
Grain Sorghum: A Conundrum for Chicken-Meat Production

Abstract
:1. Introduction
2. Background
3. Starch
4. Apices of the Sorghum Bermuda Triangle
4.1. Kafirin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response | Observations (n =) | Experiment Leverage (p =) | Dietary Kafirin Leverage (p =) | Whole Model |
|---|---|---|---|---|
| ME:GE | 10 | 0.109 | 0.034 | R2 = 0.71, p = 0.045 |
| AMEn | 13 | 0.485 | 0.013 | R2 = 0.70, p = 0.074 |
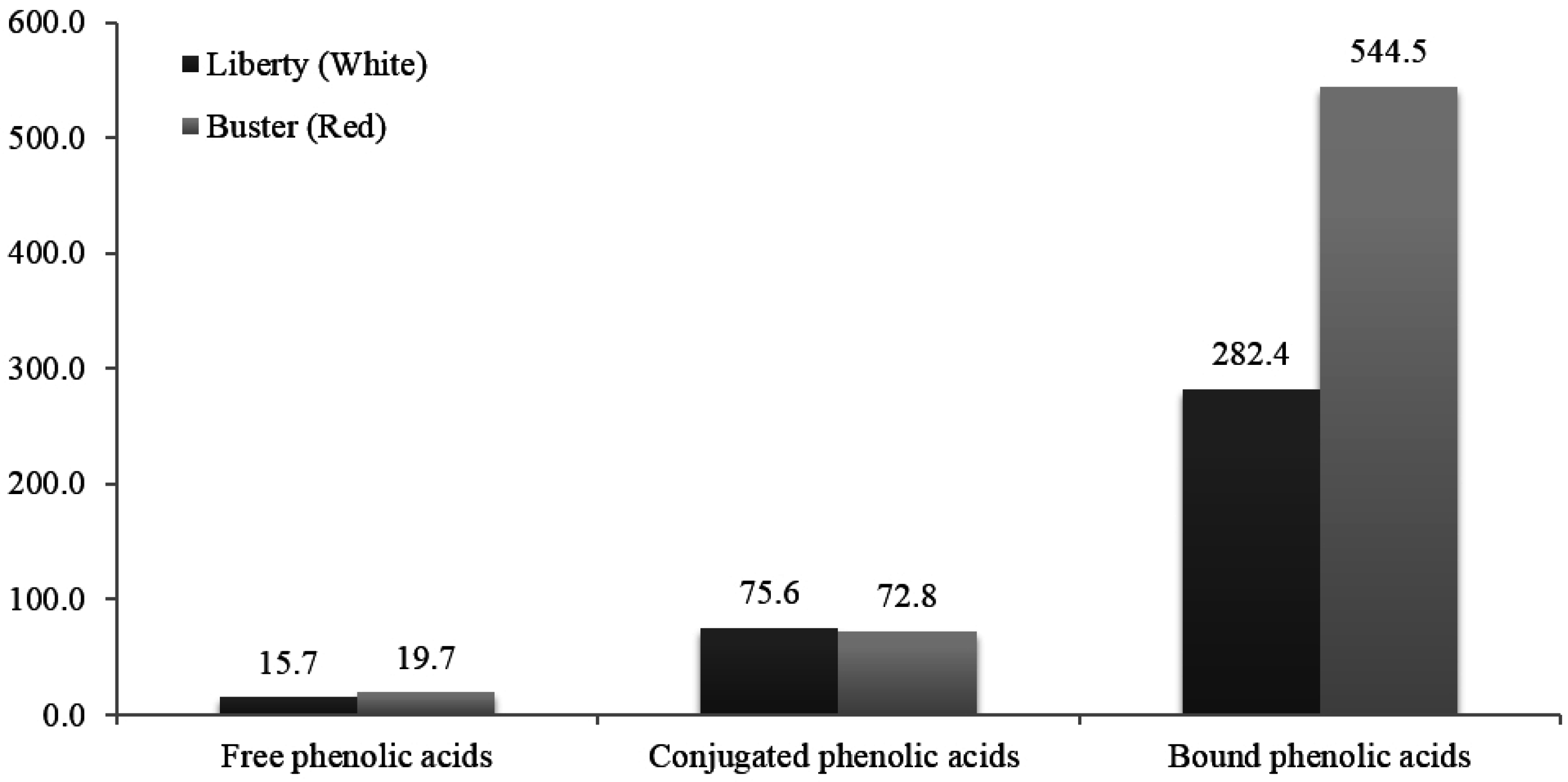
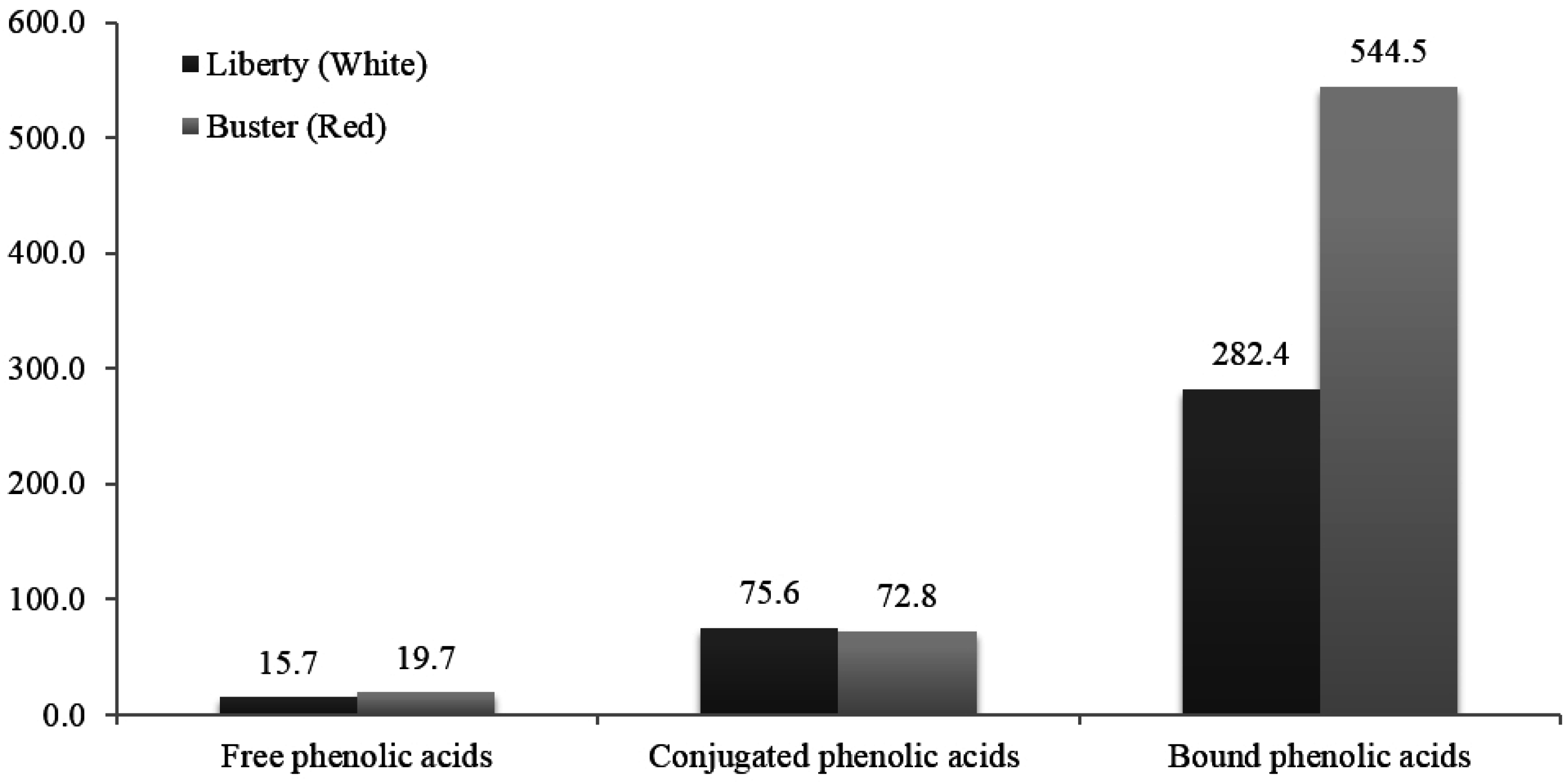
4.2. “Non-tannin” Polyphenolic Compounds and Phenolic Acids
4.3. Phytate
5. White versus Red Sorghum
| Parameter | Red Sorghums (n = 28) | White Sorghums (n = 3) |
|---|---|---|
| Starch (g/kg) | 637 | 656 |
| Protein (g/kg) | 117 | 105 |
| Kafirin index 1 | 5.48 | 4.27 |
| Phytate (g/kg) | 7.37 | 6.27 |
| “Phenolic compounds” | 6.99 | 1.53 |
| Apparent ileal starch digestibility | 0.881 | 0.908 |
| Apparent metabolisable energy (AME) | 15.42 MJ/kg | 15.87 MJ/kg |
| Range | 14.3–16.2 | 15.6–16.1 |
| N-corrected AME (AMEn) | 14.94 MJ/kg | 15.33 MJ/kg |
| Range | 14.0–15.2 | 15.2–15.5 |
6. Starch and Protein Digestive Dynamics in Sorghum-Based Broiler Diets
| Item | Rapidly Digested Starch (g/kg DM) | Slowly Digested Starch (g/kg DM) | Resistant Starch (g/kg DM) | Potential Digestible Starch (g/kg DM) | Rate of Starch Digestion (min−1) |
|---|---|---|---|---|---|
| Maize | 147 | 367 | 191 | 95.0 | 0.017 |
| Barley | 151 | 264 | 143 | 88.6 | 0.024 |
| Wheat | 181 | 379 | 53 | 92.8 | 0.035 |
| Sorghum | 119 | 342 | 275 | 70.4 | 0.018 |
| Starch Source | Jejunal Digestibility | Ileal Digestibility | Potential Digestible Starch (%) | Starch Digestion Rate (h−1) |
|---|---|---|---|---|
| Wheat | 0.882 | 0.944 | 93.9 | 2.51 |
| Corn | 0.888 | 0.969 | 96.8 | 2.55 |
| Sorghum | 0.837 | 0.953 | 95.4 | 1.81 |
| Barley | 0.898 | 0.981 | 98.5 | 2.51 |
| Peas | 0.574 | 0.804 | 85.3 | 1.03 |
| Potato starch | 0.198 | 0.329 | 38.4 | 0.55 |
7. Inclusions of Sulfite Reducing Agents
| SMBS Inclusion | Study/Sorghum | AME Response (MJ) | ME:GE Response (%) | AMEn Response (MJ) | FCR Response (%) |
|---|---|---|---|---|---|
| 1.25 | 1/#12 2 | 0.39 | 1.82 | 0.29 | 17.4 |
| 2.50 | 1/#12 2 | 0.17 | 0.54 | 0.08 | 5.67 |
| 5.00 | 1/#12 2 | 0.53 | 2.58 | 0.51 | 20.7 |
| 1.50 | 2/#13 | 0.36 | 2.22 | 0.45 | 2.47 |
| 2.25 | 2/#13 | 0.35 | 2.47 | 0.38 | 1.65 |
| 3.00 | 2/#13 | 0.29 | 1.73 | 0.31 | 2.26 |
| 3.75 | 2/#13 | 0.36 | 1.73 | 0.30 | 3.02 |
| 4.50 | 2/#13 | 0.40 | 2.47 | 0.51 | 2.33 |
| 5.25 | 2/#13 | 0.34 | 2.35 | 0.43 | 2.47 |
| 1.75 | 3/FW | −0.42 | −1.34 | −0.37 | 3.13 |
| 1.75 | 4/MP | 0.40 | 2.61 | 0.45 | 0.39 |
| 3.50 | 4/MP | 0.25 | 1.65 | 0.38 | −1.64 |
| 1.75 | 4/JM | 0.41 | 3.27 | 0.47 | −1.82 |
| 3.50 | 4/JM | 0.31 | 2.32 | 0.48 | −1.63 |
| 1.75 | 5/HP | 0.22 | 1.45 | 0.22 | 0.53 |
| 3.50 | 5/HP | 0.34 | 2.46 | 0.35 | 8.57 |
| 1.75 | 5/Tiger | 0.09 | 1.25 | 0.06 | −0.60 |
| 3.50 | 5/Tiger | 0.29 | 1.94 | 0.29 | 1.47 |
| 1.75 | 5/HFQ | 0.06 | 0.42 | 0.17 | 0.13 |
| 3.50 | 5/HFQ | 0.50 | 3.38 | 0.59 | 0.54 |
| (studies 2–5) | g/kg | ||||
| Mean | 2.84 | 0.27 MJ | 1.90% | 0.32 MJ | 1.37% |
| Median | 3.00 | 0.34 MJ | 2.22% | 0.38 MJ | 1.47% |
8. Responses of Exogenous Feed Enzymes
9. Innovative Assessments of Sorghum Quality
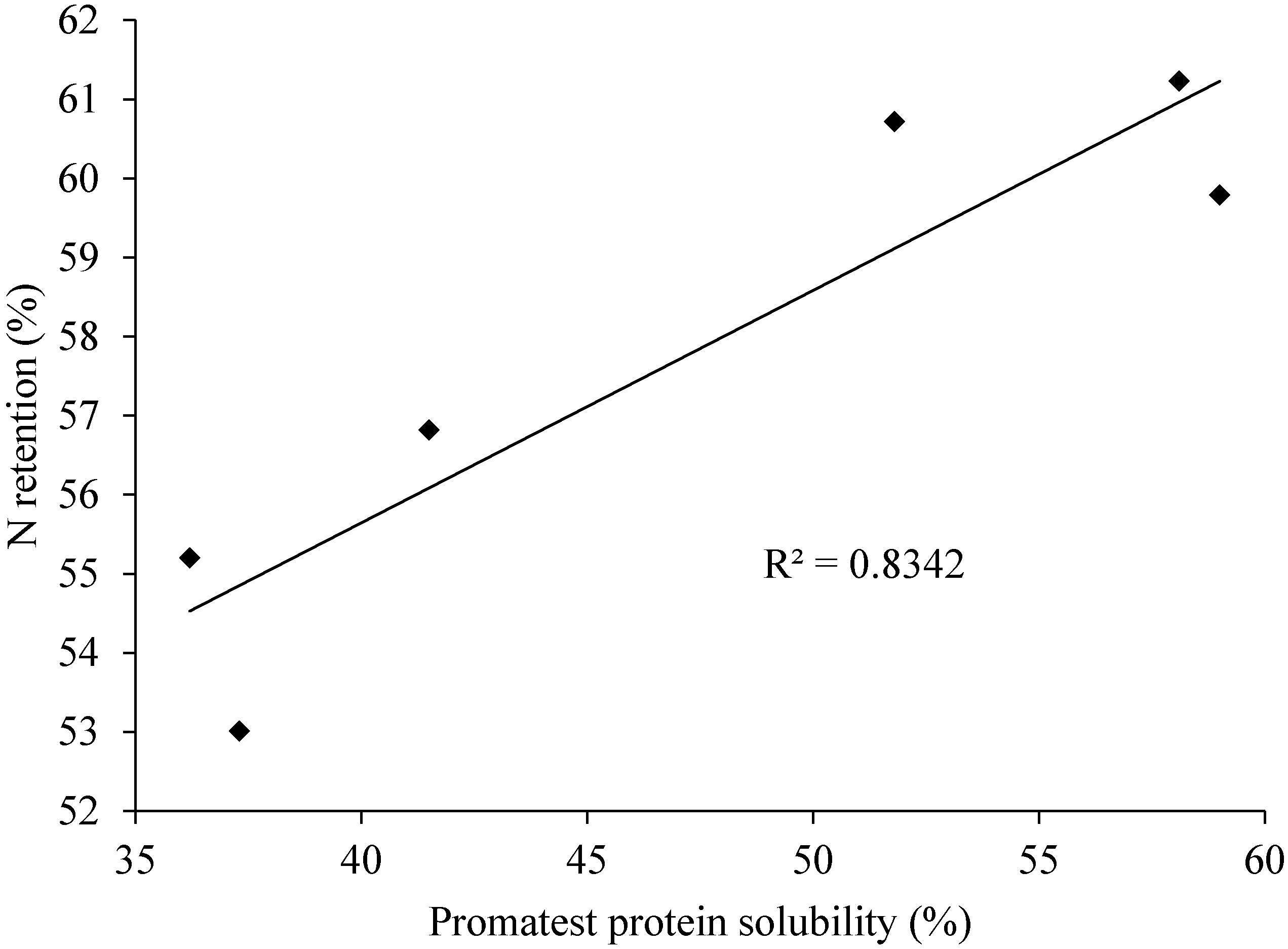
9.1. Promatest Protein Solubility
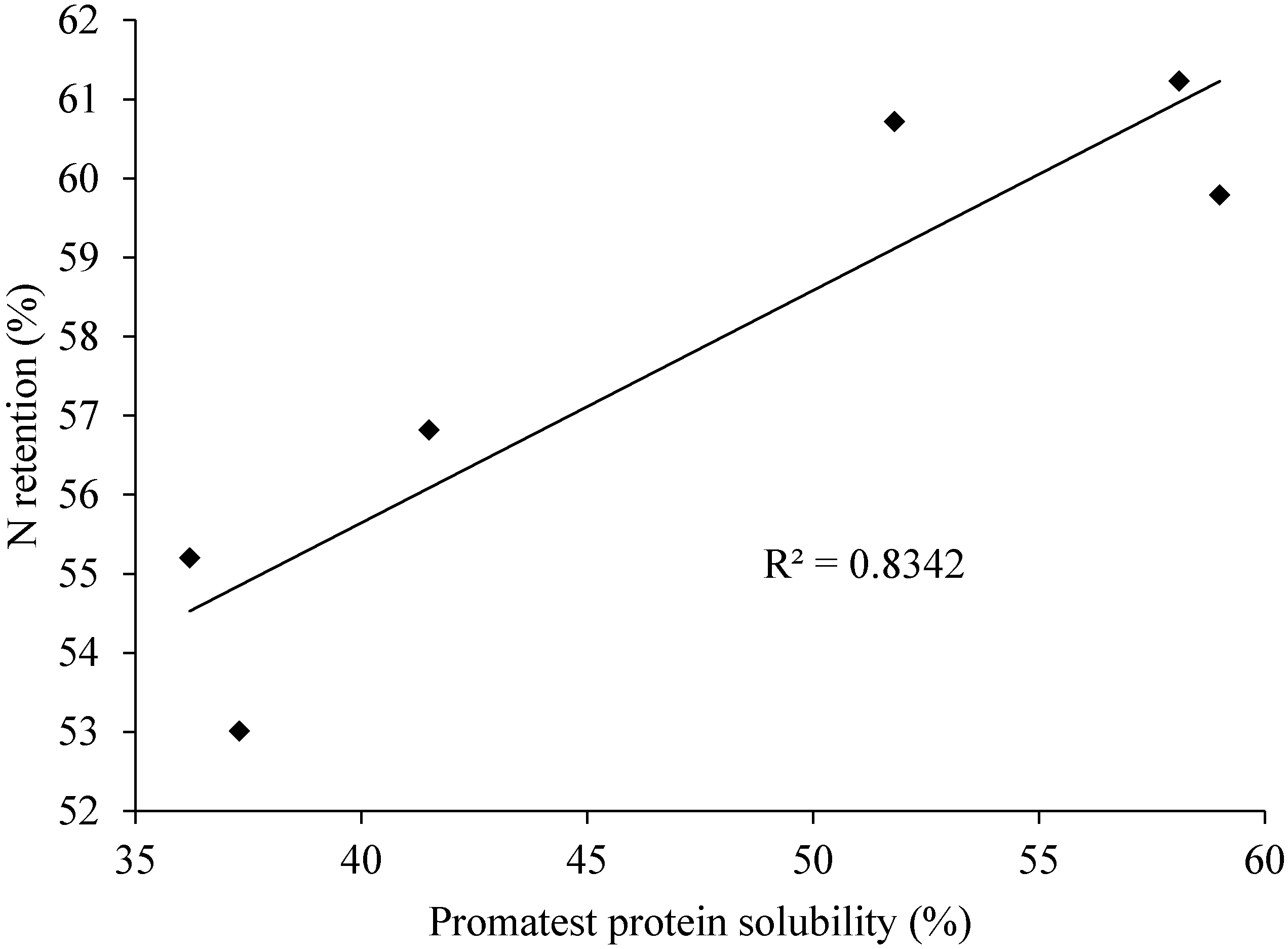
9.2. RVA Starch Pasting Profiles
| RVA Viscosity | Category | Parameter | Correlation Coefficient | Significance |
|---|---|---|---|---|
| Holding | Growth performance | FCR | r = −0.821 | p = 0.045 |
| Final | r = −0.832 | p = 0.040 | ||
| Peak | Nutrient | AME | r = 0.789 | p = 0.062 |
| Breakdown | utilisation | r = 0.774 | p = 0.071 | |
| Peak | ME:GE ratio | r = 0.800 | p = 0.056 | |
| Breakdown | r = 0.767 | p = 0.075 | ||
| Final | N retention | r = 0.880 | p = 0.021 | |
| Setback | r = 0.774 | p = 0.071 | ||
| Peak | AMEn | r = 0.835 | p = 0.039 | |
| Breakdown | r = 0.834 | p = 0.039 | ||
| Final | Protein (N) | PJ | r = 0.803 | p = 0.054 |
| Setback | digestibility | r = 0.896 | p = 0.016 | |
| Setback | DJ | r = 0.798 | p = 0.057 | |
| Setback | DI | r = 0.862 | p = 0.027 |
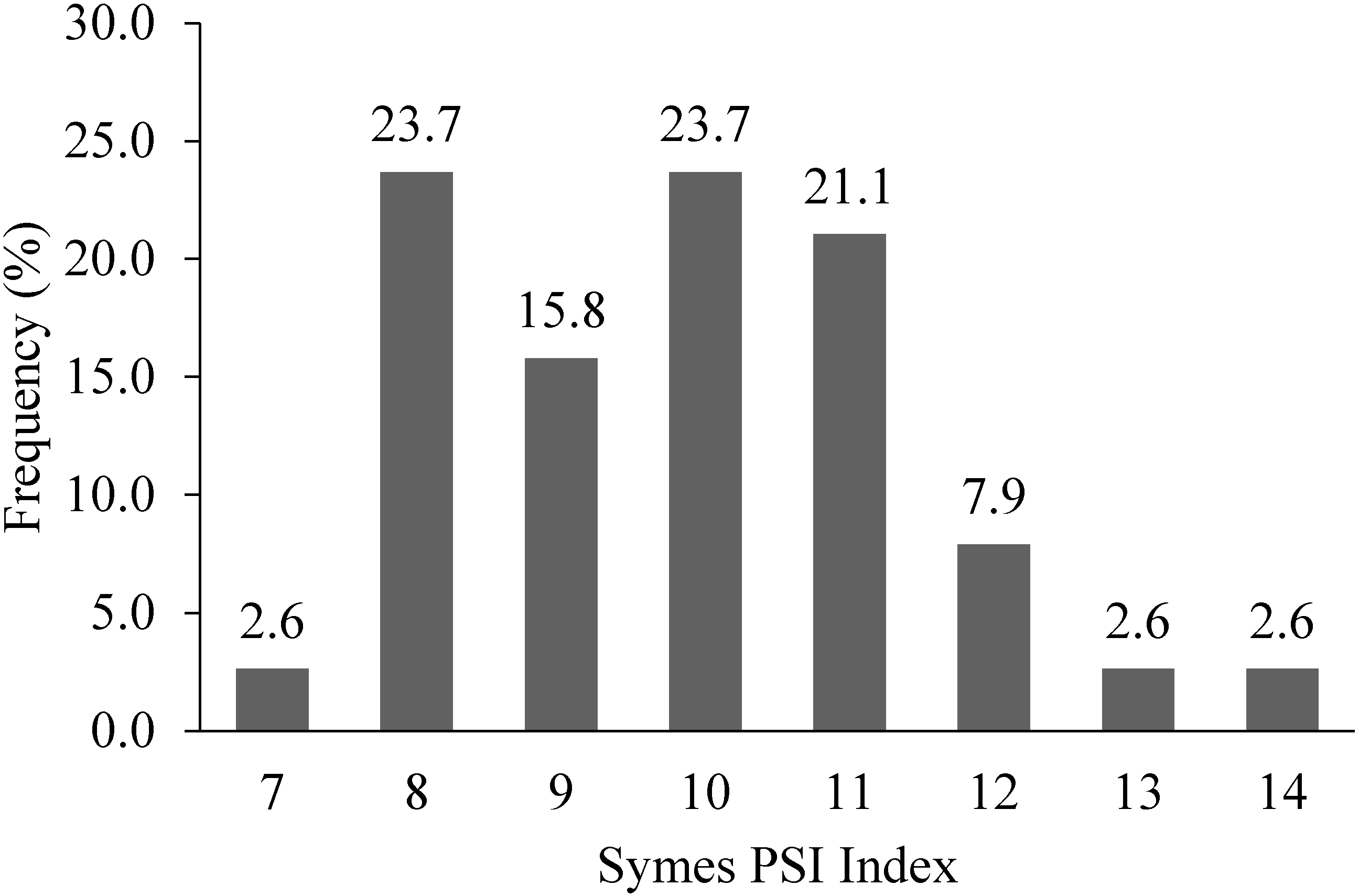
9.3. Grain Texture: Symes Particle Size Index (PSI)
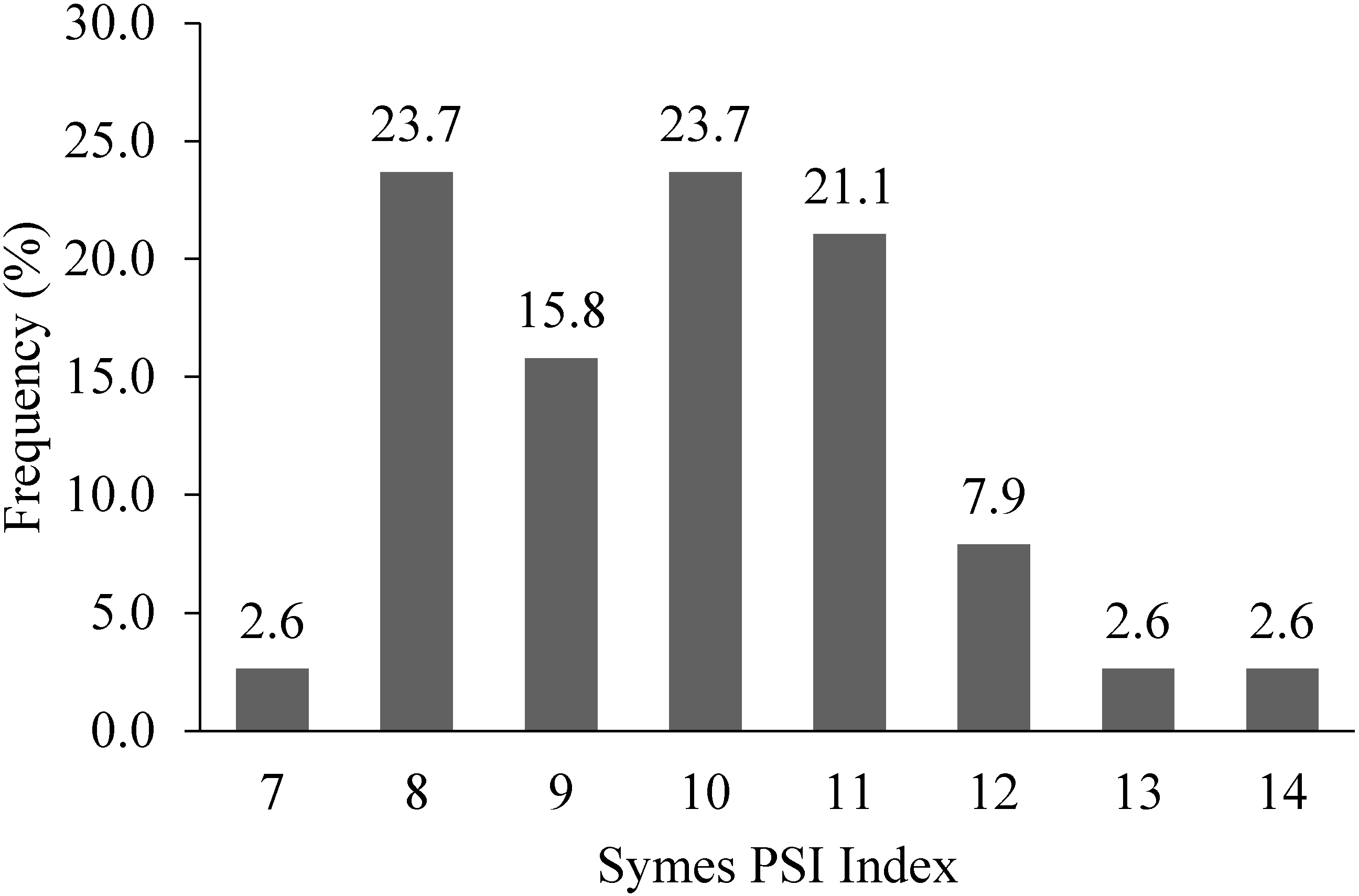
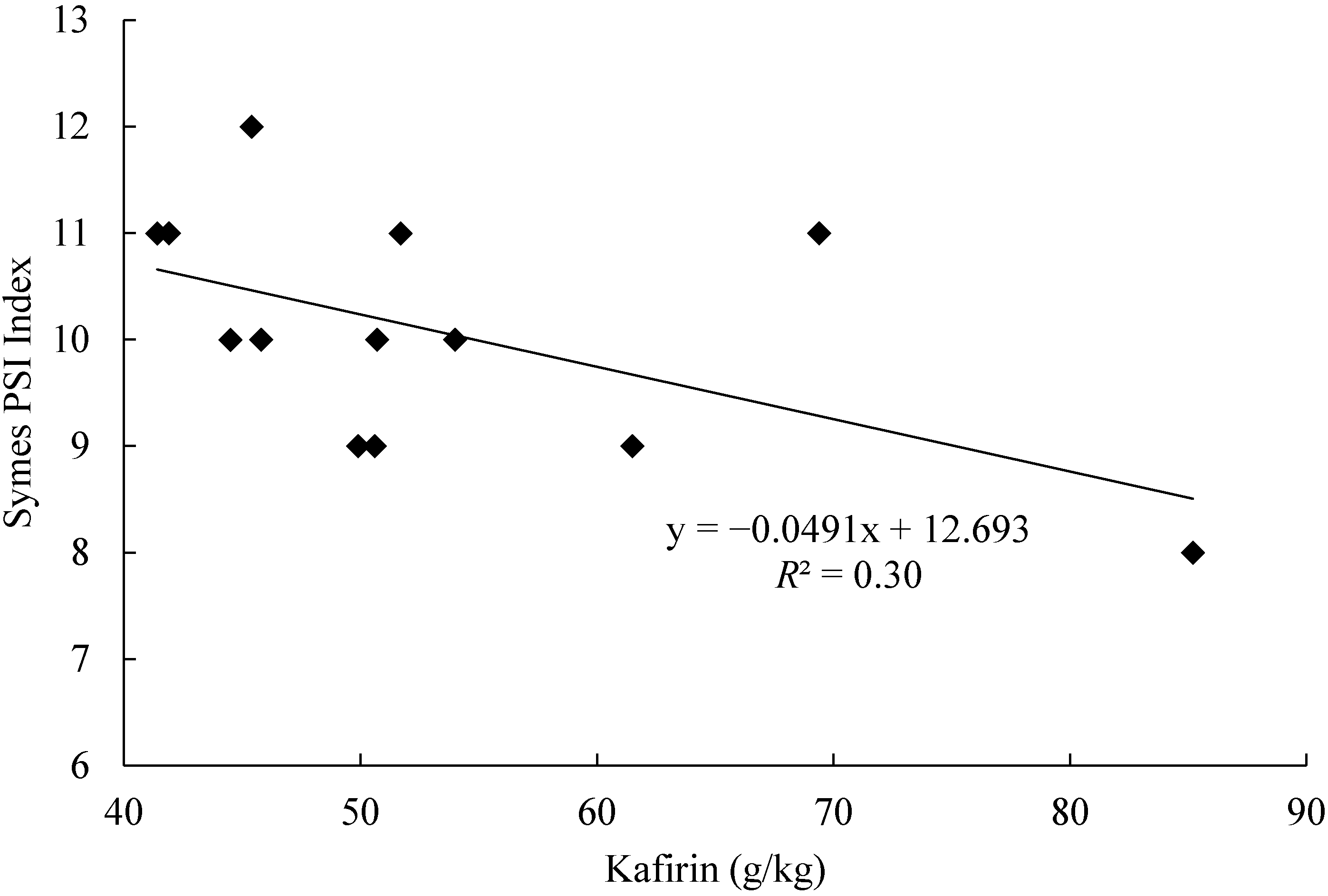
10. Implications
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Black, J.L.; Hughes, R.J.; Nielsen, S.G.; Tredrea, A.M.; MacAlpine, R.; van Barneveld, R.J. The energy value of cereal grains, particularly wheat and sorghum, for poultry. Proc. Aust. Poult. Sci. Symp. 2005, 17, 21–29. [Google Scholar]
- Selle, P.H. The protein quality of sorghum. Proc. Aust. Poult. Sci. Symp. 2011, 22, 147–160. [Google Scholar]
- Liu, S.Y.; Selle, P.H.; Cowieson, A.J. Strategies to enhance the performance of pigs and poultry on sorghum-based diets. Anim. Feed Sci. Technol. 2013, 181, 1–14. [Google Scholar] [CrossRef]
- Perez-Maldonado, R.A.; Rodrigues, H.D. Nutritional Characteristics of Sorghums from Queensland and New South Wales for Chicken Meat Production; RIRDC Publication No. 09/170; Rural Industries Research and Development Corporation: Barton, Australia, 2009. [Google Scholar]
- Bryden, W.L.; Liu, X.; Ravindran, G.; Hew, L.I.; Ravindran, V. Ileal digestible amino acids values in feedstuffs for poultry; RIRDC Publication No. 09/71; Rural Industries Research and Development Corporation: Barton, Australia, 2009. [Google Scholar]
- Waniska, R.D.; Hugo, L.F.; Rooney, L.W. Practical methods to determine the presence of tannins in sorghum. J. Appl. Poult. Res. 1992, 1, 122–128. [Google Scholar] [CrossRef]
- Liu, S.Y.; Cadogan, D.J.; Péron, A.; Truong, H.H.; Selle, P.H. Effects of phytase supplementation on growth performance, nutrient utilisation and digestive dynamics of starch and protein in broiler chickens offered maize-, sorghum- and wheat-based diets. Anim. Feed Sci. Technol. 2014, 197, 164–175. [Google Scholar] [CrossRef]
- Ao, Z.; Choct, M. Effect of early feeding and grain type on growth and performance of broilers. Proc. Aust. Poult. Sci. Symp. 2004, 16, 116–119. [Google Scholar]
- Selle, P.H.; Cadogan, D.J.; Ru, Y.J.; Partridge, G.G. Impact of exogenous enzymes in sorghum- or wheat-based broiler diets on nutrient utilization and growth performance. Int. J. Poult. Sci. 2010, 9, 53–58. [Google Scholar] [CrossRef]
- Bryden, W.L.; Selle, P.H.; Cadogan, D.J.; Liu, X.; Muller, N.D.; Jordan, D.R.; Gidley, M.J.; Hamilton, W.D. A review of the nutritive value of sorghum for broilers; RIRDC Publication No. 09/007; Rural Industries Research and Development Corporation: Barton, Australia, 2009. [Google Scholar]
- Selle, P.H.; Cadogan, D.J.; Li, X.; Bryden, W.L. Implications of sorghum in broiler chicken nutrition. Anim. Feed Sci. Technol. 2010, 156, 57–74. [Google Scholar] [CrossRef]
- Belton, P.S.; Delgadillo, I.; Halford, N.G.; Shewry, P.R. Kafirin structure and functionality. J. Cereal Sci. 2006, 44, 272–286. [Google Scholar] [CrossRef]
- Duodu, K.G.; Taylor, J.R.N.; Belton, P.S.; Hamaker, B.R. Factors affecting sorghum protein digestibility. J. Cereal Sci. 2003, 38, 117–131. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Chemistry and biochemistry of inositol polyphosphates. Rev. Pure Appl. Chem. 1966, 16, 209–224. [Google Scholar]
- Mosse, J.; Huet, J.C.; Baudet, J. The amino-acid composition of whole sorghum grain in relation to its nitrogen-content. Cereal Chem. 1988, 65, 271–277. [Google Scholar]
- Selle, P.H.; Cowieson, A.J.; Cowieson, N.P.; Ravindran, V. Protein-phytate interactions in pig and poultry nutrition: A reappraisal. Nutr. Res. Rev. 2012, 25, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Beta, T.; Corke, H. Effect of ferulic acid and catechin on sorghum and maize starch pasting properties. Cereal Chem. 2004, 81, 418–422. [Google Scholar] [CrossRef]
- Liu, S.Y.; Selle, P.H.; Cowieson, A.J. Influence of white- and red-sorghum varieties and hydrothermal component of steam-pelleting on digestibility coefficients of amino acids and kinetics of amino acids, nitrogen and starch digestion in diets for broiler chickens. Anim. Feed Sci. Technol. 2013, 186, 53–63. [Google Scholar] [CrossRef]
- Selle, P.H.; Walker, A.R.; Bryden, W.L. Total and phytate-phosphorus contents and phytase activity of australian-sourced feed ingredients for pigs and poultry. Aust. J. Exp. Agric. 2003, 43, 475–479. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41. [Google Scholar] [CrossRef]
- Truong, H.H.; Yu, S.; Peron, A.; Cadogan, D.J.; Khoddami, A.; Roberts, T.H.; Liu, S.Y.; Selle, P.H. Phytase supplementation of maize-, sorghum- and wheat-based broiler diets, with identified starch pasting properties, influences jejunal and ileal digestibilities of phytate (IP6) and sodium. Anim. Feed Sci. Technol. 2014, 198, 248–256. [Google Scholar] [CrossRef]
- Liu, S.Y.; Selle, P.H. A consideration of starch and protein digestive dynamics in chicken-meat production. Worlds Poult. Sci. J. 2015, 71, 297–310. [Google Scholar] [CrossRef]
- Giuberti, G.; Gallo, A.; Cerioli, C.; Masoero, F. In vitro starch digestion and predicted glycemic index of cereal grains commonly utilized in pig nutrition. Anim. Feed Sci. Technol. 2012, 174, 163–173. [Google Scholar] [CrossRef]
- Truong, H.H.; Liu, S.Y.; Selle, P.H. Starch utilisation in chicken-meat production: The foremost influential factors. Anim. Prod. Sci. 2015. [Google Scholar] [CrossRef]
- Sang, Y.J.; Bean, S.; Seib, P.A.; Pedersen, J.; Shi, Y.C. Structure and functional properties of sorghum starches differing in amylose content. J. Agric. Food Chem. 2008, 56, 6680–6685. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Lau, T.; Cai, N.; Singh, J.; Pedersen, J.F.; Vensel, W.H.; Hurkman, W.J.; Wilson, J.D.; Lemaux, P.G.; Buchanan, B.B. Digestibility of protein and starch from sorghum (Sorghum bicolor) is linked to biochemical and structural features of grain endosperm. J. Cereal Sci. 2009, 49, 73–82. [Google Scholar] [CrossRef]
- Taylor, J.R.N.; Emmambux, M.N. Developments in our understanding of sorghum polysaccharides and their health benefits. Cereal Chem. 2010, 87, 263–271. [Google Scholar] [CrossRef]
- Svihus, B.; Uhlen, A.K.; Harstad, O.M. Effect of starch granule structure, associated components and processing on nutritive value of cereal starch: A review. Anim. Feed Sci. Technol. 2005, 122, 303–320. [Google Scholar] [CrossRef]
- Ertl, D.; Dale, N. The metabolizable energy of waxy vs. Normal corn for poultry. J. Appl. Poult. Res. 1997, 6, 432–435. [Google Scholar] [CrossRef]
- Pirgozliev, V.R.; Rose, S.P.; Bedford, M.R. The effect of amylose: Amylopectin ratio in dietary starch on growth performance and gut morphology in broiler chickens. Arch. Fur Geflugelkunde 2010, 74, 21–29. [Google Scholar]
- Collins, N.E.; Moran, E.T.; Stilborn, H.L. Performance of broilers fed normal and waxy corn diets formulated with chick and rooster derived apparent metabolizable energy values for the grains. J. Appl. Poult. Res. 2003, 12, 196–206. [Google Scholar] [CrossRef]
- Gilding, E.K.; Frere, C.H.; Cruickshank, A.; Raba, A.K.; Prentis, P.J.; Mudge, A.M.; Mace, E.S.; Jordan, D.R.; Godwin, I.D. Allelic variation at a single gene increases food value in a drought-tolerant staple cereal. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed]
- Virupaksha, T.K.; Sastry, L.V.S. Studies on protein content and amino acid composition of some varieties of grain sorghum. J. Agric. Food Chem. 1968, 16, 199–203. [Google Scholar] [CrossRef]
- Taylor, J.R.N.; Schussler, L.; Vanderwalt, W.H. Fractionation of proteins from low-tannin sorghum grain. J. Agric. Food Chem. 1984, 32, 149–154. [Google Scholar] [PubMed]
- Selle, P.H.; Liu, S.Y.; Cowieson, A.J. Sorghum: An enigmatic grain for chicken-meat production. In Sorghum: Production, Growth Habits and Health Benefits; Parra, P.C., Ed.; Nova Publishers Inc.: Hauppauge, NY, USA, 2013; pp. 1–44. [Google Scholar]
- Henzell, R.G. Grain sorghum breeding in Australia: Current status and future prospects. In Proceedings of the 2nd Australian Sorghum Conference, Gatton, Australia, 4–6 February 1992; pp. 70–80.
- Symes, K.J. Inheritance of grain hardness in wheat as measured by particle size index. Aust. J. Agric. Res. 1965, 16, 113–123. [Google Scholar] [CrossRef]
- De Alencar Figueiredo, L.F.; Davrieux, F.; Fliedel, G.; Rami, J.F.; Chantereau, J.; Deu, M.; Courtois, B.; Mestres, C. Development of nirs equations for food grain quality traits through exploitation of a core collection of cultivated sorghum. J. Agric. Food Chem. 2006, 54, 8501–8509. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.R.; Hancock, J.D.; Bramel-Cox, P.J.; Hines, R.H.; Behnke, K.C. Effects of corn, sorghum genotype, and particle size on milling characteristics and growth performance in broiler chicks. Poult. Sci. 1994, 73, 11. [Google Scholar]
- Ravindran, V.; Hew, L.I.; Bryden, W.L. Digestible amino acids in poultry feedstuffs; RIRDC Publication No. 98/9; Rural Industries Research and Development Corporation: Barton, Australia, 1998. [Google Scholar]
- Selle, P.H.; Gill, R.J.; Downing, J.A. The vulnerability of sorghum to “moist-heat”. Proc. Aust. Poult. Sci. Symp. 2010, 21, 68–71. [Google Scholar]
- Gidley, M.J.; Flanagan, B.M.; Sharpe, K.; Sopade, P.A. Starch digestion in monogastrics—Mechanisms and opportunities. Proc. Recent Adv. Anim. Nutr. Aust. 2011, 18, 207–213. [Google Scholar]
- Rooney, L.W.; Pflugfelder, R.L. Factors affecting starch digestibility with special emphasis on sorghum and corn. J. Anim. Sci. 1986, 63, 1607–1623. [Google Scholar] [PubMed]
- Taylor, J.R.N. Non-starch polysaccharides, protein and starch: Form function and feed-highlight on sorghum. Proc. Aust. Poult. Sci. Symp. 2005, 17, 9–16. [Google Scholar]
- De Mesa-Stonestreet, N.J.; Alavi, S.; Bean, S.R. Sorghum proteins: The concentration, isolation, modification, and food applications of kafirins. J. Food Sci. 2010, 75, R90–R104. [Google Scholar] [CrossRef] [PubMed]
- Ezeogu, L.I.; Duodu, K.G.; Taylor, J.R.N. Effects of endosperm texture and cooking conditions on the in vitro starch digestibility of sorghum and maize flours. J. Cereal Sci. 2005, 42, 33–44. [Google Scholar] [CrossRef]
- Truong, H.H.; Neilson, K.A.; McInerney, B.V.; Khoddami, A.; Roberts, T.H.; Liu, S.Y.; Selle, P.H. Performance of broiler chickens offered nutritionally-equivalent diets based on two red grain sorghums with quantified kafirin concentrations as intact pellets or re-ground mash following steam-pelleting at 65 or 97 °C conditioning temperatures. Anim. Nutr. 2015, in press. [Google Scholar] [CrossRef]
- Selle, P.H.; Liu, S.Y.; Cai, J.; Cowieson, A.J. Steam-pelleting temperatures, grain variety, feed form and protease supplementation of mediumly-ground, sorghum-based broiler diets: Influences on growth performance, relative gizzard weights, nutrient utilisation, starch and nitrogen digestibility. Anim. Prod. Sci. 2013, 53, 378–387. [Google Scholar] [CrossRef]
- Liu, S.Y.; Selle, P.H.; Court, S.G.; Cowieson, A.J. Protease supplementation of sorghum-based broiler diets enhances amino acid digestibility coefficients in four small intestinal sites and accelerates their rates of digestion. Anim. Feed Sci. Technol. 2013, 183, 175–183. [Google Scholar] [CrossRef]
- Mangan, J.L. Nutritional effects of tannins in animal feeds. Nutr. Res. Rev. 1988, 1, 209–231. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Nyachoti, C.M.; Atkinson, J.L.; Leeson, S. Sorghum tannins: A review. Worlds Poult. Sci. J. 1997, 53, 5–21. [Google Scholar] [CrossRef]
- McClymont, G.L.; Duncan, D.C. Studies on nutrition of poultry. III. Toxicity of grain sorghum for chickens. Ausi. Vet. J. 1952, 28, 229–233. [Google Scholar] [CrossRef]
- Asquith, T.N.; Izuno, C.C.; Butler, L.G. Characterization of the condensed tannin (proanthocyanidin) from a group II sorghum. J. Agric. Food Chem. 1983, 31, 1299–1303. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L.W. Sorghum and millet phenols and antioxidants. J. Cereal Sci. 2006, 44, 236–251. [Google Scholar] [CrossRef]
- Thompson, L.U.; Yoon, J.H. Starch digestibility as affected by polyphenols and phytic acid. J. Food Sci. 1984, 49, 1228–1229. [Google Scholar] [CrossRef]
- Thompson, L.U.; Yoon, J.H.; Jenkins, D.J.A.; Wolever, T.M.S.; Jenkins, A.L. Relationship between polyphenol intake and blood-glucose response of normal and diabetic individuals. Am. J. Clin. Nutr. 1984, 39, 745–751. [Google Scholar] [PubMed]
- Awika, J.M.; Dykes, L.; Gu, L.W.; Rooney, L.W.; Prior, R.L. Processing of sorghum (Sorghum bicolor) and sorghum products alters procyanidin oligomer and polymer distribution and content. J. Agric. Food Chem. 2003, 51, 5516–5521. [Google Scholar] [CrossRef] [PubMed]
- Dlamini, N.R.; Taylor, J.R.N.; Rooney, L.W. The effect of sorghum type and processing on the antioxidant properties of African sorghum-based foods. Food Chem. 2007, 105, 1412–1419. [Google Scholar] [CrossRef]
- Zhu, F. Interactions between starch and phenolic compound. Trends Food Sci. Technol. 2015, 43, 129–143. [Google Scholar] [CrossRef]
- Tomasik, P.; Schilling, C.H. Complexes of starch with inorganic guests. Adv. Carbohydr. Chem. Biochem. 1998, 53, 263–343. [Google Scholar]
- Barros, F.; Awika, J.M.; Rooney, L.W. Interaction of tannins and other sorghum phenolic compounds with starch and effects on in vitro starch digestibility. J. Agric. Food Chem. 2012, 60, 11609–11617. [Google Scholar] [CrossRef] [PubMed]
- Kandil, A.; Li, J.H.; Vasanthan, T.; Bressler, D.C. Phenolic acids in some cereal grains and their inhibitory effect on starch liquefaction and saccharification. J. Agric. Food Chem. 2012, 60, 8444–8449. [Google Scholar] [CrossRef] [PubMed]
- Welsch, C.A.; Lachance, P.A.; Wasserman, B.P. Dietary phenolic-compounds—Inhibition of Na+-dependent d-glucose uptake in rat intestinal brush-border membrane-vesicles. J. Nutr. 1989, 119, 1698–1704. [Google Scholar] [PubMed]
- Elkin, R.G.; Freed, M.B.; Hamaker, B.R.; Zhang, Y.; Parsons, C.M. Condensed tannins are only partially responsible for variations in nutrient digestibilities of sorghum grain cultivars. J. Agric. Food Chem. 1996, 44, 848–853. [Google Scholar] [CrossRef]
- Beta, T.; Corke, H. Genetic and environmental variation in sorghum starch properties. J. Cereal Sci. 2001, 34, 261–268. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L.W.; Waniska, R.D.; Rooney, W.L. Phenolic compounds and antioxidant activity of sorghum grains of varying genotypes. J. Agric. Food Chem. 2005, 53, 6813–6818. [Google Scholar] [CrossRef] [PubMed]
- Selle, P.H.; Ravindran, V.; Caldwell, R.A.; Bryden, W.L. Phytate and phytase: Consequences for protein utilisation. Nutr. Res. Rev. 2000, 13, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Selle, P.H.; Cowieson, A.J.; Ravindran, V. Consequences of calcium interactions with phytate and phytase for poultry and pigs. Livest. Sci. 2009, 124, 126–141. [Google Scholar] [CrossRef]
- Doherty, C.; Faubion, J.M.; Rooney, L.W. Semiautomated determination of phytate in sorghum and sorghum products. Cereal Chem. 1982, 59, 373–378. [Google Scholar]
- Selle, P.H.; Ravindran, V.; Cowieson, A.J.; Bedford, M.R. Phytate and Phytase; CABI Publishing: Oxon, UK, 2010; pp. 160–205. [Google Scholar]
- Oatway, L.; Vasanthan, T.; Helm, J.H. Phytic acid. Food Rev. Int. 2001, 17, 419–431. [Google Scholar] [CrossRef]
- Ravindran, V.; Sands, J.S.; Hruby, M.; Partridge, G.G. Influence of Phyzyme tm xp Phytase on Nutrient Utilization in Broilers Fed Diets Containing Graded Levels of Phytate; Poultry Research Foundation: Sydney, Australia, 2005; pp. 238–241. [Google Scholar]
- Wu, Y.B.; Ravindran, V.; Hendriks, W.H. Influence off exogenous enzyme supplementation on energy utilisation and nutrient digestibility off cereals for broilers. J. Sci. Food Agric. 2004, 84, 1817–1822. [Google Scholar] [CrossRef]
- Csonka, F.A.; Murphy, J.C.; Jones, D.B. The iso-electric points of various proteins. J. Am. Chem. Soc. 1926, 48, 763–768. [Google Scholar] [CrossRef]
- Kreydiyyeh, S.I.; Baydoun, E.A.H.; Churukian, Z.M. Tea extract inhibits intestinal-absorption of glucose and sodium in rats. Comp. Biochem. Physiol. C-Pharmacol. Toxicol. Endocrinol. 1994, 108, 359–365. [Google Scholar] [CrossRef]
- Kreydiyyeh, S.I. Inhibitors in tea of intestinal absorption of phenylalanine in rats. Comp. Biochem. Physiol. C-Pharmacol. Toxicol. Endocrinol. 1996, 113, 67–71. [Google Scholar] [CrossRef]
- Liu, N.; Ru, Y.J.; Li, F.D.; Cowieson, A.J. Effect of diet containing phytate and phytase on the activity and messenger ribonucleic acid expression of carbohydrase and transporter in chickens. J. Anim. Sci. 2008, 86, 3432–3439. [Google Scholar] [CrossRef] [PubMed]
- Weurding, R.E.; Enting, H.; Verstegen, M.W.A. The relation between starch digestion rate and amino acid level for broiler chickens. Poult. Sci. 2003, 82, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Li, T.J.; Dai, Q.Z.; Yin, Y.L.; Zhang, J.; Huang, R.L.; Ruan, Z.; Deng, Z.; Xie, M. Dietary starch sources affect net portal appearance of amino acids and glucose in growing pigs. Animal 2008, 2, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Enting, H.; Pos, J.; Weurding, R.E.; Veldman, A. Starch digestion rate affects broiler performance. Proc. Aust. Poult. Sci. Symp. 2005, 17, 17–20. [Google Scholar]
- Weurding, R.E.; Veldman, A.; Veen, W.A.G.; van der Aar, P.J.; Verstegen, M.W.A. Starch digestion rate in the small intestine of broiler chickens differs among feedstuffs. J. Nutr. 2001, 131, 2329–2335. [Google Scholar] [PubMed]
- Liu, S.Y.; Selle, P.H.; Cowieson, A.J. The kinetics of starch and nitrogen digestion regulate growth performance and nutrient utilisation in coarsely-ground, sorghum-based broiler diets. Anim. Prod. Sci. 2013, 53, 1033–1040. [Google Scholar] [CrossRef]
- Van den Borne, J.; Schrama, J.W.; Heetkamp, M.J.W.; Verstegen, M.W.A.; Gerrits, W.J.J. Synchronising the availability of amino acids and glucose increases protein retention in pigs. Animal 2007, 1, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Axtell, J.D.; Kirleis, A.W.; Hassen, M.M.; Mason, N.D.; Mertz, E.T.; Munck, L. Digestibility of sorghum proteins. Proc. Natl. Acad. Sci. USA Biol. Sci. 1981, 78, 1333–1335. [Google Scholar] [CrossRef]
- Choi, S.J.; Woo, H.D.; Ko, S.H.; Moon, T.W. Confocal laser scanning microscopy to investigate the effect of cooking and sodium bisulfate on in vitro digestibility of waxy sorghum flour. Cereal Chem. 2008, 85, 65–69. [Google Scholar] [CrossRef]
- Batterman-Azcona, S.J.; Hamaker, B.R. Changes occurring in protein body structure and alpha-zein during cornflake processing. Cereal Chem. 1998, 75, 217–221. [Google Scholar] [CrossRef]
- Duodu, K.G.; Nunes, A.; Delgadillo, I.; Parker, M.L.; Mills, E.N.C.; Belton, P.S.; Taylor, J.R.N. Effect of grain structure and cooking on sorghum and maize in vitro protein digestibility. J. Cereal Sci. 2002, 35, 161–174. [Google Scholar] [CrossRef]
- Hamaker, B.R.; Kirleis, A.W.; Butler, L.G.; Axtell, J.D.; Mertz, E.T. Improving the in vitro protein digestibility of sorghum with reduing agents. Proc. Natl. Acad. Sci. USA 1987, 84, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Rom, D.L.; Shull, J.M.; Chandrashekar, A.; Kirleis, A.W. Effects of cooking and treatment with sodium bisulfite on in vitro protein digestibility and microstructure of sorghum flour. Cereal Chem. 1992, 69, 178–181. [Google Scholar]
- Selle, P.H.; Liu, S.Y.; Cai, J.; Caldwell, R.A.; Cowiesona, A.J. Preliminary assessment of including a reducing agent (sodium metabisulphite) in “all-sorghum” diets for broiler chickens. Anim. Feed Sci. Technol. 2013, 186, 81–90. [Google Scholar] [CrossRef]
- Selle, P.H.; Liu, S.Y.; Cai, J.; Caldwell, R.A.; Cowieson, A.J. Graded inclusions of sodium metabisulphite in sorghum-based diets: I. Reduction of disulphide cross-linkages in vitro and enhancement of energy utilisation and feed conversion efficiency in broiler chickens. Anim. Feed Sci. Technol. 2014, 190, 59–67. [Google Scholar] [CrossRef]
- Truong, H.H.; Cadogan, D.J.; Liu, S.Y.; Selle, P.H. Addition of sodium metabisulfite and microbial phytase, individually and in combination, to a sorghum-based diet for broiler chickens from 7 to 28 days post-hatch. Anim. Prod. Sci. 2015. [Google Scholar] [CrossRef]
- Paterson, L.; Mitchell, J.R.; Hill, S.E.; Blanshard, J.M.V. Evidence for sulfite induced oxidative reductive depolymerisation of starch polysaccharides. Carbohydr. Res. 1996, 292, 143–151. [Google Scholar] [CrossRef]
- Paterson, L.A.; Hill, S.E.; Mitchell, J.R.; Blanshard, J.M.V. Sulphite and oxidative-reductive depolymerization reactions. Food Chem. 1997, 60, 143–147. [Google Scholar] [CrossRef]
- Ravindran, V.; Cabahug, S.; Ravindran, G.; Bryden, W.L. Influence of microbial phytase on apparent ileal amino acid digestibility of feedstuffs for broilers. Poult. Sci. 1999, 78, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Cadogan, D.J.; Péron, A.; Truong, H.H.; Selle, P.H. A combination of xylanase, amylase and protease influences growth performance, nutrient utilisation, starch and protein digestive dynamics in broiler chickens offered maize-, sorghum- and wheat-based diets. Anim. Prod. Sci. 2014. [Google Scholar] [CrossRef]
- Odjo, S.; Malumba, P.; Dossou, J.; Janas, S.; Bera, F. Influence of drying and hydrothermal treatment of corn on the denaturation of salt-soluble proteins and color parameters. J. Food Eng. 2012, 109, 561–570. [Google Scholar] [CrossRef]
- Gehring, C.K.; Bedford, M.R.; Cowieson, A.J.; Dozier, W.A. Effects of corn source on the relationship between in vitro assays and ileal nutrient digestibility. Poult. Sci. 2012, 91, 1908–1914. [Google Scholar] [CrossRef] [PubMed]
- Selle, P.H.; Liu, S.Y.; Cai, J.; Cowieson, A.J. Steam-pelleting and feed form of broiler diets based on three coarsely ground sorghums influences growth performance, nutrient utilisation, starch and nitrogen digestibility. Anim. Prod. Sci. 2012, 52, 842–852. [Google Scholar] [CrossRef]
- Zhang, G.Y.; Hamaker, B.R. Sorghum (Sorghum bicolor L. Moench) flour pasting properties influenced by free fatty acids and protein. Cereal Chem. 2005, 82, 534–540. [Google Scholar] [CrossRef]
- Liu, S.Y.; Selle, P.H.; Khoddami, A.; Roberts, T.H.; Cowieson, A.J. Graded inclusions of sodium metabisulphite in sorghum-based diets: II. Modification of starch pasting properties in vitro and beneficial impacts on starch digestion dynamics in broiler chickens. Anim. Feed Sci. Technol. 2014, 190, 68–78. [Google Scholar] [CrossRef]
- Selle, P.H.; Liu, S.Y.; Cai, J.; Cowieson, A.J. Steam-pelleting temperatures and grain variety of finely-ground, sorghum-based broiler diets. I. Influence on growth performance, relative gizzard weights, nutrient utilisation, starch and nitrogen digestibility. Anim. Prod. Sci. 2014, 54, 339–346. [Google Scholar] [CrossRef]
- Chiremba, C.; Taylor, J.R.N.; Rooney, L.W.; Beta, T. Phenolic acid content of sorghum and maize cultivars varying in hardness. Food Chem. 2012, 134, 81–88. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.Y.; Fox, G.; Khoddami, A.; Neilson, K.A.; Truong, H.H.; Moss, A.F.; Selle, P.H. Grain Sorghum: A Conundrum for Chicken-Meat Production. Agriculture 2015, 5, 1224-1251. https://doi.org/10.3390/agriculture5041224
Liu SY, Fox G, Khoddami A, Neilson KA, Truong HH, Moss AF, Selle PH. Grain Sorghum: A Conundrum for Chicken-Meat Production. Agriculture. 2015; 5(4):1224-1251. https://doi.org/10.3390/agriculture5041224
Chicago/Turabian StyleLiu, Sonia Y., Glen Fox, Ali Khoddami, Karlie A. Neilson, Ha H. Truong, Amy F. Moss, and Peter H. Selle. 2015. "Grain Sorghum: A Conundrum for Chicken-Meat Production" Agriculture 5, no. 4: 1224-1251. https://doi.org/10.3390/agriculture5041224
APA StyleLiu, S. Y., Fox, G., Khoddami, A., Neilson, K. A., Truong, H. H., Moss, A. F., & Selle, P. H. (2015). Grain Sorghum: A Conundrum for Chicken-Meat Production. Agriculture, 5(4), 1224-1251. https://doi.org/10.3390/agriculture5041224