The Common Bean (Phaseolus vulgaris) Basic Leucine Zipper (bZIP) Transcription Factor Family: Response to Salinity Stress in Fertilized and Symbiotic N2-Fixing Plants

 , , and
, , and
Abstract
1. Introduction
2. Results
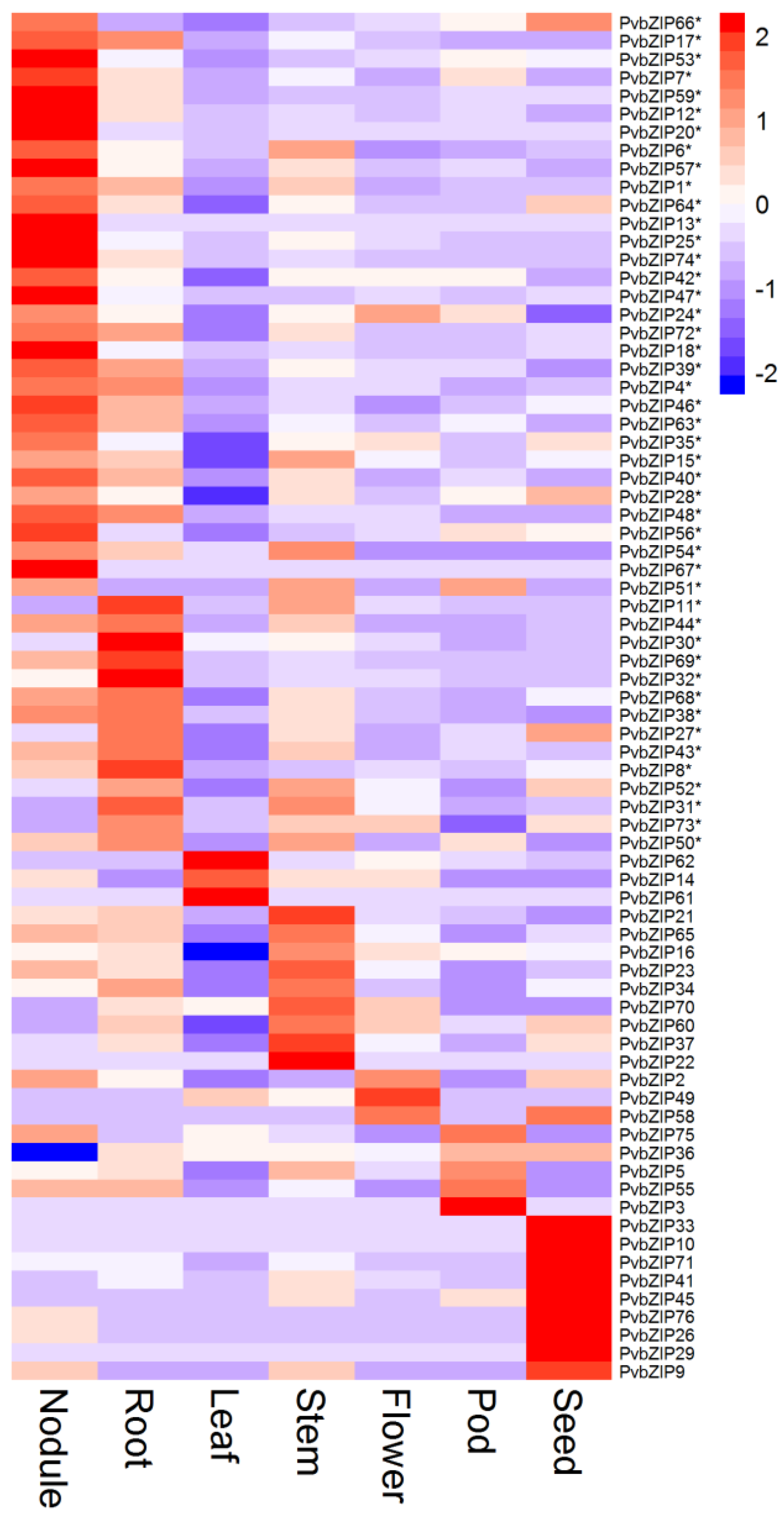
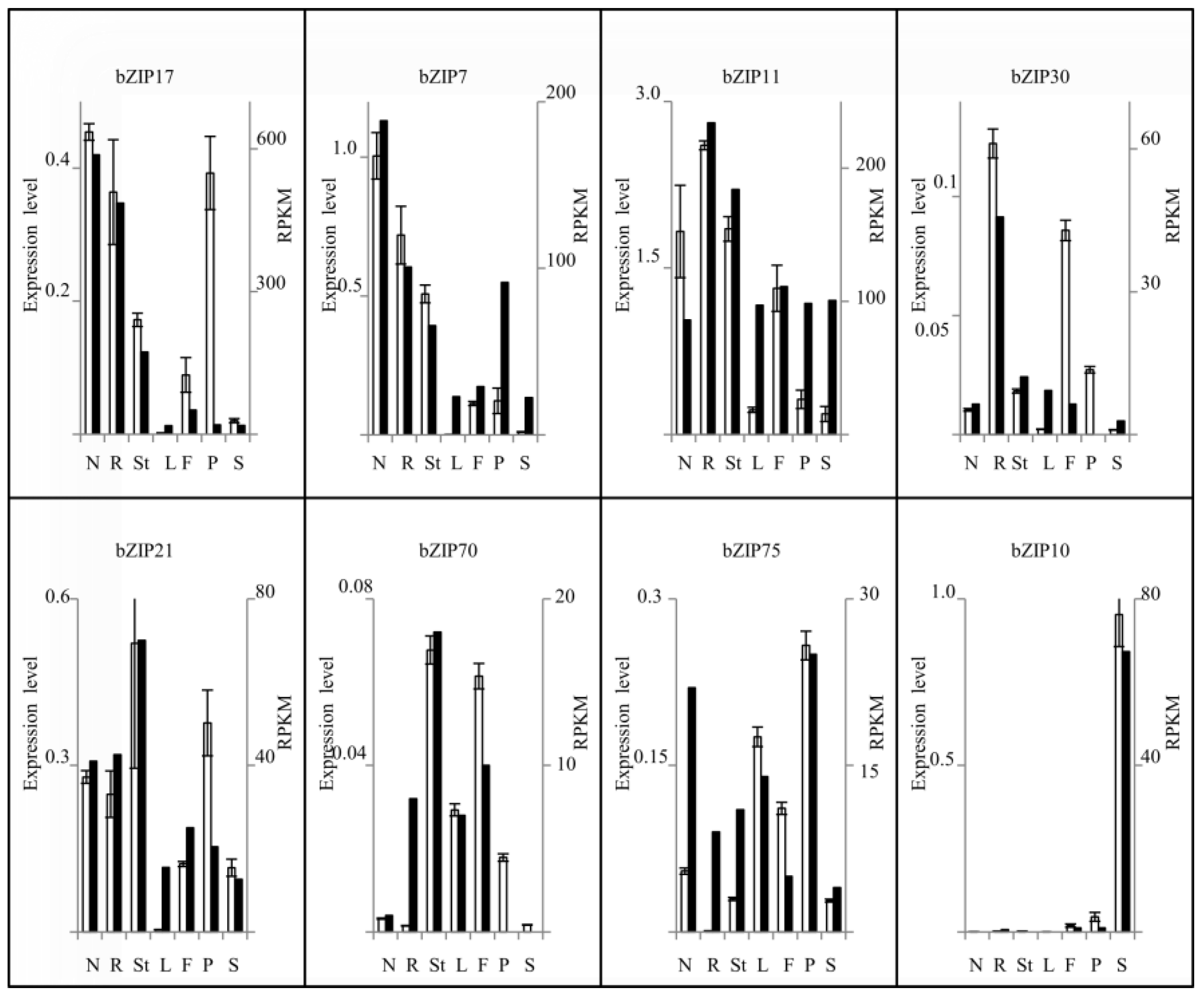
2.1. Expression Profile of P. vulgaris bZIP TF Genes in Different Plant Tissues
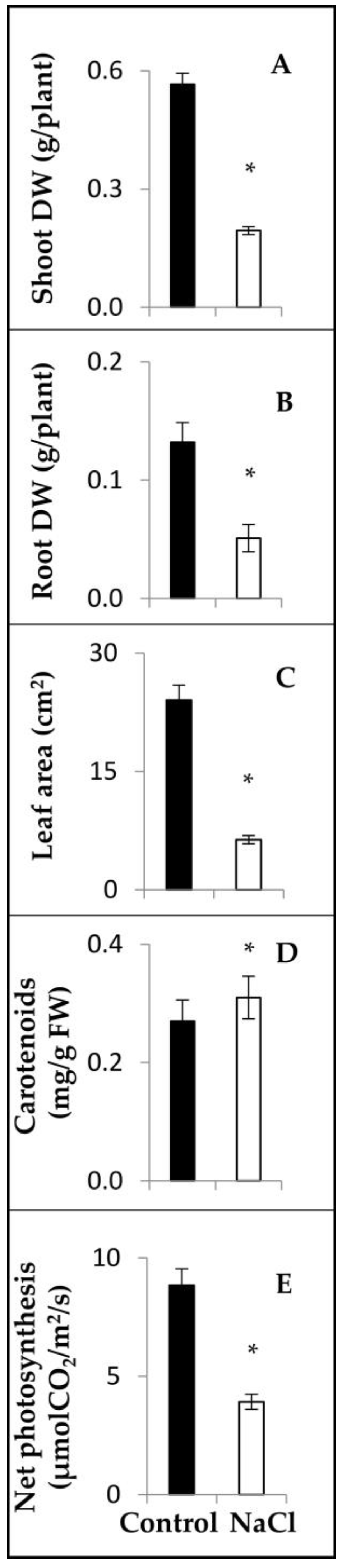
2.2. Salinity Response Profile of PvbZIP Genes from Fertilized Plants
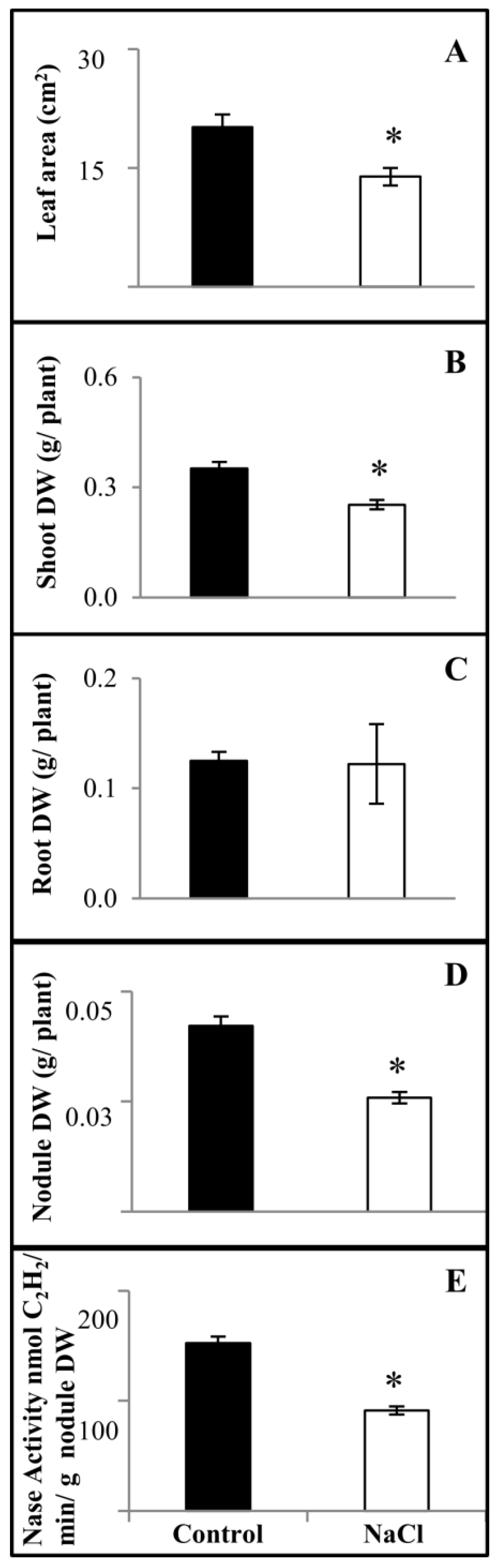
2.3. Salinity Response Profile of PvbZIP Genes from SNF Plants
2.4. Exploring the Downstream PvbZIP Regulation in Salt-Stressed Plants
3. Discussion
4. Materials and Methods
4.1. Selection of the P. vulgaris bZIP Genes
4.2. Plant Material and Growth Conditions
4.3. Phenotypic Analysis
4.4. RNA Isolation and qRT-PCR
4.5. Analysis of Genes Co-Expressed with Salt-Responsive PvbZIP Genes
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Wignarajah, K. Growth-Response of Phaseolus vulgaris to Varying Salinity Regimes. Environ. Exp. Bot. 1990, 30, 141–147. [Google Scholar] [CrossRef]
- Delgado, M.J.; Ligero, F.; Lluch, C. Effects of Salt Stress on Growth and Nitrogen-Fixation by Pea, Faba-Bean, Common Bean and Soybean Plants. Soil Biol. Biochem. 1994, 26, 371–376. [Google Scholar] [CrossRef]
- Cordovilla, M.P.; Ligero, F.; Lluch, C. Effect of salinity on growth, nodulation and nitrogen assimilation in nodules of faba bean (Vicia faba L.). Appl. Soil Ecol. 1999, 11, 1–7. [Google Scholar] [CrossRef]
- Gama, P.B.S.; Inanaga, S.; Tanaka, K.; Nakazawa, R. Physiological response of common bean (Phaseolus vulgaris L.) seedlings to salinity stress. Afr. J. Biotechnol. 2007, 6, 88. [Google Scholar]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, Y.; Cai, H.; Bai, X.; Ji, W.; Ding, X.; Zhu, Y. The Arabidopsis AtbZIP1 transcription factor is a positive regulator of plant tolerance to salt, osmotic and drought stresses. J. Plant Res. 2012, 125, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Weisshaar, B.; Droge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F.; bZIP Research Group. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Joshi, T.; Benedito, V.A.; Xu, D.; Udvardi, M.K.; Stacey, G. Legume transcription factor genes: What makes legumes so special? Plant Physiol. 2009, 151, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cheng, K.; Wan, L.; Yan, L.; Jiang, H.; Liu, S.; Lei, Y.; Liao, B. Genome-wide analysis of the basic leucine zipper (bZIP) transcription factor gene family in six legume genomes. BMC Genom. 2015, 16, 1053. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Zou, H.F.; Wei, W.; Hao, Y.J.; Tian, A.G.; Huang, J.; Liu, Y.F.; Zhang, J.S.; Chen, S.Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Hiz, M.C.; Canher, B.; Niron, H.; Turet, M. Transcriptome analysis of salt tolerant common bean (Phaseolus vulgaris L.) under saline conditions. PLoS ONE 2014, 9, e92598. [Google Scholar] [CrossRef] [PubMed]
- Broughton, W.J.; Hernández, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef]
- O’Rourke, J.A.; Iniguez, L.P.; Fu, F.; Bucciarelli, B.; Miller, S.S.; Jackson, S.A.; McClean, P.E.; Li, J.; Dai, X.; Zhao, P.X.; et al. An RNA-Seq based gene expression atlas of the common bean. BMC Genom. 2014, 15, 866. [Google Scholar] [CrossRef] [PubMed]
- Bhawna, V.S.B.; Gajula, M.P. PvTFDB: A Phaseolus vulgaris transcription factors database for expediting functional genomics in legumes. Database 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Valdes-Lopez, O.; Arenas-Huertero, C.; Ramirez, M.; Girard, L.; Sanchez, F.; Vance, C.P.; Luis Reyes, J.; Hernandez, G. Essential role of MYB transcription factor: PvPHR1 and microRNA: PvmiR399 in phosphorus-deficiency signalling in common bean roots. Plant Cell Environ. 2008, 31, 1834–1843. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Fabre, R.; Guillen, G.; Loredo, M.; Arellano, J.; Valdes-Lopez, O.; Ramirez, M.; Iniguez, L.P.; Panzeri, D.; Castiglioni, B.; Cremonesi, P.; et al. Common bean (Phaseolus vulgaris L.) PvTIFY orchestrates global changes in transcript profile response to jasmonate and phosphorus deficiency. BMC Plant Biol. 2013, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Nova-Franco, B.; Iniguez, L.P.; Valdes-Lopez, O.; Alvarado-Affantranger, X.; Leija, A.; Fuentes, S.I.; Ramirez, M.; Paul, S.; Reyes, J.L.; Girard, L.; et al. The micro-RNA72c-APETALA2-1 node as a key regulator of the common bean-Rhizobium etli nitrogen fixation symbiosis. Plant Physiol. 2015, 168, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Uribe, L.; O’Connell, M.A. A root-specific bZIP transcription factor is responsive to water deficit stress in tepary bean (Phaseolus acutifolius) and common bean (P. vulgaris). J. Exp. Bot. 2006, 57, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- González, M.C.; Pérez, N.; Cristo, E.; Rodríguez, M.; Borrás, O. Development of salinity-tolerant rice varieties using biotechnological and nuclear techniques. In Induced Plant Mutations in the Genomics Era; Shu, G.Y., Ed.; FAO (Food and Agriculture Organization of the United Nations): Rome, Italy, 2009; pp. 138–140. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Graham, P.H.; Draeger, K.J.; Ferrey, M.L.; Conroy, M.J.; Hammer, B.E.; Martinez, E.; Aarons, S.R.; Quinto, C. Acid Ph Tolerance in Strains of Rhizobium and Bradyrhizobium, and Initial Studies on the Basis for Acid Tolerance of Rhizobium tropici Umr1899. Can. J. Microbiol. 1994, 40, 198–207. [Google Scholar] [CrossRef]
- Valdes-Lopez, O.; Yang, S.S.; Aparicio-Fabre, R.; Graham, P.H.; Reyes, J.L.; Vance, C.P.; Hernandez, G. MicroRNA expression profile in common bean (Phaseolus vulgaris) under nutrient deficiency stresses and manganese toxicity. New Phytol. 2010, 187, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Shamseldin, A.; Werner, D. High salt and high pH tolerance of new isolated Rhizobium etli strains from Egyptian soils. Curr. Microbiol. 2005, 50, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; James, R.A.; Lauchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Bouhmouch, I.; Souad-Mouhsine, B.; Brhada, F.; Aurag, J. Influence of host cultivars and Rhizobium species on the growth and symbiotic performance of Phaseolus vulgaris under salt stress. J. Plant Physiol. 2005, 162, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Winicov, I. Alfin1 transcription factor overexpression enhances plant root growth under normal and saline conditions and improves salt tolerance in alfalfa. Planta 2000, 210, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Yokotani, N.; Ichikawa, T.; Kondou, Y.; Matsui, M.; Hirochika, H.; Iwabuchi, M.; Oda, K. Tolerance to various environmental stresses conferred by the salt-responsive rice gene ONAC063 in transgenic Arabidopsis. Planta 2009, 229, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Deng, C.; Ouyang, B.; Ye, Z. Molecular analysis of two salt-responsive NAC-family genes and their expression analysis in tomato. Mol. Biol. Rep. 2011, 38, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Summerfield, R.J.; Huxley, P.A.; Minchin, F.R. Plant Husbandry and Management-Techniques for Growing Grain Legumes under Simulated Tropical Conditions in Controlled Environments. Exp. Agric. 1977, 13, 81–92. [Google Scholar] [CrossRef]
- Franco, A.A.; Munns, D.N. Nodulation and growth of Phaseolus vulgaris in solution culture. Plant Soil 1982, 66, 149–160. [Google Scholar] [CrossRef]
- Abramoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with Image J. Biophoton. Int. 2004, 11, 42. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Hardy, R.W.; Holsten, R.D.; Jackson, E.K.; Burns, R.C. The acetylene-ethylene assay for n(2) fixation: Laboratory and field evaluation. Plant Physiol. 1968, 43, 1185–1207. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, G.; Ramirez, M.; Valdes-Lopez, O.; Tesfaye, M.; Graham, M.A.; Czechowski, T.; Schlereth, A.; Wandrey, M.; Erban, A.; Cheung, F.; et al. Phosphorus stress in common bean: Root transcript and metabolic responses. Plant Physiol. 2007, 144, 752–767. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.; Guillen, G.; Fuentes, S.I.; Iniguez, L.P.; Aparicio-Fabre, R.; Zamorano-Sanchez, D.; Encarnacion-Guevara, S.; Panzeri, D.; Castiglioni, B.; Cremonesi, P.; et al. Transcript profiling of common bean nodules subjected to oxidative stress. Physiol. Plant. 2013, 149, 389–407. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J.; Jenkins, J.; Shu, S.; Song, Q.; Chavarro, C.; et al. A reference genome for common bean and genome-wide analysis of dual domestications. Nat. Genet. 2014, 46, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.D.; Nacu, S. Fast and SNP-tolerant detection of complex variants and splicing in short reads. Bioinformatics 2010, 26, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Tarazona, S.; Furio-Tari, P.; Turra, D.; Pietro, A.D.; Nueda, M.J.; Ferrer, A.; Conesa, A. Data quality aware analysis of differential expression in RNA-seq with NOISeq R/Bioc package. Nucleic Acids Res. 2015, 43, e140. [Google Scholar] [CrossRef] [PubMed]
- Frith, M.C.; Fu, Y.; Yu, L.; Chen, J.F.; Hansen, U.; Weng, Z. Detection of functional DNA motifs via statistical over-representation. Nucleic Acids Res. 2004, 32, 1372–1381. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normalized Expression Level a | Expression Ratio b | ||
|---|---|---|---|
| Gene ID | NaCl | Control | NaCl/C |
| Up-regulated | |||
| PvbZIP11 | 4.1 ± 0.02 | 2.8 ± 0.31 | 1.5 |
| PvbZIP13 | 1 × 10−1 ± 0.01 | 1 × 10−2 ± 0.00 | 13.0 |
| PvbZIP20 | 2 × 10−1 ± 0.01 | 1 × 10−1 ± 0.01 | 2.7 |
| PvbZIP28 | 1.1 ± 0.04 | 0.4 ± 0.05 | 2.8 |
| PvbZIP39 | 2.2 ± 0.17 | 1.4 ± 0.04 | 1.6 |
| PvbZIP50 | 1.1 ± 0.08 | 4 × 10−2 ± 0.01 | 2.7 |
| PvbZIP51 | 2 × 10−2 ± 0.00 | 1 × 10−2 ± 0.00 | 2.0 |
| PvbZIP57 | 0.3 ± 0.01 | 0.2 ± 0.01 | 1.7 |
| PvbZIP63 | 0.2 ± 0.02 | 0.1 ± 0.00 | 2.5 |
| PvbZIP66 | 2.1 ± 0.04 | 1.2 ± 0.02 | 1.7 |
| PvbZIP74 | 0.1 ± 0.01 | 0.1 ± 0.01 | 1.9 |
| Down-regulated | |||
| PvbZIP17 | 0.5 ± 0.03 | 1.3 ± 0.08 | −2.8 |
| PvbZIP30 | 0.2 ± 0.04 | 0.5 ± 0.01 | −2.3 |
| PvbZIP32 | 0.4 ± 0.02 | 0.6 ± 0.02 | −1.8 |
| PvbZIP38 | 0.2 ± 0.01 | 0.5 ± 0.04 | −3.0 |
| PvbZIP44 | 0.6 ± 0.07 | 1.1 ± 0.02 | −1.7 |
| ROOTS | NODULES | ||||||
|---|---|---|---|---|---|---|---|
| Normalized Expression Level a | Expression Ratio b | Normalized Expression Level a | Expression Ratio b | ||||
| Gene ID | NaCl | Control | NaCl/C | Gene ID | NaCl | Control | NaCl/C |
| Up-regulated | Up-regulated | ||||||
| PvbZIP6 | 8.4 ± 0.31 | 3.8 ± 0.20 | 2.2 | PvbZIP30 | 3 × 10−1 ± 0.02 | 1 × 10−1 ± 0.02 | 2.3 |
| PvbZIP8 | 1 × 10−1 ± 0.00 | 2 × 10−2 ± 0.00 | 2.7 | Down-regulated | |||
| PvbZIP24 | 2.6 ± 0.22 | 1.3 ± 0.16 | 1.9 | PvbZIP8 | 2 × 10−3 ± 0.00 | 1 × 10−2 ± 0.00 | −4.9 |
| PvbZIP28 | 1 × 10−1 ± 0.01 | 4 × 10−2 ± 0.00 | 2.0 | PvbZIP28 | 3 × 10−3 ± 0.00 | 1 × 10−2 ± 0.00 | −2.5 |
| PvbZIP52 | 2.2 ± 0.06 | 1.4 ± 0.18 | 1.6 | PvbZIP32 | 1 × 10−1 ± 0.01 | 1 × 10−1 ± 0.01 | −1.7 |
| PvbZIP53 | 35.8 ± 4.86 | 14.0 ± 0.91 | 2.6 | PvbZIP39 | 1 × 10−1 ± 0.02 | 4 × 10−1 ± 0.02 | −4.8 |
| PvbZIP54 | 4 × 10−2 ± 0.00 | 2 × 10−2 ± 0.00 | 2.2 | PvbZIP42 | 1 × 10−1 ± 0.01 | 1 × 10−1 ± 0.01 | −1.5 |
| PvbZIP57 | 3.8 ± 0.10 | 2.1 ± 0.26 | 1.8 | PvbZIP50 | 1 × 10−3 ± 0.00 | 1 × 10−1 ± 0.02 | −13.2 |
| PvbZIP66 | 15.4 ± 2.50 | 9.0 ± 0.32 | 1.7 | PvbZIP53 | 1.8 ± 0.01 | 3.5 ± 0.01 | −1.9 |
| Down-regulated | PvbZIP56 | 3 × 10−1 ± 0.02 | 6 × 10−1 ± 0.03 | −1.9 | |||
| PvbZIP30 | 2.7 ± 0.06 | 4.9 ± 0.30 | −1.8 | PvbZIP57 | 1 × 10−2 ± 0.00 | 1 × 10−2 ± 0.00 | −1.9 |
| PvbZIP63 | 3 × 10−1 ± 0.03 | 5 × 10−1 ± 0.01 | −2.1 | ||||
| PvbZIP68 | 1 × 10−3 ± 0.00 | 4 × 10−3 ± 0.00 | −3.8 | ||||
| PvbZIP69 | 2 × 10−1 ± 0.02 | 3 × 10−1 ± 0.00 | −1.9 | ||||
| GO Identifier | Description (Molecular Function) | p | Number of Genes |
|---|---|---|---|
| GO:0016491 | oxidoreductase activity | 0.001 | 50 |
| GO:0003700 | transcription factor activity, sequence-specific DNA binding | 0.007 | 18 |
| GO:0015116 | sulfate transmembrane transporter activity | 0.003 | 3 |
| GO:0004657 | proline dehydrogenase activity | 0.001 | 2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayra, L.; Ramírez, M.; Íñiguez, L.P.; Rodés, R.; Ortega, E.; Hernández, G. The Common Bean (Phaseolus vulgaris) Basic Leucine Zipper (bZIP) Transcription Factor Family: Response to Salinity Stress in Fertilized and Symbiotic N2-Fixing Plants. Agriculture 2018, 8, 160. https://doi.org/10.3390/agriculture8100160
Ayra L, Ramírez M, Íñiguez LP, Rodés R, Ortega E, Hernández G. The Common Bean (Phaseolus vulgaris) Basic Leucine Zipper (bZIP) Transcription Factor Family: Response to Salinity Stress in Fertilized and Symbiotic N2-Fixing Plants. Agriculture. 2018; 8(10):160. https://doi.org/10.3390/agriculture8100160
Chicago/Turabian StyleAyra, Litzy, Mario Ramírez, Luis P. Íñiguez, Rosa Rodés, Eduardo Ortega, and Georgina Hernández. 2018. "The Common Bean (Phaseolus vulgaris) Basic Leucine Zipper (bZIP) Transcription Factor Family: Response to Salinity Stress in Fertilized and Symbiotic N2-Fixing Plants" Agriculture 8, no. 10: 160. https://doi.org/10.3390/agriculture8100160
APA StyleAyra, L., Ramírez, M., Íñiguez, L. P., Rodés, R., Ortega, E., & Hernández, G. (2018). The Common Bean (Phaseolus vulgaris) Basic Leucine Zipper (bZIP) Transcription Factor Family: Response to Salinity Stress in Fertilized and Symbiotic N2-Fixing Plants. Agriculture, 8(10), 160. https://doi.org/10.3390/agriculture8100160