
Taxonomy and Distribution of the Deep-Sea Batfish Genus Halieutopsis (Teleostei: Ogcocephalidae), with Descriptions of Five New Species †

Abstract
1. Introduction
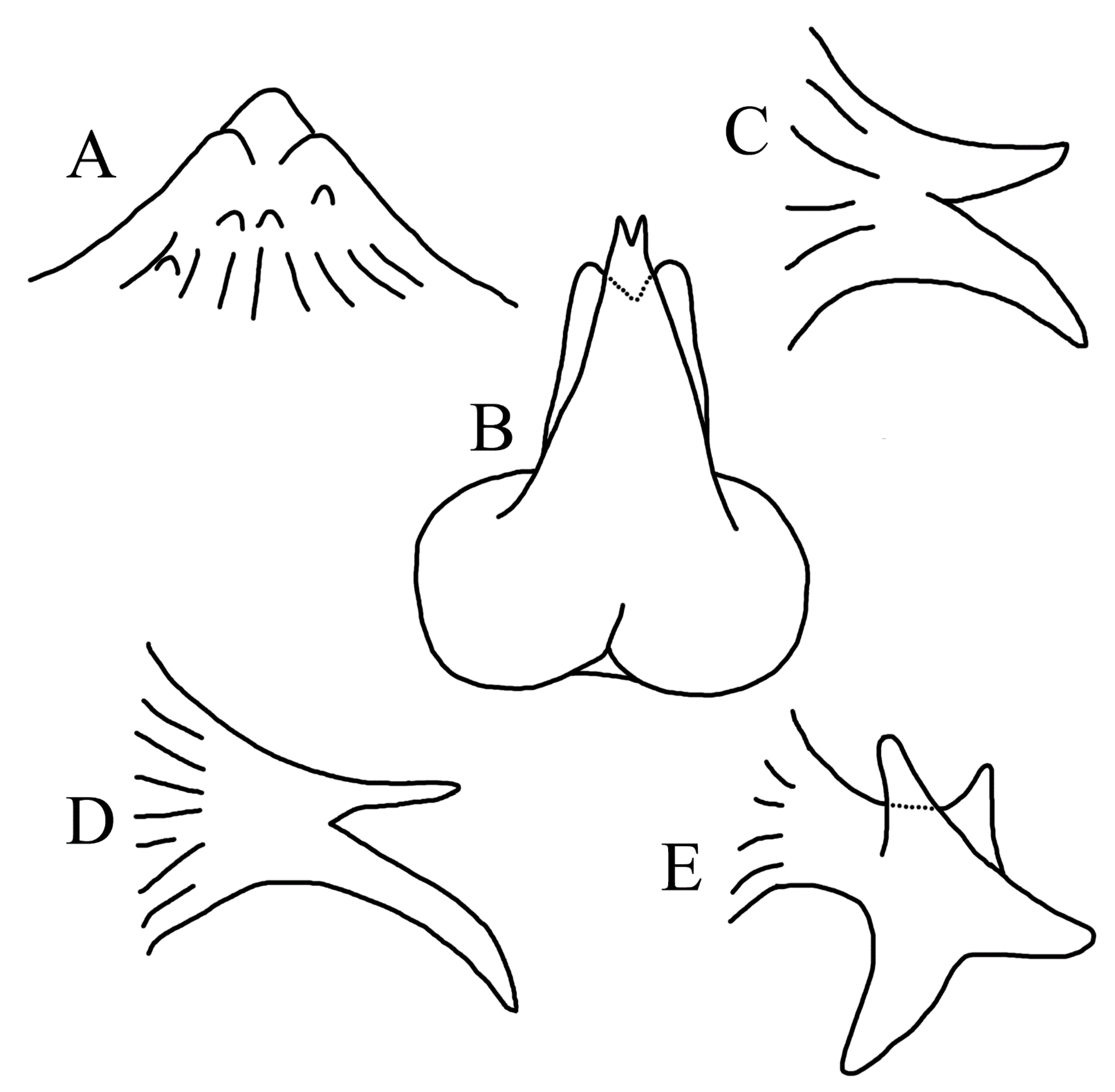
2. Materials and Methods
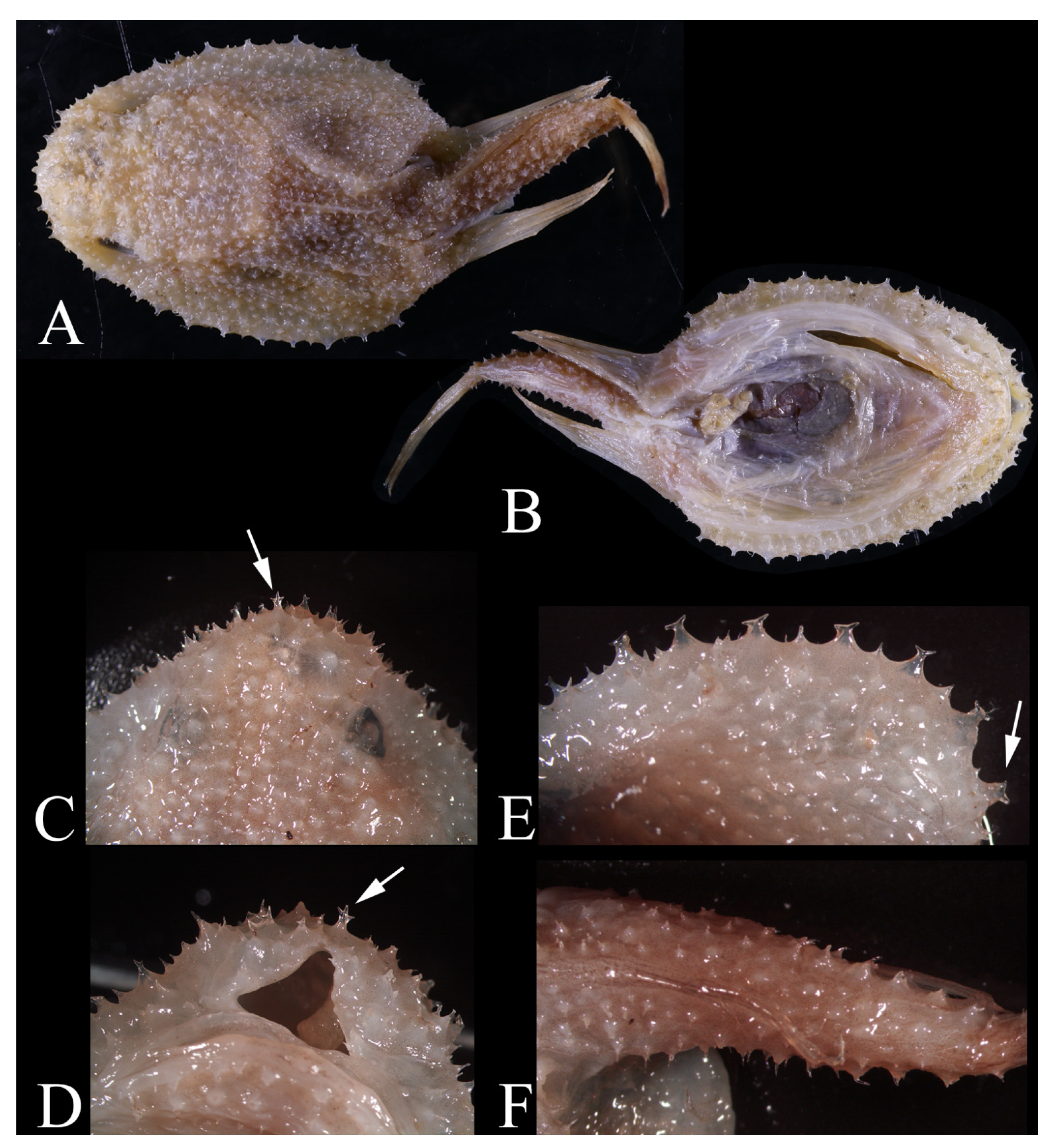
3. Results
3.1. Family Ogcocephalidae
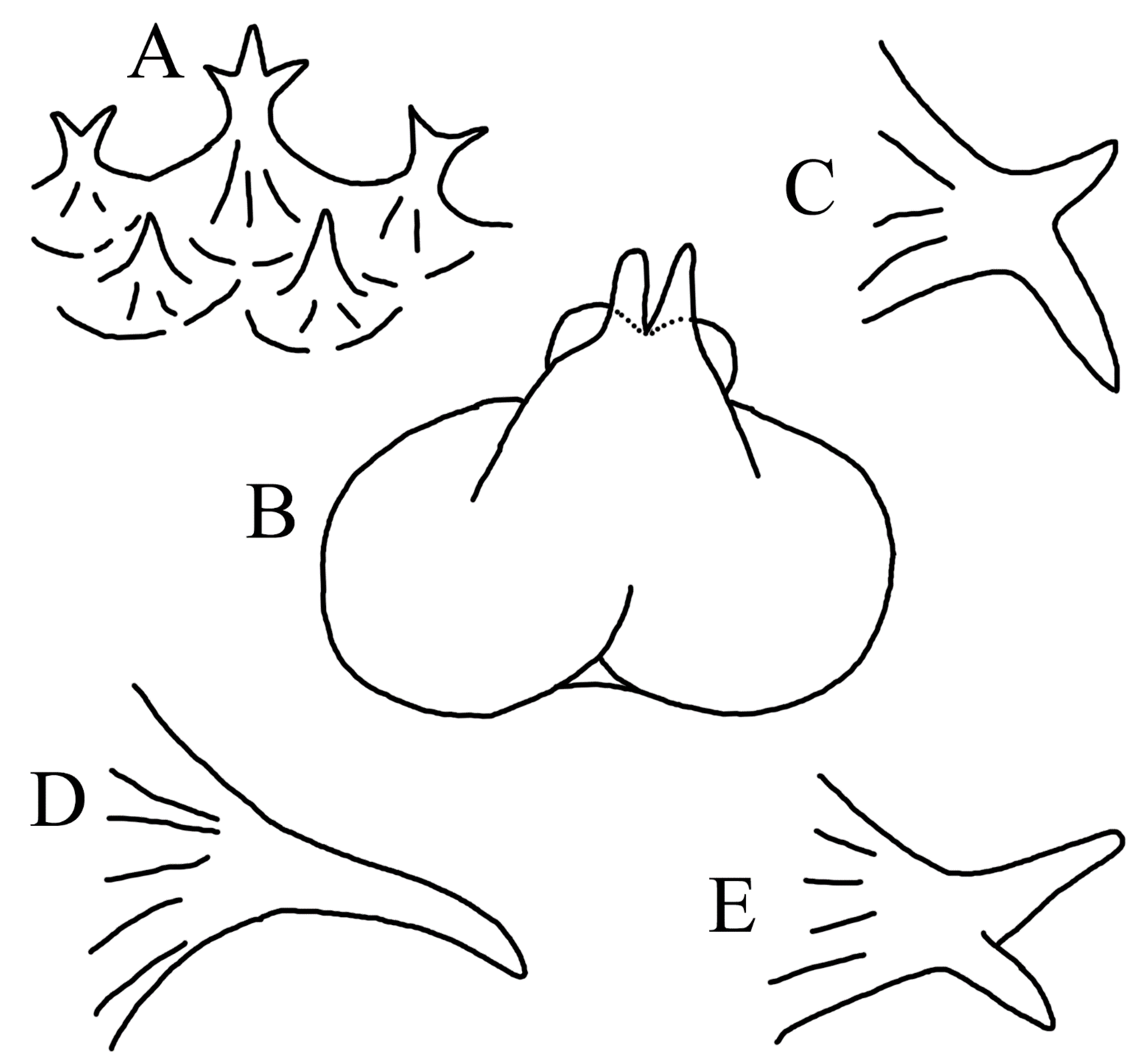
3.1.1. Halieutopsis Garman, 1899
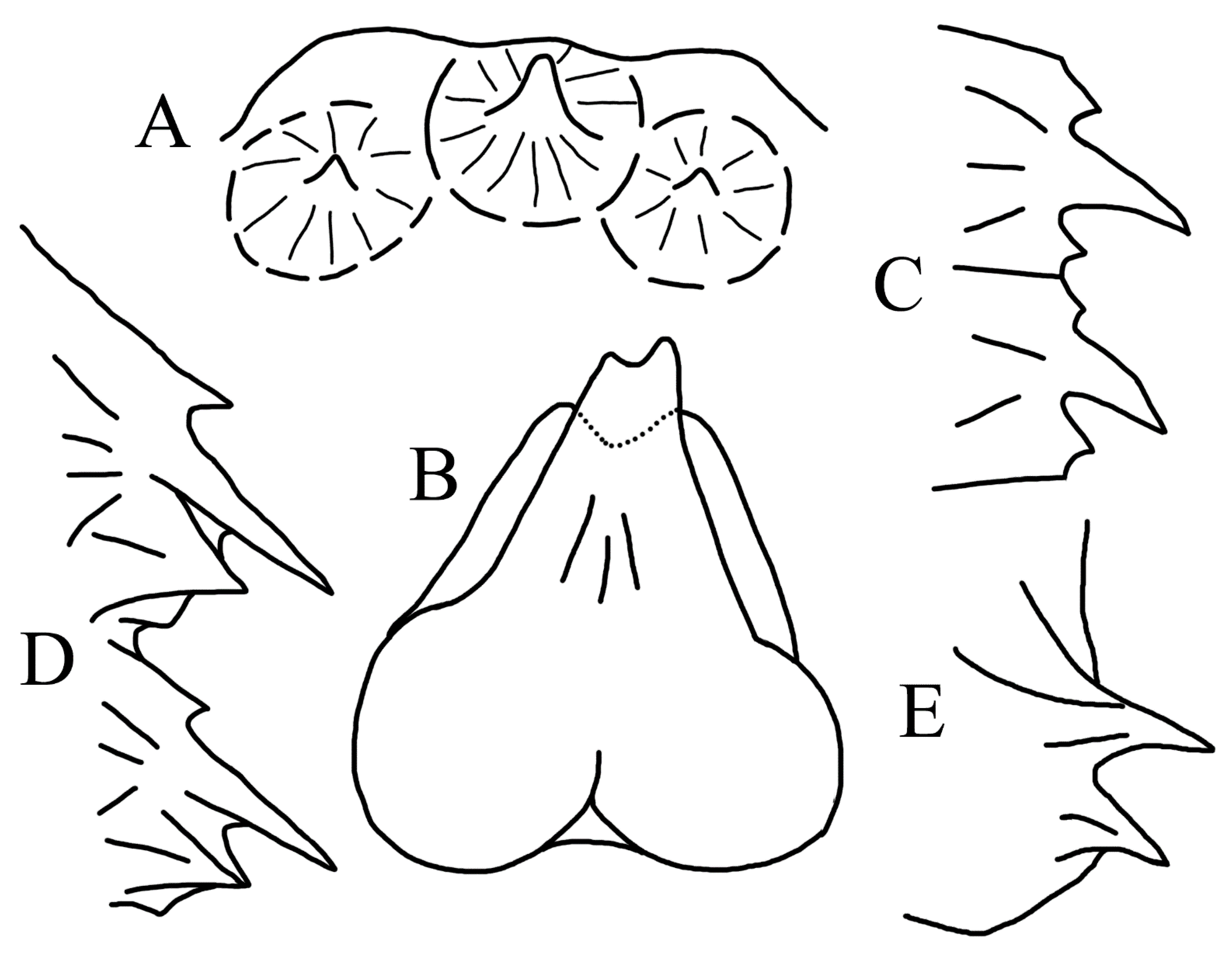
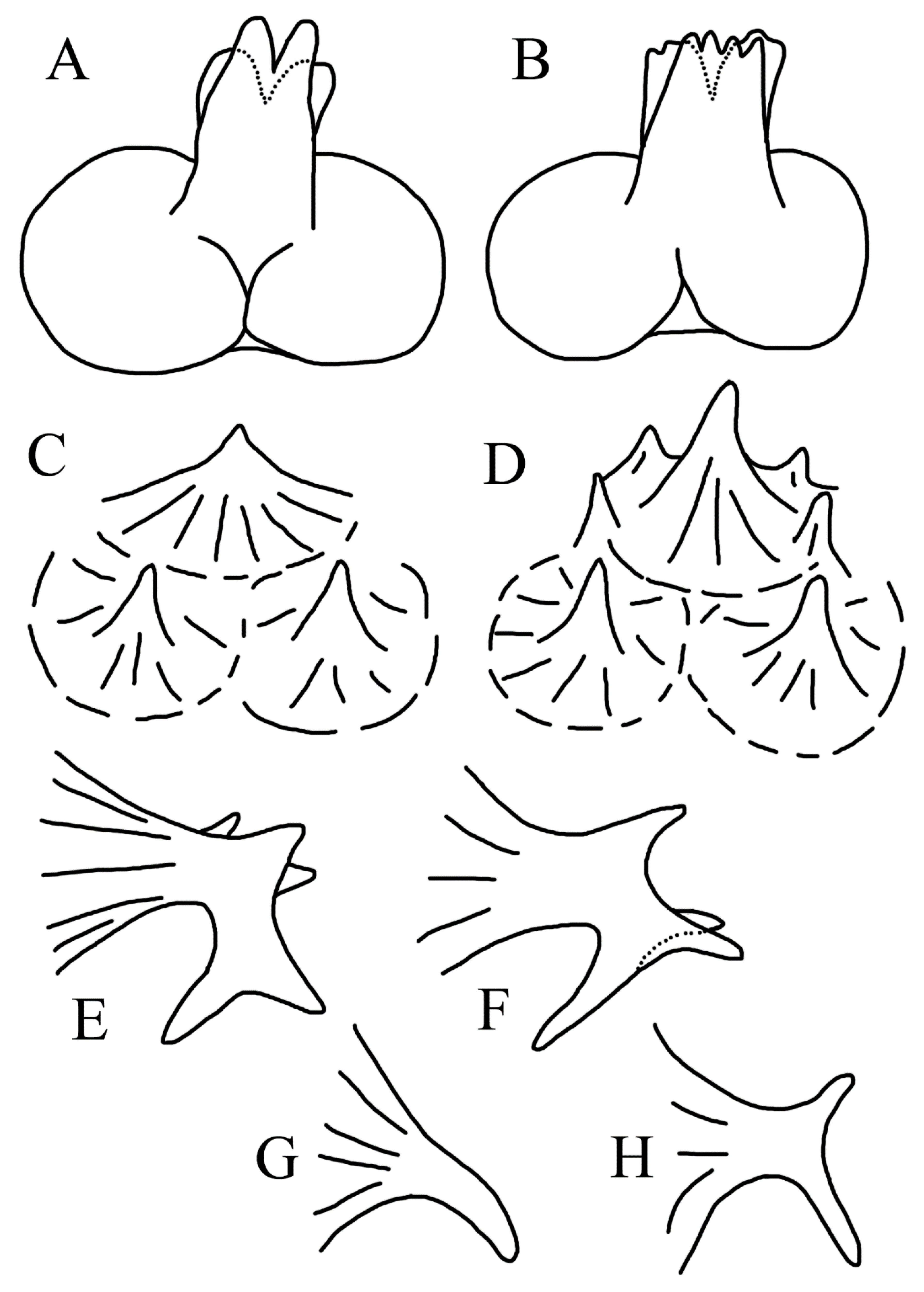
3.1.2. Key to Species of Halieutopsis
3.1.3. Halieutopsis andriashevi Bradbury, 1988
3.1.4. Halieutopsis bathyoreos Bradbury, 1988
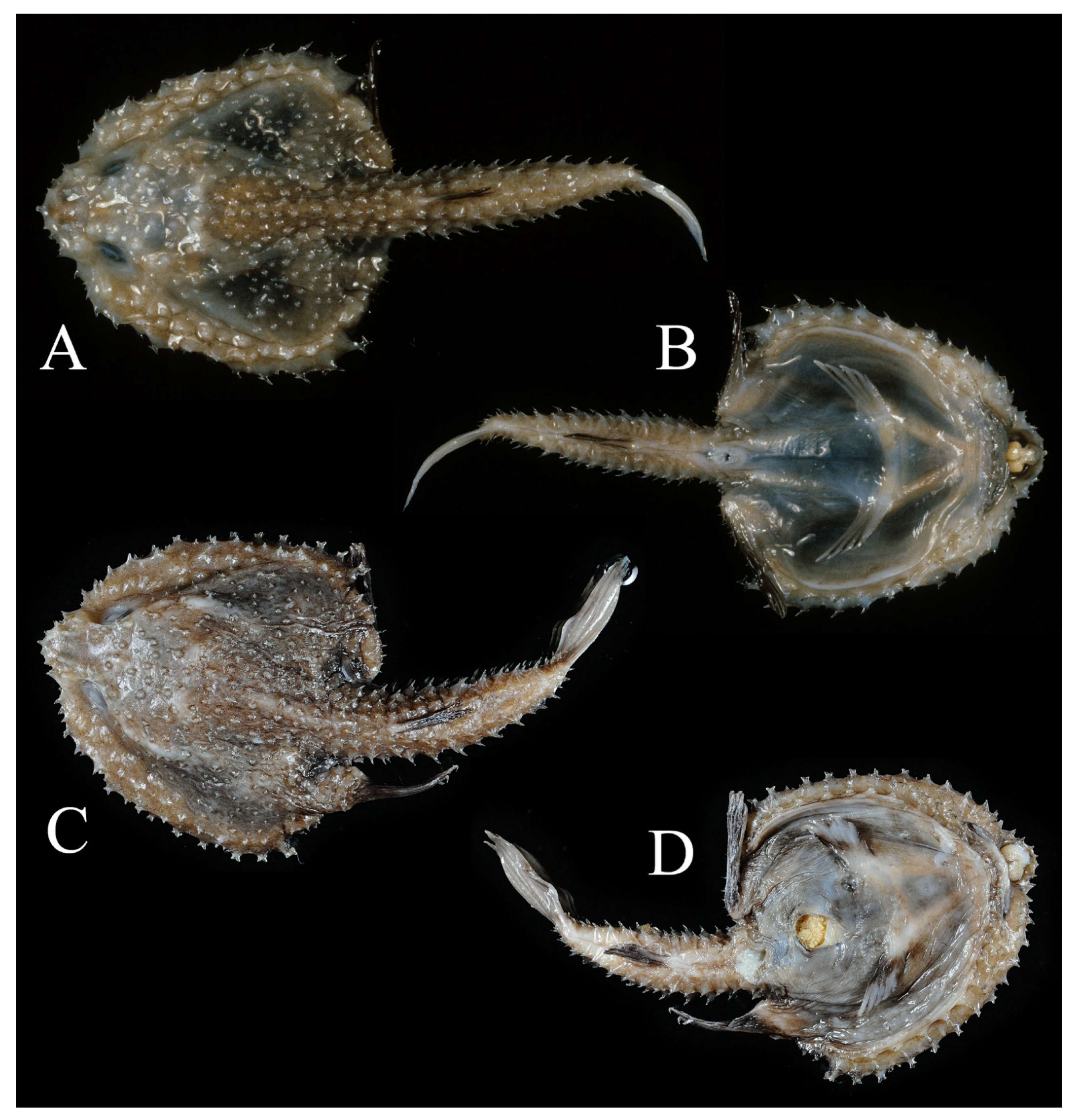
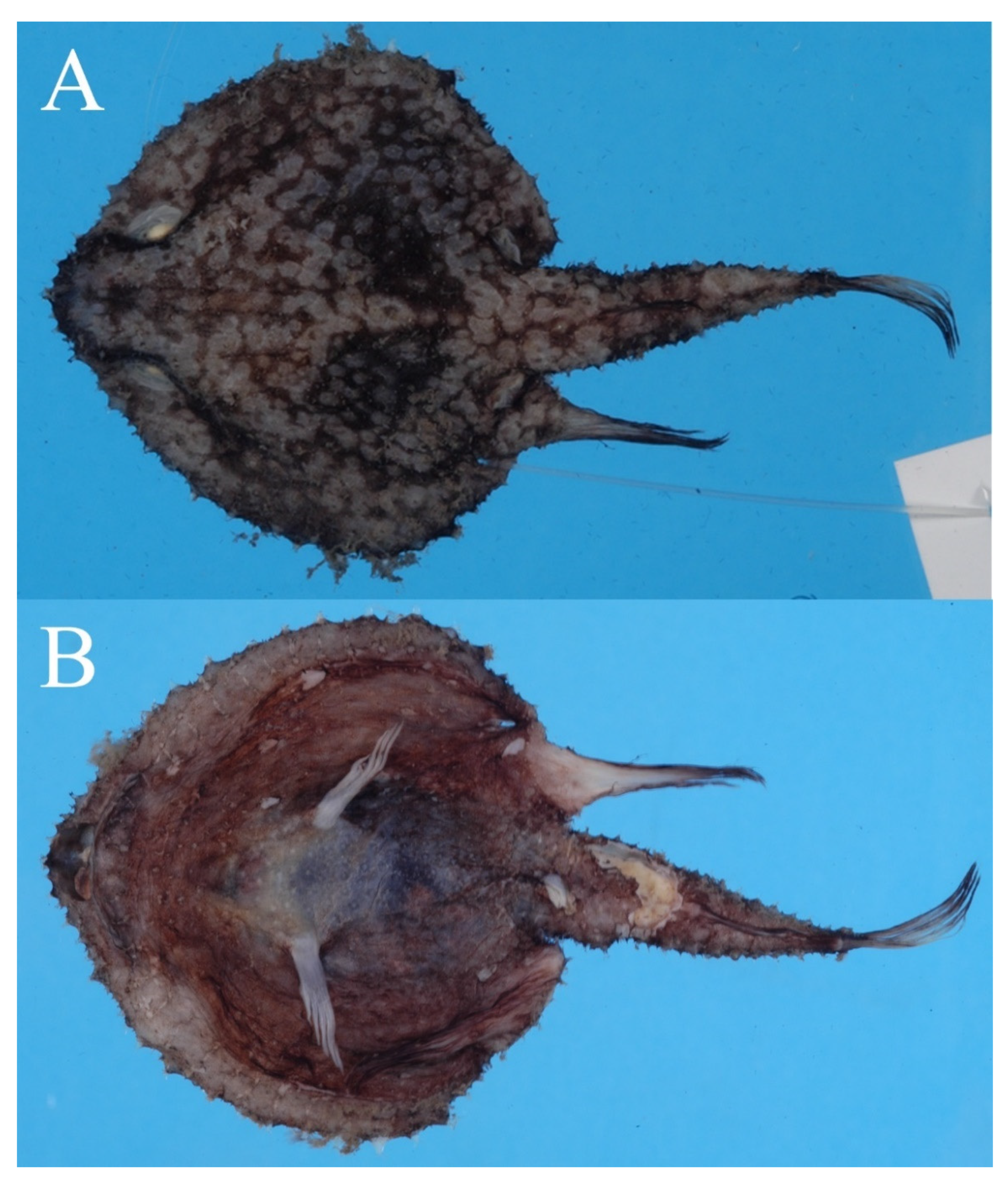
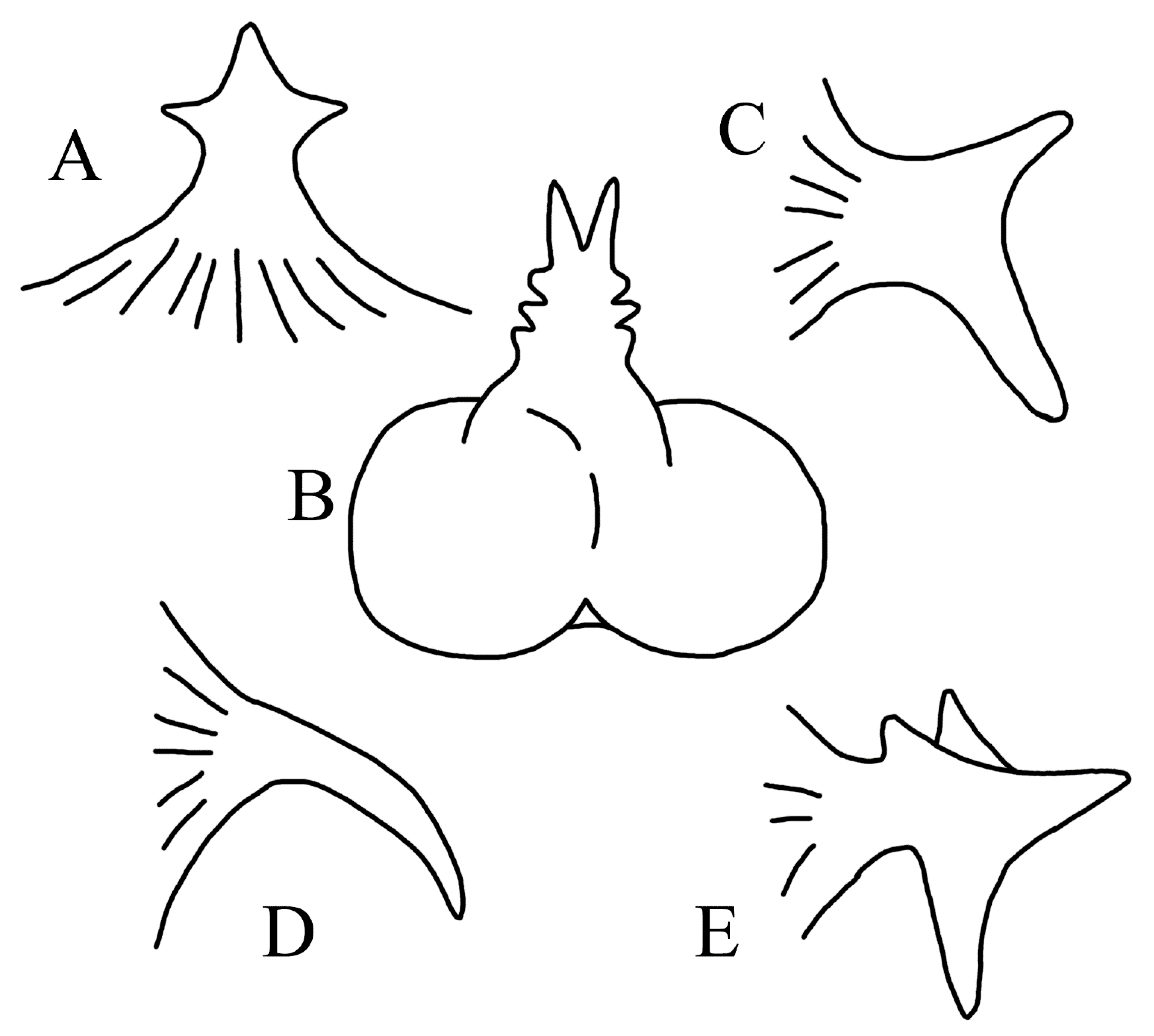
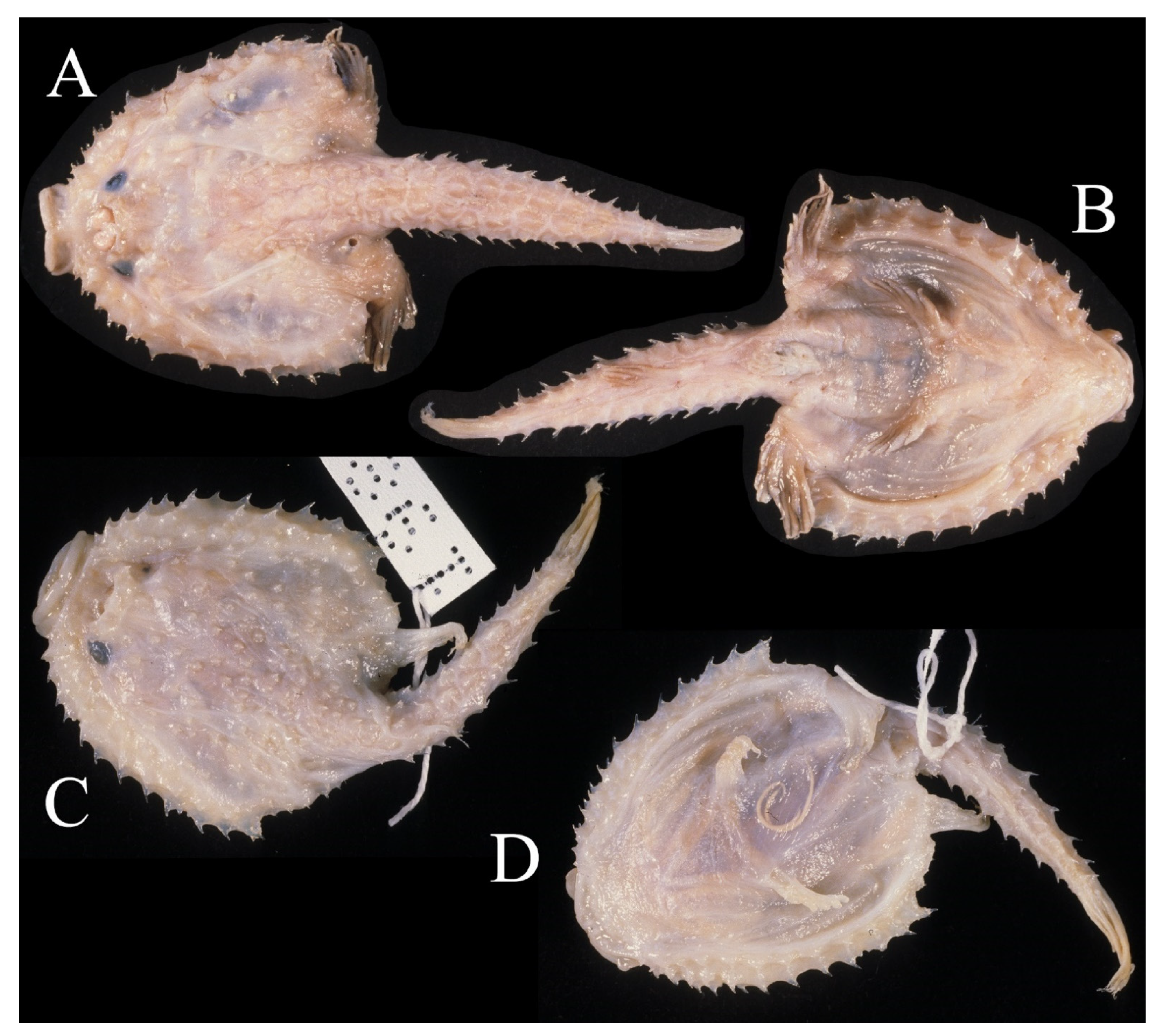
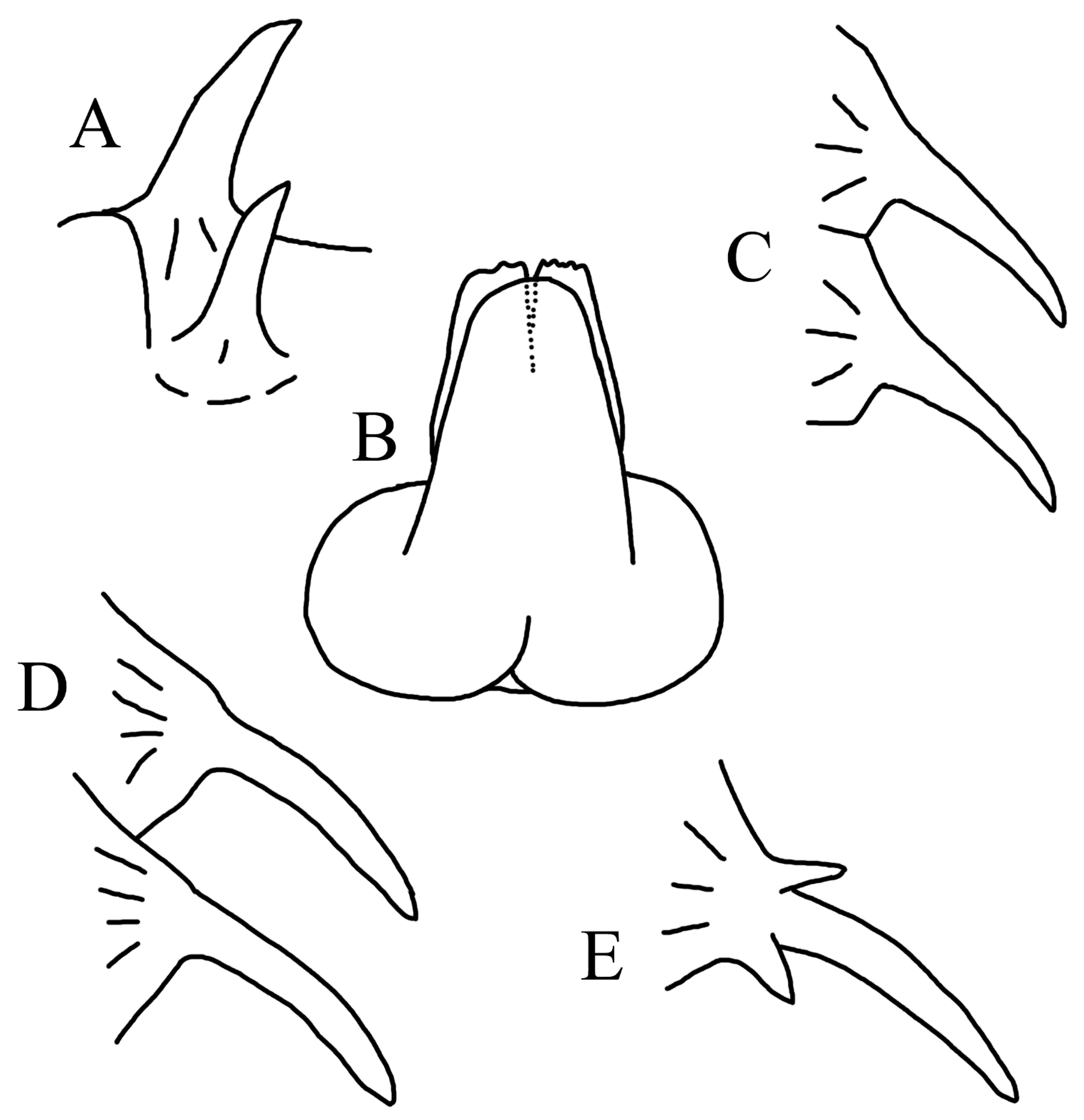
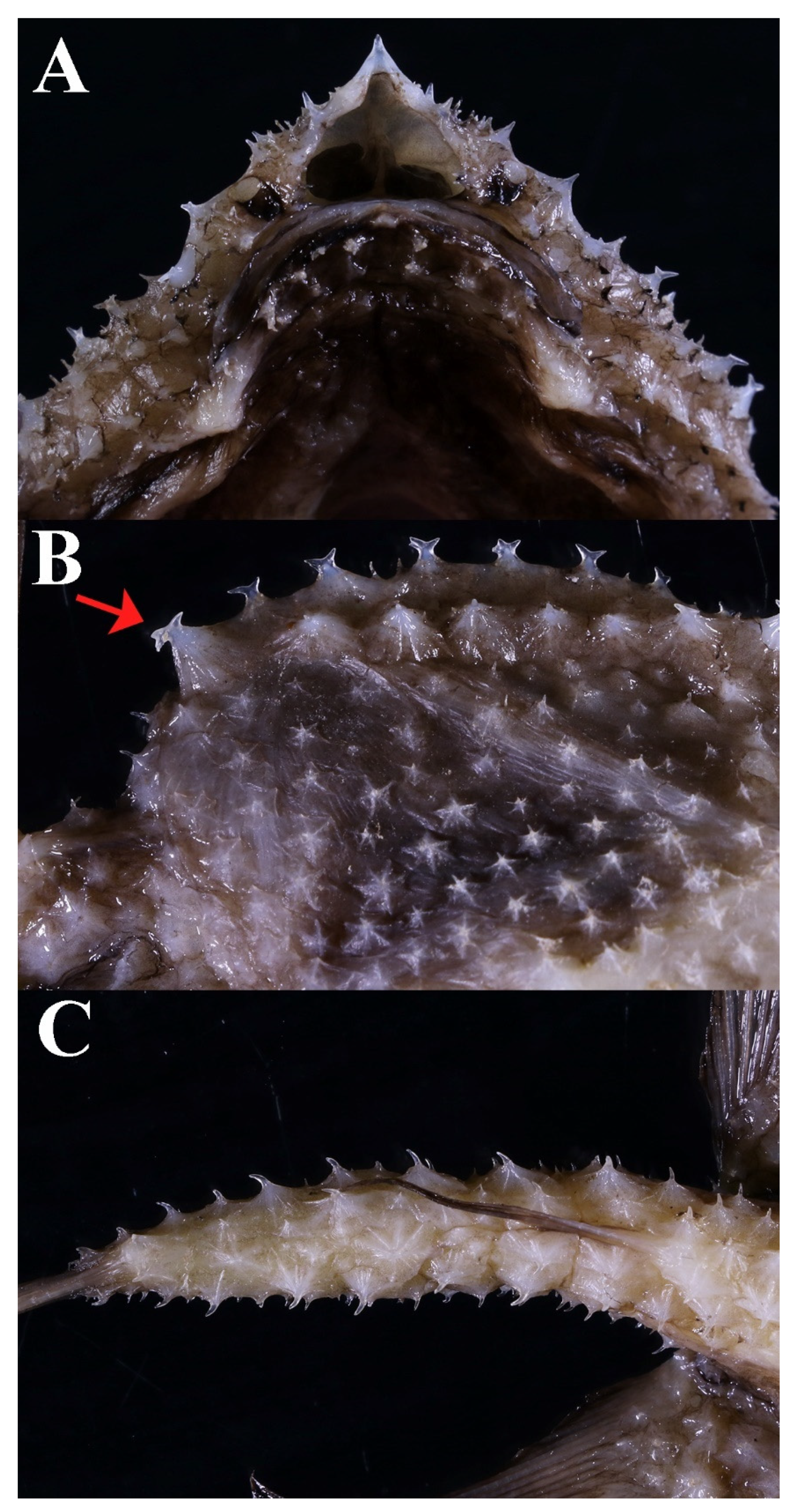
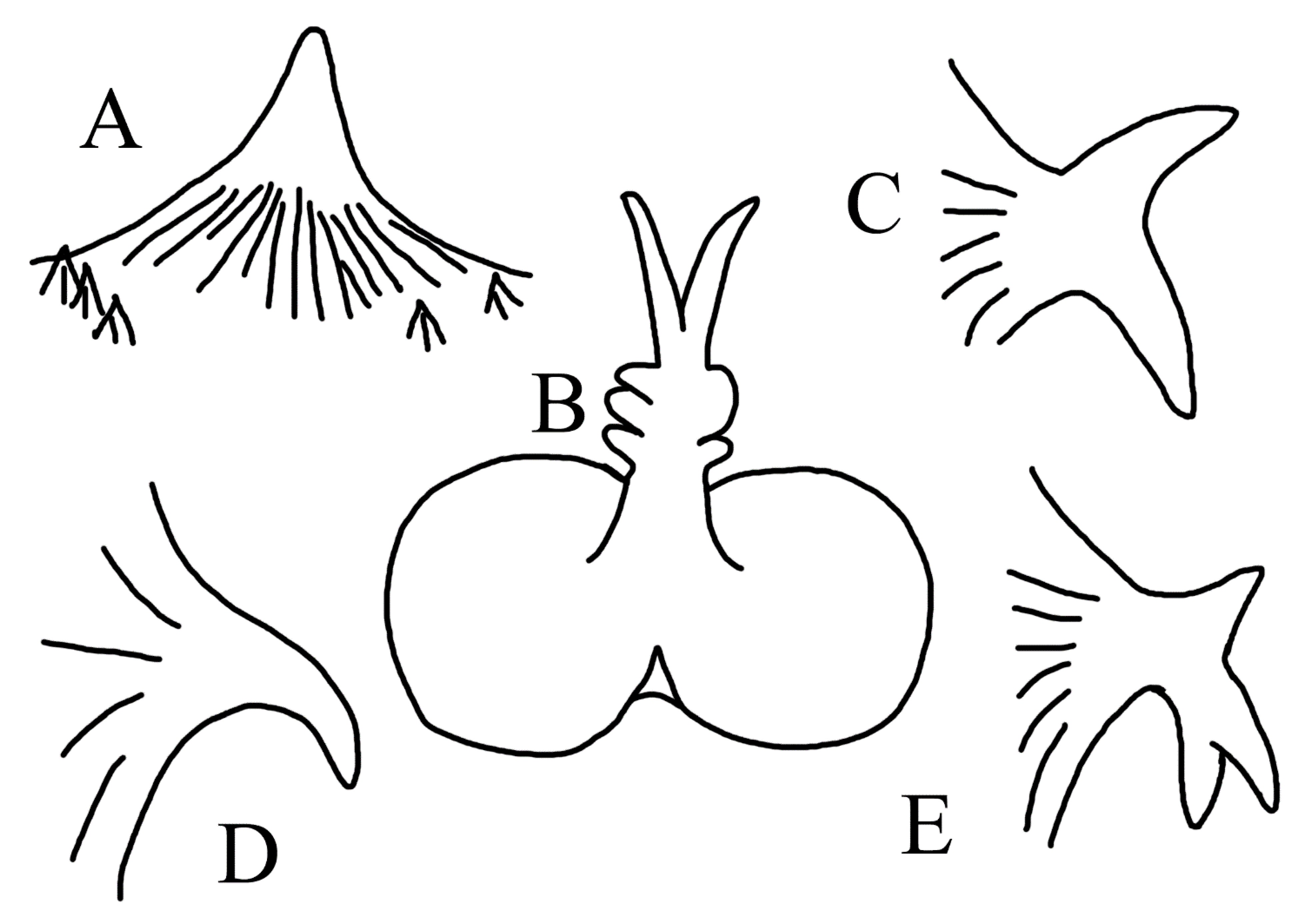
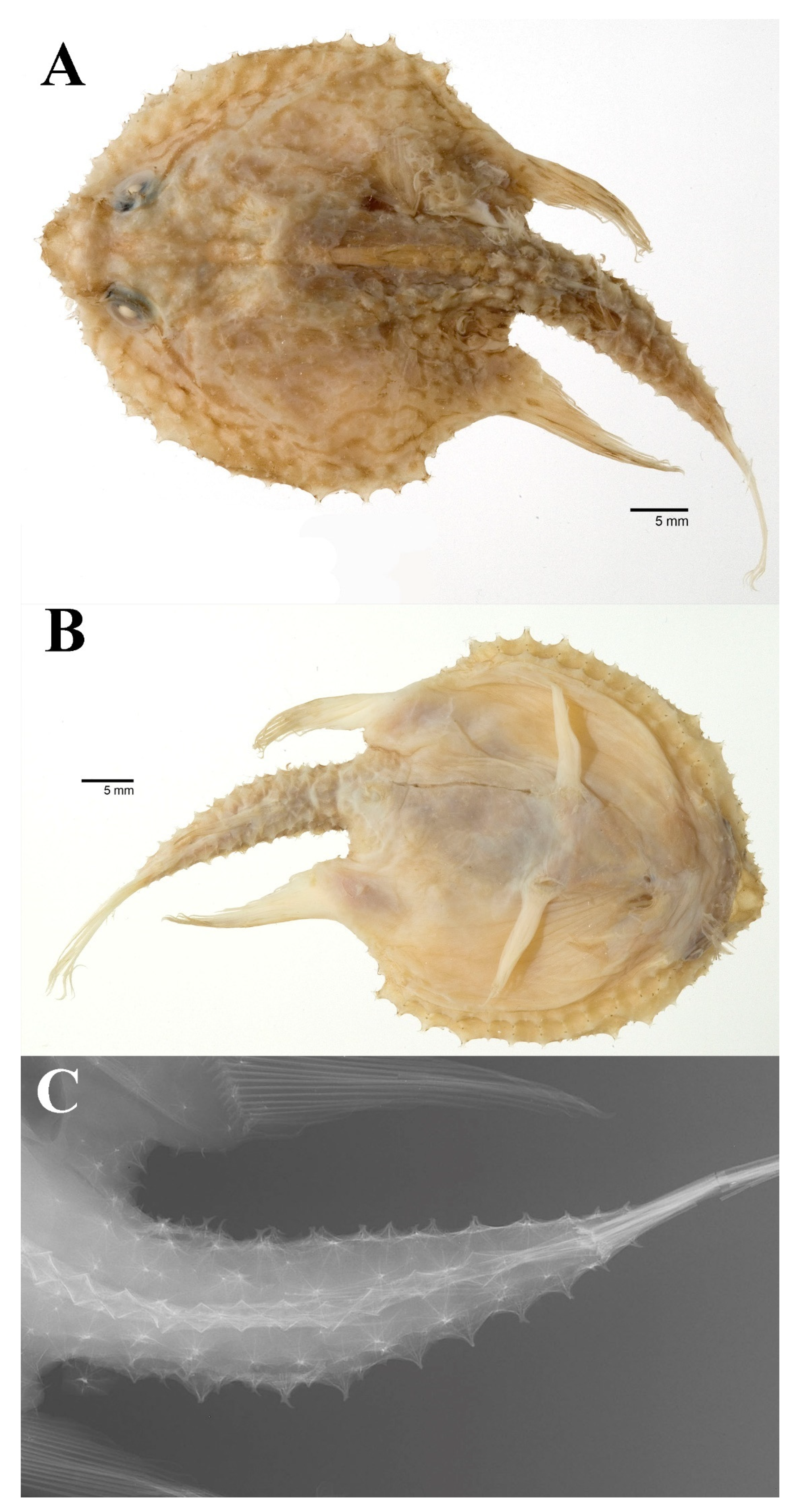
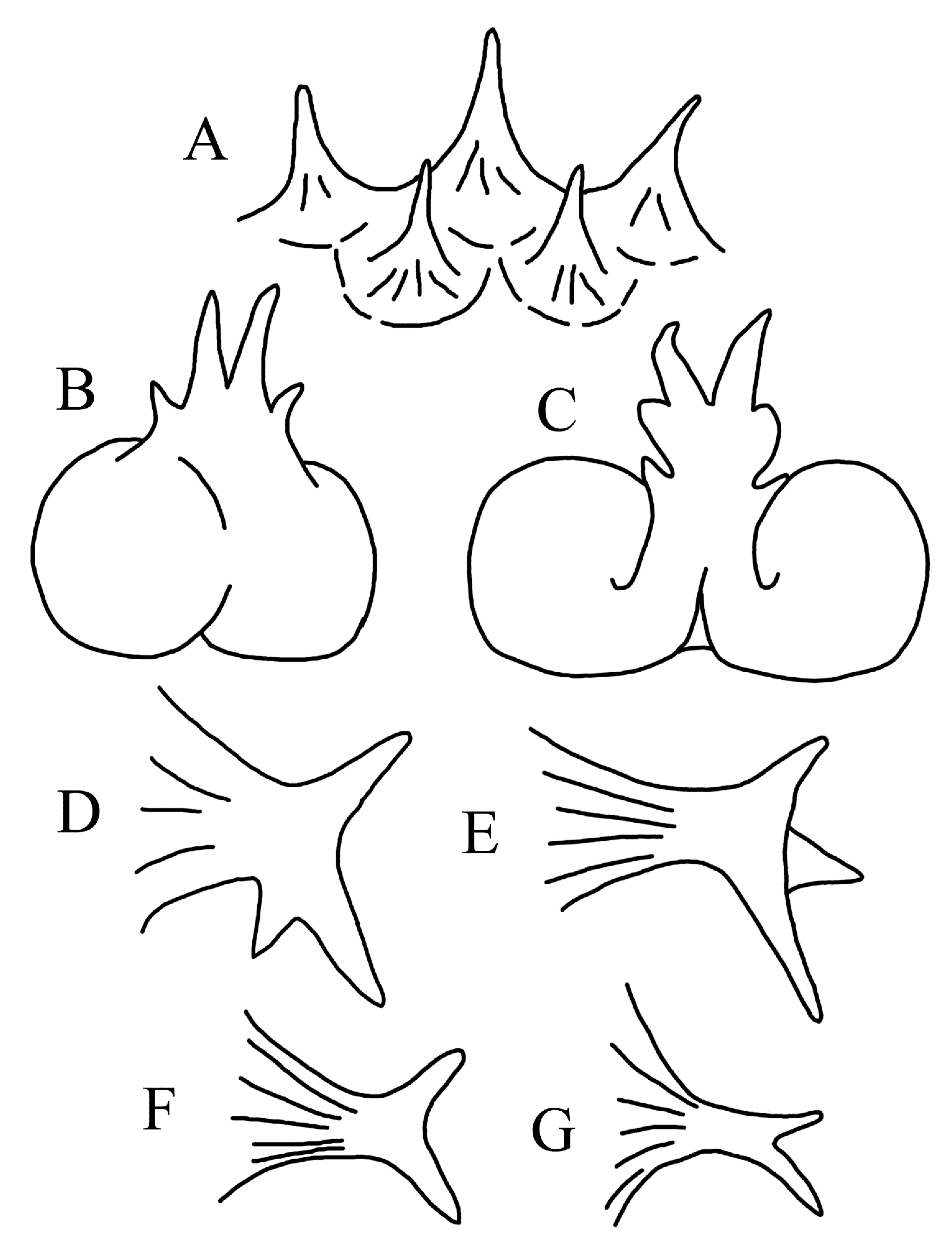
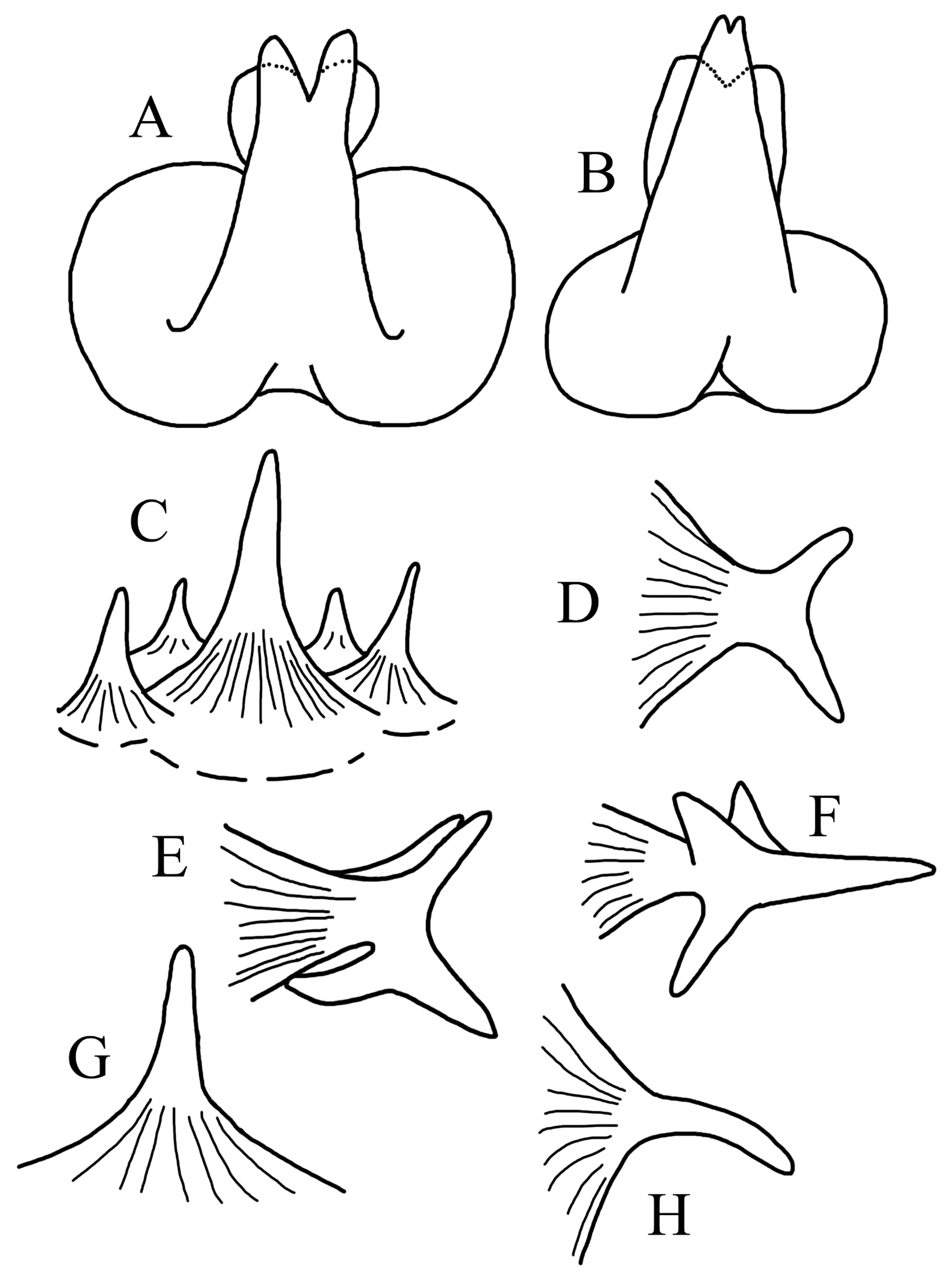
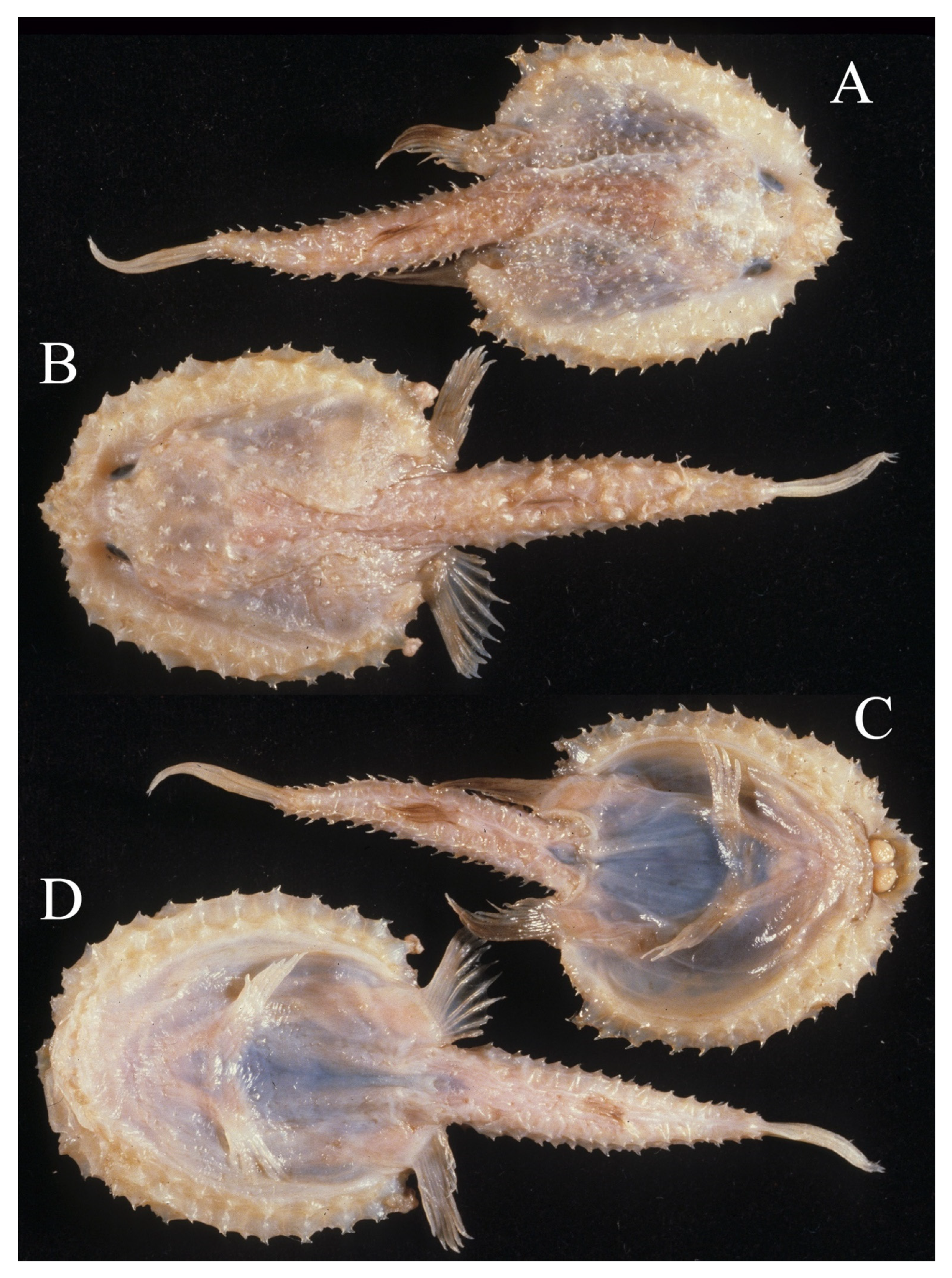
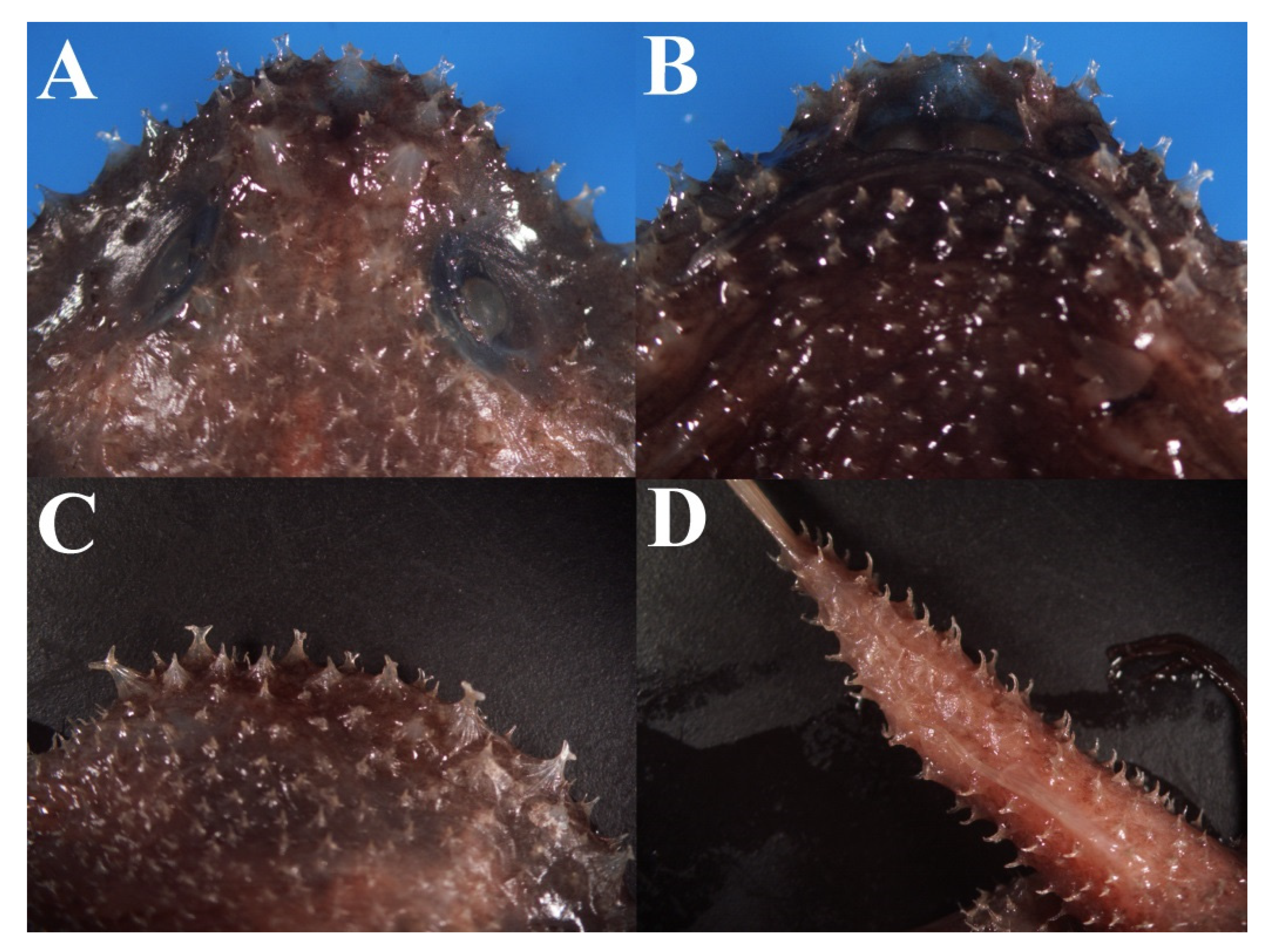
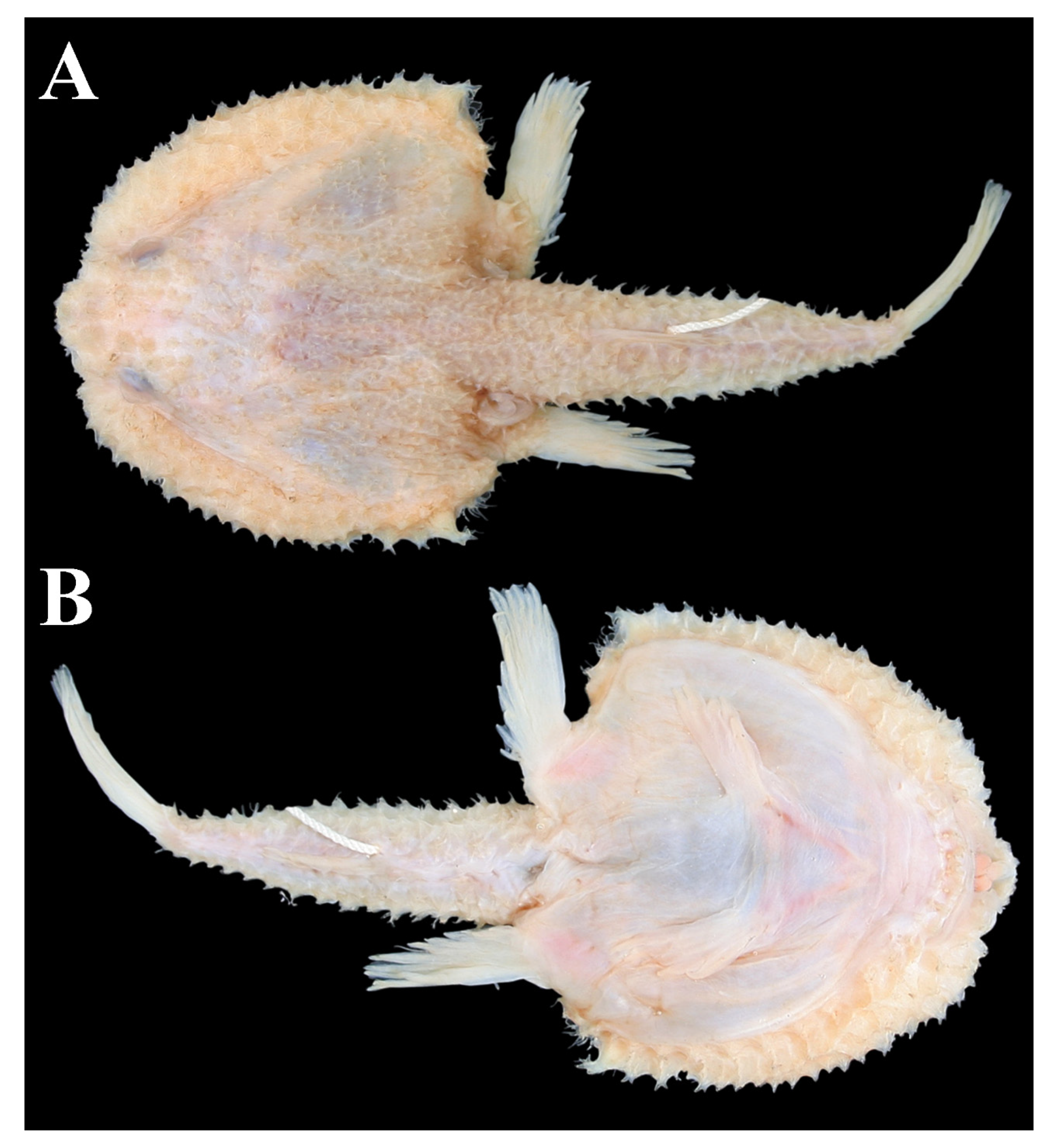
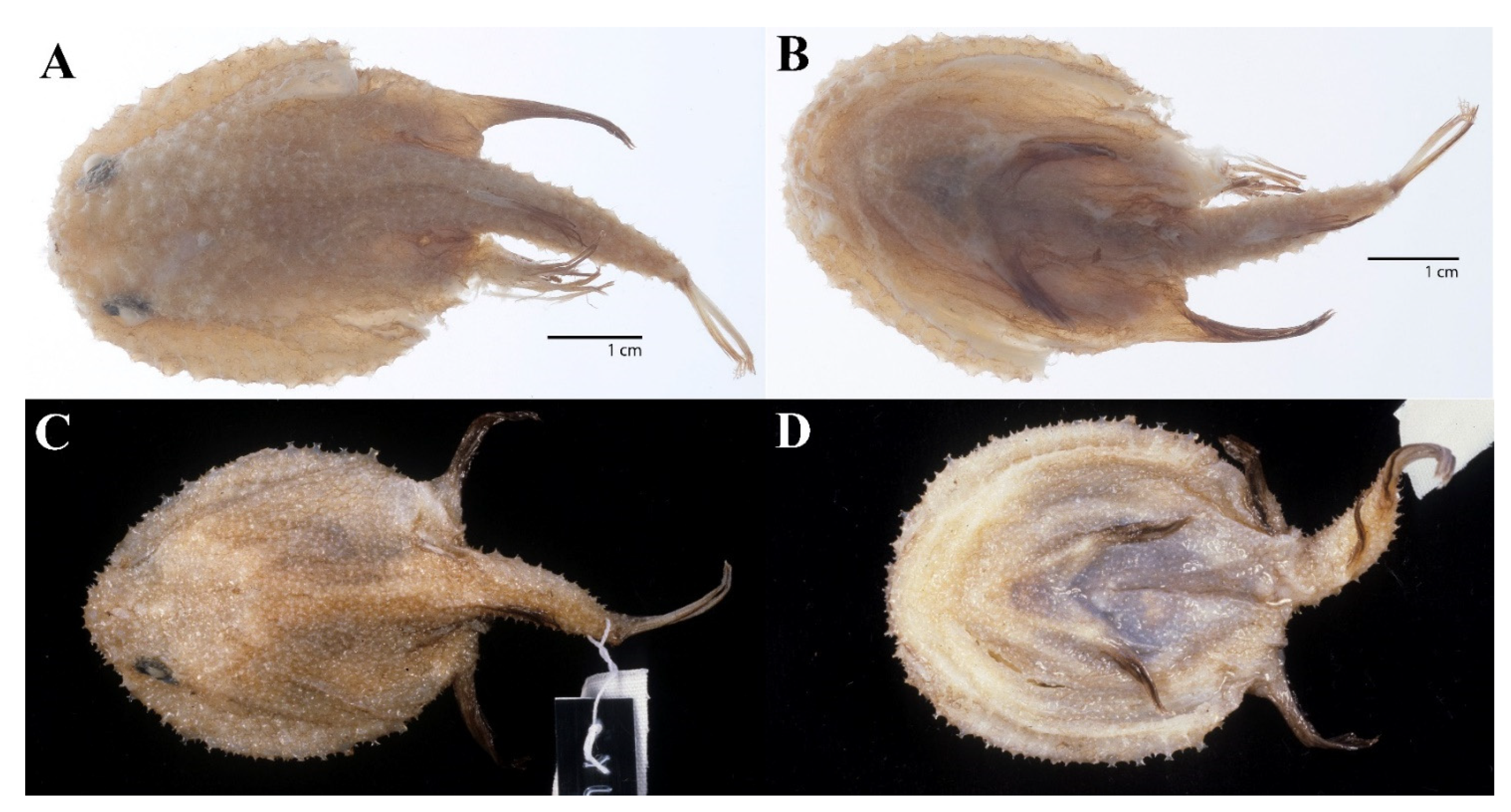
3.1.5. Halieutopsis echinoderma sp. nov.
3.1.6. Halieutopsis galatea Bradbury, 1988
3.1.7. Halieutopsis ingerorum Bradbury, 1988
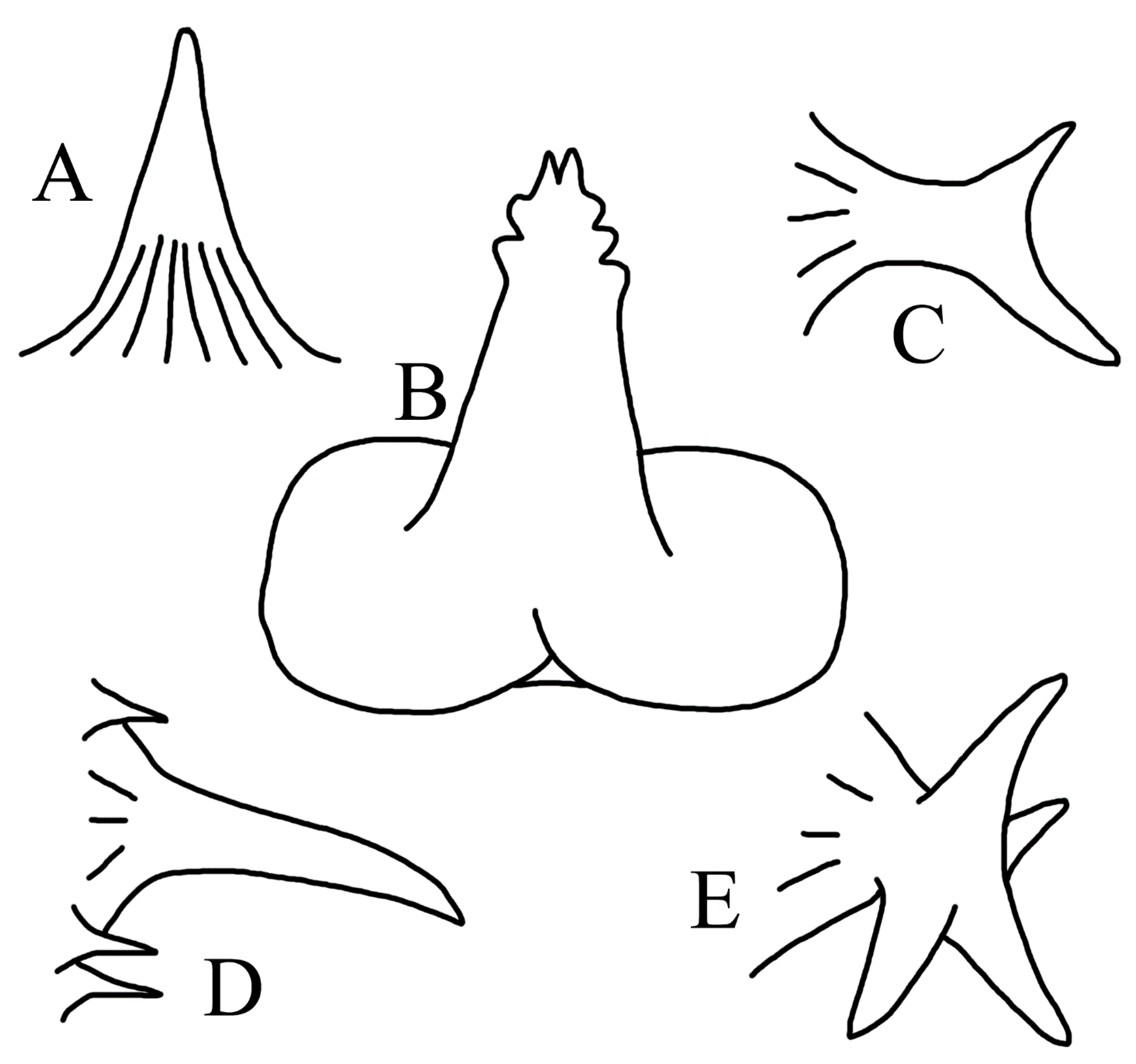
3.1.8. Halieutopsis kawaii sp. nov.
3.1.9. Halieutopsis margaretae Ho & Shao, 2007
3.1.10. Halieutopsis murrayi sp. nov.
3.1.11. Halieutopsis nasuta (Alcock, 1891)
3.1.12. Halieutopsis nudiventer (Lloyd, 1909)
3.1.13. Halieutopsis oblonga (Smith & Radcliffe, 1912)
3.1.14. Halieutopsis okamurai sp. nov.
3.1.15. Halieutopsis simula (Smith & Radcliffe, 1912)
3.1.16. Halieutopsis stellifera (Smith & Radcliffe, 1912)
3.1.17. Halieutopsis taiwanea sp. nov.
3.1.18. Halieutopsis tumifrons Garman, 1899
4. Discussion
4.1. Systematics
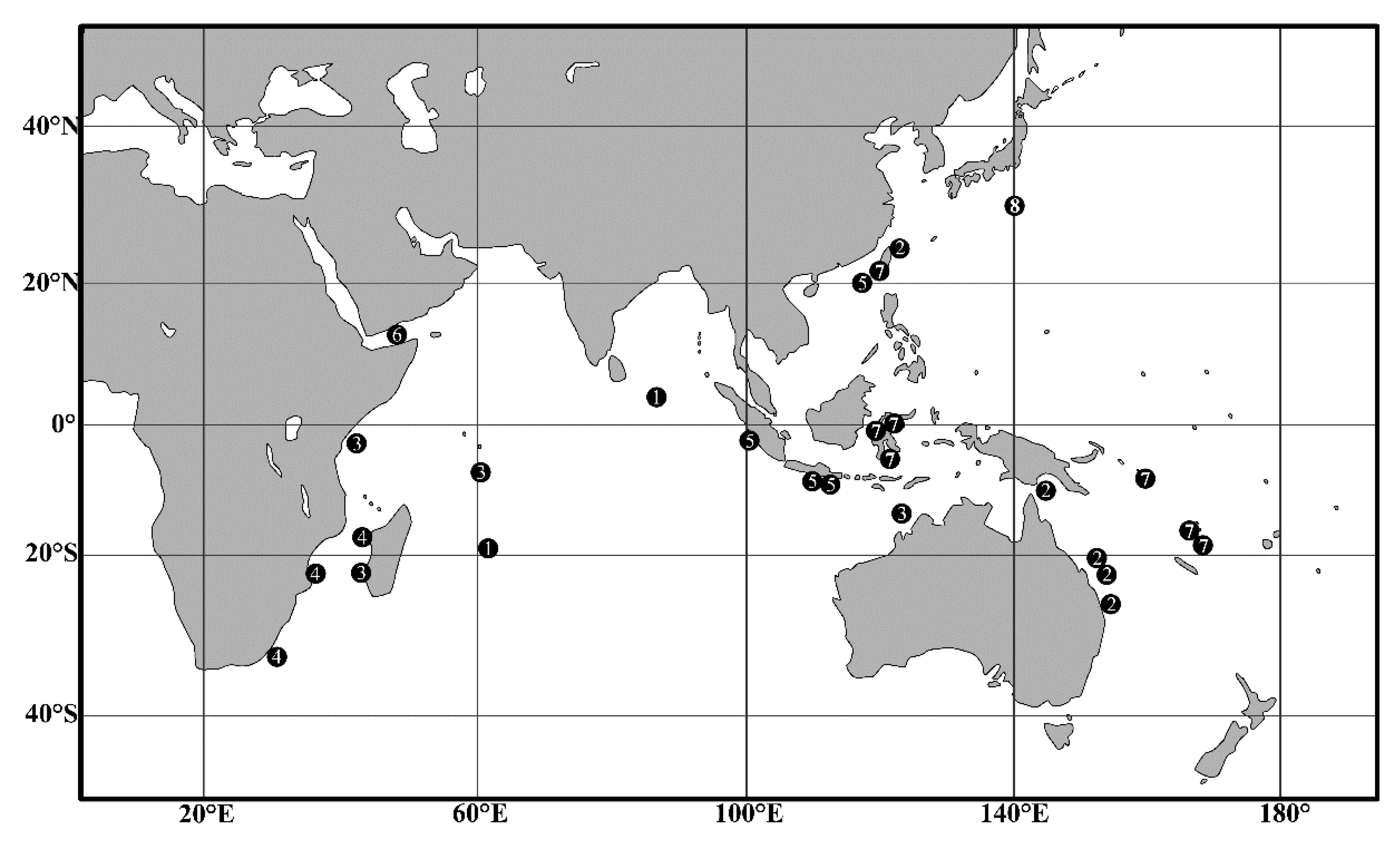
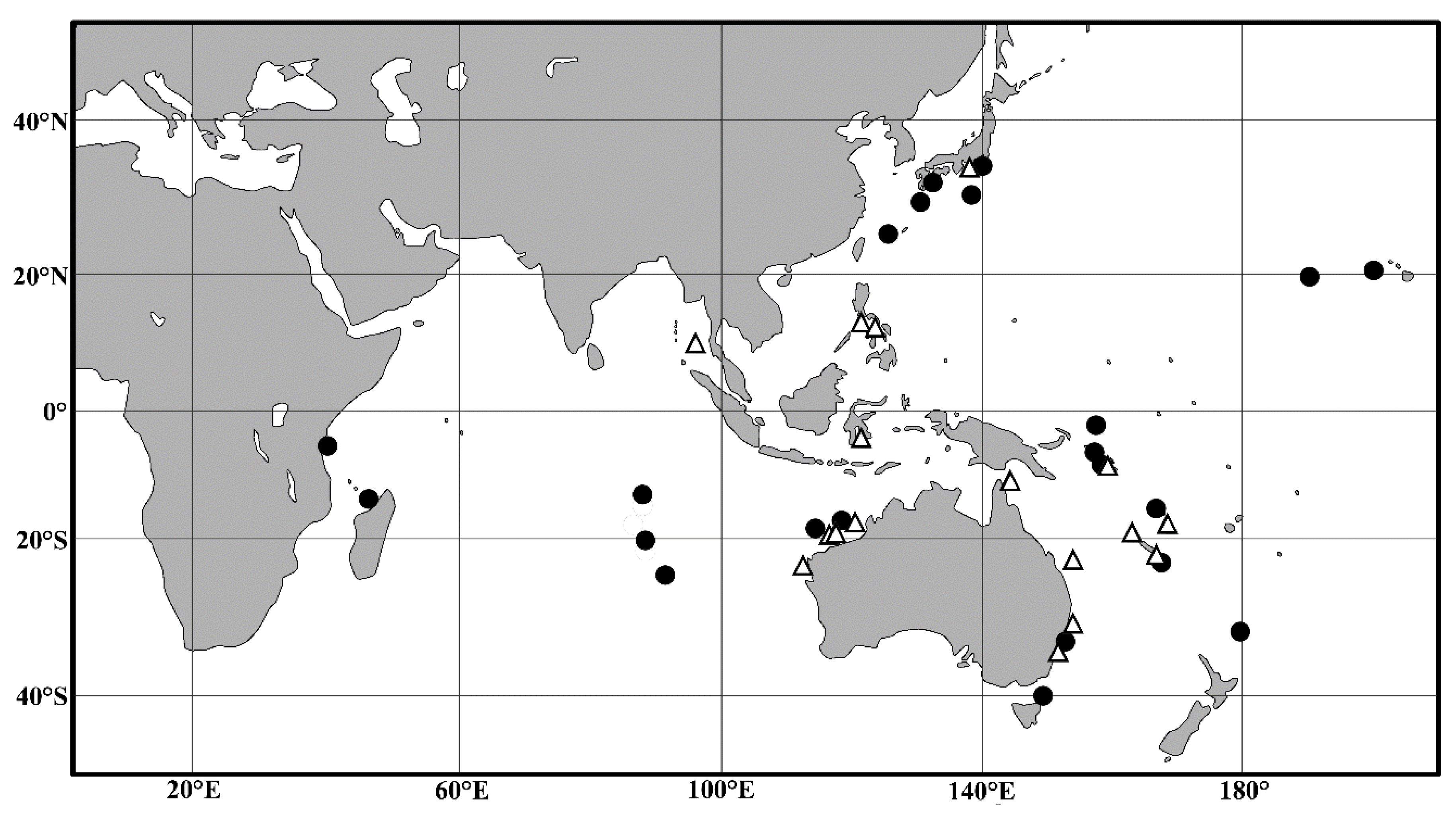
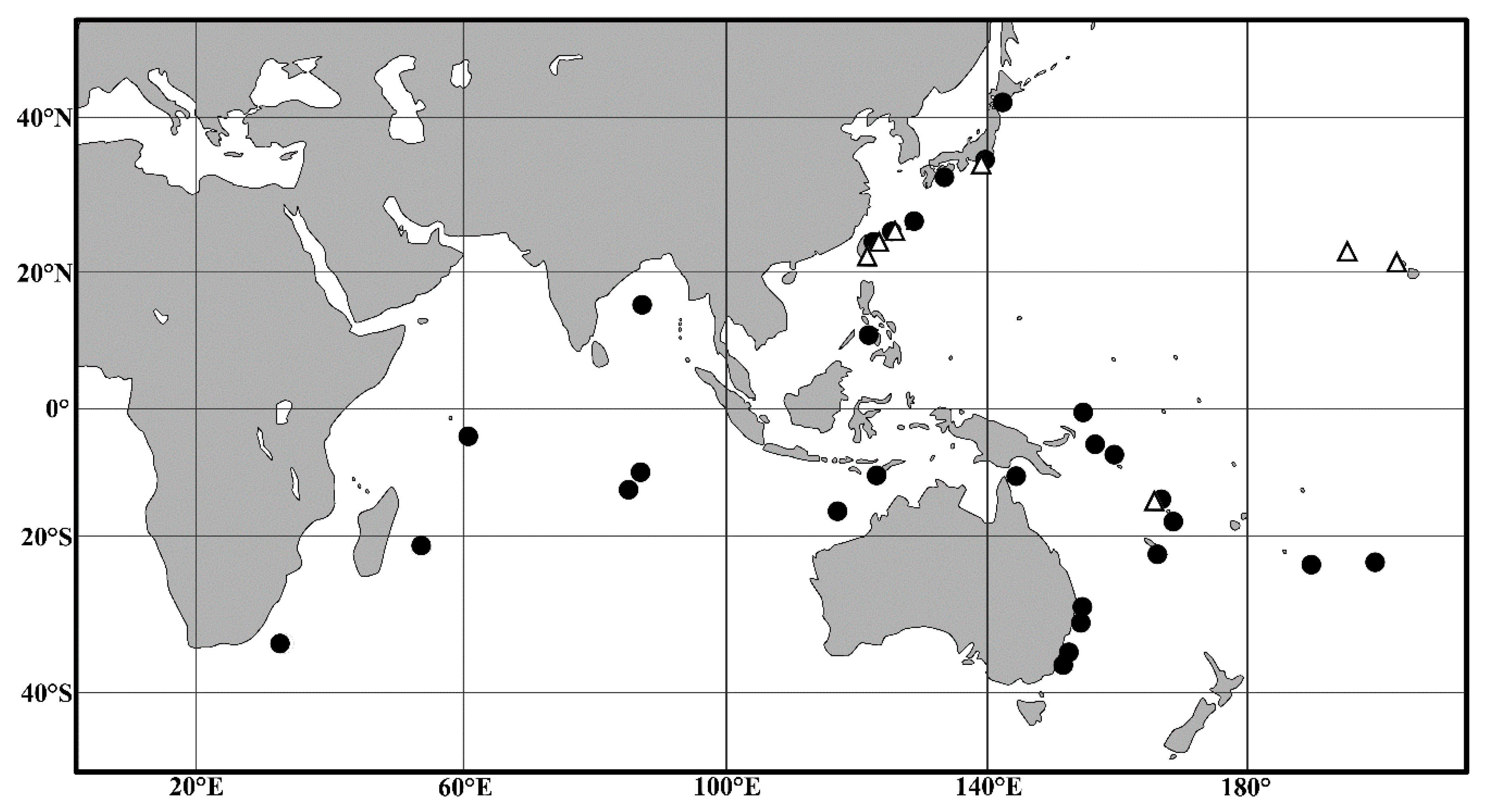
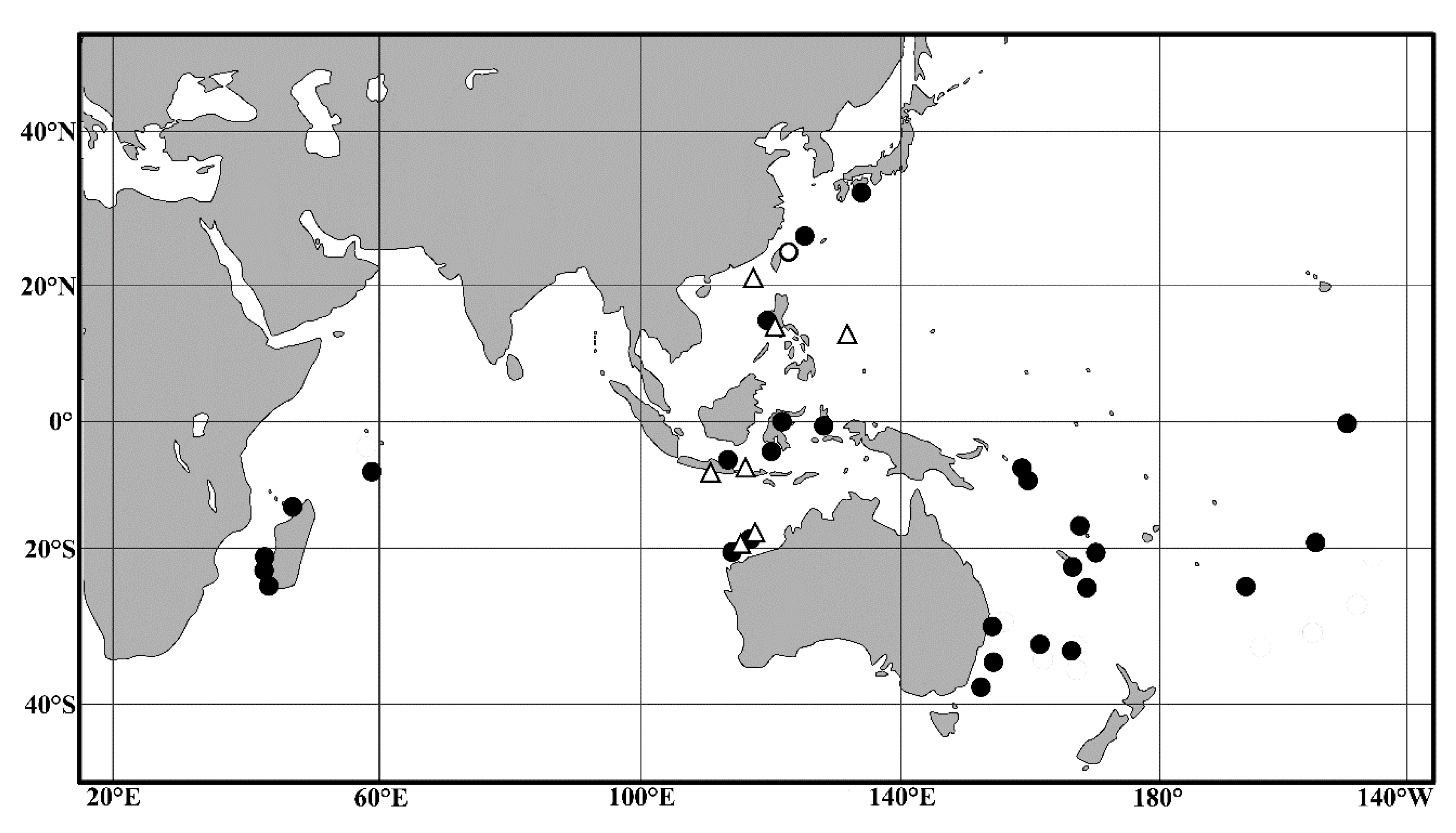
4.2. Biogeographic Distributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garman, S. The Fishes. In Reports on an Exploration off the West Coasts of Mexico, Central and South America, and off the Galapagos Islands by the U.S. Fish Commission Steamer “Albatross,” during 1891. No. XXVI. Bull. Mus. Comp. Zool. 1899, 24, 1–431. [Google Scholar]
- Bradbury, M.G. Family Ogcocephalidae Jordan 1895–batfishes. Calif. Acad. Sci. Annot. Checkl. Fishes 2003, 17, 1–17. [Google Scholar]
- Ho, H.-C.; Shao, K.-T. A New Species of Halieutopsis (Lophiiformes: Ogcocephalidae) from Western North and Eastern Central Pacific Ocean. Raffles Bull. Zool. 2007, 14, 87–92. [Google Scholar]
- Ho, H.-C.; Shao, K.-T. The Batfishes (Lophiiformes Ogcocephalidae) of Taiwan, with Descriptions of Eight New Records. J. Fish. Soc. Taiwan 2008, 35, 289–313. [Google Scholar]
- Smith, H.M.; Radcliffe, L. New Pediculate Fishes from the Philippine Islands and Contiguous Waters. Scientific Results of the Philippine Cruise of the Fisheries Steamer “Albatross,” 1907–1910. No. 16. Proc. U. S. Natl. Mus. 1902, 42, 199–214. [Google Scholar]
- Bradbury, M.G. The genera of batfishes. Copeia 1967, 2, 399–422. [Google Scholar] [CrossRef]
- Alcock, A.W. Class Pisces. In II.—Natural History Notes from H. M. Indian Marine Survey Steamer ‘Investigator’, Commander R. F. Hoskyn, R.N., Commanding.—Series II., No. 1. On the Results of Deep-Sea Dredging during the Season 1890–1891. Ann. Mag. Nat. Hist. Ser. 1891, 8, 16–34. [Google Scholar]
- Bradbury, M.G. Rare Fishes of the Deep-Sea Genus Halieutopsis: A Review with Descriptions of Four New Species (Lophiiformes: Ogcocephalidae). Fieldiana. Zool. 1988, 44, 1–22. [Google Scholar]
- Bradbury, M.G. A Review of the Fish Genus Dibranchus with Descriptions of New Species and a New Genus Solocisquama (Lophiiformes, Ogcocephalidae). Proc. Calif. Acad. Sci. 1999, 51, 259–310. [Google Scholar]
- Lloyd, R.E. A Description of the Deep-Sea Fish Caught by the R. I. M. S. Ship ‘Investigator’ since the Year 1900, with Supposed Evidence of Mutation in Malthopsis. Mem. Indian Mus. 1909, 2, 139–180. [Google Scholar]
- Ride, W.D.J.L.; Cogger, H.G.; Dupuis, C.; Kraus, O.; Minelli, A.; Thompson, F.C.; Tubbs, P.K. International Code of Zoological Nomenclature; International Trust for Zoological Nomenclature: London, UK, 1999; Available online: http://www.nhm.ac.uk/hosted-sites/iczn/code/ (accessed on 1 October 2021).
- Brauer, A. Diagnosen Von Neuen Tiefseefischen, Welche Von Der Valdivia-Expedition Gesammelt Sind. Zool. Anz. 1902, 25, 277–298. [Google Scholar]
- Shimazaki, M.; Endo, H.; Yabe, M. Redescription of a Rare Deep-Sea Batfish, Halieutopsis bathyoreos (Lophiiformes: Ogcocephalidae). Ichthy. Res. 2004, 51, 120–125. [Google Scholar] [CrossRef]
- Yamakawa, T. Fishes of the Okinawa Trough and the Adjacent Waters. The Intensive Research of Unexploited Fishery Resources on Continental Slopes; Okamura, O., Kitajima, T., Eds.; Japan Fisheries Resource Conservation Association: Tokyo, Japan, 1984; Volume I, pp. 1–414. [Google Scholar]
- Iwamoto, T.; Cailliet, G.M.; Cohen, D.M.; Pietsch, T.W.; Tucker, T.; Larson, R.J.; Martin, M.L.; Margaret, G. Bradbury (1927–2010). Copeia 2011, 4, 599–605. [Google Scholar] [CrossRef]
- Bradbury, M.G. Batfishes. In Species Identification Guide for Fisheries Purposes. The Living Marine Resources of the Western Central Pacific. Batoid Fishes, Chimeras and Bony Fishes Part 1 (Elopidae to Linophrynidae); Carpenter, K.E., Niem, V.H., Eds.; FAO: Rome, Italy, 1999; pp. 1398–2068. [Google Scholar]
- Bradbury, M.G.; Randall, J.E.; Lim, K.K.P. A Checklist of the Fishes of the South China Sea. Raffles Bull. Zool. 2002, 8, 569–667. [Google Scholar]
- Ho, H.-C.; Roberts, C.D.; Shao, K.-T. Revision of Batfishes (Lophiiformes: Ogcocephalidae) of New Zealand and Adjacent Waters, with Description of Two New Species of the Genus Malthopsis. Zootaxa 2013, 3626, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.-C. 115 Family Ogcocephalidae. In The Fishes of New Zealand; Roberts, C.D., Stewart, A.L., Struthers, C.D., Eds.; Te Papa Press: Wellington, New Zealand, 2015; pp. 896–902. [Google Scholar]
- Norman, J.R. Fishes. The John Murray Expedition 1933–1934. Sci. Rep. John Murray Exped. 1939, 7, 1–116. [Google Scholar]
- Brauer, A. Die Tiefsee-Fische. I. Systematischer Teil. In Wissenschaftliche Ergebnisse Der Deutschen Tiefsee-Expedition “Valdivia” 1898–1899; Chun, C., Ed.; Verlag von Gustav Fischer: Jena, Germany, 1906; pp. 1–432. [Google Scholar]
- Ho, H.-C.; Shao, K.-T. Annotated Checklist and Type Catalog of Fish Genera and Species Described from Taiwan. Zootaxa 2011, 2957, 1–74. [Google Scholar] [CrossRef]
- Alcock, A. Illustrations of the Zoology of H.M. Indian Marine Surveying Steamer Investigator, under the command of Commander T. H. Heming, R. N., Fishes,—Part V; Alcock, A., Anderson, A.R.S., Eds.; Office of the Superintendent of Government Printing: Calcutta, India, 1898; plates XVIII–XXIV. [Google Scholar]
- Alcock, A. A Descriptive Catalogue of the Indian Deep-Sea Fishes in the Indian Museum: Being a Revised Account of the Deep-Sea Fishes Collected by the Royal Indian Marine Survey Ship Investigator; Trustees of the Indian Museum: Calcutta, India, 1899; p. 1211. [Google Scholar]
- Weber, M. Die Fische der Siboga-Expedition; E.J. Brill, Ltd.: Leiden, The Netherlands, 1913; pp. 1–710. [Google Scholar]
- Weber, M.; de Beaufort, L.F. The Fishes of the Indo-Australian Archipelago. XI. Scleroparei, Hypostomides, Pediculati, Plectognathi, Opisthomi, Discocephali, Xenopterygii; E.J. Brill, Ltd.: Leiden, The Netherlands; pp. 1–481.
- Shao, K.-T.; Ho, H.-C.; Lin, P.-L.; Lee, P.-F.; Lee, M.-Y.; Tsai, C.-Y.; Liao, Y.-C.; Lin, Y.-C. A Checklist of the Fishes of Southern Taiwan, Northern South China Sea. Raffles Bull. Zool. 2008, 19, 233–271. [Google Scholar]
- Okamura, O.; Machida, Y.; Mochizuki, K.; Yamakawa, T. First Record of the Deep-Sea Batfish Halieutopsis stellifera from Japan. Rep. USA March Biol. Ins. Kochi Univ. 1987, 9, 201–205. [Google Scholar]
- Bradbury, M.G. A Revision of the Fish Genus Ogcocephalus with Descriptions of New Species from the Western Atlantic Ocean (Ogcocephalidae; Lophiiformes). Proc. Calif. Acad. Sci. 1980, 42, 229–285. [Google Scholar]
- Springer, V.G. Pacific Plate Biogeography, with Special Reference to Shorefishes. Smithson. Contrib. Zool. 1982, 367, 1–182. [Google Scholar] [CrossRef][Green Version]
- Peters, W.C.H. Uber Eine Neue, Mit Halieutaea Verwandte Fischgattung, Dibranchus, Aus Dem Atlantischen Ocean. Ber. Akad. Wiss. Berlin 1876, 736–742. [Google Scholar]






































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
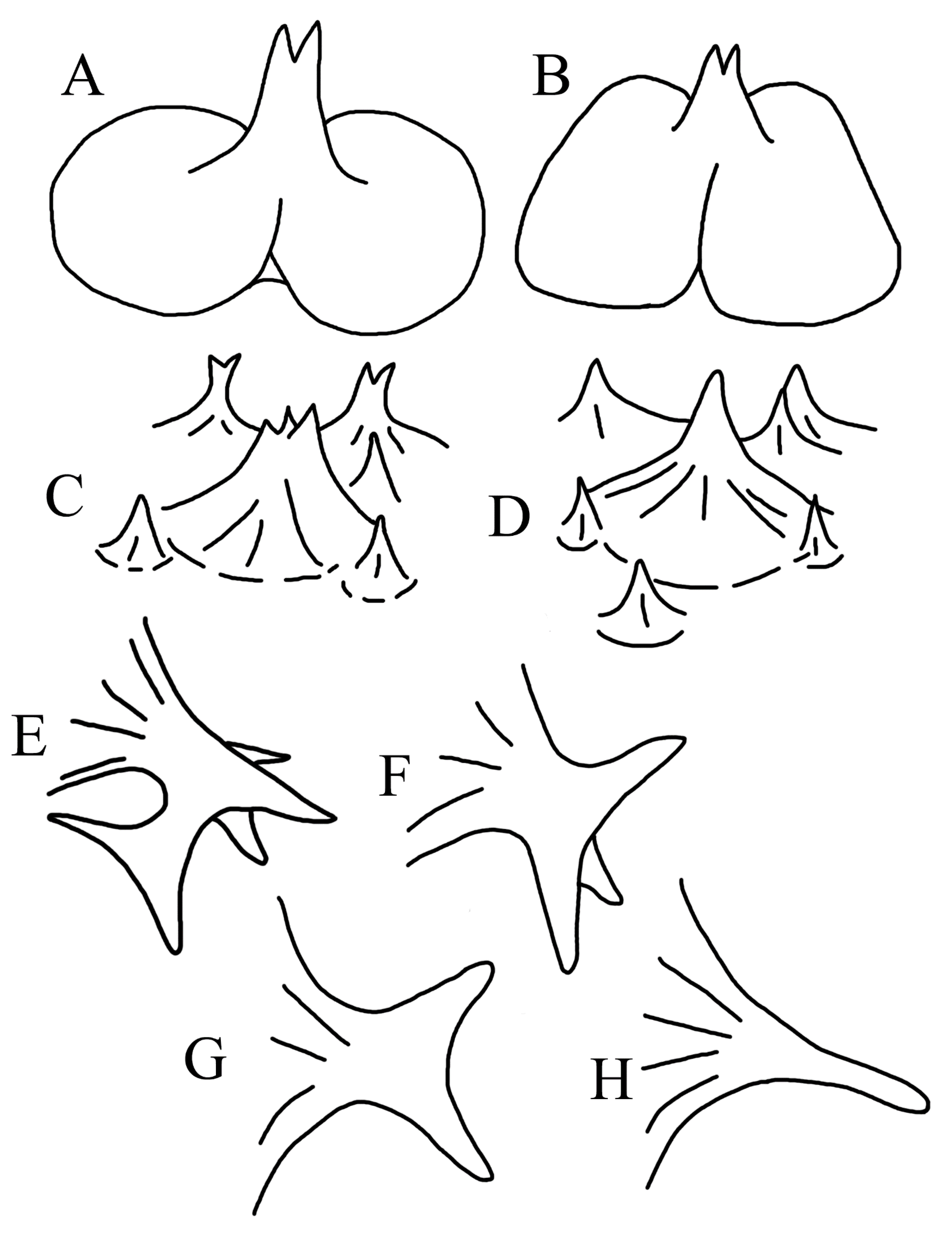{kind=link}
{kind=link}
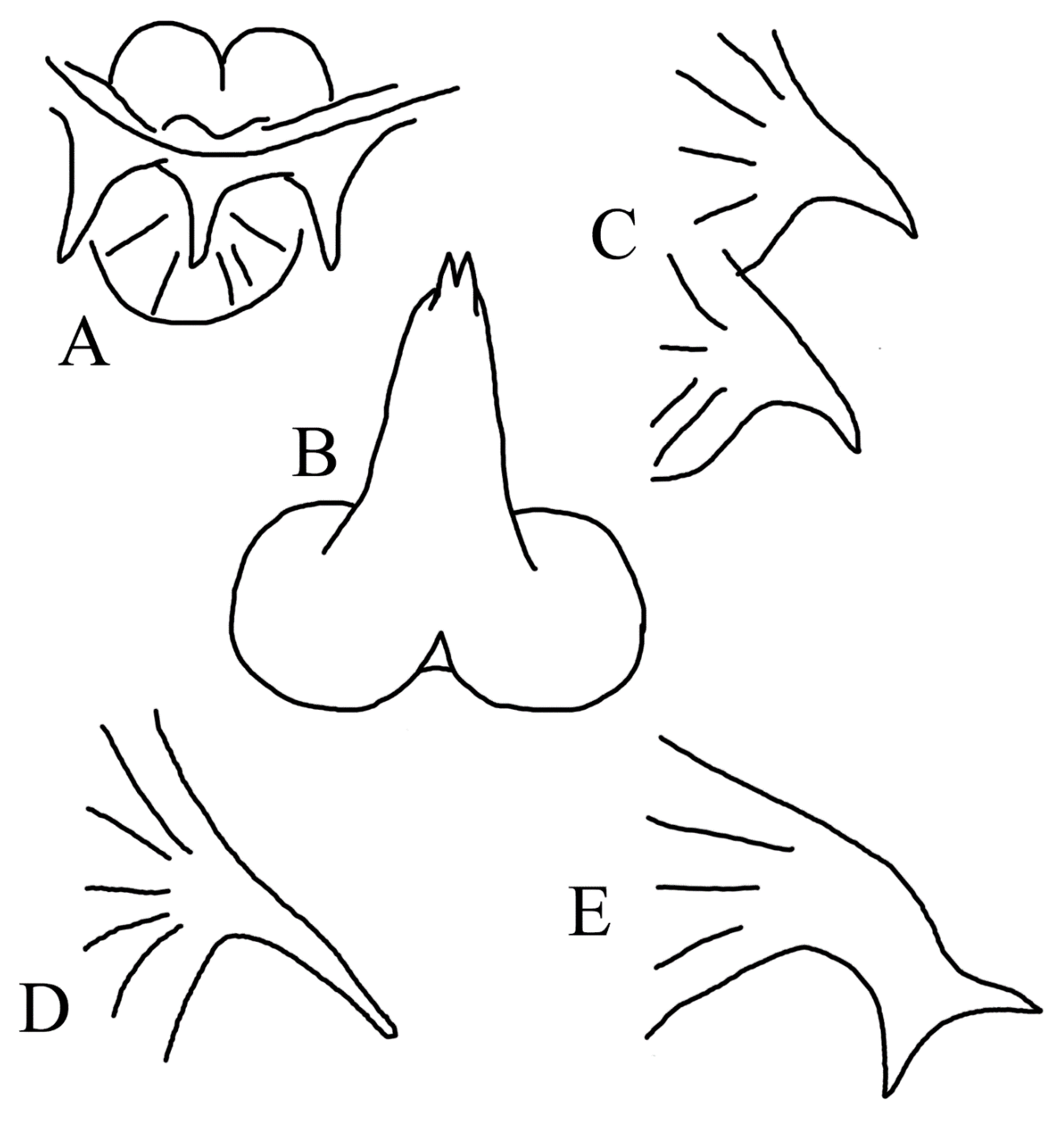{kind=link}
| Dorsal-Fin Rays | Pectoral-Fin Rays | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | 4 | 5 | 6 | 7 | 13 | 14 | 15 | 16 | 17 | ||
| H. andriashevi | 2 | 1* | 1 | 2 | 2* | ||||||
| H. bathyoreos | 33 | 3 | 26 | 4* | 4 | 12 | 46* | 2* | |||
| H. echinoderma sp. nov. | 7 | 1 | 6* | 12* | 2 | ||||||
| H. galatea | 5 | 1* | 4 | 2* | 4 | 4 | |||||
| H. ingerorum | 3 | 1 | 1* | 1 | 2* | 4 | |||||
| H. kawaii sp. nov. | 5 | 1* | 4 | 3 | 5 | 0 | 2* | ||||
| H. margaretae | 7 | 2 | 4 | 1 | 9 | 5 | |||||
| H. murrayi sp. nov. | 1 | 1* | 2* | ||||||||
| H. nasuta | 27 | 5 | 17 | 4 | 1 | 2 | 10 | 38 | 4 | ||
| H. nudiventer | 20 | 4 | 15 | 1 | 5 | 12 | 7 | 4 | |||
| H. oblonga | 8 | 1 | 6 | 1 | 1 | 8 | 5 | 2 | |||
| H. okamurai sp. nov. | 1 | 1* | 2* | ||||||||
| H. simula | 5 | 2 | 3* | 2 | 4* | 4 | |||||
| H. stellifera | 33 | 8 | 23* | 2 | 34* | 30 | 2 | ||||
| H. taiwanea sp. nov. | 3 | 1 | 2* | 4 | 2* | ||||||
| H. tumifrons | 2 | 1 | 1 | 2 | 2 | ||||||
| Suprorbital | Preopercular | Subopercular | |||||||||||||||||||||
| n | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 2 | 3 | 4 | 5 | 5 | 6 | 7 | |||||||||
| H. andriashevi | 2 | 1* | 1* | 0 | 2 | 3* | 1 | 4* | |||||||||||||||
| H. bathyoreos | 33 | 4* | 28 | 1 | 23* | 10 | 30* | 3 | |||||||||||||||
| H. echinoderma sp. nov. | 7 | 1 | 1 | 4* | 8 | 14* | 1 | 11* | 2 | ||||||||||||||
| H. galatea | 5 | 3* | 7* | 10* | 10* | ||||||||||||||||||
| H. ingerorum | 3 | 4* | 2 | 6* | 6* | ||||||||||||||||||
| H. kawaii sp. nov. | 5 | 6 | 4* | 10* | 6 | 4* | |||||||||||||||||
| H. margaretae | 7 | 2 | 10* | 2 | 2 | 12* | 2 | 12* | |||||||||||||||
| H. murrayi sp. nov. | 1 | 2* | 2* | 2* | |||||||||||||||||||
| H. nasuta | 27 | 2 | 25 | 27 | 6 | 46 | 2 | 18 | 34 | 2 | |||||||||||||
| H. nudiventer | 20 | 4 | 18 | 14 | 4 | 34 | 6 | 36 | 4 | ||||||||||||||
| H. oblonga | 8 | 3 | 10 | 2 | 1 | 3 | 9 | 6 | 8 | 8 | |||||||||||||
| H. okamurai sp. nov. | 1 | 2* | 2* | 2* | |||||||||||||||||||
| H. simula | 5 | 2 | 2 | 6* | 4* | 3 | 3 | 8* | 2 | ||||||||||||||
| H. stellifera | 33 | 2 | 2 | 62* | 3* | 39* | 24 | 4* | 62 | ||||||||||||||
| H. taiwanea sp. nov. | 3 | 6* | 6* | 6 | |||||||||||||||||||
| H. tumifrons | 2 | 1 | 3 | 4 | 2 | 2 | |||||||||||||||||
| DBS | Ventral | Tail | |||||||||||||||||||||
| n | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |||
| H. andriashevi | 2 | 4* | 2 | 0 | 1* | 1* | 1 | 1 | 1* | 1* | |||||||||||||
| H. bathyoreos | 33 | 2 | 31* | 2 | 21 | 10* | 18 | 12 | 2 | 1* | 1* | ||||||||||||
| H. echinoderma sp. nov. | 7 | 14* | 9* | 2* | 3 | 1 | 5 | 7* | 1* | ||||||||||||||
| H. galatea | 5 | 10* | 1 | 6* | 3* | 4 | 2 | 2 | 0 | 0 | 1* | 1* | |||||||||||
| H. ingerorum | 3 | 2 | 4* | 5* | 1* | 2 | 2 | 0 | 2* | ||||||||||||||
| H. kawaii sp. nov. | 6 | 8* | 2 | 6* | 2 | 4 | 5* | 1* | |||||||||||||||
| H. margaretae | 7 | 6 | 8 | 3* | 11* | 6 | 8* | ||||||||||||||||
| H. murrayi sp. nov. | 1 | 2* | 2* | 2* | |||||||||||||||||||
| H. nasuta | 27 | 4 | 50 | 3 | 26 | 25 | 7 | 20 | 23 | 2 | 1 | 1 | |||||||||||
| H. nudiventer | 20 | 40 | 36 | 4 | 9 | 23 | 4 | 4 | |||||||||||||||
| H. oblonga | 8 | 12 | 4 | 6 | 8 | 2 | 2 | 10 | 4 | ||||||||||||||
| H. okamurai sp. nov. | 1 | 2* | 1* | 1* | 1* | 1* | |||||||||||||||||
| H. simula | 5 | 3* | 7 | 2* | 5* | 2 | 1 | 5 | 3 | 0 | 1* | 1* | |||||||||||
| H. stellifera | 33 | 62* | 4 | 1 | 32 | 32* | 1 | 6 | 56* | 4 | 2 | ||||||||||||
| H. taiwanea sp. nov. | 3 | 6* | 1 | 5* | 4* | ||||||||||||||||||
| H. tumifrons | 2 | 1 | 1 | 2 | 4 | 1 | 1 | 2 | |||||||||||||||
| H. andriashevi | H. bathyoreos | H. galatea | ||||||
|---|---|---|---|---|---|---|---|---|
| HT * | NT | HT * | NT | HT | HT + NT | |||
| SL (mm) | 46.1 | 51.1 | 62.6 | 42.1–73.8 (n = 22) | 53.0 | 41.9-83.9 (n = 4) | ||
| % SL | Range (mean ± SD) | Range (mean ± SD) | ||||||
| Skull length | 24.9 | 27.0 | 24.9 | 24.2–29.5 (26.8 ± 1.5) | 28.1 | 28.1–32.2 (30.2 ± 1.9) | ||
| Head width | 17.3 | 22.7 | 24.0 | 21.3–29.7 (25.7 ± 2.1) | 24.3 | 24.3–30.3 (27.7 ± 2.7) | ||
| Head depth | - | 24.3 | - | 22.2–29.4 (24.8 ± 1.9) | 28.1 | 26.6–29.8 (27.9 ± 1.4) | ||
| Eye diameter | 6.1 | 8.8 | 8.5 | 6.7–10.0 (8.3 ± 1.0) | 7.0 | 8.9–12.5 (10.4 ± 1.6) | ||
| Interorbital width | 9.7 | 9.6 | 112 | 9.1–12.1 (11.0 ± 0.9) | 22.5 | 11.7–12.4 (11.9 ± 0.3) | ||
| Illicial trough width | - | 9.0 | - | 9.3–13.3 (11.0 ± 1.1) | 11.7 | 6.0–10.4 (8.3 ± 2.0) | ||
| Mouth width | 17.3 | 17.2 | 19.3 | 18.5–23.3 (21.8 ± 1.4) | 8.9 | 22.5–25.1 (23.6 ± 1.1) | ||
| Upper jaw length | 11.5 | 10.8 | 11.5 | 10.3–13.2 (12.1 ± 0.9) | 11.7 | 11.7–15.9 (14.2 ± 1.9) | ||
| Predorsal length | 57.3 | 65.2 | 56.4 | 58.0–68.3 (62.6 ± 3.1) | 70.4 | 65.0–70.4 (67.3 ± 2.2) | ||
| Pre-anus length | 52.6 | 56.6 | 52.4 | 50.4–61.8 (55.9 ± 3.7) | 57.4 | 57.4–64.5 (61.8 ± 3.0) | ||
| Preanal length | 70.1 | 72.0 | 69.7 | 65.5–74.0 (71.5 ± 2.8) | 71.1 | 71.1–76.6 (74.8 ± 2.4) | ||
| Tail length | 47.4 | 43.4 | 47.6 | 39.2–49.6 (44.1 ± 3.7) | 42.6 | 35.5–42.6 (39.2 ± 2.2) | ||
| Disk margin | 34.4 | 38.2 | 41.4 | 38.8–46.3 (42.8 ± 2.7) | 43.0 | 43.0–46.6 (44.2 ± 1.6) | ||
| Dorsal-fin length | - | 15.1 | - | 12.5–18.8 (16.3 ± 2.0) | 20.2 | 17.7–27.8 (24.6 ± 4.5) | ||
| Pectoral-fin length | 20.4 | 18.0 | 20.3 | 17.9–23.9 (21.6 ± 1.9) | 27.5 | 19.2–28.4 (22.0 ± 4.1) | ||
| Anal-fin length | - | 15.1 | - | 13.5–19.2 (16.1 ± 1.8) | 20.8 | 18.4–20.8 (19.6 ± 1.1) | ||
| Caudal-fin length | - | 24.7 | - | 23.4–30.2 (26.2 ± 2.1) | 31.5 | 25.6–31.5 (28.1 ± 2.5) | ||
| Meristics | n = 33 | n = 5 | ||||||
| Dorsal-fin rays | 4 | 5 | 6 | 4–6 ( mainly 5) | 4 | 4–5 | ||
| Pectoral-fin rays | 15 | 14 | 15;16 | 13–16 (15) | 14 | 14–16 | ||
| Anal-fin rays | 4 | 4 | 4 | 4–5 (4) | 4 | 4 | ||
| Lateral-line neuromasts | ||||||||
| SS | 3;4 | 6 | 4 | 4–6 (5) | 7;8 | 6–8 | ||
| PMS | - | 0 | - | 0 | - | 1 | ||
| CS | - | 8 | - | 8–9 (8) | 8 | 8 | ||
| BS | 0? | 9 | 8 | 8 | 9;10 | 9 | ||
| SOS | 5 | 5 | 5 | 5–6 (5) | 6 | 5–6 | ||
| POS | 2 | 3 | 2 | 2–3 (2) | 3 | 3 | ||
| DBSS | 3 | 3 | 3 | 2–3 (3) | 3 | 3 | ||
| PBSS | 3 | 2 | 3 | 3 | 3 | 3 | ||
| VS | 3 | 1 | 3 | 1–3 (2–3) | 3;4 | 3–5 | ||
| TS | 8;10 | 10;11 | 13;14 | 10–13 (10–11) | 14;15 | 8–10 | ||
| H. echinoderma sp. nov. | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Holotype | Paratypes | ||||||||
| SL (mm) | 74.0 | 69.4 | 66.1 | 60.9 | 54.7 | 54.0 | 45.0 | ||
| % SL | Range (Mean ± SD) | ||||||||
| Skull length | 31.1 | 31.3 | 31.9 | 32.7 | 32.9 | 34.1 | 31.6 | 31.1–34.1 (32.2 ± 1.1) | |
| Head width | 27.0 | 32.4 | 25.0 | 29.2 | 30.3 | 26.7 | 26.9 | 25.0–32.4 (28.2 ± 2.6) | |
| Head depth | 25.7 | 32.4 | 29.0 | 27.1 | 30.7 | 31.7 | 28.0 | 25.7–32.4 (29.2 ± 2.5) | |
| Eye diameter | 11.2 | 12.2 | 9.5 | 11.2 | 12.1 | 10.9 | 11.1 | 9.5–12.2 (11.2 ± 0.9) | |
| Interorbital width | 13.5 | 14.4 | 13.8 | 14.6 | 13.7 | 13.5 | 16.2 | 13.5–16.2 (14.3 ± 1.0) | |
| Illicial trough width | 9.5 | 9.8 | 11.0 | 11.0 | 9.5 | 11.1 | - | 9.5–11.1 (10.3 ± 0.8) | |
| Mouth width | 25.4 | 25.6 | 24.1 | 24.8 | 27.4 | 26.5 | 25.6 | 24.1–27.4 (25.6 ± 1.1) | |
| Upper jaw length | 13.4 | 15.3 | 13.8 | 15.8 | 15.5 | 14.4 | 13.8 | 13.4–15.8 (14.6 ± 1.0) | |
| Predorsal length | 68.2 | 66.7 | 69.9 | 69.3 | 71.3 | 66.3 | 70.4 | 66.3–71.3 (68.9 ± 1.9) | |
| Pre-anus length | 64.7 | 65.1 | 64.8 | 66.8 | 68.9 | 63.1 | 68.0 | 63.1–68.9 (65.9 ± 2.1) | |
| Preanal length | 78.4 | 73.8 | 80.0 | 78.0 | 78.8 | 76.3 | 79.6 | 73.8–80.0 (77.8 ± 2.2) | |
| Tail length | 35.3 | 34.9 | 35.2 | 33.2 | 31.1 | 36.9 | 32.0 | 31.1–36.9 (34.1 ± 1.9) | |
| Disk margin | 43.5 | 49.0 | 46.6 | 44.5 | 45.5 | 49.4 | 45.8 | 43.5–49.4 (46.3 ± 2.2) | |
| Dorsal-fin length | 14.9 | 15.1 | 16.5 | 15.6 | 18.5 | 17.2 | 15.3 | 14.9–18.5 (16.2 ± 1.3) | |
| Pectoral-fin length | 26.2 | 27.4 | 26.8 | 29.2 | 25.6 | 26.5 | 27.6 | 25.6–29.2 (27.0 ± 1.2) | |
| Anal-fin length | 16.2 | 17.9 | 17.4 | 18.2 | 19.6 | 21.1 | 19.6 | 16.2–21.1 (18.6 ± 1.6) | |
| Caudal-fin length | 24.7 | 28.0 | 25.7 | 24.8 | 26.1 | 26.9 | 30.0 | 24.7–30.0 (26.6 ± 1.9) | |
| Meristics | Range | ||||||||
| Dorsal-fin rays | 5 | 5 | 4 | 5 | 5 | 5 | 5 | 4–5 | |
| Pectoral-fin rays | 16 | 16 | 17 | 16 | 16 | 16 | 16 | 16–17 | |
| Anal-fin rays | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| Lateral-line neuromasts | |||||||||
| SS | 5 | 5 | 5 | 6 | 6 | 5 | 5 | 5–6 | |
| PMS | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CS | 8 | 8 | 8 | 9 | 8 | 8 | 8 | 8–9 | |
| BS | 8 | 8 | 8 | 9 | 8 | 8 | 8 | 8–9 | |
| SOS | 6 | 6 | 5;6 | 6 | 6 | 6;7 | 6;7 | 5–7 | |
| POS | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |
| DBSS | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |
| PBSS | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |
| VS | 4;5 | 5 | 3 | 3;4 | 3 | 3 | 3 | 3–5 | |
| TS | 9 | 9;10 | 10 | 10;11 | 8;9 | 9;10 | 10 | 8–10 | |
| H. ingeroum | H. taiwanea sp. nov. | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Holotype* | Nontypes | Holotype | Paratypes | ||||||
| SL (mm) | 44.2 | 78.8 | 66.3 | 78.3 | 70.0 | 67.5 | |||
| % SL | Mean ± SD | Mean ± SD | |||||||
| Skull length | 27.6 | 27.2 | 26.8 | 27.2 ± 0.4 | 22.7 | 22.6 | 22.8 | 22.7 ± 0.1 | |
| Head depth | - | 21.3 | 19.3 | 20.3 ± 1.4 | 20.3 | 18.4 | 17.0 | 18.6 ± 1.6 | |
| Head width | 21.0 | 26.4 | 25.5 | 24.3 ± 2.9 | 22.5 | 23.7 | 22.8 | 23.0 ± 0.6 | |
| Eye diameter | 8.6 | 5.7 | 6.2 | 6.8 ± 1.5 | 5.2 | 6.1 | 4.9 | 5.4 ± 0.6 | |
| Interorbital width | 12.7 | 11.3 | 11.5 | 11.8 ± 0.7 | 11.0 | 10.3 | 8.7 | 10.0 ± 1.1 | |
| Illicial trough width | - | 9.3 | 9.2 | 9.2 ± 0.0 | 8.8 | 9.6 | 8.4 | 8.9 ± 0.6 | |
| Mouth width | 18.6 | 17.5 | 18.4 | 18.2 ± 0.6 | 13.7 | 15.3 | 13.5 | 14.1 ± 1.0 | |
| Upper jaw length | 12.4 | 10.8 | 12.2 | 11.8 ± 0.9 | 10.0 | 10.6 | 9.3 | 10.0 ± 0.6 | |
| Predorsal length | 61.5 | 63.7 | 62.0 | 62.4 ± 1.1 | 57.3 | 55.9 | 58.4 | 57.2 ± 1.3 | |
| Pre-anus length | 56.1 | 58.1 | 57.6 | 57.3 ± 1.0 | 55.4 | 49.9 | 51.1 | 52.1 ± 2.9 | |
| Preanal length | 72.2 | 72.0 | 66.8 | 70.3 ± 3.0 | 67.3 | 66.6 | 69.5 | 67.8 ± 1.5 | |
| Tail length | 43.9 | 41.9 | 42.4 | 42.7 ± 1.0 | 44.6 | 50.1 | 48.9 | 47.9 ± 2.9 | |
| Disk margin | 43.9 | 44.0 | 43.4 | 43.8 ± 0.3 | 38.1 | 35.7 | 35.6 | 36.4 ± 1.4 | |
| Dorsal-fin length | - | 14.3 | 12.5 | 13.4 ± 1.3 | 14.8 | 14.7 | 11.4 | 13.6 ± 1.9 | |
| Pectoral-fin length | - | 17.9 | 20.1 | 27.0 ± 13.9 | 17.9 | 18.7 | 18.4 | 18.3 ± 0.4 | |
| Anal-fin length | - | 13.7 | 15.7 | 14.7 ± 1.4 | 13.0 | - | 14.4 | 13.7 ± 1.0 | |
| Caudal-fin length | - | 21.8 | 24.9 | 23.4 ± 2.2 | 19.7 | 24.1 | 22.1 | 22.0 ± 2.2 | |
| Meristics | Range | Range | |||||||
| Dorsal-fin rays | 5 | 4 | 6 | 4–6 | 5 | 4 | 5 | 4–5 | |
| Pectoral-fin rays | 14 | 15 | 15 | 14–15 | 16 | 15 | 15 | 15–16 | |
| Anal-fin rays | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| Lateral-line neuromasts | |||||||||
| SS | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |
| PMS | - | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| CS | - | 8 | 8 | 8 | 8 | 8 | 8 | 8 | |
| BS | 9;10 | 9 | 9 | 9–10 | 9 | 9 | 9 | 9 | |
| SOS | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | |
| POS | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| DBSS | 2 | 3 | 3 | 2–3 | 3 | 3 | 3 | 3 | |
| PBSS | - | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |
| VS | 2;3 | 2 | 2 | 2–3 | 2;2 | 2;2 | 1;2 | 1–2 | |
| TS | 13 | 10;11 | 10 | 10–13 | 14 | 14 | - | 14 | |
| H.kawaii sp. nov. | |||||||
|---|---|---|---|---|---|---|---|
| Holotype | Paratypes | ||||||
| SL (mm) | 79.4 | 83.6 | 57.8 | 56.8 | 37.2 | ||
| % SL | Range (Mean ± SD) | ||||||
| Skull length | 34.0 | 32.1 | 29.9 | 30.5 | 32.5 | 29.9–34.0 (31.8 ± 1.6) | |
| Head width | 25.8 | 25.8 | 24.0 | 27.3 | 27.7 | 24.0–27.7 (26.1 ± 1.4) | |
| Head depth | 25.8 | 27.2 | 20.9 | 23.4 | 27.7 | 20.9–27.7 (25.0 ± 2.8) | |
| Eye diameter | 13.2 | 12.0 | 11.6 | 12.5 | 14.0 | 11.6–14.0 (12.7 ± 1.0) | |
| Interorbital width | 11.0 | 11.0 | 9.9 | 10.4 | 11.3 | 9.9–11.3 (10.7 ± 0.6) | |
| Illicial trough width | 10.5 | 9.3 | 8.5 | 10.2 | 8.3 | 8.3–10.5 (9.4 ± 1.0) | |
| Iiilcial trough length | 8.1 | 8.1 | 6.1 | 7.0 | 8.3 | 6.1–8.3 (7.5 ± 1.0) | |
| Mouth width | 23.3 | 23.2 | 20.8 | 23.4 | 26.3 | 20.8–26.3 (23.4 ± 2.0) | |
| Upper jaw length | 14.1 | 14.0 | 11.9 | 12.9 | 13.4 | 11.9–14.1 (13.3 ± 0.9) | |
| Predorsal length | 66.4 | 67.1 | 63.3 | 63.2 | 66.9 | 63.2–67.1 (65.4 ± 2.0) | |
| Pre-anus length | 65.6 | 62.6 | 57.6 | 59.0 | 58.6 | 57.6–65.6 (60.7 ± 3.3) | |
| Preanal length | 75.6 | 76.3 | 72.7 | 73.9 | 73.1 | 72.7–76.3 (74.3 ± 1.6) | |
| Tail length | 34.4 | 37.4 | 42.4 | 41.0 | 41.4 | 34.4–42.4 (39.3 ± 3.3) | |
| Disk margin | 41.7 | 42.1 | 41.9 | 44.7 | 43.5 | 41.7–44.7 (42.8 ± 1.3) | |
| Dorsal-fin length | 22.4 | 17.5 | 15.4 | 16.7 | 21.8 | 15.4–22.4 (18.8 ± 3.1) | |
| Pectoral-fin length | 29.1 | 28.8 | 27.2 | 27.3 | 32.3 | 27.2–32.3 (28.9 ± 2.1) | |
| Anal-fin length | 22.7 | 18.7 | 17.3 | 18.7 | 21.5 | 17.3–22.7 (19.8 ± 2.2) | |
| Caudal-fin length | 29.8 | 27.8 | 27.7 | 29.0 | 30.9 | 27.7–30.9 (29.0 ± 1.4) | |
| Meristics | Range | ||||||
| Dorsal-fin rays | 4 | 5 | 5 | 5 | 5 | 4–5 | |
| Pectoral-fin rays | 16 | 13;14 | 14 | 14 | 13 | 13–16 | |
| Anal-fin rays | 4 | 4 | 4 | 4 | 4 | 4 | |
| Lateral-line neuromasts | |||||||
| SS | 7 | 6 | 7 | 6 | 6 | 6–7 | |
| PMS | 1 | 1 | 1 | 1 | 1 | 1 | |
| CS | 8 | 8 | 8 | 8 | 8 | 8 | |
| BS | 9 | 9 | 9 | 9 | 9 | 9 | |
| SOS | 6 | 5 | 5 | 5 | 6 | 5–6 | |
| POS | 3 | 3 | 3 | 3 | 3 | 3 | |
| DBSS | 3 | 3 | 3 | 3 | 3 | 3 | |
| PBSS | 3 | 3 | 3 | 3 | 3 | 3 | |
| VS | 3;3 | 4;4 | 3;2 | 3;3 | 2;3 | 2–4 | |
| TS | 12;11 | 11;11 | 11;11 | 10;10 | 10;10 | 10–12 | |
| H. margaretae | H. murrayi sp. nov. | H. nasuta | |||||
|---|---|---|---|---|---|---|---|
| Types* | Nontypes | Holotype | Nontypes | ||||
| SL (mm) | 38.0–66.0 | 42.2 | 44.2 | 48.7 | 20.5–71.6 (n = 21) | ||
| in %SL | Range (n = 5) | Range (mean ± SD) | |||||
| Skull length | 28.7–31.6 | 29.6 | 28.3 | 33.5 | 28.7–36.3 (32.5 ± 1.7) | ||
| Head width | 21.3–24.2 | - | 23.8 | 26.9 | 24.6–30.8 (26.8 ± 1.7) | ||
| Head depth | - | - | 23.3 | 24.4 | 25.7–32.3 (28.7 ± 1.6) | ||
| Eye diameter | 7.9–11.7 | 10.0 | 10.0 | 11.1 | 9.7–12.9 (11.3 ± 0.9) | ||
| Interorbital width | 8.3–10.5 | 9.7 | 8.6 | 8.2 | 10.3–13.3 (12.2 ± 0.8) | ||
| Illicial trough width | - | 6.6 | 7.2 | 7.0 | 8.8–12.9 (10.0 ± 0.9) | ||
| Mouth width | 20.9–23.3 | 23.0 | 22.9 | 21.4 | 22.2–28.6 (25.9 ± 1.7) | ||
| Upper jaw length | 10.8–13.2 | 14.9 | 12.7 | 12.5 | 13.4–17.5 (16.1 ± 1.1) | ||
| Predorsal length | 67.2–76.6 | 65.9 | 64.5 | 64.3 | 64.1–71.0 (66.7 ± 1.8) | ||
| Pre-anus length | 53.0–60.5 | 56.4 | 55.0 | 56.3 | 61.2–66.9 (63.9 ± 1.8) | ||
| Preanal length | 69.7–76.3 | 73.9 | 72.2 | 72.5 | 74.9–80.3 (77.1 ± 1.5) | ||
| Tail length | 39.5–47.0 | 43.6 | 45.0 | 43.7 | 33.1–38.8 (36.1 ± 1.8) | ||
| Disk margin | 38.3–42.1 | 42.7 | 40.0 | 42.5 | 43.4–48.4 (45.6 ± 1.2) | ||
| Dorsal-fin length | - | 17.1 | 17.6 | 19.1 | 16.1–21.0 (18.3 ± 1.4) | ||
| Pectoral-fin length | - | 24.6 | 23.1 | 31.6 | 25.8–31.5 (28.9 ± 1.6) | ||
| Anal-fin length | - | 19.4 | 18.8 | 19.7 | 14.8–20.6 (17.9 ± 1.5) | ||
| Caudal-fin length | - | 30.8 | 27.8 | 31.0 | 24.3–30.9 (27.4 ± 1.8) | ||
| Meristics | n = 27 | ||||||
| Dorsal-fin rays | 4–6 | 5 | 5 | 5 | 4–5 (mainly 5) | ||
| Pectoral-fin rays | 14–15 | 15 | 15 | 14;14 | 14–16 (15) | ||
| Anal-fin rays | 4 | 4 | 4 | 4 | 4 | ||
| Lateral-line neuromasts | |||||||
| SS | 6 | 6 | 6 | 7 | 6–8 (7) | ||
| PMS | 1 | 1 | 1 | 1 | 2 | ||
| CS | 8 | 8 | 8 | 8 | 7–8 | ||
| BS | - | 11 | 10 | 9 | 8 | ||
| SOS | 6 | 5 | 6 | 6;7 | 5–7 (6) | ||
| POS | 3 | 3 | 2 | 3 | 3–4 (3) | ||
| DBSS | 2–3 | 3 | 3 | 3 | 3 | ||
| PBSS | 3 | 3 | 3 | 3 | 3 | ||
| VS | 1–2 | 2;2 | 2;2 | 2;3 | 2–4 (3–4) | ||
| TS | 8–9 | 8;8 | 8;9 | 9 | 8–10 | ||
| H. nudiventer | H. oblonga | H. okamurai sp. nov. | |||
|---|---|---|---|---|---|
| Non-Types | Non-Types | Holotype | |||
| SL (mm) | 38.3–77.5 (n = 28) | 32.1–62.9 (n = 8) | 65.0 | ||
| in %SL | Range (mean ± SD) | Range (mean ± SD) | |||
| Skull length | 22.8-30.7 (28.5 ± 2.4) | 30.2–36.1 (33.6 ± 2.2) | 27.4 | ||
| Head width | 23.3–30.7 (27.7 ± 2.3) | 26.3–30.5 (28.5 ± 1.8) | 24.5 | ||
| Head depth | 21.6–28.6 (24.6 ± 2.1) | 27.0–34.1 (30.7 ± 3.1) | 25.8 | ||
| Eye diameter | 7.2–11.3 (9.6 ± 1.2) | 11.4–13.3 (12.3 ± 0.7) | 7.1 | ||
| Interorbital width | 8.5–11.8 (9.7 ± 0.9) | 11.4–13.8 (12.7 ± 0.9) | 12.5 | ||
| Illicial trough width | 7.5–11.1 (9.2 ± 0.8) | 6.7–10.3 (8.8 ± 1.7) | 8.8 | ||
| Mouth width | 17.5–29.1 (24.1 ± 2.3) | 21.5–29.3 (25.4 ± 3.3) | 19.4 | ||
| Upper jaw length | 10.8–16.5 (14.0 ± 1.9) | 14.6–18.7 (16.3 ± 1.9) | 10.6 | ||
| Predorsal length | 61.1–67.1 (64.2 ± 1.9) | 59.5–72.0 (66.1 ± 4.9) | 62.2 | ||
| Pre-anus length | 54.0–62.6 (58.7 ± 2.5) | 56.8–64.8 (59.3 ± 3.4) | 51.7 | ||
| Preanal length | 71.3–76.5 (73.6 ± 1.5) | 73.3–79.3 (75.9 ± 2.9) | 70.0 | ||
| Tail length | 37.4–46.0 (35.8 ± 2.5) | 35.2–43.2 (40.7 ± 3.4) | 48.3 | ||
| Disk margin | 40.9–49.9 (44.6 ± 2.5) | 42.0–57.4 (47.8 ± 6.0) | 41.7 | ||
| Dorsal-fin length | 15.1–18.6 (17.1 ± 1.2) | 19.6–26.9 (22.6 ± 3.2) | 14.6 | ||
| Pectoral-fin length | 18.9–25.8 (22.0 ± 2.2) | 26.8–37.2 (31.2 ± 4.1) | 20.0 | ||
| Anal-fin length | 14.6–19.8 (16.9 ± 1.7) | 20.3–25.7 (22.9 ± 2.1) | 14.2 | ||
| Caudal-fin length | 22.8–31.5 (26.2 ± 3.1) | 26.7–37.5 (32.1 ± 5.4) | 17.1 | ||
| Meristics | n = 20 | ||||
| Dorsal-fin rays | 4–6 (mainly 5) | 5 | 5 | ||
| Pectoral-fin rays | 13–16 (15–16) | 14–17 | 15;15 | ||
| Anal-fin rays | 4 | 4 | 4 | ||
| Lateral-line neuromasts | |||||
| SS | 3–6 | 6 | 6 | ||
| PMS | 0–1 | 0 | 1 | ||
| CS | 8 | 8 | 8 | ||
| BS | 8–9 | 8–9 | 9 | ||
| SOS | 5–6 (5) | 5–6 | 5 | ||
| POS | 2–3 (2) | 2–4 | 3 | ||
| DBSS | 3 | 3–4 | 3 | ||
| PBSS | 3 | 3 | 3 | ||
| VS | 1–2 (1) | 3–5 | 1;2 | ||
| TS | 8–11 (8–9) | 8–10 (9) | 10;11 | ||
| H. simula | H. stellifera | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| HT + NT* | NT | HT + NT* | NT | |||||||
| SL (mm) | 48.8–65.3 (n = 2) | 67.1 | 61.5 | 60.9 | 41.0 | 24.0–71.2 (n = 4) | 36.9–103.1 (n = 28) | |||
| % SL | mean ± SD | Range (mean ± SD) | ||||||||
| Skull length | 30.1–34.0 | 31.4 | 34.1 | 34.0 | 34.1 | 33.4 ± 1.3 | 29.9–40.8 | 28.9–39.3 (33.2 ± 2.6) | ||
| Head width | 26.3–28.9 | 28.3 | 30.9 | 29.4 | 36.3 | 31.2 ± 3.6 | 24.3–25.3 | 25.6–33.1 (29.3 ± 1.9) | ||
| Head depth | - | 27.6 | 31.2 | 29.2 | 34.1 | 30.5 ± 2.8 | - | 28.0–39.6 (31.9 ± 2.6) | ||
| Eye diameter | 13.0–13.3 | 11.3 | 10.9 | 12.6 | 12.4 | 11.8 ± 0.8 | 10.5–15.4 | 8.0–14.6 (11.1 ± 1.5) | ||
| Interorbital width | 20.3–21.4 | 19.7 | 20.3 | 20.4 | 22.7 | 20.8 ± 1.3 | 14.0–19.2 | 14.0–20.6 (17.5 ± 1.8) | ||
| Illicial trough width | - | 11.3 | 10.7 | - | 11.5 | 11.2 ± 0.4 | - | 8.7–13.2 (11.1 ± 1.5) | ||
| Mouth width | 30.3–31.8 | 25.2 | 29.3 | 31.7 | 34.1 | 30.1 ± 3.8 | 22.8–28.0 | 23.6–35.5 (28.1 ± 2.8) | ||
| Upper jaw length | 17.3–18.4 | 16.2 | 17.6 | 16.4 | 21.0 | 17.8 ± 2.2 | 15.6–20.8 | 13.0–24.1 (17.9 ± 2.6) | ||
| Predorsal length | 79.6 (?) | 64.8 | 70.2 | 63.5 | 63.2 | 65.4 ± 3.3 | 66.1–76.7 | 62.3–70.4 (66.9 ± 2.4) | ||
| Pre-anus length | 58.8–63.9 | 62.7 | 67.3 | 59.1 | 74.1 | 65.8 ± 6.5 | 59.4–69.6 | 62.1–71.4 (66.5 ± 2.6) | ||
| Preanal length | 77.0–77.5 | 76.8 | 77.4 | 75.7 | 80.0 | 77.5 ± 1.8 | 71.7–77.4 | 72.8–80.9 (77.2 ± 2.0) | ||
| Tail length | - | 37.3 | 32.7 | 40.9 | 25.9 | 34.2 ± 6.5 | - | 28.6–37.9 (33.5 ± 2.6) | ||
| Disk margin | 45.0–45.1 | 46.2 | 46.5 | 44.8 | 51.2 | 47.2 ± 2.8 | 41.5–47.9 | 41.4–51.9 (46.8 ± 2.6) | ||
| Dorsal-fin length | - | 23.2 | 22.3 | 19.5 | 22.2 | 21.8 ± 1.6 | - | 19.3–29.9 (24.6 ± 2.4) | ||
| Pectoral-fin length | 31.3–32.6 | 28.5 | 30.6 | 28.7 | 34.4 | 30.5 ± 2.7 | 28.0–34.7 | 26.5–39.8 (32.4 ± 3.7) | ||
| Anal-fin length | - | 24.9 | 22.8 | 22.2 | 27.1 | 24.2 ± 2.2 | - | 17.6–29.0 (23.1 ± 2.7) | ||
| Caudal-fin length | - | 29.5 | 31.2 | 27.9 | 35.4 | 31.0 ± 3.2 | - | 30.8–36.0 (33.9 ± 2.3) | ||
| Meristics | Range | n = 33 | ||||||||
| Dorsal-fin rays | 6 | 5 | 6 | 6 | 5 | 5–6 | 5–6 | 5–7 (mainly 6) | ||
| Pectoral-fin rays | 14–15 | 14 | 15 | 16 | 16 | 14–16 | 14–16 | 15–17 | ||
| Anal-fin rays | - | 4 | 4 | 4 | 4 | 4 | - | 4 | ||
| Lateral-line neuromasts | ||||||||||
| SS | 6–8 | 8 | 9 | 7 | 9 | 6–9 | 6–8 | 8 | ||
| PMS | - | 1 | 1 | - | 1 | 1 | 1 | |||
| CS | 8 (HT) | 8 | 8 | 8 | 8 | 8 | 8 (HT) | 8–9 | ||
| BS | 9–10 | 10 | 14 | 10 | 13 | 9–14 | 9–10 | 9–14 (11–12) | ||
| SOS | 5–6 | 7 | 6 | 6 | 6 | 5–7 | 5–6 | 5–6 (6) | ||
| POS | 3–4 | 4;5 | 3 | 5 | 4 | 3–5 | 3–4 | 3–6 (4–5) | ||
| DBSS | 3–5 | 3;4 | 4 | 4 | 4 | 3–5 | 3–5 | 4–5 | ||
| PBSS | 3 (HT) | 3 | 3 | 3 | 3 | 3 | 3 (HT) | 3 | ||
| VS | 3;4 (HT) | 5;6 | 3;4 | 4;4 | 4;5 | 3–6 | 4 (HT) | 3–4 | ||
| TS | 7;8 (HT) | 8;9 | 9 | 8 | 8 | 7–9 | 11-13 | 8–10 (9) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, H.-C. Taxonomy and Distribution of the Deep-Sea Batfish Genus Halieutopsis (Teleostei: Ogcocephalidae), with Descriptions of Five New Species. J. Mar. Sci. Eng. 2022, 10, 34. https://doi.org/10.3390/jmse10010034
Ho H-C. Taxonomy and Distribution of the Deep-Sea Batfish Genus Halieutopsis (Teleostei: Ogcocephalidae), with Descriptions of Five New Species. Journal of Marine Science and Engineering. 2022; 10(1):34. https://doi.org/10.3390/jmse10010034
Chicago/Turabian StyleHo, Hsuan-Ching. 2022. "Taxonomy and Distribution of the Deep-Sea Batfish Genus Halieutopsis (Teleostei: Ogcocephalidae), with Descriptions of Five New Species" Journal of Marine Science and Engineering 10, no. 1: 34. https://doi.org/10.3390/jmse10010034
APA StyleHo, H.-C. (2022). Taxonomy and Distribution of the Deep-Sea Batfish Genus Halieutopsis (Teleostei: Ogcocephalidae), with Descriptions of Five New Species. Journal of Marine Science and Engineering, 10(1), 34. https://doi.org/10.3390/jmse10010034