack ice in the Arctic and Antarctic. In Antarctic Biology in a Global Context, Proceedings of the VIIIth SCAR International Biology Symposium, Vrije Universitet, Amsterdam, The Netherlands, 27 August—1 September 2001; Huiskes, A.H.L., Gieskes, W.W.C., Rozema, J., Schorna, R.M.L., van der Vies, S.M., Wolff, W.J., Eds.; Backhuys Pub: Leiden, The Netherlands, 2003; pp. 182–186. [Google Scholar]
- Olsen, L.M.; Laney, S.R.; Duarte, P.; Kauko, H.M.; Fernandez-Mendez, M.; Mundy, C.J.; Rösel, A.; Meyer, A.; Itkin, P.; Cohen, L.; et al. The seeding of ice algal blooms in Arctic pack ice: The multiyear ice seed repository hypothesis. J. Geophys. Biogeosci. 2017, 122, 1529–1548. [Google Scholar] [CrossRef]
- Kohlbach, D.; Hop, H.; Wold, A.; Schmidt, K.; Smik, L.; Belt, S.T.; Keck Al-Habahbeh, A.; Woll, M.; Graeve, M.; Dabrowska, A.M.; et al. Multiple Trophic Markers Trace Dietary Carbon Sources in Barents Sea Zooplankton During Late Summer. Front. Mar. Sci. 2021, 7, 610248. [Google Scholar] [CrossRef]
- Flores, H.; David, C.; Ehrlich, J.; Hardge, K.; Kohlbach, D.; Lange, B.A.; Niehoff, B.; Nöthig, E.-M.; Peeken, I.; Metfies, K. Sea-ice properties and nutrient concentration as drivers of the taxonomic and trophic structure of high-Arctic protist and metazoan communities. Polar Biol. 2019, 42, 1377–1395. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Date | Position | Ice Thickness, Sampling Site (cm) | Snow Depth (cm) | Ice Type and Thickness | Chlorophyll (mg m−2) | Cell Numbers (×106 m−2) | Nitrate (µmol L−1) | Phosphate (µmol L−1) | Silicate (µmol L−1) | Under-Ice Irradiance (µmol m−2 s−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1986 | 17–19 April | 74°54′–75°54′ N 30°45′–32°59′ E | 100–110 | 5–14 | Annual, flat and rafted (80–150) | 11.8 (1.31–17.7), n = 4 | 2065 (163–5695) n = 4 | 10.3 (9.5–11.0) n = 2 | 0.83 (0.75–0.91) n = 2 | 6.1 (5.7–6.5) n = 2 | 9 (5–14) n = 3 |
| 1990 | 20–28 April | 76°46′–77°03′ N 23°44′–24°27′ E | 100–140 | 20–30 | Annual, flat and rafted and thicker multi-year floes | 0.45 (0.31–0.65), n = 4 | 41 (35–46) n = 2 | - | 0.85 (0.6–1.1) n = 2 | 2.5 (1.5–3.5) n = 2 | 34 (23–45) n = 2 |
| 1995 | 15–21 June | 77°30′–78°05′ N 34°17′–34°36′ E | 100–200 | 5–10 | Annual, flat and rafted (100–200) | 0.11 (0.03–0.21), n = 6 | 300 (8–560) n = 5 | 2.0 (1.5–2.6) n = 3 | 0.26 (0.2–0.3) n = 3 | 1.63 (1.1–2.0) n = 3 | - |
| 1996 | 15–25 May | 75°02′–75°14′ N 24°54′–26°27′ E | 40–200 | 10–30 | Annual, flat and rafted, with caves (40–200) | 7.9 (0.80–39.0), n = 17 | 2520 (190–15,400) n = 17 | 6.3 (4.3–8.1) n = 4 | 0.47 (0.4–0.6) n = 4 | 2.75 (1.4–3.9) n = 4 | 28 (1–58) n = 7 |
| 1996 | 28 July–09 August | 74°40′–81°34′ N 25°52′–34°50′ E | 150–200 | 5–15 | Old annual, rafted, with caves and large brine channels (150–340) | 2.3 (0.03–9.60), n = 10 | 800 (81–2140) n = 10 | Below detection | 0.08 (0.04–0.2) n = 4 | 0.22 (0.05–0.4) n = 4 | 6 (2–19) n = 5 |
| 1997 | 20–25 May | 76°10′–76°30′ N 29°56′–30°13′ E | 30–80 | 2–20 | Annual, flat, some rafted (30–300) | 0.18 (0.04–0.48), n = 20 | 82 (18–270) n = 18 | Below detection | 0,07 (0.06–0.09) n = 4 | 0.82 (0.6–1.3) n = 4 | 72 (10–152) n = 10 |
| 2001 | 09–16 April | 74°56′–75°20′ N 19°17′–23°59′ E | 60–120 | 5–20 | Annual, flat, thicker floes between thin new ice | 0.26 (0.01–0.67), n = 8 | - | 9.9 (9.0 -11.0) n = 4 | 0.85 (0.83–0.87) n = 4 | 5.52 (5.5–5.9) n = 4 | 14 (3–31) n = 4 |
| 2003 | 12–26 May | 79°31′ 80°49′ N 04°51′–11°37′ E | 40 | 10–15 | Thin, annual ice and thick, rafted multi-year ice (40–300) | 0.21 (0.17–0.26), n = 4 | 47 (33–60) n = 2 | 2.2 (2.1–2.4) n = 2 | 0.22 (0.20–0.23) n = 2 | 3.36 (3.1–3.6) n = 2 | 13 (12–14) n = 2 |
| 2003 | 23–25 August | 80°51′–81°03′ N 28°22′–28°39′ E | 110–270 | 2–20 | Multi-year ice (100–200) | 5.58 (2.46–8.70), n = 2 | 850 (120–1260) n = 2 | 1.5 (0.5–2.5) n = 2 | 0.22 (0.20–0.23) n = 2 | 2.05 (1.8–2.3) n = 2 | 29 (13–45) n = 2 |
| 2004 | 23–31 May | 77°03′–77°13′ N 29°17′–29°52′ E | 100–150 | 10–30 | Large, annual floe, flat (100–150) | 16.3 (6.78–35.4), n = 26 | 1610 (670–2850) n = 16 | 1.3 (0.0–3.1) n = 5 | 0.19 (0.09–0.32) n = 5 | 2.86 (2.3–3.6) n = 5 | 16 (5–41) n = 11 |
| Area | Time of Year | Dominating and Common Species |
|---|---|---|
| Outermost ice zone, 0–30 cm (Zone A) | March–April | Centric diatoms: Thalassiosira spp., Porosira glacialis, Bacterosira bathyomphala, Chaetoceros spp. |
| Average thick ice, 40–90 cm (Zone B) | April–June | Pennate diatoms: Nitzschia frigida, N. promare, Fragilariopsis oceanica, Navicula kariana, Pseudogomphonema arcticum, Fossulaphycus arcticus, Synedropsis hyperborea, Navicula vanhoeffenii, Centric diatoms: Porosira glacialis, Schinodiscus bioculatus |
| Thick annual ice, 100–120 cm (Zone C) | May–July | Pennate diatoms: Nitzschia frigida, N. promare, Fossulaphycus arcticus, Centric diatoms: Melosira arctica, Attheya septentrionalis |
| Multi-year ice, 150–200 cm (Zone D) | Probably all year | Centric diatoms: Melosira arctica, Attheya septentrionalis Pennate diatoms: Ppseudogomphonema arcticum, Synedropsis hyperborea |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hegseth, E.N.; von Quillfeldt, C. The Sub-Ice Algal Communities of the Barents Sea Pack Ice: Temporal and Spatial Distribution of Biomass and Species. J. Mar. Sci. Eng. 2022, 10, 164. https://doi.org/10.3390/jmse10020164
Hegseth EN, von Quillfeldt C. The Sub-Ice Algal Communities of the Barents Sea Pack Ice: Temporal and Spatial Distribution of Biomass and Species. Journal of Marine Science and Engineering. 2022; 10(2):164. https://doi.org/10.3390/jmse10020164
Chicago/Turabian StyleHegseth, Else Nøst, and Cecilie von Quillfeldt. 2022. "The Sub-Ice Algal Communities of the Barents Sea Pack Ice: Temporal and Spatial Distribution of Biomass and Species" Journal of Marine Science and Engineering 10, no. 2: 164. https://doi.org/10.3390/jmse10020164
APA StyleHegseth, E. N., & von Quillfeldt, C. (2022). The Sub-Ice Algal Communities of the Barents Sea Pack Ice: Temporal and Spatial Distribution of Biomass and Species. Journal of Marine Science and Engineering, 10(2), 164. https://doi.org/10.3390/jmse10020164