Photosynthesis of Sago Palm (Metroxylon sagu Rottb.) Seedling at Different Air Temperatures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Culture Conditions
2.2. Diurnal Leaf Gas Exchange
2.3. Assimilation Rate vs. CO2 Concentration (A/Ci Curve)
2.4. Photosynthetic Rate vs. Irradiance
2.5. Chlorophyll Content
2.6. Statistical Analysis
3. Results
3.1. Diurnal Leaf Gas Exchange
3.2. Photosynthetic Activity and Chlorophyll Content
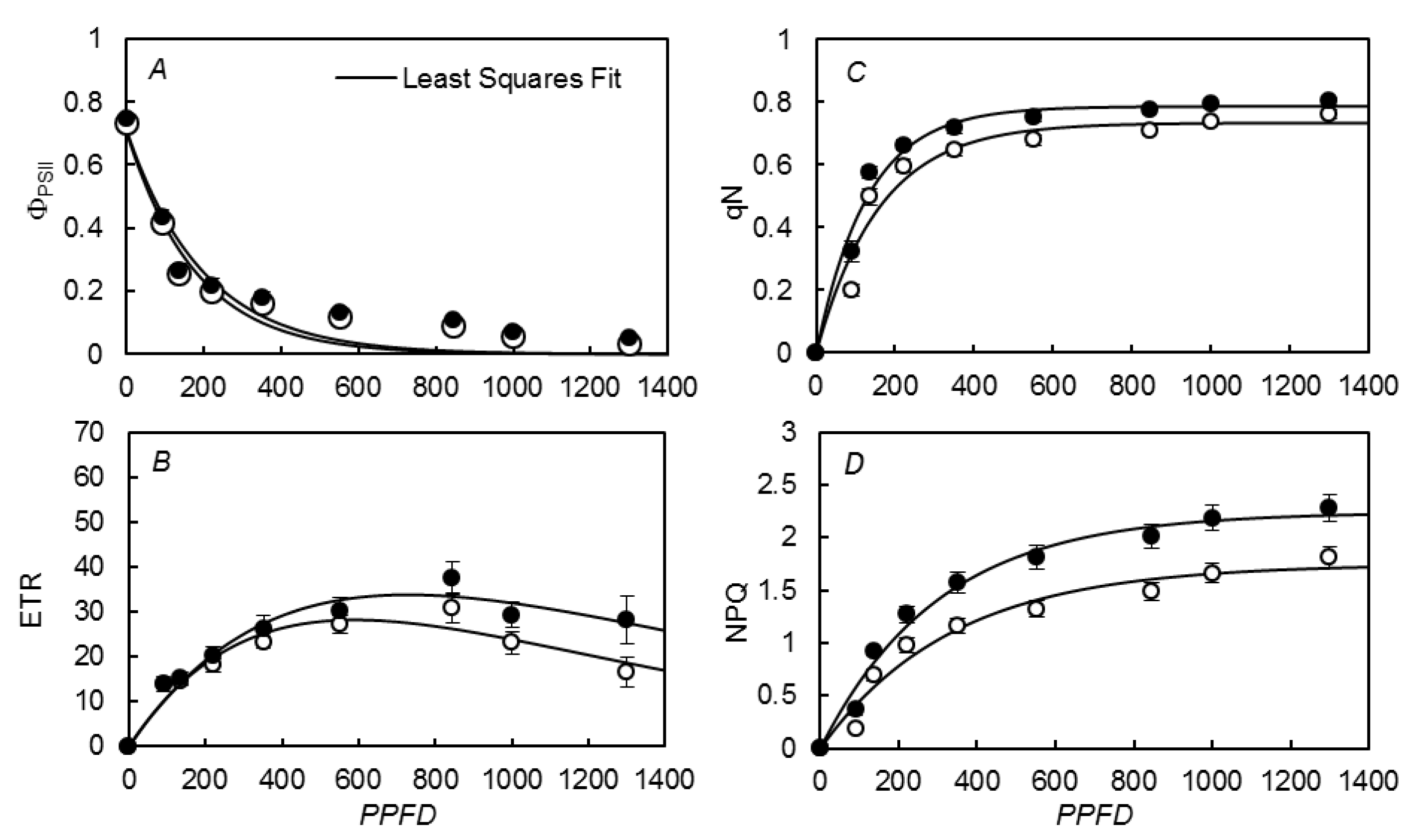
3.3. Photosynthetic Rate vs. Irradiances
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Larcher, W. Physiological Plant Ecology; Springer: Berlin/Heidelberg, Germany, 2003; p. 504. [Google Scholar]
- Ehara, H. Geographycal distributon and specification of Metroxylon palms. Jpn. J. Trop. Agric. 2005, 50, 229–233. [Google Scholar]
- Ehara, H.; Susanto, S.; Mizota, C.; Shohei, H.; Tadashi, M. Sago palm (Metroxylon sagu, Areaceae) production in the eastern archipelago of Indonesia: Variation in morphological charactersitics and pith-dry matter yield. Econ. Bot. 2000, 54, 197–206. [Google Scholar] [CrossRef]
- Flach, M.; Braber, K.D.; Frederix, M.J.J. Temperature and relative humidity requirement of young sago palm seedlings. In Proceedings of the Third International Sago Symposium; Yamada, N., Kainuma, K., Eds.; The Sago Palm Research Fund: Tokyo, Japan, 1986; pp. 139–143. [Google Scholar]
- Schuiling, D.L. Growth and Development of True Sago Palm (Metroxylon sagu Rottboll) with Special Fererence to Accumulate of Starch in the Trunk, a Study on Morphology, Genetic Variation and Ecophysiology and Their Implications for Cultivation. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2009; pp. 3–9. [Google Scholar]
- Okazaki, M.; Kimura, S.D. Ecology of the sago palm. In The Sago Palm: The Food and Environmental Challenges of the 21st Century; The Society of Sago Palm Studies, Ed.; Kyoto University Press: Kyoto, Japan, 2015; pp. 41–60. [Google Scholar]
- Yamori, W.; Caemmerer, S.V. Effect of rubisco activase deficiency on the temperature response of CO2 assimilation rate and rubisco activation state: Insights from transgenic tobacco with reduced amount of rubisco activase. Plant Physiol. 2009, 151, 2072–2082. [Google Scholar] [CrossRef] [PubMed]
- Paredes, M.; Quiles, M.J. The effect of cold stress on photosynthesis in hibiscus plant. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Irawan, A.F.; Yamamoto, Y.; Miyazaki, A.; Tetsushi, Y. Effect of various ranges of controlled air temperatures on the early growth of sago palm (Metroxylon sagu Rottb.) seedlings. Trop. Agric. Dev. 2011, 55, 68–74. [Google Scholar]
- Ehara, H.; Komada, C.; Morita, O. Germination characteristics of sago palm seeds and spine emergence in seedlings produced from spineless palm seeds. Principes 1998, 42, 212–217. [Google Scholar]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B. 2014, 137, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Hew, C.S.; Krotkov, G.; David, T. Effects of temperature on photosynthesis and CO2 evolution in light and darkness by green leaves. Plant Physiol. 1968, 44, 671–677. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 345, 659–668. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Caemmerer, V.S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting photosynthetic carbon response curves for C3 leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D. What gas exchange data can tell us about photosynthesis. Plant Cell Environ. 2016, 39, 1161–1163. [Google Scholar] [CrossRef] [PubMed]
- Klughammer, C.; Schreiber, U. Complementary PSII quantum yields calculated from simple fluorescence parameters measured by pam fluorometry and the saturation pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, R.J.; Bunthawin, S. The use of pulse amplitude modulation (PAM) fluorometry to measure photosynthesis in a CAM orchid, dendrobium SPP. (D. CV. Viravuth pink). Int. J. Plant Sci. 2010, 171, 575–585. [Google Scholar] [CrossRef]
- Apichatmeta, K.; Sudsiri, C.J.; Ritchie, R.J. Photosynthesis of oil palm (Elaeis guineensis). Sci. Hortic. Amst. 2017, 214, 34–40. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts: Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic membranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2012; pp. 106–130. [Google Scholar]
- Koyama, K.; Takemoto, S. Morning reduction of photosynthetic capacity before midday depression. Sci. Rep. 2014. [Google Scholar] [CrossRef] [PubMed]
- Cen, Y.P.; Sage, R.F. The regulation of ribulose-1,5-bisposphate carboxylase activity in response to variation in temperature and atmospheric CO2 partial pressure in sweet potato. Plant Physiol. 2005, 139, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Camejo, D.; Rodríguez, P.; Morales, M.A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An Overview. Environ. Exp. Bot. 2007, 6, 199–223. [Google Scholar] [CrossRef]
- Beck, W.A.; Redman, R. Seasonal variation in the production of plant pigments. Plant Physiol. 1940, 15, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Uchida, N.; Kobayashi, S.; Yasuda, T.; Yamaguchi, T. Photosynthetic characteristics of sago palm, Metroxylon rumphii Martius. Jpn. J. Trop. Agric. 1990, 34, 176–180. [Google Scholar]
- Salvucci, M.E.; Crafts-Brandner, S.J. Inhibition of photosynthesis by heat stress: The activation state of rubisco as a limiting factor in photosynthesis. Physiol. Plant. 2004, 120, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Marchand, F.L.; Mertens, S.; Kockelbergh, F.; Beyens, L.; Nijs, I. Performance of high arctic tundra plants improved during but deteriorated after exposure to a simulated extreme temperature event. Glob. Chang. Biol. 2005, 11, 2078–2089. [Google Scholar] [CrossRef]
- Ruban, A.V.; Horton, P. Spectroscopy of non-photochemical and photochemical quenching of chlorophyll fluorescence in leaves; evidence for a role of the light harvesting complex of photosystem II in the regulation of energy dissipation. Photosynth. Res. 1994, 40, 181–190. [Google Scholar] [CrossRef] [PubMed]
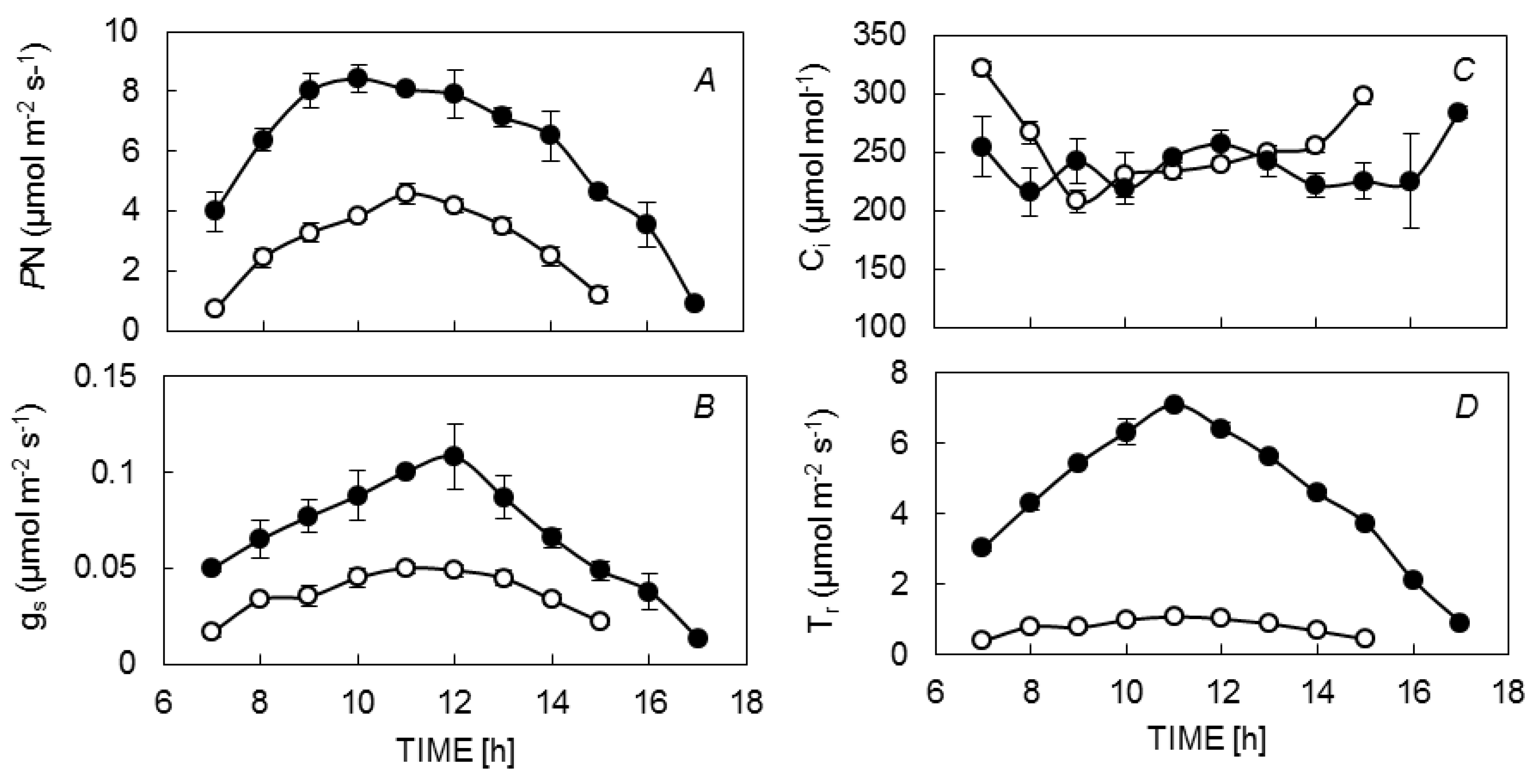


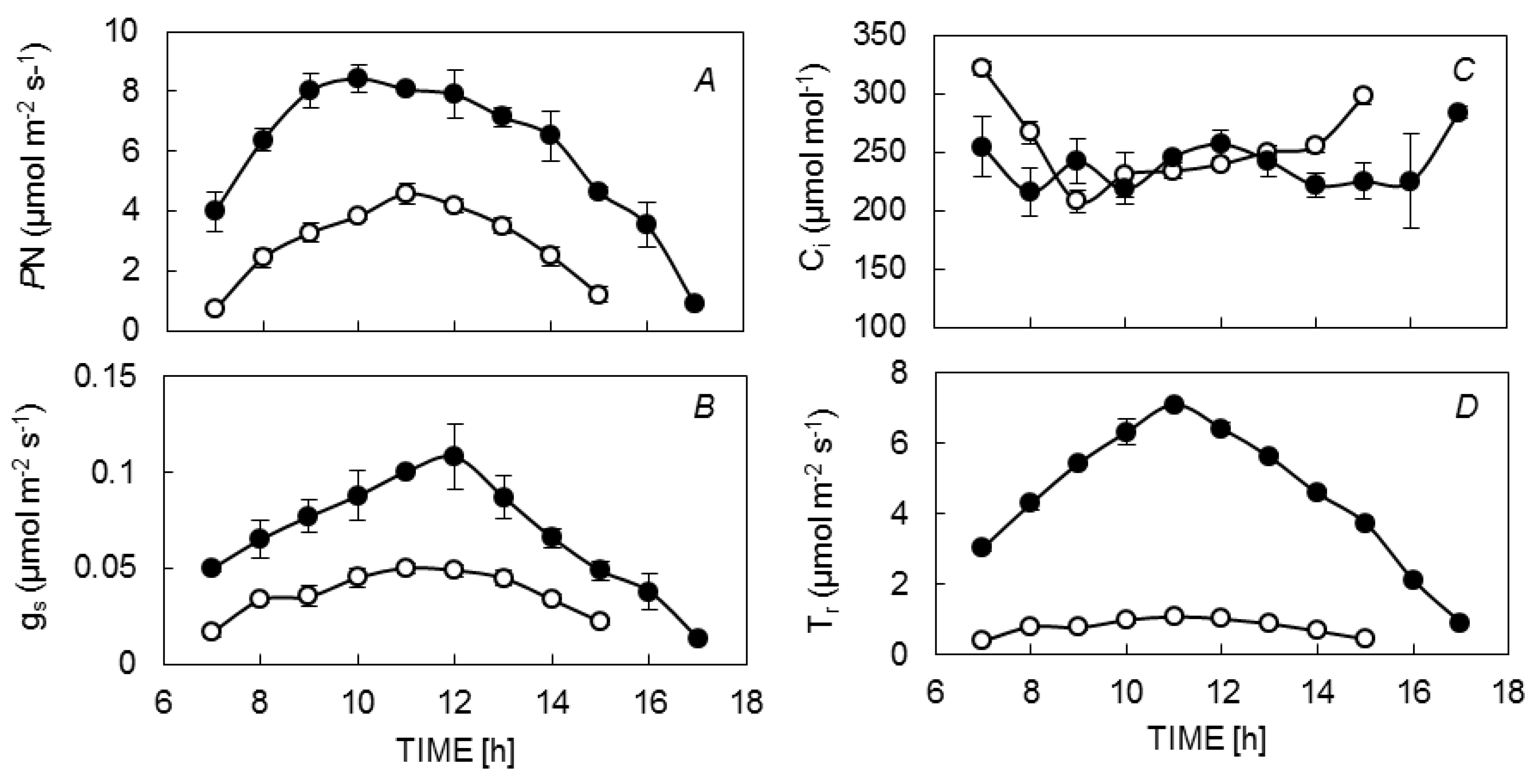{kind=link}
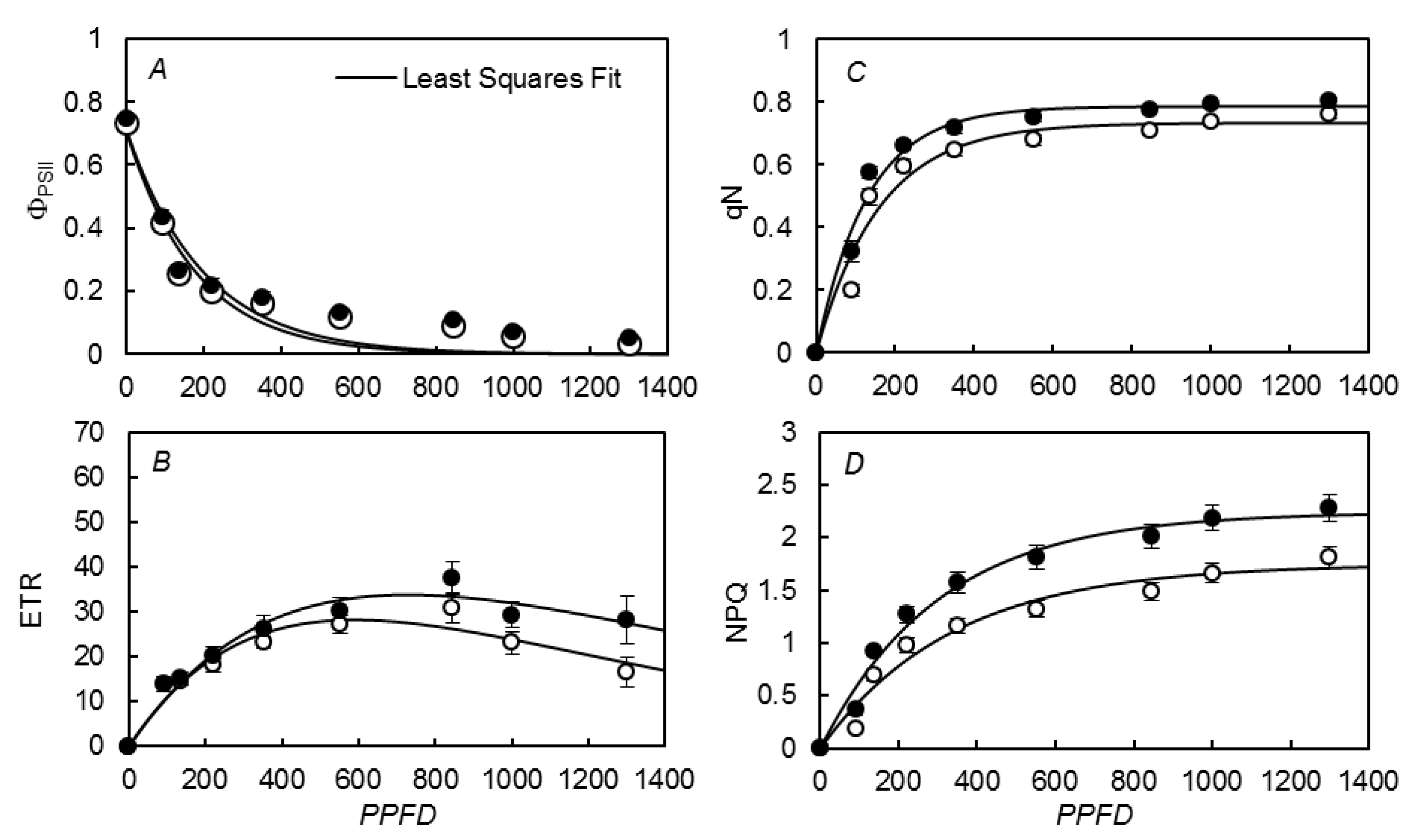{kind=link}
{kind=link}
| Parameters | 25–29 °C | 29–33 °C |
|---|---|---|
| PN [µmol (CO2) m−2 s−1] | 5.66 ± 0.91 | 8.62 ± 0.24 * |
| J [µmol m−2 s−1)] | 88.2 ± 10.98 | 114.1 ± 5.59 ns |
| Vcmax [µmol m−2 s−1)] | 40.1 ± 10.23 | 99.3 ± 10.86 * |
| SPAD | 46.2 ± 1.56 | 62.3 ± 1.91 ** |
| Chl a [µg g−1] | 838.1 ± 34.39 | 987.3 ± 99.04 ns |
| Chl b [µg g−1] | 258.5 ± 10.56 | 319.2 ± 11.45 * |
| Chl a + b [µg g−1] | 1096.6 ± 35.9 | 1306.5 ± 102.8 ns |
| Carotenoid [µg g−1] | 223.7 ± 2.91 | 255.0 ± 2.91 ns |
| Parameters | 25–29 °C | 29–33 °C |
|---|---|---|
| Maximum yield (ΦPSIImax) | 0.70 ± 0.013 | 0.70 ± 0.025 ns |
| Optimum PPFD (PPFDopt) | 587.00 ± 29.81 | 725.59 ± 33.32 * |
| Maximum electron transport rate (ETRmax) | 28.28 ± 2.53 | 33.91 ± 3.76 ns |
| Asymptotic Photosynthetic efficiency (αo) | 0.13 ± 0.006 | 0.13 ± 0.009 ns |
| qNmax | 0.73 ± 0.015 | 0.79 ± 0.014 ns |
| NPQmax | 1.75 ± 0.08 | 2.25 ± 0.13 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azhar, A.; Makihara, D.; Naito, H.; Ehara, H. Photosynthesis of Sago Palm (Metroxylon sagu Rottb.) Seedling at Different Air Temperatures. Agriculture 2018, 8, 4. https://doi.org/10.3390/agriculture8010004
Azhar A, Makihara D, Naito H, Ehara H. Photosynthesis of Sago Palm (Metroxylon sagu Rottb.) Seedling at Different Air Temperatures. Agriculture. 2018; 8(1):4. https://doi.org/10.3390/agriculture8010004
Chicago/Turabian StyleAzhar, Aidil, Daigo Makihara, Hitoshi Naito, and Hiroshi Ehara. 2018. "Photosynthesis of Sago Palm (Metroxylon sagu Rottb.) Seedling at Different Air Temperatures" Agriculture 8, no. 1: 4. https://doi.org/10.3390/agriculture8010004
APA StyleAzhar, A., Makihara, D., Naito, H., & Ehara, H. (2018). Photosynthesis of Sago Palm (Metroxylon sagu Rottb.) Seedling at Different Air Temperatures. Agriculture, 8(1), 4. https://doi.org/10.3390/agriculture8010004