
Mycotoxins: Producing Fungi and Mechanisms of Phytotoxicity



Abstract
:1. Introduction

| Name | Abbreviation | Species/Genus/Group | Chemical Group |
|---|---|---|---|
| citreoviridin | CIV | Penicillium citreo-viride | pyranone derivative |
| citrinin | CTN | Penicillium, Monascus, Aspergillus terreus | benzopyran compound |
| cyclopiazonic acid | CPA | Penicillium, Aspergillus | indole tetramic acid |
| frequentin | FRE | P. frequentans | carbocyclic compound |
| gliotoxin | GT | A. fumigatus | epipolythiodioxopiperazine |
| mycophenolic acid | MPA | Penicillium | meroterpenoid compound |
| ochratoxin A | OTA | Aspergillus, Penicillium | benzopyran compound |
| palitantin | PAL | Penicillium | cyclohexane derivative |
| patulin | PAT | Penicillium, Aspergillus | benzopyran compound |
| penicillic acid | PA | Penicillium, Aspergillus | isopropylidene tetronic acid |
| penitrem A | PNT | Penicillium | indole diterpene alkaloid |
| roquefortine | RQF | Penicillium | diketopiperazine compound |
| rubratoxin B | RB | Penicillium rubrum | alpha, beta unsaturated lactone |
| viomellein | VIM | Penicillium | benzopyran compound |
| 3-acetyldeoxynivalenol | 3-ADON | Fusarium | trichothecenes, sesquiterpenoid |
| 4-deoxynivalenol | DON | Fusarium | trichothecenes, sesquiterpenoid |
| beauvericin | BEA | Fusarium | hexadepsipeptide compound |
| diacetoxyscirpeno | DAS | Fusarium | trichothecenes, sesquiterpenoid |
| enniatin | ENN | Fusarium | cyclic depsipeptide |
| fumonisins | e.g., FB1 | Fusarium | monoterpenes |
| fusaric acid | FA | Fusarium | picolinic acid derivative, carboxylic acid |
| HT-2 toxin | - | Fusarium | trichothecenes, sesquiterpenoid |
| moniliformin | MON | Fusarium | cyclobutane compound, dione |
| nivalenol | NIV | Fusarium | trichothecenes, sesquiterpenoid |
| T-2 toxin | - | Fusarium | trichothecenes, sesquiterpenoid |
| zearalenone/F-2 toxin | ZEA | Fusarium | estrogenic compound |
| aflatoxin | e.g., AFB1 | Aspergillus | difuranocoumarin derivative |
| tenuazonic acid | TA | Alternaria tenuis | 3-acetyltetramic acid |
| cytochalasin | e.g., CB | Phoma | polyketide-amino acid hybrid |
2. Mycotoxins Produced by Aspergillus and Penicillium spp.
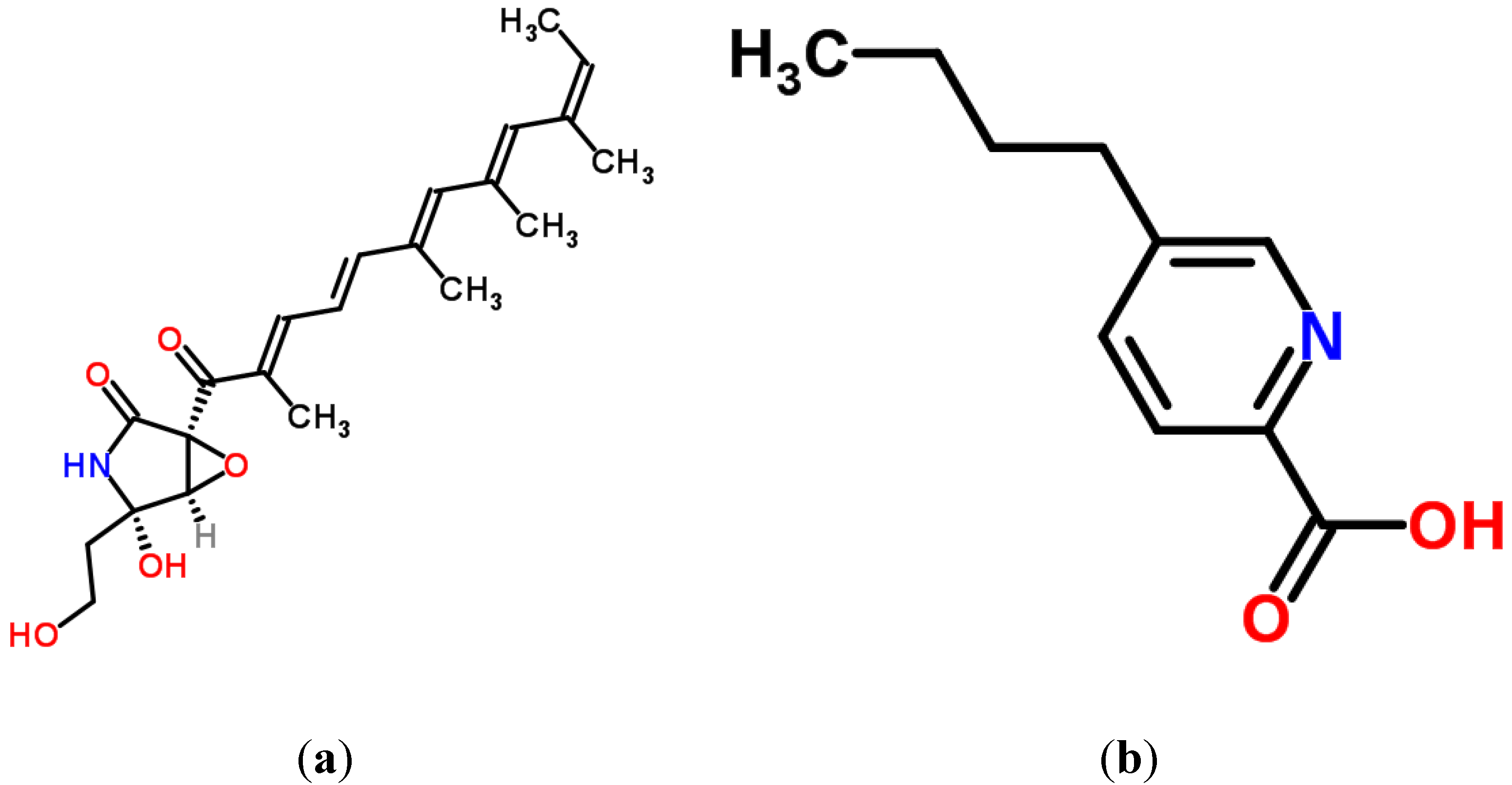
2.1. Aflatoxins
2.1.1. Historical Perspective
2.1.2. Producing Fungal Species
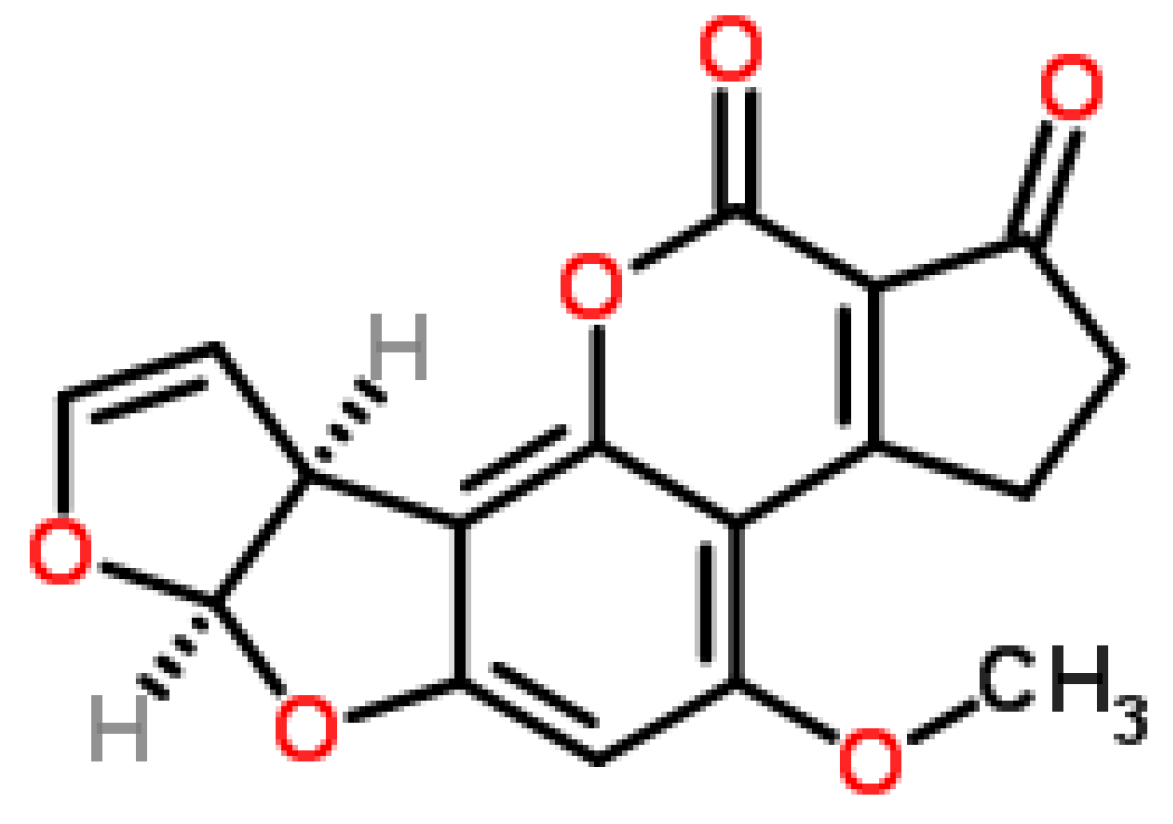
2.1.3. Phytotoxicity
2.1.4. Mechanism of Action
2.2. Citrinin
2.2.1. Historical Perspective and Producing Fungal Species
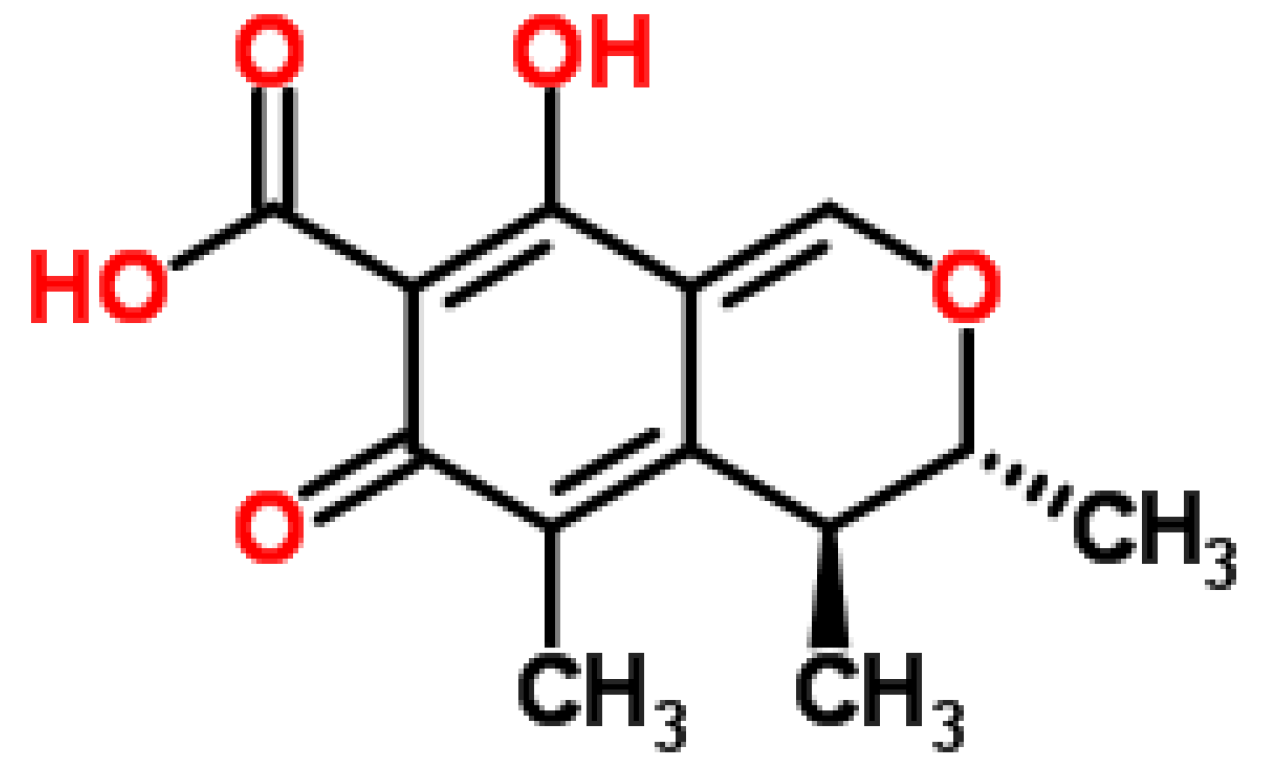
2.2.2. Phytotoxicity
2.2.3. Mechanism of Action
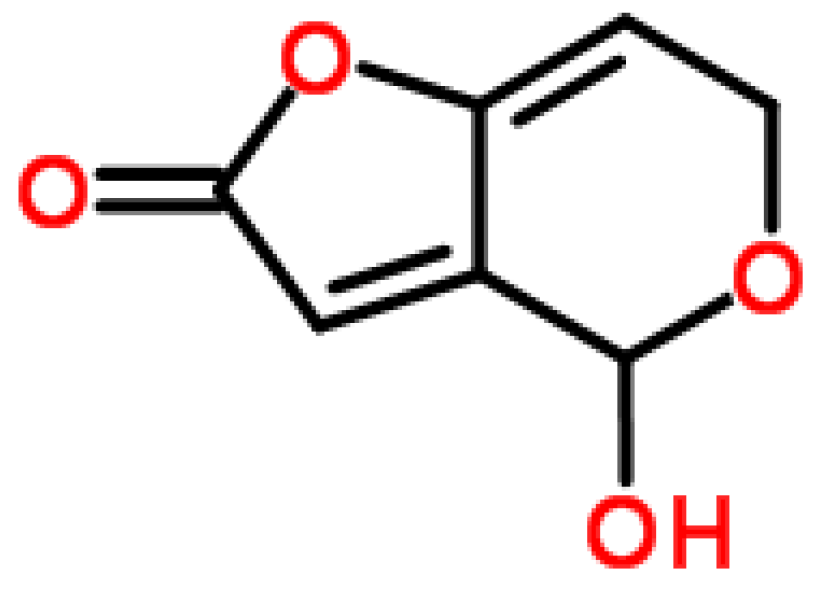
2.3. Patulin
2.3.1. Historical Perspective
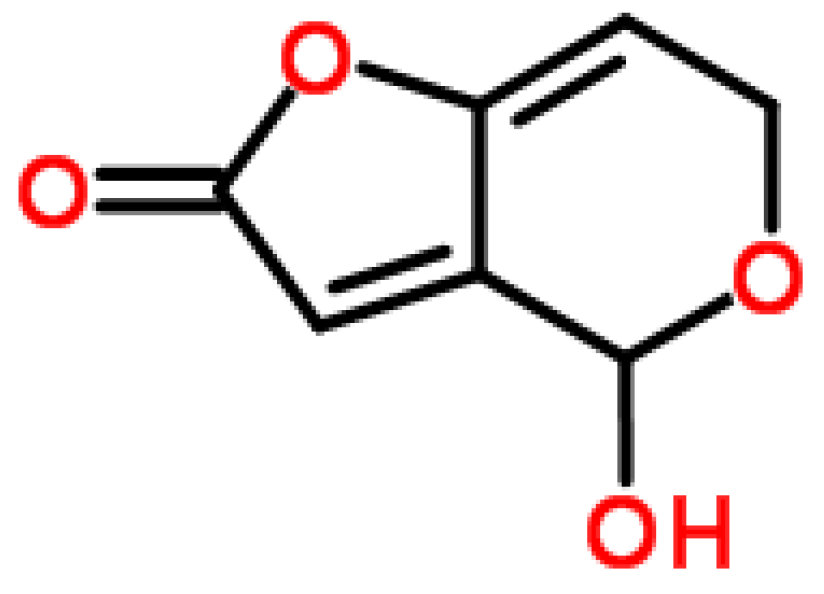
2.3.2. Producing Fungal Species
2.3.3. Phytotoxicity
2.3.4. Mechanism of Action
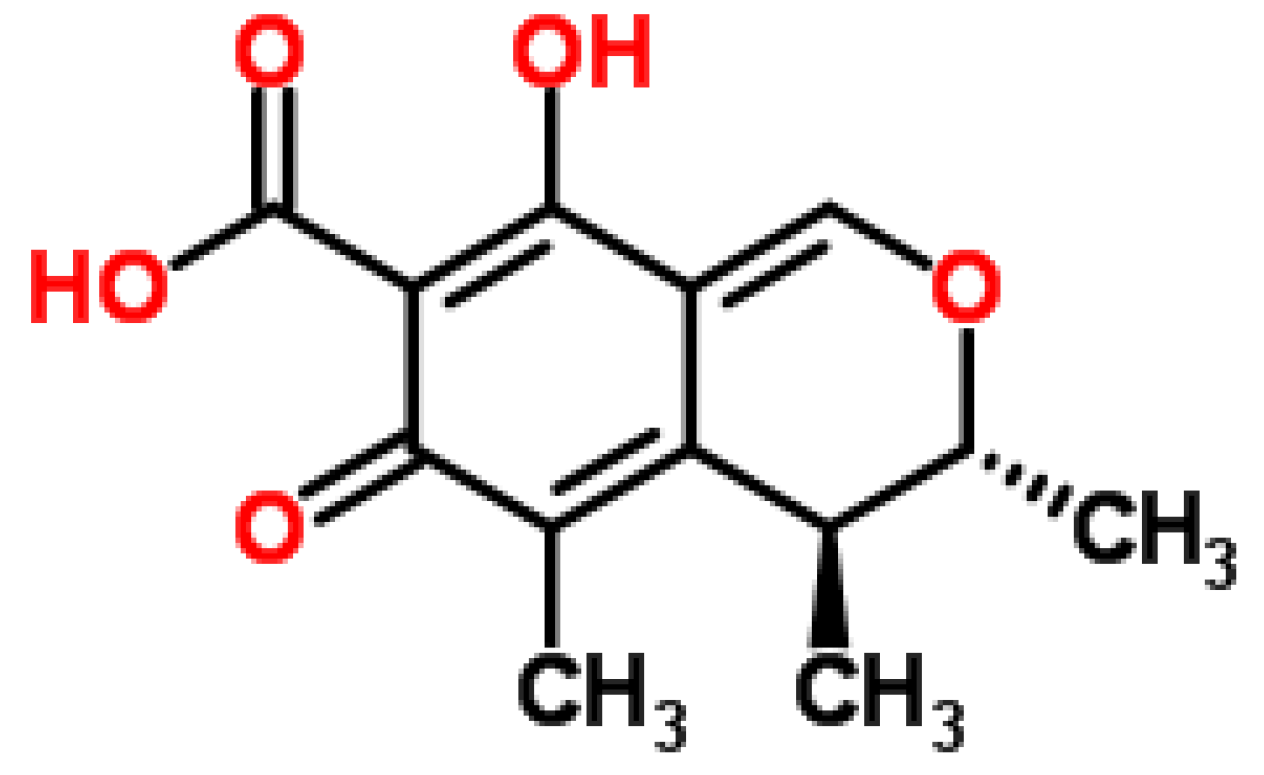
2.4. Penicillic Acid
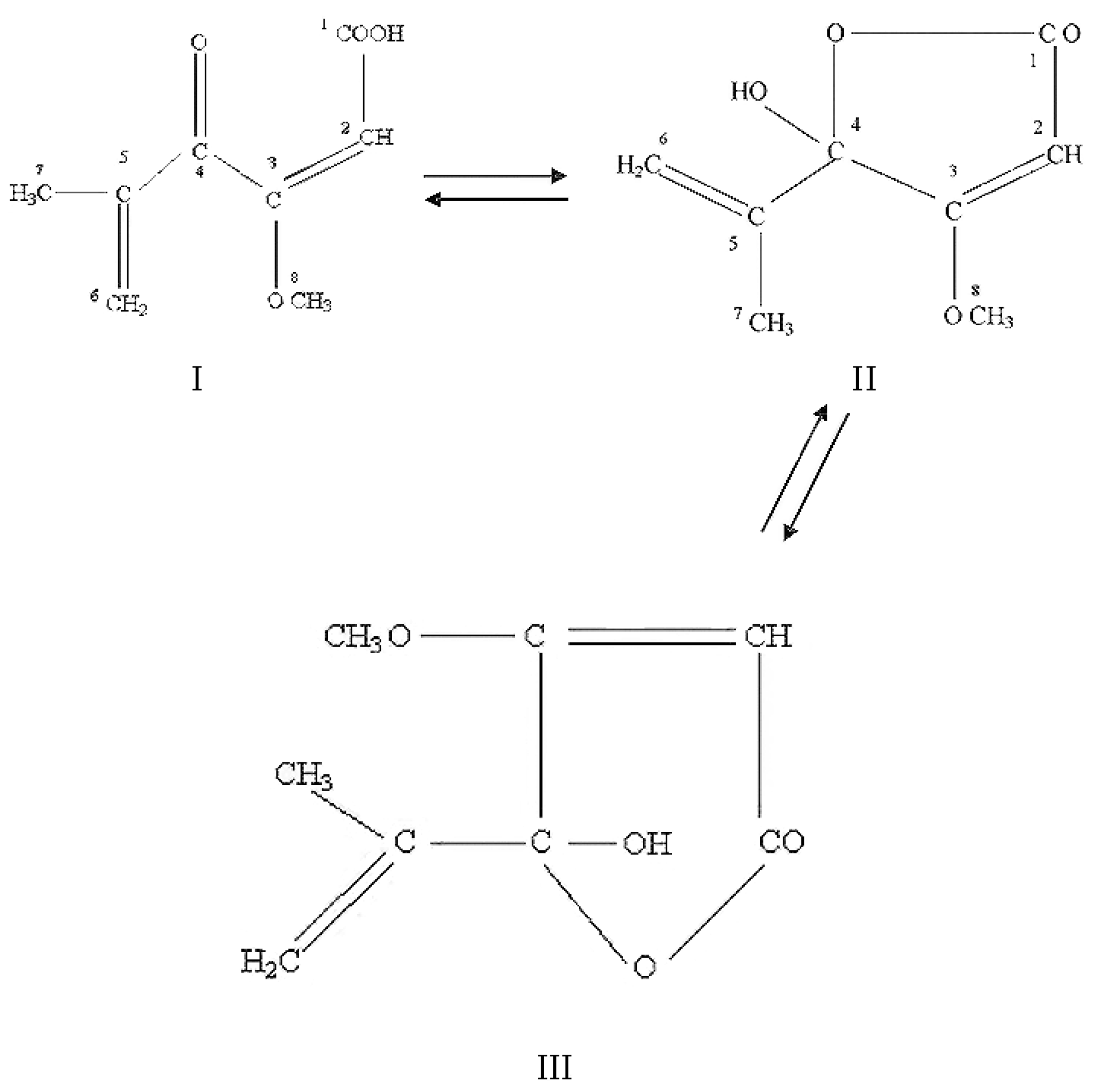
2.4.1. Historical Perspective and Tautomeric Formulae
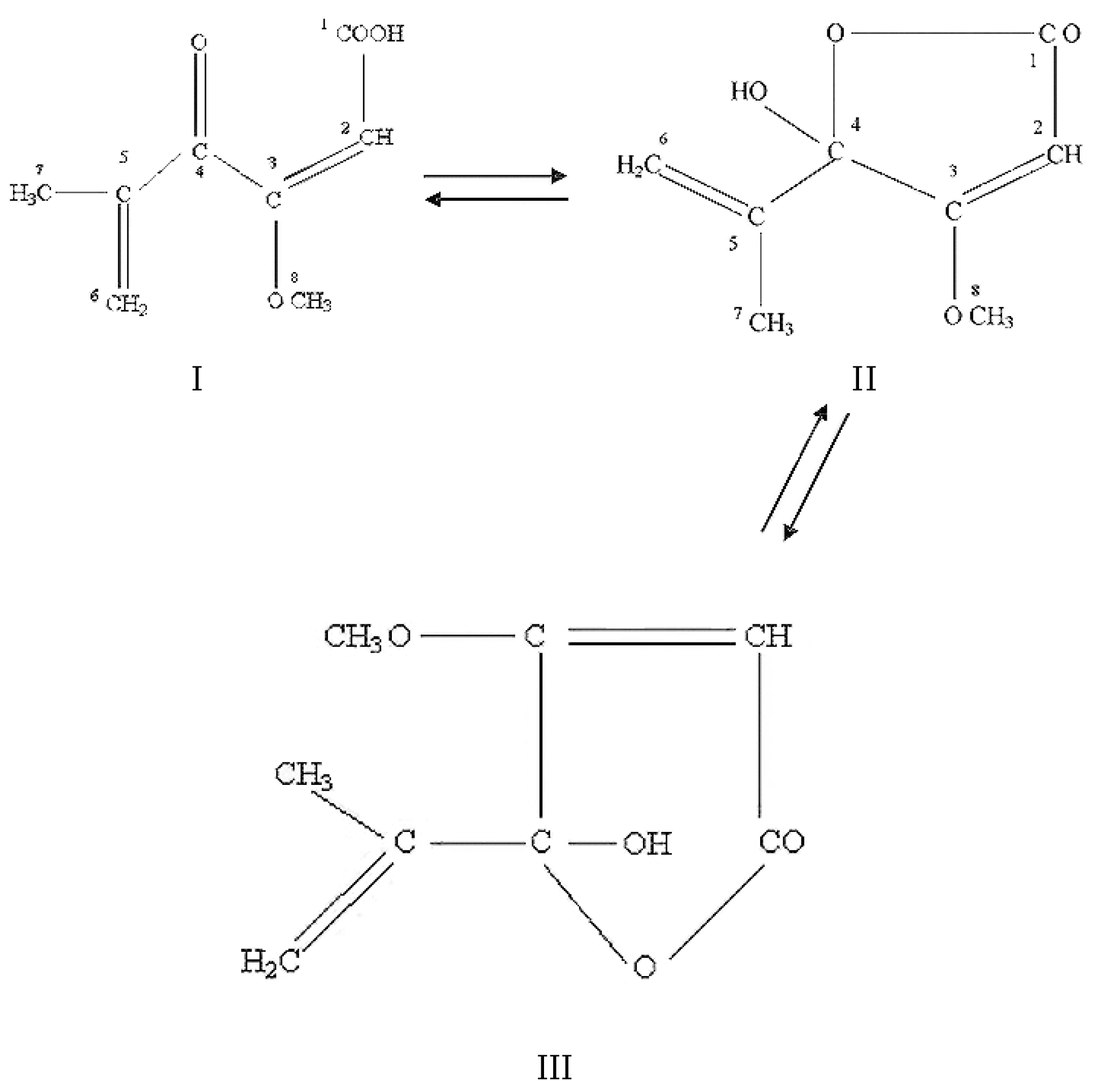
2.4.2. Producing Fungal Species
2.4.3. Phytotoxicity
2.4.4. Mechanism of Action
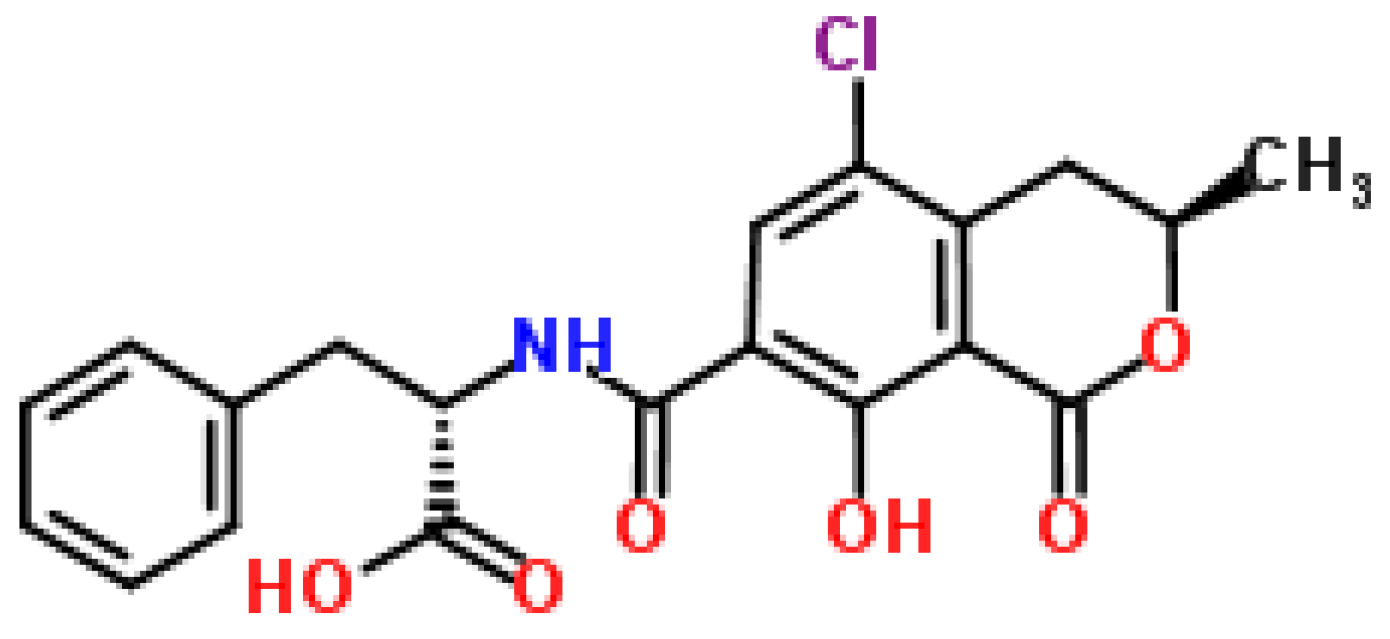
2.5. Ochratoxin
2.5.1. Historical Perspective and Producing Fungal Species
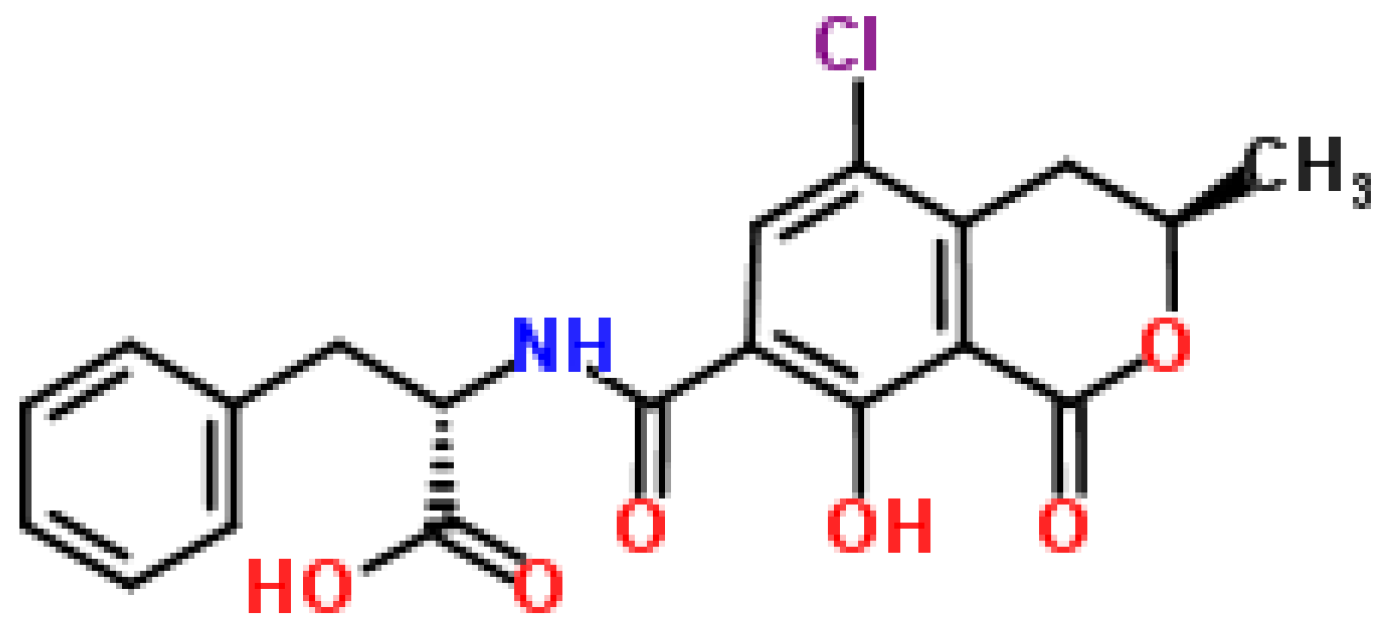
2.5.2. Phytotoxicity
2.5.3. Mechanism of Action
3. Mycotoxins Produced by Dematiaceous Hyphomycetes
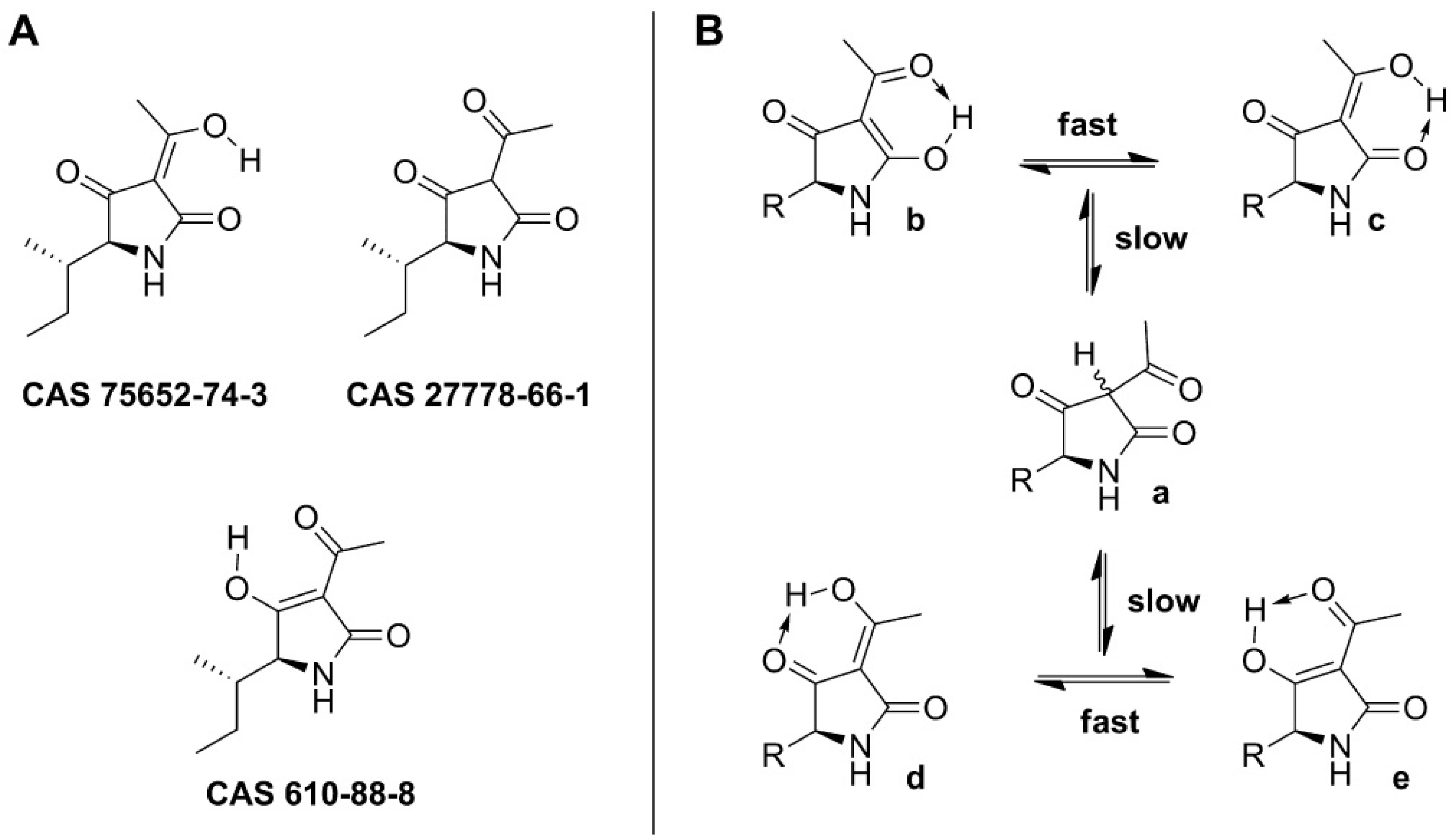
3.1. Tenuazonic Acid
3.1.1. Historical Perspective and Tautomeric Formulae
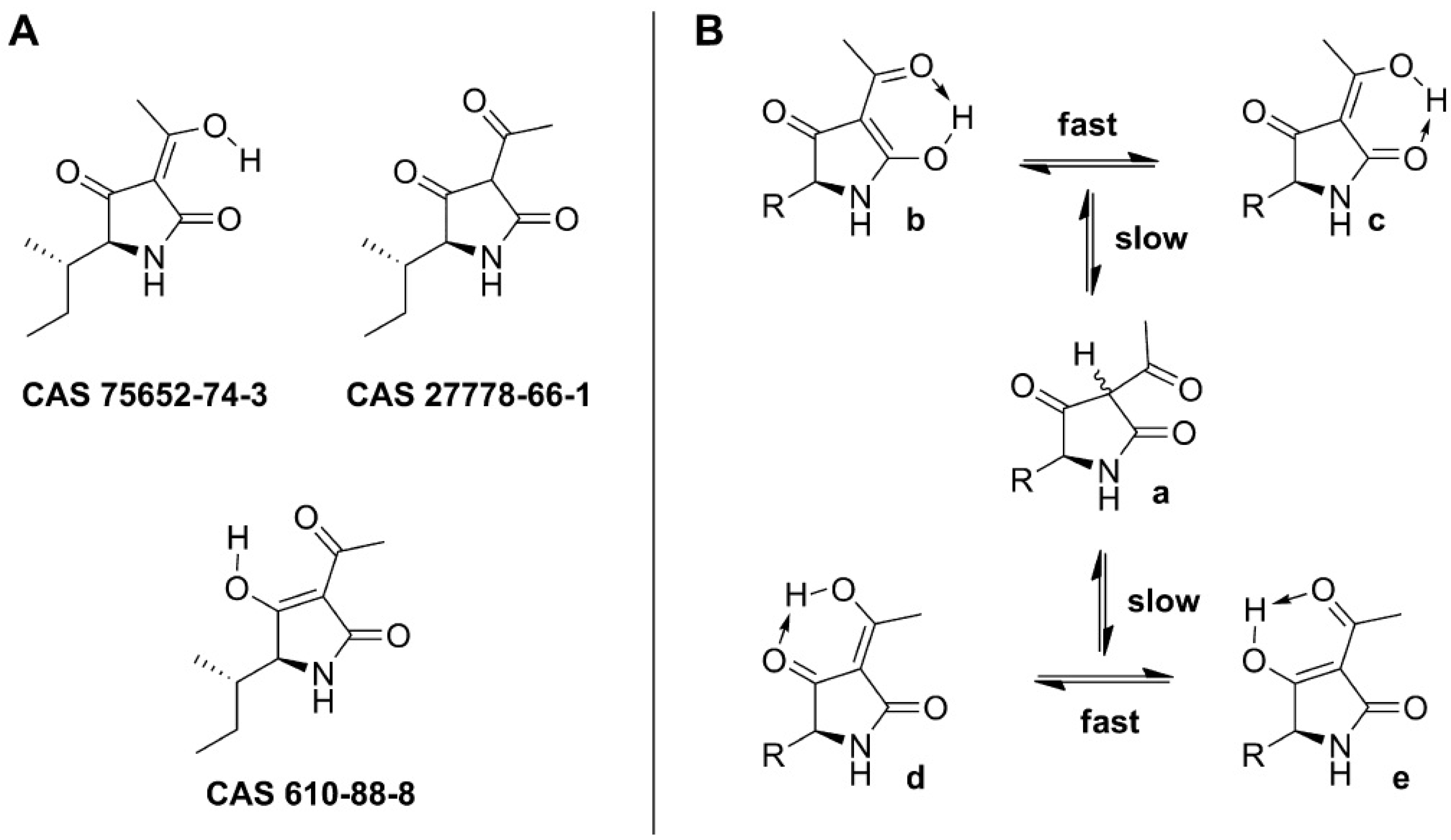
3.1.2. Phytotoxicity
3.1.3. Mechanism of Action
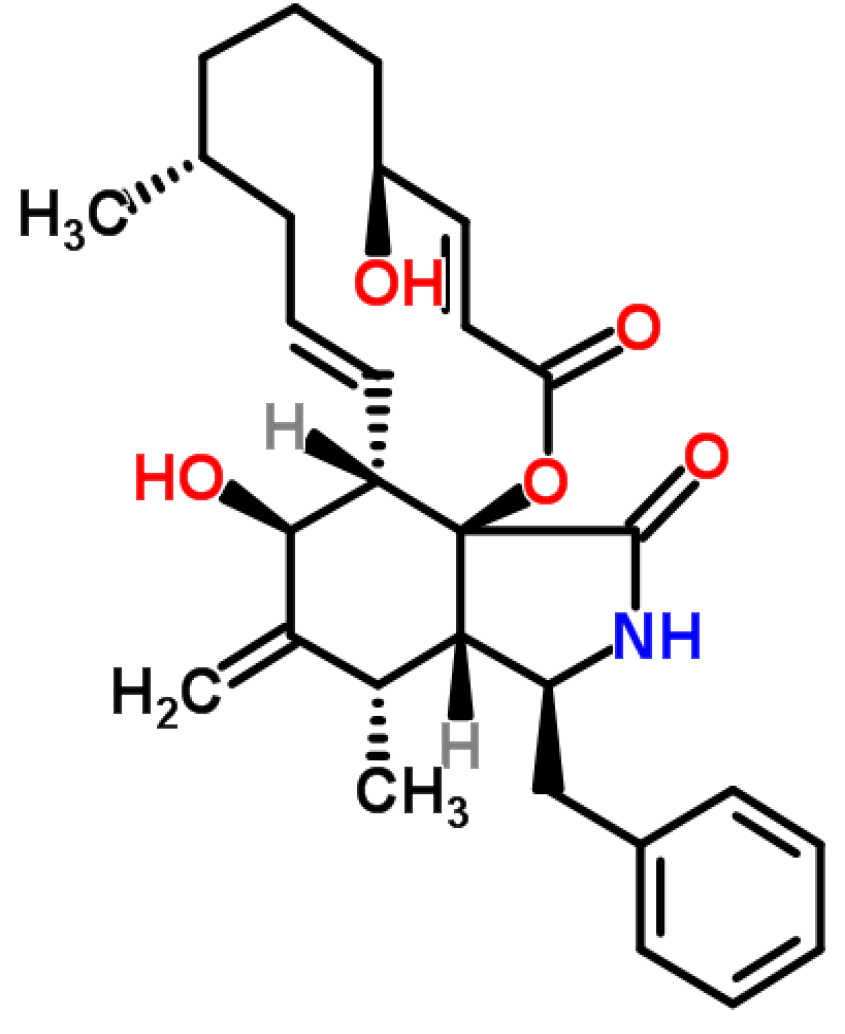
3.2. Cytochalasins
3.2.1. Historical Perspective and Producing Fungal Species
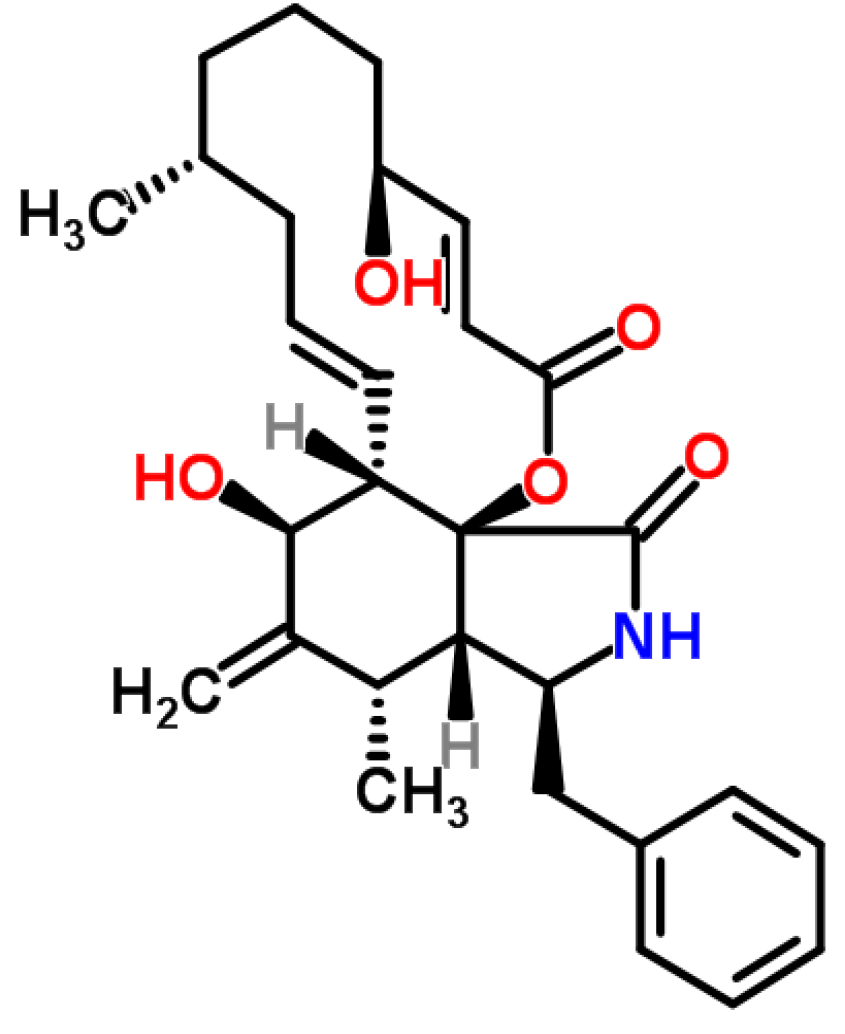
3.2.2. Phytoxicity and Mechanism of Action
4. Mycotoxins Produced by Fusarium and Other Species
4.1. Trichothecenes
4.1.1. Historical Perspective and Producing Fungal Species
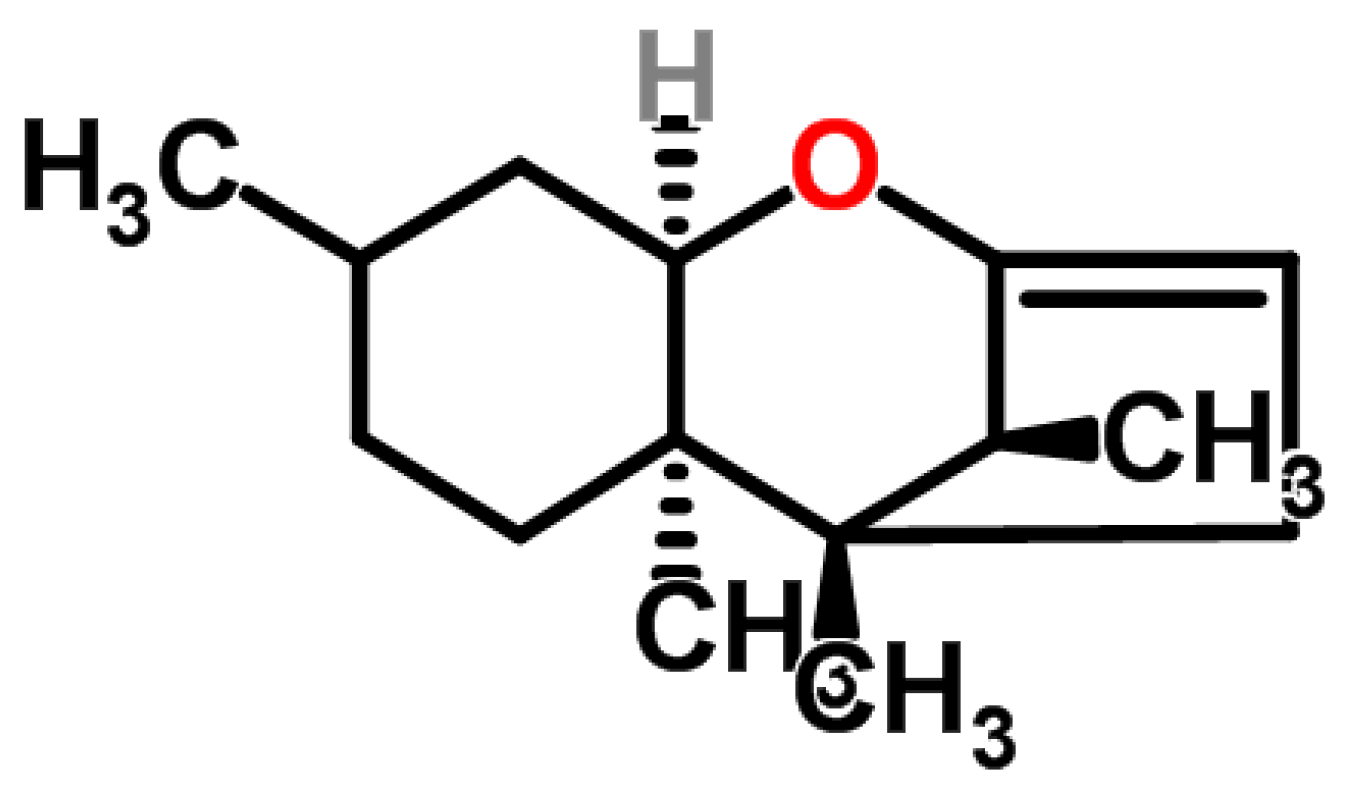
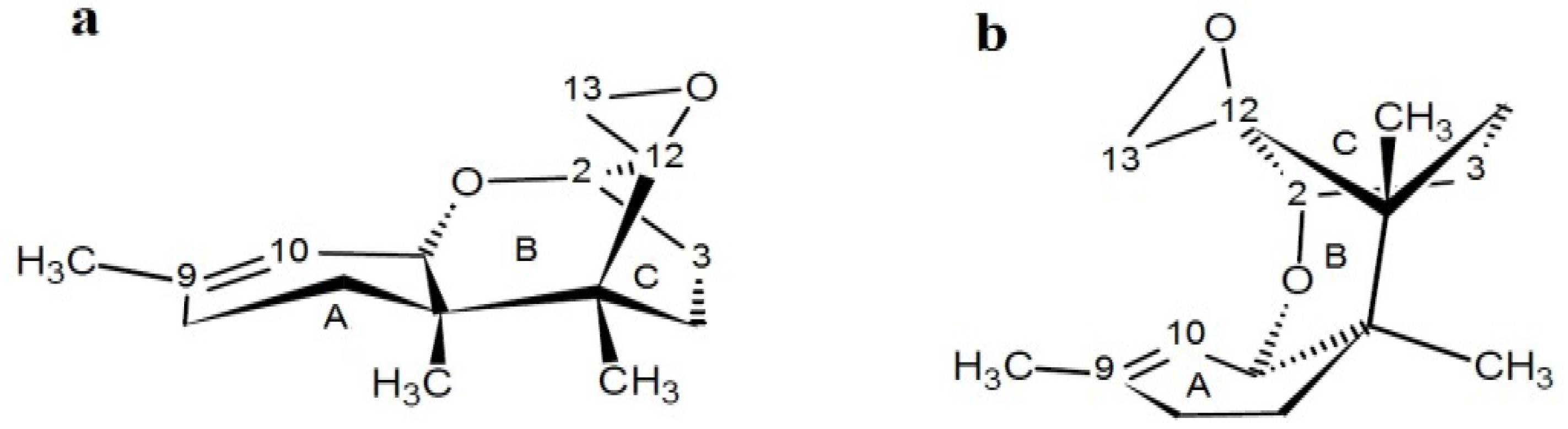
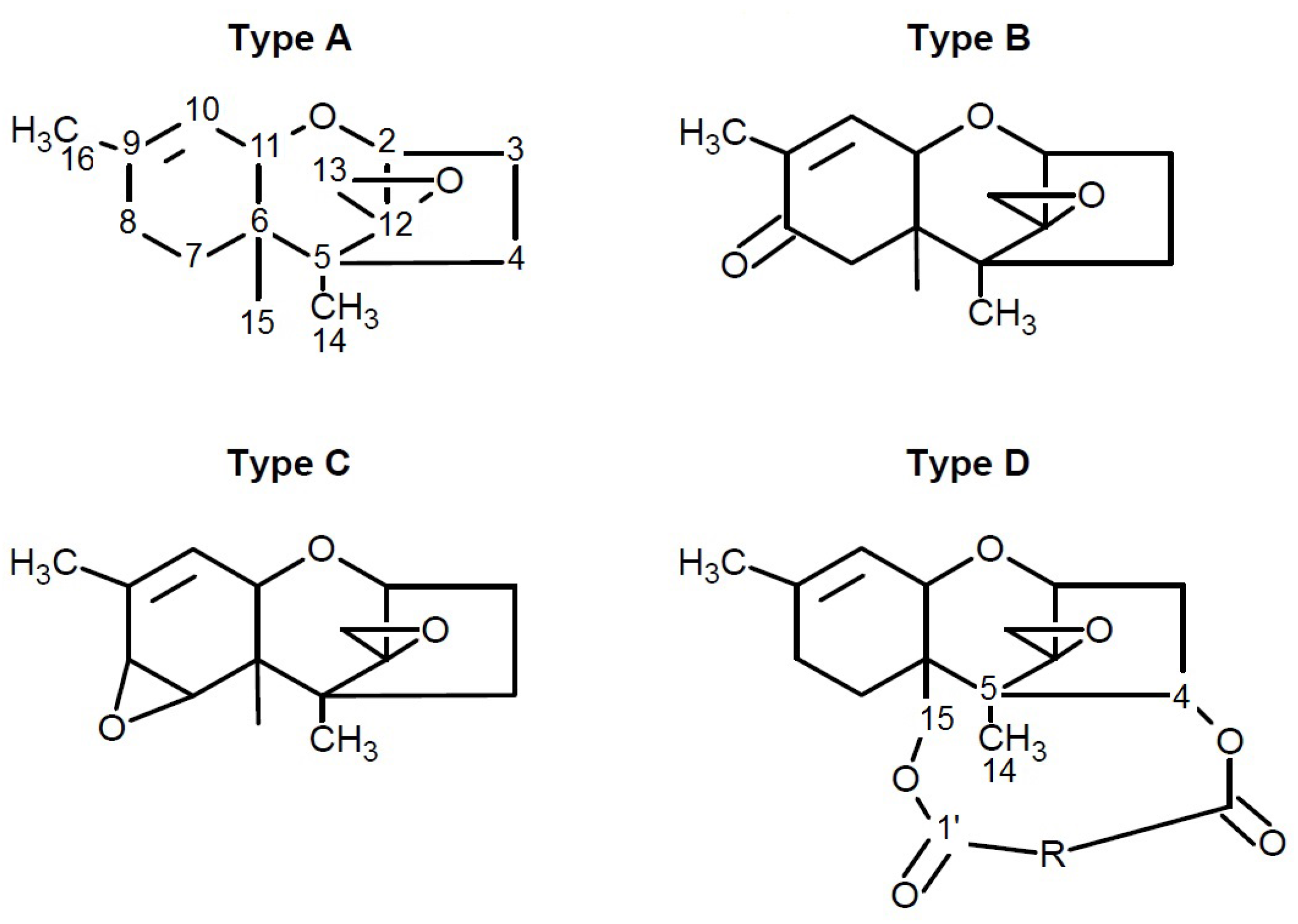
4.1.2. Phytotoxicity and Mechanism of Action
4.1.3. Deoxynivalenol (Vomitoxin) (DON) and Nivalenol (NIV)
4.1.4. Diacetyoxyscirpenol (DAS)
4.1.5. T-2 Toxin
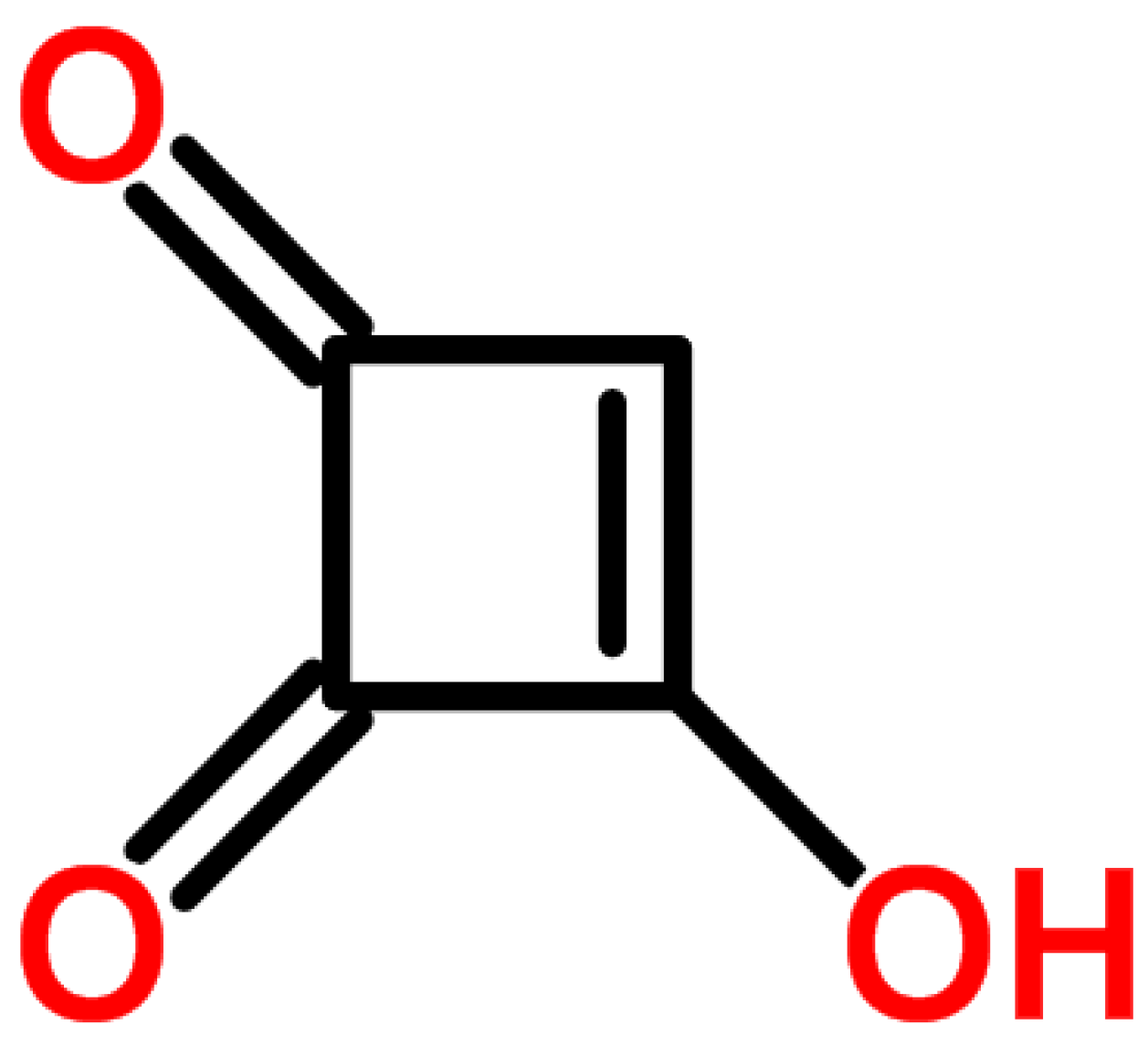
4.2. Moniliformin (MON)
4.2.1. Historical Perspective and Producing Fungal Species
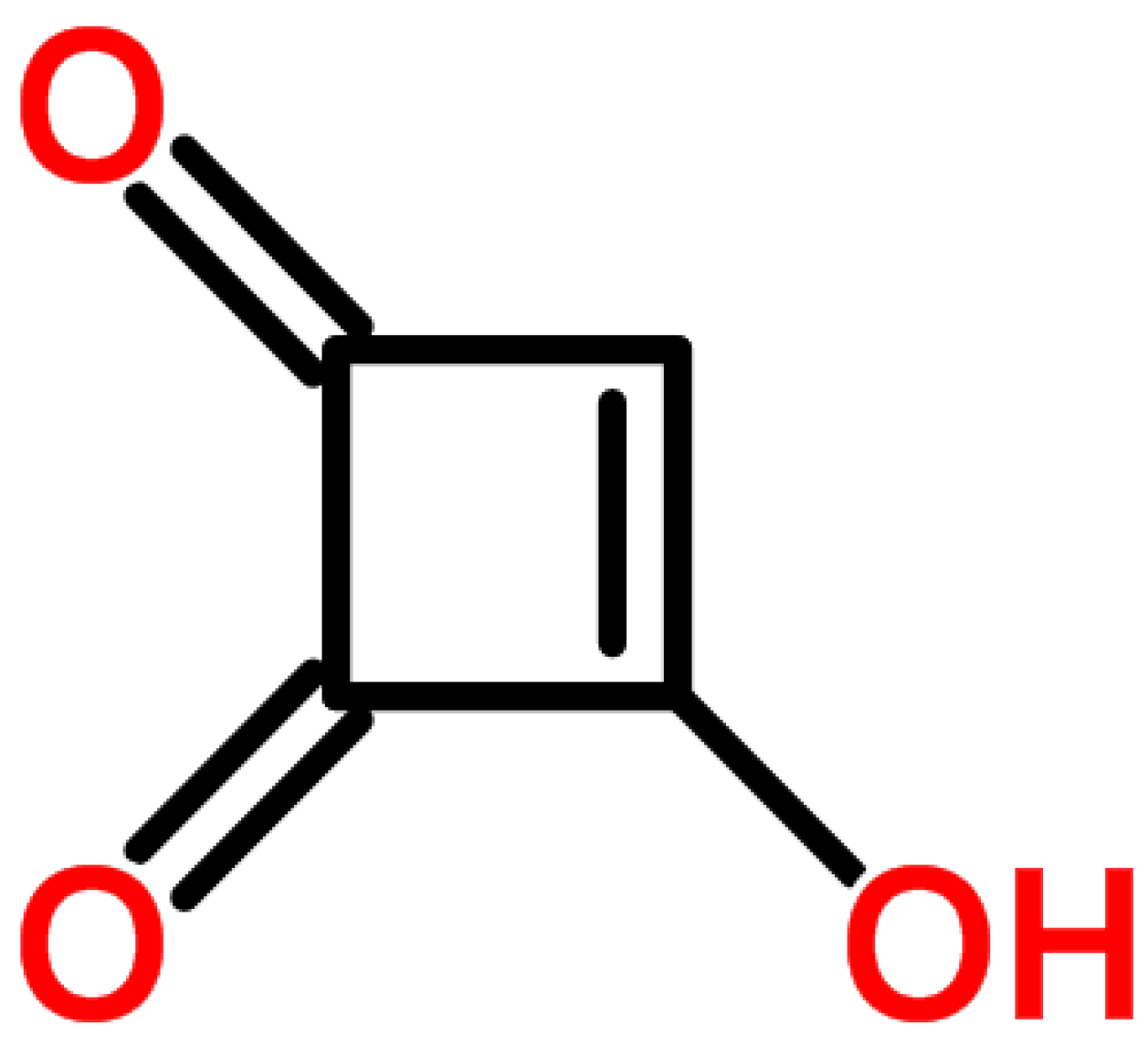
4.2.2. Phytotoxicity and Mechanism of Action
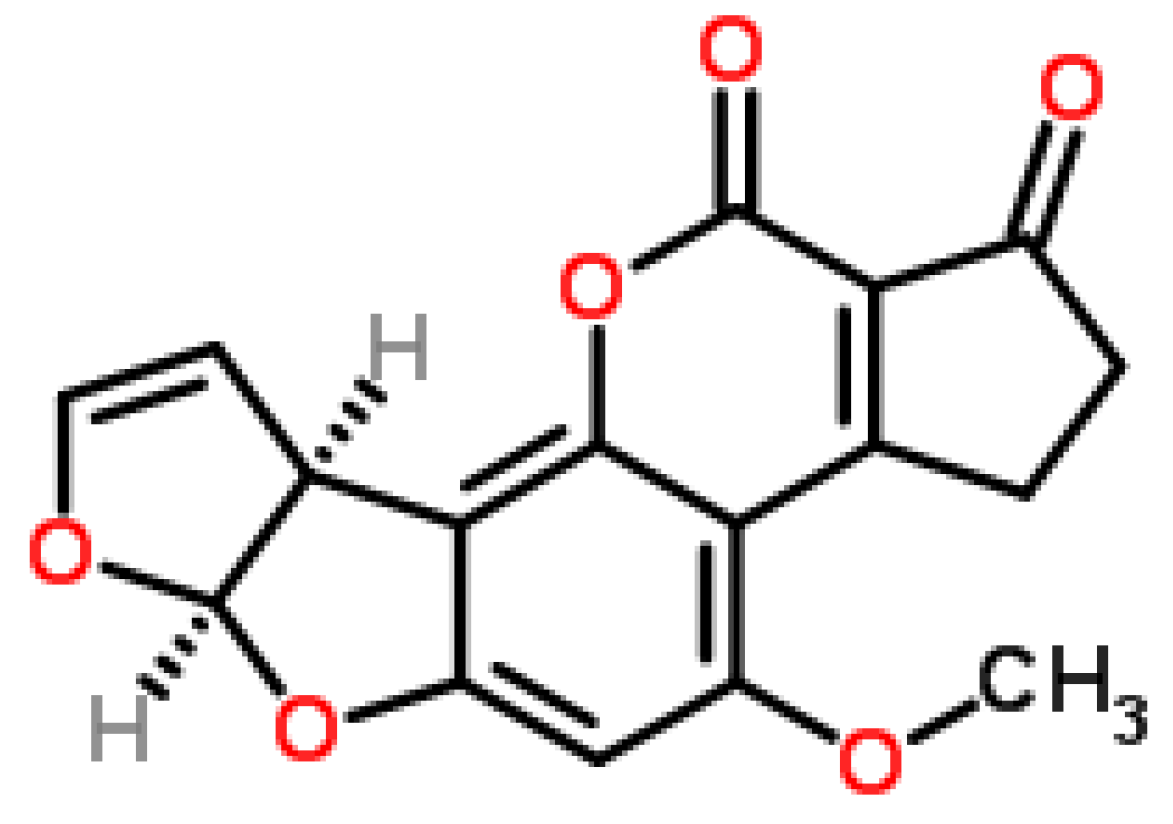
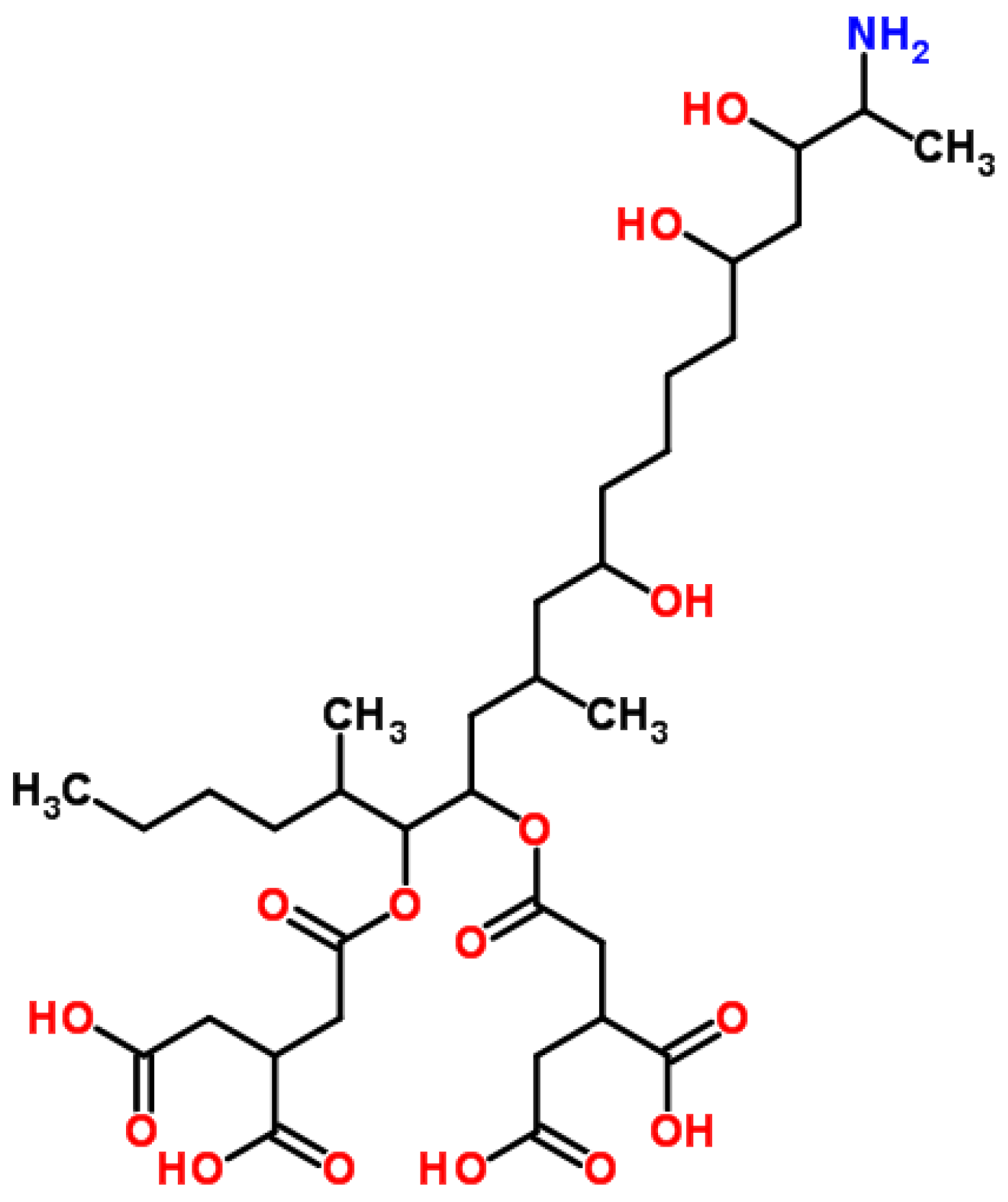
4.3. Fumonisin B1 (FB1)
4.3.1. Historical Perspective and Producing Fungal Species
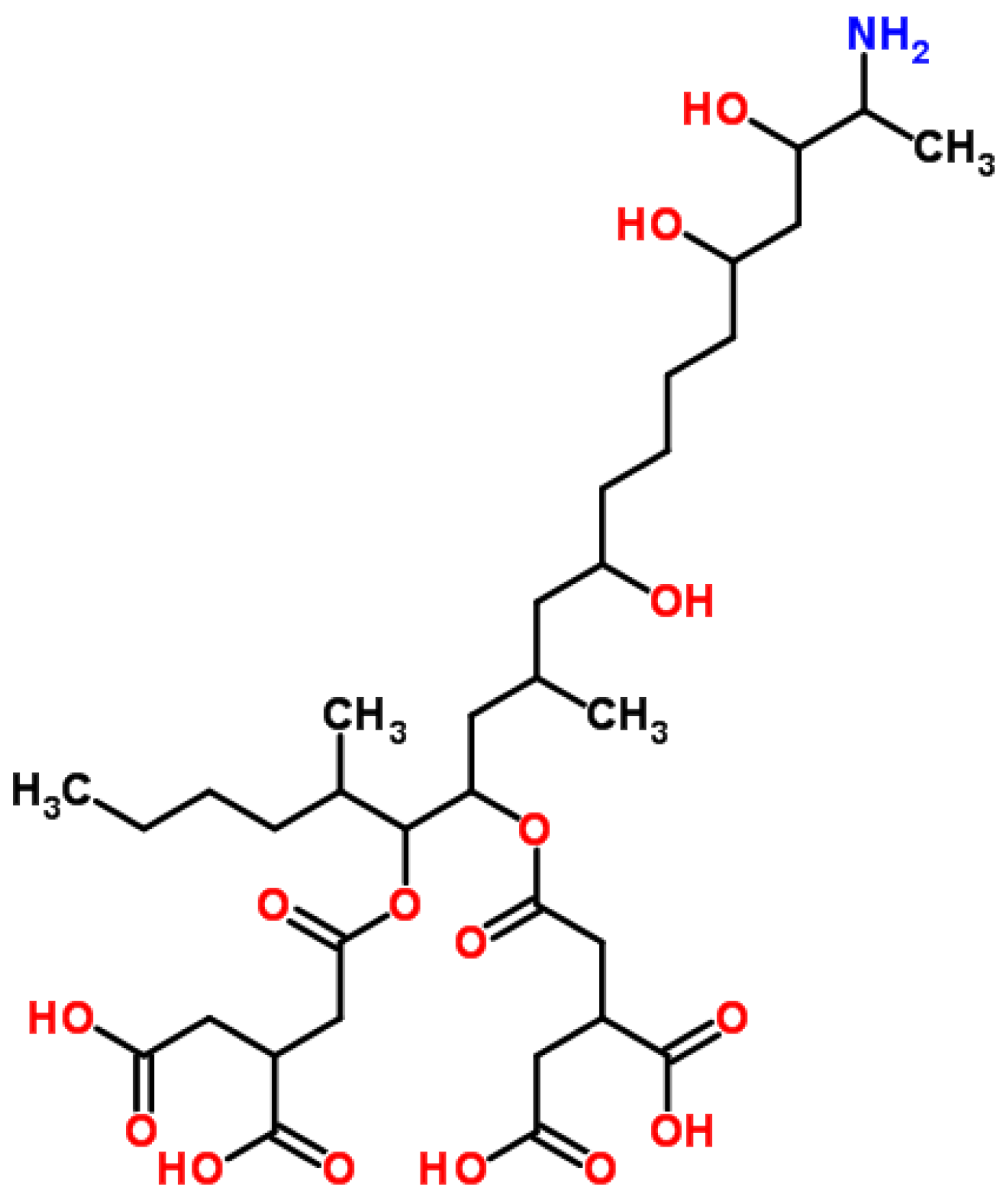
4.3.2. Phytotoxicity
4.3.3. Mechanism of Action
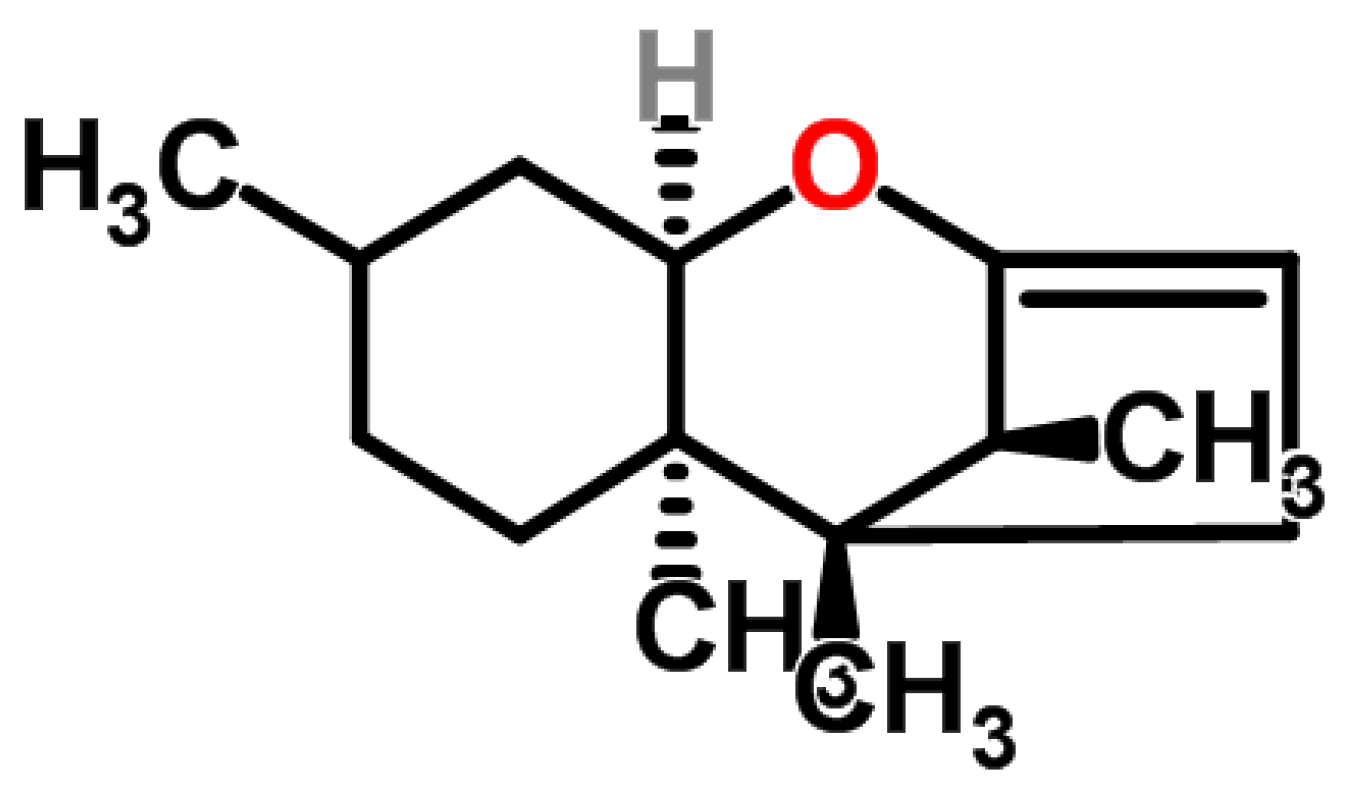
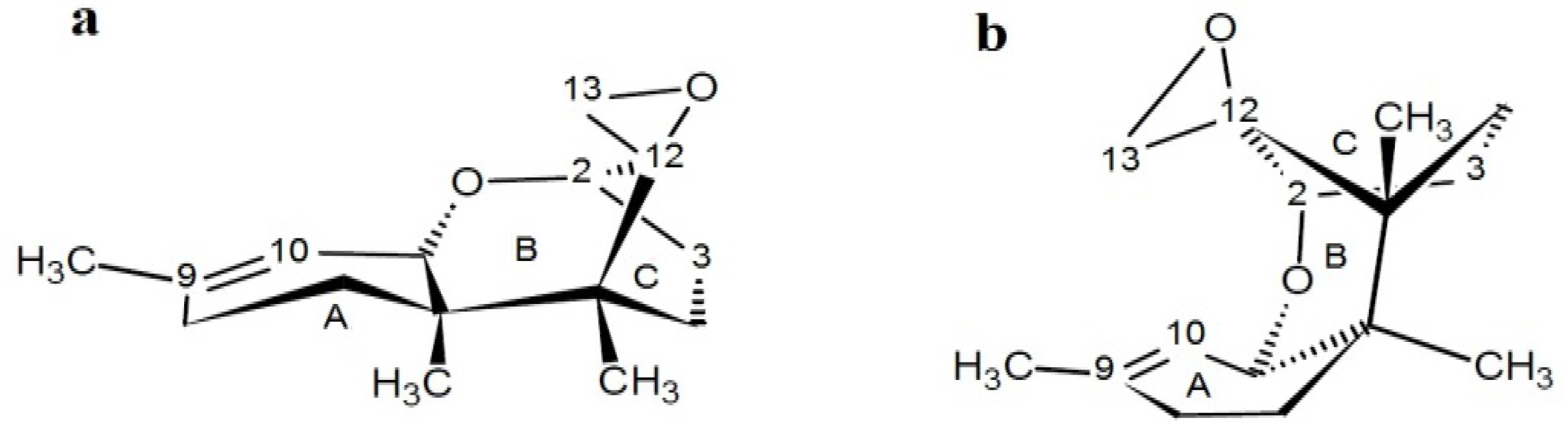
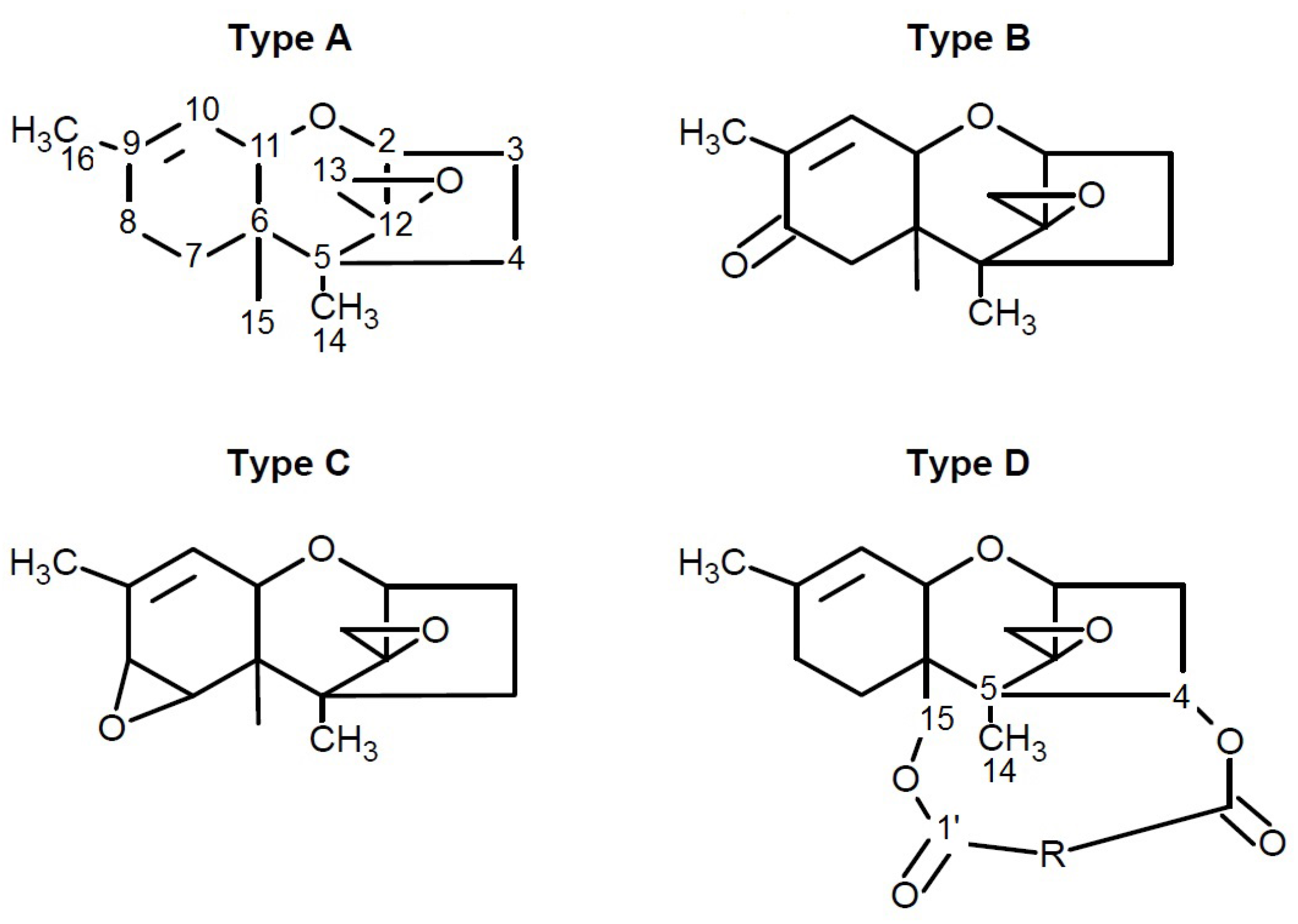
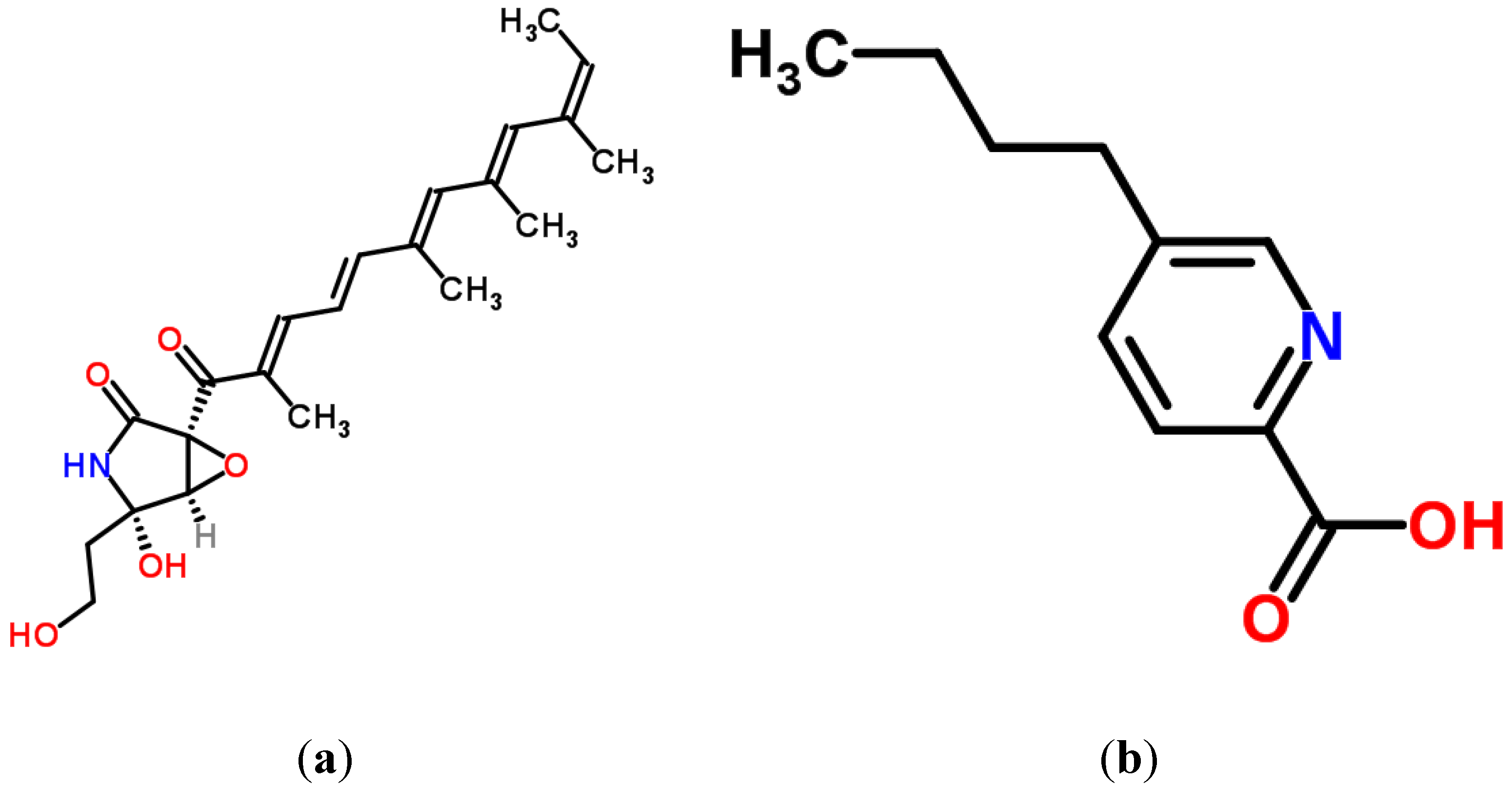
4.4. Fusarins and Fusaric Acid (FA)
4.4.1. Historical Perspective and Producing Fungal Species
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.4.2. Phytotoxicity and Mechanism of Action
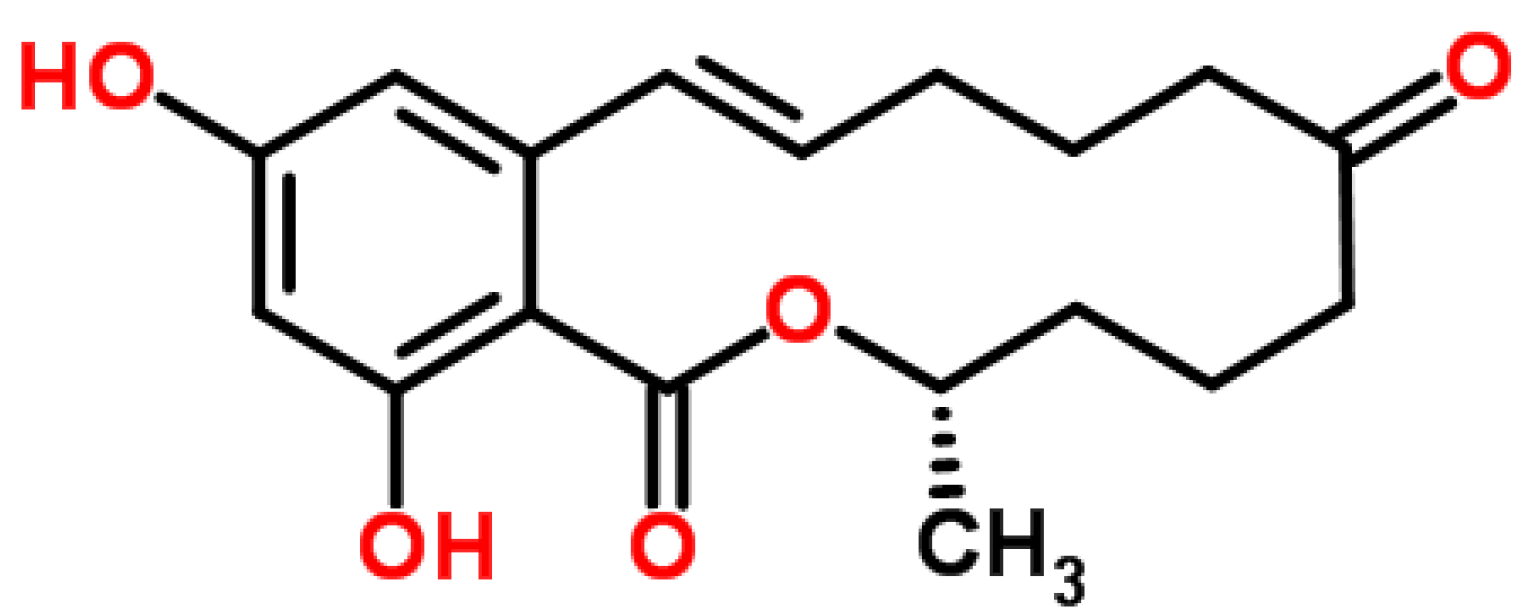
4.5. Zearalenone (ZEA)
4.5.1. Historical Perspective
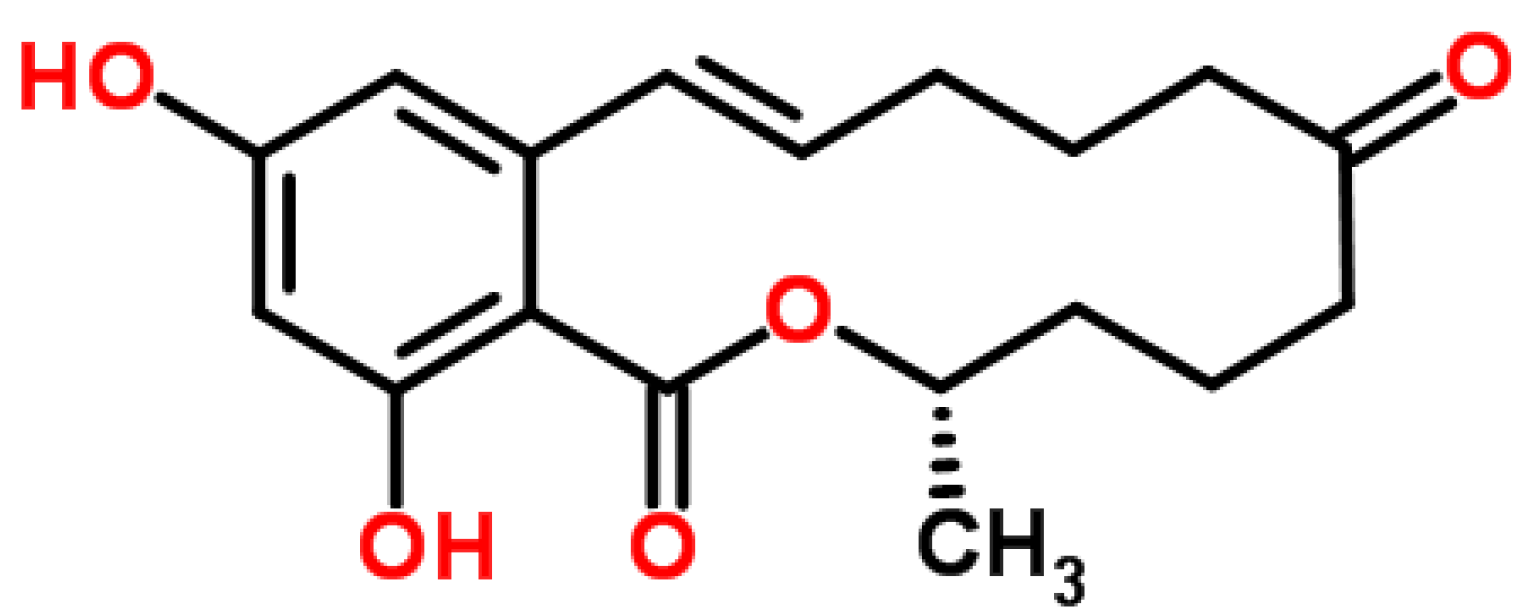
4.5.2. Producing Fungal Species
4.5.3. Phytotoxicity and Mechanism of Action
5. Remarks on the Phytotoxic Properties of Mycotoxins
| Mycotoxin | Dose | Affected Plants | Phytotoxic Effects | Ref. |
|---|---|---|---|---|
| Aflatoxin | >10 μg·mL−1 | Lepidium sativum | Maximal reduction in the growth rate of hypocotyls after germination | [40] |
| 0.83 μg·mL−1 | Barley | LD50 seedling viability | [47] | |
| 1.74 μg·mL−1 | Wheat | LD50 seedling viability | [47] | |
| 2.75 μg·mL−1 | Sorghum | LD50 seedling viability | [47] | |
| CTN | 10−3–10−4 M | Bean, cotton, sorghum | Symptoms of wilting | [63] |
| PAT | 25 μg·mL−1 | Maize | 5% inhibition of radicle emergence | [17] |
| 100 μg·mL−1 | Wheat | Decreases in internodal elongation, floret number, seed weight, and seed number | [96] | |
| PA | 250–500 μg·mL−1 | Picea glehnii | Browning of Picea glehnii roots without root destruction | [16] |
| Lettuce | Growth inhibition of lettuce seedlings | [16] | ||
| 500 μg·mL−1 | Corn | Growth of the main root was reduced 50% | [18] | |
| OTA | 1–2 mM | Arabidopsis thaliana | Formation of macroscopic lesions on leaves within 1–2 days. | [144] |
| 50 μM | Arabidopsis thaliana | Blocking of root elongation | [145] | |
| TA | 0.12 mM | Rice | Leaf browning and 80% root growth inhibition | [161] |
| 100 μg·mL−1 | Datura innoxia, rye, wheat, lettuce and green gram | Growth inhibition of germinating seeds | [160] | |
| 200 μg·mL−1 | Datura innoxia | Localized chlorotic spots turned into necrotic after 24 h | [160] | |
| CB | 10−3 M | Bromus tectorum | Coleoptile elongation reduction to 34% of the control | [203] |
| CE | 5 ppm | Apple | Limpness of shoot | [197] |
| 10−4 M | Striga hermonthica | LD50 seedling viability | [199] | |
| Pyrenophoric acid | 10−3 M | Cheatgrass | Coleoptile length reduction relative to the control by approximately half | [202] |
| Pyrichalasin H | 1 μg·mL−1 | Rice | Characteristic curling of the shoot | [204] |
| Zygosporin D | 1 nmol/plant | Rice | Reduction of the second leaf sheath length of treated plants to 41% of control | [205] |
| DON | 2 μg·mL−1 | Wheat | 50% inhibition of root growth | [241] |
| 10 μg·mL−1 | Wheat | 50% inhibition of shoot growth | [241] | |
| 50 μg·mL−1 | Wheat | Complete inhibition of germination | [244] | |
| 10–25 μg·mL−1 | Wheat | Plantlet growth reduction | [244] | |
| 10 and 25 μg·mL−1 | Maize | Significant inhibition of both primary root and shoot elongation of germinating embryos | [246] | |
| NIV | 48 μg·mL−1 | Wheat | 50% inhibition of root growth | [241] |
| 100 μg·mL−1 | Wheat | 50% inhibition of shoot growth | [241] | |
| DAS | 1.26 μg·mL−1 | Barley | LD50 seedling viability | [47] |
| 3.98 μg·mL−1 | Wheat | LD50 seedling viability | [47] | |
| 10 μg·mL−1 | Sorghum | LD50 seedling viability | [47] | |
| T-2 toxin | 1 μg·mL−1 | Wheat | Significant inhibition of root and shoot growth of germinating seedlings | [239] |
| 1 μg·mL−1 | Maize | Significant inhibition of callus growth | [20] | |
| 6.7 μg·mL−1 | Duckweed (Lemna minor L.) | Decrease of leaf growth rate | [255] | |
| MON | 10 μg·mL−1 | Wheat | A significant decrease in leaf mass | [239] |
| 10–100 μg·mL−1 | Maize | Significant reduction in fresh mass accumulation | [20] | |
| 66.7 μg·mL−1 | Duckweed (L. minor L.) | Growth reduction to 84% of the control | [255] | |
| FB1 | 1000 μg·mL−1 | Soybeans (Glycine max L.) | Necrosis and wilting | [281] |
| 100 ppm | Maize | Radicle elongation inhibition | [282] | |
| 1–100 μM | Tomato | Leaves necrosis | [280] | |
| 10–200 μg·mL−1 | Jimsonweed | Chlorosis and necrosis and reduction in height and biomass | [281] | |
| FA | 10−3 M | Tomato | Symptoms of wilting | [301] |
| 10−5 M | Arabidopsis thaliana | Toxic effects such as alteration of cell growth, mitochondrial activity and membrane permeability | [306] | |
| ZEA | 5 μg·mL−1 | Maize | Inhibitory effect on root and shoot elongation and fresh mass accumulation of germinating, embryos | [146] |
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bennett, J.W. Mycotoxins, mycotoxicoses, mycotoxicology and mycopathologia. Mycopathologia 1987, 100, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Bentley, R. What’s in a name? Microbial secondary metabolism. Adv. Appl. Microbiol. 1989, 34, 1–28. [Google Scholar]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Legan, J.D. Cereals and cereal products. In The Microbiological Safety and Quality of Food; Lund, B.M., Baird-Parker, T.C., Gould, G.W., Eds.; Aspen Publishers Inc.: Gaithersburg, MD, USA, 2000; pp. 759–783. [Google Scholar]
- Miller, J.D. Fungi and mycotoxins in grain: Implications for stored product research. J. Stored Prod. Res. 1995, 31, 1–16. [Google Scholar] [CrossRef]
- Lillehoj, E.B.; Zuber, M.S. Aflatoxin problem in corn and possible solutions. In Proceedings of the 30th Annual Corn and Sorghum Research Conference; Chicago, IL, USA, 1975; pp. 230–250. Available online: http://naldc.nal.usda.gov/naldc/download.xhtml?id=29471&content=PDF (accessed on 21 July 2015). [Google Scholar]
- Llewellyn, G.C.; O’Donnell, W.A.; Dashek, W.V. Aflatoxin infuences on seed germination and root elongation by two cultivars of Glycine max and uptake of 65Zn-ZnCl2 by the cultivars. Mycopathologia 1984, 86, 129–136. [Google Scholar] [CrossRef]
- McLean, M.; Berjak, P.; Watt, M.P.; Dutton, M.F. The effects of aflatoxin B1 on immature germinating maize (Zea mays) embryos. Mycopathologia 1992, 119, 181–190. [Google Scholar] [CrossRef]
- Anderson, H.W.; Nehring, E.W.; Wichser, W.R. Aflatoxin contamination of corn in the field. J. Agr. Food Chem. 1975, 23, 775–782. [Google Scholar] [CrossRef]
- Mertz, D.; Lee, D.; Zuber, M.; Lillehoj, E. Uptake and metabolism of aflatoxin by Zea mays. J. Agric. Food Chem. 1980, 28, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Filtenborg, O. Classification of terverticillate penicillia based on profiles of mycotoxins and other secondary metabolites. Appl. Environ. Microbiol. 1983, 46, 1301–1310. [Google Scholar] [PubMed]
- Frisvad, J.C.; Thrane, U. Mycotoxin production by food borne fungi. In Introduction to Food-Borne Fungi; Samson, R.A., Hoekstra, E.S., Frisvad, J.C., Filtenborg, O., Eds.; Centraalbureau Voor Schimmelcultures: Baarn, The Netherlands, 1996; pp. 251–260. [Google Scholar]
- Müller, H.M.; Amend, R. Formation and disappearance of mycophenolic acid, patulin, penicillic acid and PR toxin in maize silage inoculated with Penicillium roqueforti. Arch. Tierernahr. 1997, 50, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium. A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–174. [Google Scholar]
- Yamaji, K.; Fukushi, Y.; Hashidoko, Y.; Yoshida, T.; Tahara, S. Characterization of antifungal metabolites produced by Penicillium species isolated from seeds of Picea glehnii. J. Chem. Ecol. 1999, 25, 1643–1654. [Google Scholar] [CrossRef]
- Yamaji, K.; Fukushi, Y.; Hashidoko, Y.; Tahara, S. Penicillium frequentans from Picea glehnii seedling roots as a possible biological control agent against damping-off. Ecol. Res. 2005, 20, 103–107. [Google Scholar]
- Ismaiel, A.A.; Papenbrock, J. The effects of patulin from Penicilllium vulpinum on seedling growth, root tip ultrastructure and glutathione content of maize. Eur. J. Plant Pathol. 2014, 139, 497–509. [Google Scholar] [CrossRef]
- Keromnes, J.; Thouvenot, D. Role of penicillic acid in the phytotoxicity of Penicillium cyclopium and Penicillium canescens to the germination of corn seeds. Appl. Environ. Microbiol. 1985, 49, 660–663. [Google Scholar] [PubMed]
- Koteswara Rao, V.; Girisham, S.; Reddy, S.M. Influence of different species of Penicillium and their culture filtrates on seed germination and seedling growth of sorghum. J. Biochem. Technol. 2014, 5, 832–837. [Google Scholar]
- Van Asch, M.A.J.; Rijkenberg, F.H.F.; Coutinho, T.A. Phytotoxicity of fumonisin B1, moniliformin and T-2 toxin in corn callus cultures. Phytopathology 1992, 82, 1330–1332. [Google Scholar] [CrossRef]
- Nelson, E.P.; Dignani, M.C.; Anaissie, E.J. Taxonomy, biology, and clinical aspects of Fusarium species. Clin. Microbiol. Rev. 1994, 7, 479–504. [Google Scholar] [PubMed]
- McLean, M. The phytotoxicity of Fusarium metabolites: An update since 1989. Mycopathologia 1996, 133, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Hohn, T.M. Mycotoxins in plant pathogenesis. Mol. Plant-Microbe Interact. 1997, 10, 147–152. [Google Scholar] [CrossRef]
- Kühl, H. Pharmazeutische Zentrahalle. für Deutschland. In Historical Aspects of Mycotoxicology and Developments in Aflatoxins, Proceedings of the International Symposium on Mycotoxins; Cairo, Egypt, 6–8 September 1981, Mirocha, C.J., Ed.; Food and Drug Administration: RockVille, MD, USA; National Research Center: Cairo, Egypt, 1983. [Google Scholar]
- Blount, W.P. Turkey X Disease. J. Br. Turkey Fed. 1961, 9, 52–61. [Google Scholar]
- Asplin, F.D.; Carnaghan, R.B.A. The toxicity of certain groundnut meals for poultry with special reference to their effect on ducklings and chickens. Vet. Rec. 1961, 73, 1215–1218. [Google Scholar]
- Sargeant, K.; O’Kelly, J.; Carnaghan, R.B.A.; Alleroft, R. The assay of a toxic principle in certain groundnut meals. Vet. Rec. 1961, 73, 1219–1222. [Google Scholar]
- Sargeant, K.; Sheridan, A.; O’Kelly, J.; Carnaghan, R.B.A. Toxicity associated with certain samples of groundnuts. Nature 1961, 192, 1096–1097. [Google Scholar] [CrossRef]
- Sargeant, K.; Carnaghan, R.B.A.; Alleroft, R. Toxic products in Groundnuts: Chemistry and Origin. Chem. Ind. 1963, 31, 53–55. [Google Scholar]
- Asao, T.; Buchi, G.; Abdel Kader, M.M.; Chang, S.B.; Wick, E.; Wogan, G.N. Aflatoxins A and G. J. Amer. Chem. Soc. 1963, 85, 1706–1707. [Google Scholar] [CrossRef]
- Richard, J.L.; Payne, G.A.; Desjardin, A.E.; Maragos, C.; Norred, W.P.; Pestka, J.J. Mycotoxins, risks in plant, animal and human systems. In CAST Task Force Report 139; Council for Agricultural Science and Technology: Ames, IA, USA, 2003; pp. 101–103. [Google Scholar]
- Ismaiel, A.A.; Tharwat, N.A. Antifungal activity of silver ion on ultrastructure and production aflatoxin B1 and patulin by two mycotoxigenic strains, Aspergillus flavus OC1 and Penicillium vulpinum CM1. J. Mycol. Méd. 2014, 24, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Chipley, J.R.; Uraih, N. Inhibition of Aspergillus growth and aflatoxin release by derivatives of benzoic acid. Appl. Environ. Microbiol. 1980, 40, 352–357. [Google Scholar] [PubMed]
- Paranagama, P.A.; Abeysekera, K.H.T.; Abeywickrama, K.; Nugaliyadde, L. Fungicidal and anti-aflatoxigenic effects of the essential oil of Cymbopogon citratus (DC.) Stapf. (lemongrass) against Aspergillus flavus Link. isolated from stored rice. Lett. Appl. Microbiol. 2003, 37, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, Y.L.; Shashikala, J. Inhibition of aflatoxin B1 production of Aspergillus flavus, isolated from soybean seeds by certain natural plant products. Lett. Appl. Microbiol. 2006, 43, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Gnonlonfin, G.J.B.; Adjovi, Y.; Gbaguidi, F.; Gbenou, J.; Katerere, D.; Brimer, L.; Sanni, A. Scopoletin in cassava products as an inhibitor of aflatoxin production. J. Food Saf. 2011, 4, 553–558. [Google Scholar] [CrossRef]
- Ismaiel, A.A.; Ghaly, M.F.; El-Naggar, A.K. Milk kefir: Ultrastructure, antimicrobial activity, and efficacy on aflatoxin B1 production by Aspergillus flavus. Curr. Microbiol. 2011, 62, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Pratiwi, C.; Rahayu, W.P.; Lioe, H.N.; Herawati, D.; Broto, W.; Ambarwati, S. The effect of temperature and relative humidity for Aspergillus flavus BIO 2237 growth and aflatoxin production on soybeans. Int. Food Res. J. 2015, 22, 82–87. [Google Scholar]
- ChemSpider. Available online: http://www.chemspider.com (accessed on 13 July 2015).
- Crisan, E.V. Effects of aflatoxin on seedling growth and ultrastructure in plants. Appl. Microbiol. 1973, 12, 991–1000. [Google Scholar]
- Sinha, K.K.; Kumari, P. Some physiological abnormalities induced by aflatoxin B1 in mung seeds (Vigna radiata var. Pusa Baishakhi). Mycopathologica 1990, 110, 77–79. [Google Scholar] [CrossRef]
- Sinha, K.S.; Sinha, A.K. Effect of aflatoxin B1 on germination index and seedling growth in wheat varieties. Mycopathologica 1993, 123, 165–169. [Google Scholar] [CrossRef]
- Adekunle, A.A.; Bassir, O. The effects of afatoxinB1 and G1 on the germination and leaf colour of cowpea (Vigna sinensis). In Mycopathologia et Mycologia Applicata; Klower Academic Publisher: Ibadan, Nigeria, 1997; pp. 1–12. [Google Scholar]
- Samuel, A.T.; Valentine, I.T. Effect of total aflatoxin on the growth characteristics and chlorophyll level of sesame (Sesamum indicum L.). N. Y. Sci. J. 2014, 7, 8–13. [Google Scholar]
- Reiss, J. Effects of mycotoxins on higher plants, algae, fungi and bacteria. In Mycotoxic Fungi, Mycotoxins, Mycotoxicoses: An Encyclopaedic Handbook Volume 3. Mycotoxicoses of Man and Plants: Mycotoxin Control and Regulatory Practices; Wyllie, T.D., Morehouse, L.G., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1978; pp. 119–143. [Google Scholar]
- McLean, M.; Watt, M.P.; Berjak, P.; Dutton, M.F. Aflatoxin B1—Its effects on an in vitro plant system. Food Addit. Contam. 1995, 12, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Hasan, H.A.H. Phytotoxicity of pathogenic fungi and their mycotoxins to cereal seedling viability. Mycopathologia 1999, 148, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Fadl-Allah, E.M.; Mahmoud, M.A.-H.; Abd El-Twab, M.H.; Helmey, R.K. Aflatoxin B1 induces chromosomal aberrations and 5S rDNA alterations in durum wheat. J. Assoc. Arab Univ. Basic Appl. Sci. 2011, 10, 8–14. [Google Scholar] [CrossRef]
- Slowatizky, I.; Mayer, A.M.; Plojakoff-Mayber, A. The effect of aflatoxin on greening of etiolated leaves. Isr. J. Bot. 1969, 18, 31–36. [Google Scholar]
- Tripathi, R.K.; Misra, R.S. Mechanism of inhibition of maize seed germination by aflatoxin BI. In Mycotoxins in Food and Feed; Bilgrami, K.S., Prasad, T., Sinha, K.K., Eds.; Allied Press: Bhagalpur, India, 1983; pp. 129–141. [Google Scholar]
- McLean, M.; Watt, M.P.; Berjak, P.; Dutton, M.F.; Snyman, C. Immunocytochemical localisation of aflatoxin B1: A preliminary investigation. Commun. Electron Microsc. Soc. South. Afr. 1992, 22, 73–74. [Google Scholar]
- Vogelstein, B.; Kinzler, K.W. P53 function and dysfunction. Cell 1992, 70, 523–526. [Google Scholar] [CrossRef]
- Lane, D.P. P53, guardian of the genome. Nature 1992, 358, 15–16. [Google Scholar] [CrossRef] [PubMed]
- Symonds, H.; Krall, L.; Remington, L.; Saenz-Robles, M.; Lowe, S.; Jacks, T.; van Dyke, T. P53 dependency apoptosis suppresses tumor growth and progression in vivo. Cell 1994, 78, 703–711. [Google Scholar] [CrossRef]
- Hetherington, A.C.; Raistrick, H. Studies in the Biochemistry of Micro-organisms. On the production and chemical constitution of a new yellow colouring matter, citrinin, produced from glucose by Penicillium citrinum. Trans. Roy. Soc. (Lond.) B 1931, 220, 269–295. [Google Scholar] [CrossRef]
- Ciegler, A.; Vesonder, R.F.; Jackson, L.K. Production and biological activity of patulin and citrinin from Penicillium expansum. Appl. Environ. Microbiol. 1977, 33, 1004–1006. [Google Scholar] [PubMed]
- Bragulat, M.R.; Martinez, E.; Castella, G.; Cabanes, F.J. Ochratoxin A and citrinin producing species of the genus Penicillium from feedstuffs. Int. J. Food Microbiol. 2008, 126, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.S.; Kirby, L.K.; Beasley, J.N.; Johnson, Z.B.; Ciegler, A. The effect of drying method and storage time on citrinin activity in corn. Poult. Sci. 1985, 64, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.M.; van Walbeek, W.; Kennedy, B.; Anyeti, B. Mycotoxins (ochratoxin A, citrinin and sterigmatocystin) and toxigenic fungi in grains and other agricultural products. J. Agric. Food Chem. 1972, 20, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Sago, Y.; Zheng, Y.; Nakagawa, H.; Kushiro, M. Mycotoxins in rice. Int. J. Food Microbiol. 2007, 119, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Vella, F.; Kpodo, K.; Sorensen, A.K.; Jakobsen, M. The occurrence of mycotoxins in fermented maize products. Food Chem. 1995, 56, 147–153. [Google Scholar]
- White, A.G.; Truelove, B. The effects of aflatoxin B1, citrinin, and ochratoxin A on amino acid uptake and incorporation by cucumber. Can. J. Bot. 1972, 50, 2659–2664. [Google Scholar] [CrossRef]
- Damodaran, C.; Kathirvel-Pandian, S.; Seeni, S. Citrinin, a phytotoxin? Cell. Mol. Life Sci. 1975, 31, 1415–1417. [Google Scholar] [CrossRef]
- Betina, V. Mycotoxins: Chemical, Biological and Environmental Aspects; Elsevier: New York, NY, USA, 1989. [Google Scholar]
- Maćıas, M.; Ulloa, M.; Gamboa, A.; Mata, R. Phytotoxic compounds from the new coprophilous fungus Guanomyces polythrix. J. Nat. Prod. 2000, 63, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Chagas, G.M.; Wambier Kluppel, M.L.; Campello, A.P.; Buchi, D.F.; Oliveira, M.B.M. Alterations induced by citrinin in cultured kidney cells. Cell Struct. Funct. 1994, 19, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Chagas, G.M.; Oliveira, M.B.M.; Campello, A.P.; Kluppel, M.L.W. Mechanism of citrinin-induced dysfunction of mitochondria. IV. Effect on Ca2+ transport. Cell Biochem. Funct. 1995, 13, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Chain, E.; Florey, H.W.; Jennings, M.A. An antibacterial substance produced by Penicillium claviforme. Brit. J. Exp. Pathol. 1942, 23, 202–205. [Google Scholar]
- Birkinshaw, H.; Bracken, A.; Michael, S.E.; Raistrick, H. Patulin in the common cold. II. Biochemistry and chemistry. Lancet 1943, 245, 625–630. [Google Scholar] [CrossRef]
- Florey, H.W.; Jennings, M.A.; Philpot, F.J. Claviformin from Aspergillus giganteus Wehm. Nature (London) 1944, 153, 139. [Google Scholar] [CrossRef]
- Singh, J. Patulin. In Antibiotics; Gottlieb, D., Shaw, P.D., Eds.; Springer Verlag: New York, NY, USA, 1967; pp. 621–630. [Google Scholar]
- Lovett, J.; Peeler, J.T. Effect of pH on the thermal destruction kinetics of patulin in aqueous solution. J. Food Sci. 1973, 38, 1094–1095. [Google Scholar] [CrossRef]
- Moake, M.M.; Padilla-Zakour, O.I.; Worobo, R.W. Comprehensive review of patulin control methods in foods. Compr. Rev. Food Sci. F 2005, 4, 8–21. [Google Scholar] [CrossRef]
- Gye, W.E. Patulin in the common cold. III. Preliminary trial in the common cold. Lancet 1943, 245, 630–631. [Google Scholar]
- Hopkins, J. The toxicological hazards of patulin. Food Cosmet. Toxicol. 1993, 31, 455–459. [Google Scholar]
- Stansfeld, J.H.; Francis, A.E.; Stuart-Harris, C.H. Laboratory and clinical trials of patulin. Lancet 1944, 244, 370–372. [Google Scholar] [CrossRef]
- Sant’Ana, A.S.; Rosenthal, A.; Massaguer, P.R. The fate of patulin in apple juice processing: A review. Food Res. Int. 2008, 41, 441–453. [Google Scholar] [CrossRef]
- Steiman, R.; Seigle-Murandi, F.; Sage, L.; Krivobok, S. Production of patulin by Micromycetes. Mycopathologia 1989, 105, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Welke, J.E.; Hoeltz, M.; Dottori, H.A.; Noll, I.B. Patulin accumulation in apples during storage by Penicillium expansum and Penicillium griseofulvum strains. Braz. J. Microbiol. 2011, 42, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, H.; Mizuhara, Y.; Uekane, K.; Hagihara, M. A crystalline antibacterial substance from Penicillium leucopus and four other strains of Penicillia, and Aspergillus clavatus, and its probable identity with patulin. Jpn. J. Med. 1947, 1, 97–100. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–242. [Google Scholar]
- Ciegler, A. Patulin. In Mycotoxins in Human and Animal Health; Rodricks, J.V., Hesseltine, C.W., Mehlman, M.A., Eds.; Pathotox Publishers Inc: Park Forest South, IL, USA, 1977; pp. 609–624. [Google Scholar]
- Barta, J.; Mecir, R. Antibacterial activity of Penicillium divergens Bainier. Experienfia 1948, 4, 277–278. [Google Scholar] [CrossRef]
- Fang-Chao, X.; Chun-Lin, L.; Xiang-Zheng, Y.; Yi-Lun, C. Isolation of main patulin-producing strains from storage pears and their ITS sequences analysis and identification. Food Ferment. Ind. 2011, 37, 54–57. [Google Scholar]
- Varga, J.; Due, M.; Frisvad, J.C.; Samson, R.A. Taxonomic revision of Aspergillus section Clavati based on molecular, morphological and physiological data. Stud. Mycol. 2007, 59, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.; Mateo, R.; Querol, A.; Huerta, T.; Hernández, E. Mycotoxins and mycotoxigenic moulds in nuts and sunflower seeds for human consumption. Mycopathologia 1991, 115, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.; Dombrink-Kurtzman, M.A. Patulin. In Microbiology of Fruits and Vegetables; Sapers, G.M., Gorny, J.R., Yousef, A.E., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 281–311. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Varga, J.; Frisvad, J.C. Polyphasic taxonomy of the heat resistant ascomycete genus Byssochlamysand its Paecilomycesanamorphs. Persoonia 2009, 22, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Rovira, R.; Ribera, F.; Sanchis, V.; Canela, R. Improvements in the quantitation of patulin in apple juice by high performance liquid chromatography. J. Agric. Food Chem. 1993, 41, 214–216. [Google Scholar] [CrossRef]
- Sanderson, P.G.; Spotts, R.A. Postharvest decay of winter pear and apple fruit caused by species of Penicillium. Phytopathology 1995, 85, 103–110. [Google Scholar] [CrossRef]
- Morales, H.; Marin, S.; Rovira, A.; Ramos, A.J.; Sanchis, V. Patulin accumulation in apples by Penicillium expansum during postharvest stages. Lett. Appl. Microbiol. 2007, 44, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Sommer, N.F.; Buchanan, J.R.; Fortlage, R.J. Production of patulin by Penicillium expansum. Appl. Microbiol. 1974, 28, 589–593. [Google Scholar] [PubMed]
- European Union. Commission Regulation No. 1425/2003 Amending Commission Regulation No.1881/2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. European Union: Brussels, Belgium, 2006. [Google Scholar]
- Börner, H. The formation of antibiotic and antiphytotoxic compounds in soil and their possible use for the self-incompatibility of apples in the nursery. Mitt. Biol. Bundesanst. Land Forstwirt. Berlin-Dahlem 1963, 108, 27–29. [Google Scholar]
- McCalla, T.M.; Guenzi, W.D.; Norstadt, F.A. Microbial studies of phytotoxic substances in the stubble-mulch system. Z. Allg. Mikrobiol. 1963, 3, 202–210. [Google Scholar] [CrossRef]
- Ellis, J.R.; McCalla, T.M. Effects of patulin and method of application on growth stages of wheat. Appl. Microbiol. 1973, 25, 562–566. [Google Scholar] [PubMed]
- Liu, B.H.; Wu, T.S.; Yu, F.Y.; Su, C.C. Induction of oxidative stress response by the mycotoxin patulin in mammalian cells. Toxicol. Sci. 2007, 95, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Polacco, J.C.; Sands, D.C. The mycotoxin patulin inhibits respiration of higher plant cells. Plant Sci. Lett. 1977, 9, 121–128. [Google Scholar] [CrossRef]
- Fliege, R.; Metzler, M. The mycotoxin patulin induces intra- and intermolecular protein cross links in vitro involving cysteine, lysine, and histidine side chains, and α-amino groups. Chem. Biol. Interact. 1999, 123, 85–103. [Google Scholar] [CrossRef]
- Ashoor, S.H.; Chu, F.S. Inhibition of alcohol and lactic dehydrogenases by patulin and penicillic acid in vitro. Food Cosmet. Toxicol. 1973, 11, 617–624. [Google Scholar] [CrossRef]
- Ashoor, S.H.; Chu, F.S. Inhibition of muscle aldolase by penicillic acid and patulin in vitro. Food Cosmet. Toxicol. 1973, 11, 995–1000. [Google Scholar] [CrossRef]
- Alsberg, G.L.; Black, O.F. Contributions to the study of maize deterioration: Biochemical and toxicological investigations of Penicillium puberulum and P. stoloniferum. U.S. Dep. Agric. Bureau Plant Ind. Bull. 1913, 270, 1–47. [Google Scholar]
- Oxford, A.E.; Raistrick, H. Studies in the biochemistry of micro-orgaisms. I- Erythritol, a metabolic product of Penicillium brevi-compactum Dierckx and P. cyclopium Westling. Biochem. J. 1935, 29, 1599. [Google Scholar] [PubMed]
- Birkinshaw, J.H.; Oxford, A.E.; Raistrick, H. Studies in the biochemistry of microorganisms. XL VIII. Penicillic acid, a metabolite product of Penicillium puberulum Bainier and P. cyclopium Westling. Biochem. J. 1936, 30, 394–411. [Google Scholar] [PubMed]
- Oxford, A.E.; Raistrick, H.; Smith, G. Anti-bacterial substances from moulds. Part II. Penicillic acid, a metabolic product of Penicillium puberulum Bainier and Penicillium cyclopium Westling. Chem. Ind. 1942, 10, 22–24. [Google Scholar]
- Bentley, R.; Keil, J.G. Tetronic acid biosynthesis in molds II. Formation of penicillic acid in Penicillium cyclopium. J. Biol. Chem. 1962, 237, 867–873. [Google Scholar] [PubMed]
- Shaw, E. A synthesis of protoanemonin; the tautomerism of acetylacrylic acid and of penicillic acid. J. Am. Chem. Soc. 1946, 68, 2510–2513. [Google Scholar] [CrossRef] [PubMed]
- Macri, A.; Dancea, Z.; Baba, A.I. Mycotoxin Involvement in Oncogenesis: Bibliographical Investigations; Buletinul Universitii de stiinte Agricole si Medicina Veterinara Cluj Nopoca Seria Medicina Veterinara: Cluj-Nopoca, Romania, 2002; Volume 58, pp. 640–644. [Google Scholar]
- He, J.; Wijeratne, E.M.K.; Bashyal, B.P.; Zhan, X.J.; Seliga, C.J.; Liu, M.X.; Pierson, E.E.; Pierson, L.S., III; van Etten, H.D.; Gunatilaka, A.A.L. Cytotoxic and other metabolites of Aspergillus inhibiting the rhizosphere of Sonoran desert plants. J. Nat. Prod. 2004, 67, 1985–1991. [Google Scholar] [CrossRef] [PubMed]
- Ezzat, S.M.; El-Sayed, A.E.; Abou El-Hawa, M.I.; Ismaiel, A.A. Morphological and ultrastructural studies for the biological action of penicillic acid on some bacterial species. Res. J. Microbiol. 2007, 2, 303–314. [Google Scholar]
- Nonaka, K.; Chiba, T.; Suga, T.; Asami, Y.; Iwatsuki, M. Coculnol, a new penicillic acid produced by a coculture of Fusarium solani FKI-6853 and Talaromyces sp. FKA-65. J. Antibiot. 2015. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, A.A. Effect of Acidic Microbial Toxins on Morphology and Ultrastructure of Bacteria. Ph.D. Thesis, Zagazig University, Zagazig, Egypt, September 2007. [Google Scholar]
- Bresler, G.; Brizzio, S.B.; Vaamonde, G. Mycotoxin-producing potential of fungi isolated from amaranth seeds in Argentina. Int. J. Food Microbiol. 1995, 25, 101–108. [Google Scholar] [CrossRef]
- Kang, S.W.; Park, C.H.; Hong, S.I.; Kim, S.W. Production of penicillic acid by Aspergillus sclerotiorum CGF. Biores. Technol. 2007, 98, 191–197. [Google Scholar] [CrossRef] [PubMed]
- SohYoung, O.; IllMin, C.; SuBong, P.; Seunghum, Y. Survey and control of the occurrence of mycotoxins from post harvest. I. Mycotoxins produced by Penicillium isolates from corn and wheat cereals. Korean J. Plant Pathol. 1998, 14, 700–704. [Google Scholar]
- Gedek, P.; Bauer, J.; Schreiber, H. Mycotoxin production of silage deteriorating molds. Wien Tierarztl Monatsschr 1981, 68, 299–301. [Google Scholar]
- Lund, F.; Frisvad, J.C. Chemotaxonomy of Penicillium aurantiogriseum and related species. Mycol. Res. 1994, 98, 481–492. [Google Scholar] [CrossRef]
- Hesseltine, C.W. Natural occurrence of mycotoxins in cereals. Mycopathol. Mycol. Appl. 1974, 53, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Lindenfelser, L.A.; Ciegler, A. Penicillic acid production in submerged culture. Appl. Environ. Microbiol. 1977, 34, 553–556. [Google Scholar] [PubMed]
- Andersen, B. Consistent production of phenolic compounds by Penicillium brevicompactum for chemotaxonomic characterization. Antonie Leuwenhoek 1991, 60, 115–123. [Google Scholar] [CrossRef]
- Tuthill, D.E.; Frisvad, J.C. Eupenicillium bovifimosum, a new species from dry cow manure in Wyoming. Mycologia 2002, 94, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Garza, S.; Canela, R.; Vinas, I.; Sanchis, V. Effects of potassium sorbate on growth and penicillic acid production by Aspergillus ochraceus and Penicillium aurantiogriseum. Zentralbl Mikrobiol. 1993, 148, 343–350. [Google Scholar] [PubMed]
- Ciegler, A. Bioproduction of ochratoxin A and penicillic acid by members of the Aspergillus ochraceus group. Can. J. Microbiol. 1972, 18, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Negishi, R.; Nagai, H.; Dmitrnok, A.; Kobyahi, H. Three new chlorine containing antibiotics from a marine-derived fungus Aspergillus ostianus collected in Pohnpei. J. Antibiot. (Tokyo) 2003, 56, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Bastin, R.; van Roey, G. Action of some antibiotics and inhibiting substances on cellular stretching. P. Pharm. Belg. 1954, 9, 112–127. [Google Scholar]
- Reiss, J. Inhibitory action of the mycotoxins patulin and penicillic acid on urease. Food Cosmet. Toxicol. 1979, 17, 145–146. [Google Scholar] [CrossRef]
- Tashiro, F.; Hirai, K.; Ueno, Y. Inhibitory effects of carcinogenic mycotoxins on deoxyribonucleic acid-dependent ribonucleic acid polymerase and ribonuclease H. Appl. Environ. Microbiol. 1979, 38, 191–196. [Google Scholar] [PubMed]
- Ciegler, A.; Mintzlaff, H.J.; Weisleder, D.; Leistner, L. Potential production and detoxification of penicillic acid in mold-fermented sausage (Salami). Appl. Microbiol. 1972, 24, 114–119. [Google Scholar] [PubMed]
- Sassa, T.; Hayakari, S.; Ikeda, M.; Miura, Y. Plant growth inhibitors produced by fungi. I. Isolation and identification of penicillic acid and dihydropenicillic acid. Agr. Biol. Chem. 1971, 35, 2130–2131. [Google Scholar] [CrossRef]
- Martínez-Luis, S.; González, M.C.; Ulloa, M.; Mata, R. Phytotoxins from the fungus Malbranchea aurantiaca. Phytochemistry 2005, 66, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Eqbal, M.M. Effect of penicillic acid on some biochemical changes in germinating maize seeds. Geobios 2002, 29, 161–163. [Google Scholar]
- Fondy, T.P.; Everse, J.; Driscou, G.A.; Castillo, F.; Stolzenbach, F.E.; Kaplan, N.O. The comparative enzymology of lactic dehydrogenases. IV. Function of sulfhydryl groups in lactic dehyrogenases and the sequence around the essential group. J. Biol. Chem. 1965, 240, 4219–4234. [Google Scholar] [PubMed]
- Ciegler, A.; Kurtzman, C.P. Penicillic acid production by blue-eye fungi on various agricultural commodities. Appl. Microbiol. 1970, 20, 761–764. [Google Scholar] [PubMed]
- Kurtzman, C.P.; Ciegler, A. Mycotoxin from a blue-eye mold of corn. Appl. Microbiol. 1970, 20, 204–207. [Google Scholar] [PubMed]
- Van Der Merwe, K.J.; Steyne, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- Horie, Y. Productivity of ochratoxin A of Aspergillus carbonarius in Aspergillus section Nigri. Nippon Kingakukai Kaiho 1995, 36, 73–76. [Google Scholar]
- Abarca, M.L.; Bragulat, M.R.; Sastella, G.; Cabanes, F.J. Ochratoxin A production by strains of Aspergillus niger var niger. Appl. Environ. Microbiol. 1994, 60, 2650–2652. [Google Scholar] [PubMed]
- Ciegler, A.; Fennell, D.J.; Mintzlaff, H.J.; Leistner, L. Ochratoxin synthesis by Penicillium species. Naturwissenschaften 1972, 59, 365–366. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; González-Salgado, A.; González-Jaén, M.A.T.; Vázquez, C.; Patiño, B. ITS-based detection and quantification of Aspergillus ochraceus and Aspergillus westerdijkiae in grapes and green coffee beans by real-time quantitative PCR. Int. J. Food Microbiol. 2009, 131, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Lund, F.; Frisvad, J.C. Penicillium verrucosum in wheat and barley indicates presence of ochratoxin A. J. Appl. Microbiol. 2003, 95, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Bogs, C.; Battilani, P.; Geisen, R. Development of a molecular detection and differentiation system for ochratoxin A producing Penicillium species and its application to analyse the occurrence of Penicillium nordicum in cured meats. Int. J. Food Microbiol. 2006, 107, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Beardall, J.; Miller, J.D. Natural occurrence of mycotoxins other than aflatoxin in Africa, Asia and South America. Mycotox. Res. 1994, 10, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.J.; White, S.; Dobson, A.D.W. Mycotoxins in agriculture and food safety. Ir. J. Agr. Food Res. 2000, 39, 235–244. [Google Scholar]
- Wang, Y.; Peng, X.; Xu, W.; Luo, Y.; Zhao, W. Transcript and protein profiling analysis of OTA-induced cell death reveals the regulation of the toxicity response process in Arabidopsis thaliana. J. Exp. Bot. 2011, 29, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.L.; Xu, W.T.; Wang, Y.; Huang, K.L.; Liang, Z.H.; Zhao, W.W.; Luo, Y.B. Mycotoxin ochratoxin A-induced cell death and changes in oxidative metabolism of Arabidopsis thaliana. Plant Cell Rep. 2010, 29, 153–161. [Google Scholar] [CrossRef] [PubMed]
- McLean, M. The phytotoxicity of selected mycotoxins on mature, germinating Zea mays embryos. Mycopathologia 1996, 132, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Ringot, D.; Chango, A.; Schneider, Y.; Larondelle, Y. Toxicokinetics and toxicodynamics of ochratoxin A, an update. Chem. Biol. Interact. 2006, 159, 18–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, W.; Hao, J.; Xu, W.; Luo, Y.; Wu, W. Changes in biosynthesis and metabolism of glutathione upon ochratoxin A stress in Arabidopsis thaliana. Plant Physiol. Biochem. 2014, 79, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Rosett, T.; Sankhala, R.H.; Stickings, C.E.; Taylor, M.E.U.; Thomas, R. Biochemistry of microorganisms. CIII. Metabolites of Alternaria tenuis auct: Culture filtrate products. Biochem. J. 1957, 67, 390–400. [Google Scholar] [PubMed]
- Stickings, C.E. Studies in the biochemistry of micro-organisms. 106. Metabolites of Alternaria tenius auct: The structure of tenuazonic acid. Biochem. J. 1959, 72, 332–340. [Google Scholar] [PubMed]
- Nolte, M.J.; Steyn, P.S.; Wessels, P.L. Structural investigations of 3-acylpyrrolidine-2,4-diones by nuclear magnetic resonance spectroscopy and X-ray crystallography. J. Chem. Soc. Perkin Trans. 1 1980. [Google Scholar] [CrossRef]
- Mikula, H.; Horkel, E.; Hans, P.; Hametner, C.; Fröhlich, J. Structure and tautomerism of tenuazonic acid—A synergetic computational and spectroscopic approach. J. Hazard. Mater. 2013, 250–251, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Shigeura, H.T.; Gordon, C.N. The biological activity of tenuazonic acid. Biochemistry 1963, 2, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Aver’yanov, A.A.; Lapikova, V.P.; Lebrun, M.H. Tenuazonic Acid, Toxin of Rice Blast Fungus, Induces Disease Resistance and Reactive Oxygen Production in Plants. Russ. J. Plant Pathol. 2007, 54, 749–754. [Google Scholar] [CrossRef]
- Kinoshita, T.; Renbutsu, Y.; Khan, I.D.; Kohmojo, K.; Nishimura, S. Distribution of tenuazonic acid production in the genus Alternaria and its pathological evaluation. Ann. Phytopathol. Soc. (Jpn.) 1972, 38, 397–404. [Google Scholar] [CrossRef]
- Umetsu, N.; Kaji, J.; Tamari, K. Investigation on the toxin production by several blast fungus strains and isolation of tenuazonic acid as a novel toxin. Agric. Biol. Chem. 1972, 36, 859–866. [Google Scholar] [CrossRef]
- Umetsu, N.; Kaji, J.; Aoyama, K.; Tamari, K. Toxins in blast diseased rice plants. Agri. Biol. Chem. 1974, 38, 1867–1874. [Google Scholar] [CrossRef]
- Davis, N.D.; Diener, U.L.; Morgan-Jones, G. Tenuazonic acid production Alternaria alternata and Alternaria tenuissima isolated from cotton. Appl. Environ. Microbiol. 1977, 34, 155–159. [Google Scholar] [PubMed]
- Steyn, P.S.; Rabie, C.J. Characterization of magnesium and calcium tenuazonate from Phoma sorghina. Phytochemistry 1976, 15, 1977–1979. [Google Scholar] [CrossRef]
- Janardhanan, K.K.; Husain, A. Phytotoxic activity of tenuazonic acid isolated from Alternaria alternata (Fr.) Keissler causing leaf blight of Datura innoxia Mill. and its effect on host metabolism. J. Phytopathol. 1984, 111, 305–311. [Google Scholar] [CrossRef]
- Lebrun, M.H.; Nicolas, L.; Boutar, M.; Gaudemer, F.; Ranomenjanahary, S.; Gaudemer, A. Relationships between the structure and the phytotoxicity of the fungal toxin tenuazonic acid. Phytochemistry 1988, 27, 77–84. [Google Scholar] [CrossRef]
- Carrasco, L.; Vazquez, D. Survey of inhibitors in different steps of protein synthesis by mammalian ribosomes. J. Antibiot. 1972, 25, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Barbacid, M.; Vazquez, D. (3H) anisomycin binding to eukaryotic ribosomes. J. Mol. Biol. 1974, 84, 603–623. [Google Scholar] [CrossRef]
- Barbacid, M.; Vazquez, D. Binding of [acetyl-14C] trichodermin to the peptidyl transferase centre of eukaryotic ribosomes. Eur. J. Biochem. 1974, 44, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Park, P. Effect of the host-specific toxin and other toxic metabolites by Alternaria kikuchiana on ultrastructure of leaf cells of Japanese pear. Ann. Phytopath. Soc. (Jpn.) 1977, 43, 15–25. [Google Scholar] [CrossRef]
- Binder, M.; Tamm, C.; Turner, W.B.; Minato, H. Nomenclature of a class of biologically active mould metabolites: The cytochalasins, phomins, and zygosporins. J. Chem. Soc. Perkin Trans. 1 1973, 11, 1146–1147. [Google Scholar] [CrossRef] [PubMed]
- Rothweiler, W.; Tamm, C. islation and structure of phomin. Expertientia 1966, 22, 750–752. [Google Scholar] [CrossRef]
- Rothweiler, W.; Tamm, C. Isolierung and struktur der Antibiotica phomin und 5-Dehydrophomin. Helv. Chim. Acta 1970, 53, 696–724. [Google Scholar] [CrossRef] [PubMed]
- Aldridge, D.C.; Armstrong, J.J.; Speake, R.N.; Turner, W.B. The structure of cytochalasins A and B. J. Chem. Soc. C 1967. [Google Scholar] [CrossRef]
- Cole, R.J.; Jarvis, B.B.; Schweikert, M.A. Handbook of Secondary Fungal Metabolites; Academic Press: San Diego, CA, USA, 2003; Chapters 14–20; Volume III, pp. 199–560. [Google Scholar]
- Lindholm, C.; Norppa, H.; Hayashi, M.; Sorsa, M. Induction of micronuclei and anaphase aberrations by cytochalasin B in human lymphocyte cultures. Mutat. Res. 1991, 260, 369–375. [Google Scholar] [CrossRef]
- Fenech, M.; Morley, A.A. Measurement of micronuclei inlymphocytes. Mutat. Res. 1985, 147, 29–36. [Google Scholar] [CrossRef]
- Falck, G.; Catalan, J.; Norppa, H. Influence of culture time on the frequency and contents of human lymphocytes micronuclei with and without cytochalasin B. Mutat. Res. 1997, 392, 71–79. [Google Scholar] [CrossRef]
- Minissi, S.; Gustavino, B.; Degrassi, F.; Tanzarela, C.; Rizzoni, M. Effect of cytochalasin B on the induction of chromosome missegregation by colchicine at low concentrations in human lymphocytes. Mutagenesis 1999, 14, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.I.; Liu, H.F. The blockage of survivin and securin expression increases the cytochalasin B-induced cell death and growth inhibition in human cancer cells. Mol. Pharmacol. 2006, 69, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, A.A. Morphogenesis and Physiological Activity of Some Microorganisms as Affected by the Mycotoxin Cytochalasin. Master Thesis, Zagazig University, Zagazig, Egypt, March 2005. [Google Scholar]
- Scott, P.M.; Harwig, J.; Chen, Y.K.; Kennedy, B.P.C. Cytochalasins A and B from strains of Phoma exigua var. exigua and formation of cytochalasin B in potato gangrene. J. Gen. Microbiol. 1975, 87, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Capasso, R.; Evidente, A.; Ritieni, A.; Radazzo, G.; Vurro, M.; Bottalico, A. Ascochalasin, a new cytochalasin from Ascochyta heteromorpha. J. Nat. Prod. 1988, 51, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Capio, E.R.; Tate, M.E.; Wallwork, H. Phytotoxic metabolites from Drechslera wirreganensis and D. campanulata. Aust. Plant Pathol. 2004, 33, 23–28. [Google Scholar] [CrossRef]
- Carlier, M.F.; Ciquet, P.; Pantalont, D.; Korn, E.D. Interaction of cytochalasins D with actin filaments in the presence of ADP and ATP. J. Biol. Chem. 1986, 261, 2041–2050. [Google Scholar] [PubMed]
- Zholkevich, V.N.; Chugunova, T.W. Interaction between cytoskeletal proteins, biomediators and phytohormones in the regulation of water transport in plants. Doklady Biol. Sci. 1995, 341, 163–165. [Google Scholar]
- Seagull, R.W. Cytoskeletal stability affects fiber initiation. Int. J. Plant Sci. 1998, 159, 590–598. [Google Scholar] [CrossRef]
- Decognet, V.; Citharel, J. Production of cytochalasins by Phoma exigua var. linicola. Asp. Appl. Biol. 1991, 28, 107–110. [Google Scholar]
- Huang, R.F.; Wang, X.C. Effects of cytochalasin B on stomatal movement of Vicia faba. Acta Phytophysiol. Sin. 1996, 22, 404–408. [Google Scholar]
- Teng, X.Y.; Zhuang, L.X. The role of actin tip growth of pollen tube. Acta Phytophysiol. Sin. 1997, 23, 239–244. [Google Scholar]
- Kadota, A.; Wada, M. Cytoskeletal aspects of nuclear migration during tip growth in the fern Adiantum protonemal cell. Protoplasma 1995, 188, 170–179. [Google Scholar] [CrossRef]
- Stitt, A.W.; Fairweather, I.; Johnston, C.F. Fasciola hepatica: Disruption of spermatogenesis by the microfilament inhibitor cytochalasin B. Parasitol. Res. 1991, 77, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Tamagnini, R.P.; Romano, B.; Frenguelli, G.; Bisogni, A.M. Effect of cytochalasin B on germinating seeds of Psium sativum L. respiration, ultrastructure of cotyledon cells, storage substances, acid phosphatase activity. J. Botanico Italiano 1983, 117, 227–236. [Google Scholar] [CrossRef]
- Abdrakhimova, I.R.; Kholkhlova, L.P. Modifiers of cytoskeleton proteins and regulation of electron transport pathways of plant respiration. Doklady Biol. Sci. 2000, 372, 273–275. [Google Scholar]
- Thomas, D.; des, S.; Lager, N.M.; Manavathu, E.K. Cytochalasin B: Effect on root morphogenesis in Allium cepa. Can. J. Bot. 1973, 51, 2269–2273. [Google Scholar] [CrossRef]
- Eleftheriou, E.P.; Palevitz, B.A. The effect of cytochalasin D on preprophase band organization in root tip cells of Allium. J. Cell Sci. 1992, 103, 989–998. [Google Scholar]
- Behboodi, B.S.; Samadi, L. Mitotic apparatus movement and cell wall pattering in root apical meristem of Allium cepa. Pakistan J. Bot. 2002, 34, 405–410. [Google Scholar]
- Siugimoto, M.; Toyada, K.; Ichinose, Y.; Yamada, T.; Shiraishi, T. Cytochalasin A inhibits the binding of phenylalanine ammonia lyase mRNA to ribosomes during induction of phytoalexin in pea seedlings. Plant Cell Physiol. 2000, 411, 234–238. [Google Scholar] [CrossRef]
- Quader, H.; Liebe, S. Actin filament-independent formation of tubular endoplasmic reticulum in onion epidermis cell. J. Plant Physiol. 1995, 145, 71–77. [Google Scholar] [CrossRef]
- Gravis, C.; Newcomb, W.; Simmonds, D.H. Rearrangement of the actin filament and microtubule cytoskeleton during induction of microspore embryogenesis in Brassica napus L.C.V. topas. Protoplasma 2001, 213, 194–202. [Google Scholar] [CrossRef]
- Traas, J.A.; Burgain, S.; Dumas De vaul, R. The organization of the cytoskeleton during meiosis in egg plant (Solanum melongena (L.)): Microtubules and F-actin are both necessary for coordinated meiotic division. J. Cell Sci. 1989, 92, 541–550. [Google Scholar]
- Sawai, K.; Okuno, T.; Fujioka, H.; Furuyo, M. The relation between the phytotoxicity of cytochalasin E and its molecular structure. Ann. Phytopathol. Soc. Jpn. 1983, 49, 262–265. [Google Scholar] [CrossRef]
- Kanematsu, S.; Hayashi, T.; Kudo, A. Isolation of Rosellinia necatrix mutants with impaired cytochalasin E production and its pothogenicity. Ann. Phytopathol. Soc. Jpn. 1997, 63, 425–431. [Google Scholar] [CrossRef]
- Zonno, M.C.; Vurro, M. Effect of fungal toxins on germination of Striga hermonthica seeds. Weed Res. Oxf. 1999, 39, 15–20. [Google Scholar] [CrossRef]
- Kshirsagar, A.; Reid, A.J.; Maccol, S.M.; Saunders, V.A.; Whalky, A.J.S.; Evans, E.H. The effect of fungal metabolites on leaves as detected by chlorophyll fluorescence. New Phytol. 2001, 151, 451–457. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Vurro, M.; Zonno, M.C.; Matto, A. Cytochalasins Z1, Z2 and Z3, three 24-Oxa(14)cytochalasins produced by Pyrenophora semeniperda. Phytochemistry 2002, 60, 45–53. [Google Scholar] [CrossRef]
- Masi, M.; Meyer, S.; Cimmino, A.; Andolfi, A.; Evidente, A. Pyrenophoric Acid, a Phytotoxic sesquiterpenoid Penta-2,4-dienoic acid produced by a potential Mycoherbicide, Pyrenophora semeniperda. J. Nat. Prod. 2014, 77, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Evidente, A.; Meyer, S.; Nicholson, J.; Munoz, A. Effect of strain and cultural conditions on the production of cytochalasin B by the potential mycoherbicide Pyrenophora semeniperda (Pleosporaceae, Pleosporales). Biocontrol Sci. Technol. 2014, 24, 53–64. [Google Scholar] [CrossRef]
- Nukina, M. The blast disease fungi and their metabolic products. J. Pestic. Sci. 1999, 24, 239–298. [Google Scholar] [CrossRef]
- Fujii, Y.; Tani, H.; Ichinoe, M.; Nakajima, H. Zygosporin D and Two New Cytochalasins Produced by the fungus Metarrhizium anisopliae. J. Nat. Prod. 2000, 63, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.J.; Cox, R.H. The trichothecenes. In Handbook of Toxic Fungal Metabolites; Academic Press: Toronto, ON, Canada, 1981; pp. 152–263. [Google Scholar]
- Wilkins, K.; Nielsen, K.; Din, S. Patterns of volatile metabolites and nonvolatile trichothecenes produced by isolates of Stachybotrys, Fusarium, Trichoderma, Trichothecium and Memnoniella. Environ. Sci. Pollut. Res. 2003, 10, 162–166. [Google Scholar] [CrossRef]
- Trapp, S.C.; Hohn, T.M.; McCormick, S.; Jarvis, B.B. Characterization of the gene cluster for biosynthesis of macrocyclic trichothecenes in Myrothecium roridum. Mol. Gen. Genet. 1998, 257, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, B.B.; Mazzola, E.P. Macrocyclic and other novel trichothecenes: Their structure, synthesis, and biological significance. Acc. Chem. Res. 1982, 15, 388–395. [Google Scholar] [CrossRef]
- Freeman, G.G.; Morrison, R.I. Trichothecin: An antifungal metabolic product of Trichothecium roseum Link. Nature 1948, 162, 30. [Google Scholar] [CrossRef] [PubMed]
- Brian, P.W.; Dawkins, A.W.; Grove, J.F.; Hemming, H.G.; Lowe, D.; Norris, G.L.F. Phytotoxic compounds produced by Fusarium equiseti. J. Exp. Bot. 1961, 12, 1–12. [Google Scholar] [CrossRef]
- Tatsuno, T.M.; Saito, M.; Enomoto, M.; Tsunoda, H. Nivalenol, a toxic principle of Fusarium nivale. Chem. Pharm. Bull. 1968, 16, 2519–2520. [Google Scholar] [CrossRef] [PubMed]
- Bamburg, J.R.; Riggs, N.V.; Strong, F.M. The structures of toxins from two strains of Fusarium tricinctum. Tetrahedron 1968, 24, 3329. [Google Scholar] [CrossRef]
- Marasus, W.F.O.; Nelson, P.E.; Toussoun, T.A. Toxigenic Fusarium Species; Pennsylvania State University Press: University Park, PA, USA, 1984. [Google Scholar]
- Vesonder, R.F.; Hesseltine, C.W. Vomitoxin: Natural occurrence on cereal grains and significance as a refusal and emetic factor to swine. Process Biochem. 1980, 44, 12–14. [Google Scholar]
- Savard, M.E.; Blackwell, B.A.; Greenhalgh, R. An 1H nuclear magnetic resonance study of derivatives of 3-hydroxy-12,13-epoxytrichothec-9-enes. Can. J. Chem. 1987, 65, 2254–2262. [Google Scholar] [CrossRef]
- Savard, M.E.; Blackwell, B.A. Spectral characteristics of secondary metabolites from Fusarium Fungi. In Mycotoxins in Grain; Miller, J.D., Trenholm, H.L., Eds.; Eagen Press: St. Paul, MI, USA, 1994; pp. 59–228. [Google Scholar]
- Jarvis, B.B.; Mazzocchi, D.B.; Ammon, H.L.; Mazzola, E.P.; Flippen-Anderson, J.L.; Gilardi, R.D.; George, C.F. Conformational effects in trichothecenes: Structures of 15-hydroxy C4 and C8 ketones. J. Org. Chem. 1990, 55, 3660–3662. [Google Scholar] [CrossRef]
- Shank, R.A.; Foroud, N.A.; hazendonk, P.; Eudes, F.; Blackwell, A. Current and future experimental strategies for structural analysis of trichothecene mycotoxins-A prospectus. Toxins 2011, 3, 1518–1553. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, R.; Blackwell, B.A.; Savard, M.E. The NMR spectra of trichothecenes and related fungal metabolites. Tetrahedron 1989, 45, 2373–2383. [Google Scholar] [CrossRef]
- Schollenberger, M.; Drochner, W.; Müller, H.-M. Fusarium toxins of the scirpentriol subgroup: A review. Mycopathologia 2007, 164, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y. Toxicological features of T-2 toxin and related trichothecenes. Toxicol. Sci. 1984, 4, S124–S132. [Google Scholar] [CrossRef]
- Jarvis, B.B.; Midiwo, J.O.; Bean, G.A.; Aboul-Nasr, M.B.; Barnos, C.S. The mystery of trichothecene antibiotics in Baccharis species. J. Nat. Prod. 1988, 51, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, M.D.; Romero, N.; Reddy, P.V.; White, J.F., Jr. A hypocrealean epibiont on meristems of Baccharis coridifolia. Mycologia 1997, 89, 375–382. [Google Scholar] [CrossRef]
- Ueno, Y. Trichothecenes: Chemical, Biological, and Toxicological Aspects; Elsevier: Amsterdam, The Nerthlands, 1983. [Google Scholar]
- Cutler, H.G. Trichothecenes and their role in the expression of plant disease. In Biotechnology for Crop Protection; Hedin, P.A., Menn, J.J., Hollingworth, R.M., Eds.; Maple Press: York, PA, USA, 1988; pp. 50–72. [Google Scholar]
- Cheeke, P.R. Natural Toxicant in Feeds, Forages, and Poisonous Plants; Interstate Publishers Inc.: Danville, IL, USA, 1998. [Google Scholar]
- CAST (Council for Agriculture Science and Technology). Mycotoxins Risks in Plant, Animal, and Human Systems; CAST: Ames, IA, USA, 2003. [Google Scholar]
- Harris, L.J.; Desjardins, A.E.; Plattner, R.D.; Nicholson, P.; Butler, G.; Young, J.C.; Weston, G.; Proctor, R.H.; Holn, T.M. Possible role of trichothecenes mycotoxins in virulence of Fusarium graminearum on maize. Plant Dis. 1999, 83, 954–960. [Google Scholar] [CrossRef]
- Wang, H.; Hwang, S.F.; Eudes, F.; Chang, K.F.; Howard, R.J.; Turnbull, G.D. Trichothecenes and aggressiveness of Fusarium graminaearum causing seeding blight and root rot in cereals. Plant Pathol. 2006, 55, 224–230. [Google Scholar] [CrossRef]
- Abbas, H.K.; Johnson, B.B.; Shier, W.T.; Tak, H.; Jarvis, B.B.; Boyette, C.D. Phytotoxicity and mammalian cytotoxicity of macrocyclic trichothecene mycotocins from Myrothecium verrucaria. Phytochemistry 2002, 59, 309–313. [Google Scholar] [CrossRef]
- Zonno, M.C.; Vurro, M. Inhibition of germination of Orobanche ramose seeds by Fusarium toxins. Phytoparasitica 2002, 30, 519–524. [Google Scholar] [CrossRef]
- Abbas, H.K.; Duke, S.O.; Merrill, A.H., Jr.; Wang, E.; Shier, W.T. Phytotoxicity of australifungin, AAL-toxins and fumonisin B 1 to Lemna pausicostata. Phytochemistry 1998, 47, 1509–1514. [Google Scholar] [CrossRef]
- Morin, L.; Gianotti, A.F.; Lauren, D.R. Trichothecene production and pathogenicity of Fusarium tumidum, a candidate bioherbicides for gorse and broom in New Zealand. Mycol. Res. 2000, 104, 993–999. [Google Scholar] [CrossRef]
- Tanaka, T.; Abbas, H.K.; Duke, S.O. Structure–dependent phytotoxicity of fumonisins, and related compound in a duckweed bioassay. Phytochemistry 1993, 33, 779–785. [Google Scholar] [CrossRef]
- Vesonder, R.F.; Golinski, P. Metabolites of Fusarium. In Fusarium Mycotoxins, Taxonomy and Pathogenicity; Chelkowski, J., Ed.; Elsevier Publishers: Amsterdam, The Nertanlands, 1989; pp. 1–39. [Google Scholar]
- Atanassov, Z.; Nakamura, C.; Mori, N.; Kaneda, C.; Kato, H.; Jin, Y.-Z.; Yoshizawa, T.; Murai, K. Mycotoxin production and pathogenicity of Fusarium species and wheat resistance to Fusarium head blight. Can. J. Bot. 1994, 72, 161–167. [Google Scholar] [CrossRef]
- Snijders, C.H.A.; Krechting, C.F. Inhibition of deoxynivalenol translocation and fungal colonization in Fusarium head blight resistant wheat. Can. J. Bot. 1992, 70, 1570–1576. [Google Scholar] [CrossRef]
- Wakulinski, W. Phytotoxicity of the secondary metabolites of fungi causing wheat head fusariosis (head blight). Acta Physiol. Plant 1989, 11, 301–306. [Google Scholar]
- Rocha, L.G.; Almeida, J.R.G.S.; Macedo, R.O.; Barbosa-Filho, J.M. A review of natural products with antileishmanial activity. Phytomedicine 2005, 12, 514–535. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Otani, M. Effects of Fusarium mycotoxins on the growth of shoots and roots at germination in some Japanese wheat cultivars. Cereal Res. Commun. 1990, 18, 229–232. [Google Scholar]
- Cossette, F.; Miller, J.D. Phytotoxic effect of deoxynivalenol and gibberella ear rot resistance of corn. Nat. Toxins 1995, 3, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Bottalico, A.; Leario, P.; Visconti, A. Some experimental data on the phytotoxicity of mycotoxins. Phytopath. Medit. 1980, 19, 196–198. [Google Scholar]
- Bandurska, H.; Chelkowski, J.; Wisniewska, H. Free proline accumulation in wheat seedlings influenced by Fusarium culmorum infection and the pathogen metabolite deoxynivaienol. Acta Physiol. Plant 1994, 16, 111–116. [Google Scholar]
- Menke-Milczarek, I.; Zimny, J. Phytotoxicity of deoxynivalenol to wheat calli. Mycotox. Res. 1991, 7, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Packa, D. Cytogenetic changes in plant cells as influenced by mycotoxins. Mycotox. Res. 1991, 7, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.F.; Bilgrami, K.S.; Masood, A. Cytotoxic effects of DON and T-2 toxin on plant cells. Mycopathologia 1993, 124, 95–97. [Google Scholar] [CrossRef]
- Miller, J.D. Effects of Fusarium graminearum metabolites on wheat cells. In Phytotoxins and Plant Pathogenesis; Granti, A., Durbin, R.D., Ballio, A., Eds.; NATO ASI Series, Springer Netherlands: Heidelberg, Germany, 1989; pp. 449–452. [Google Scholar]
- Snijders, C.H.A. Breeding for resistance to Fusarium in wheat and maize. In Mycotoxins in Grain. Compounds other than Aflatoxin; Miller, J.D., Trenholm, H.L., Eds.; Eagan Press: St. Paul, MN, USA, 1994; pp. 37–58. [Google Scholar]
- Scott, P.M. The natural occurrence of trichothecenes. In Trichothecene Mycotoxins: Pathophysiologic Effects; Beasley, V.R., Ed.; CRC Press: Boca Raton, FL, USA, 1989; pp. 1–26. [Google Scholar]
- Miller, J.D. Epidemiology of Fusarium ear diseases in cereals. In Myeotoxins in Grain. Compounds Other than Aflatoxin; Miller, J.D., Trenholm, H.L., Eds.; Eagan Press: St. Paul, MN, USA, 1994; pp. 19–36. [Google Scholar]
- Thompson, W.L.; Wannemacher, R.W. Structure-function relationship of 12,13-epoxytrichothecene mycotoxins in cell culture: Comparison to whole animal lethality. Toxicon 1986, 24, 985–994. [Google Scholar] [CrossRef]
- Terse, P.S.; Madhyastha, M.S.; Zurovac, O.; Stringfellow, D.; Marquardt, R.R.; Kemppainen, B.W. Comparison of in vitro and in vivo biological activity of mycotoxins. Toxicon 1993, 31, 913–919. [Google Scholar] [CrossRef]
- Chang, N.T.; Xue, Y.L. Effect of T-2 toxin on wheat leaf protoplasts. Chin. Sci. Bull. 1990, 35, 431–434. [Google Scholar]
- Vesonder, R.F.; Labeda, D.P.; Peterson, R.E. Phytotoxie activity of selected water-soluble metabolites of Fusarium against Lemna minor L. (Duckweed). Mycopathologia 1992, 118, 185–189. [Google Scholar] [CrossRef]
- Cole, R.J.; Kirksey, J.W.; Cutler, H.G.; Doubnik, B.L.; Peckham, J.C. Toxin from Fusarium moniliforme: Effects on plants and animals. Science 1973, 179, 1324–1326. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, H.R.; Ciegler, A.; Vesonder, R.F. Moniliformin, a metabolite of Fusarium moniliforme NRRL 6322: Purification and toxicity. Appl. Environ. Microbiol. 1979, 37, 11–13. [Google Scholar] [PubMed]
- Burmeister, H.R.; Grove, M.D.; Kwolek, W.F. Moniliformin and butenolide: Effect on mice of high-level, long-term oral intake. Appl. Environ. Microbiol. 1980, 40, 1142–1144. [Google Scholar] [PubMed]
- Springer, J.P.; Clardy, J.; Cole, R.J.; Kirksey, J.W.; Hill, R.K.; Carlson, R.M.; Isidor, J.L. Structure and synthesis of moniliformin, a novel cyclobutane microbial toxin. J. Am. Chem. Soc. 1974, 96, 2267–2268. [Google Scholar] [CrossRef] [PubMed]
- Steyn, M.; Thiel, P.G.; van Schalkwyk, G.C. Isolation and purification of moniliformin. J. Assoc. Off. Anal. Chem. 1978, 61, 578–580. [Google Scholar] [PubMed]
- Booth, C. The Genus Fusarium; Commonwealth Mycological Institute: Kew, UK, 1971. [Google Scholar]
- Rable, C.J.; Lubben, A.; Louw, A.I.; Rathbone, E.B.; Steyn, P.S.; Vleggaar, R. Moniliformin, a mycotoxin from Fusarium fusarioides. J. Agric. Food Chem. 1978, 26, 375–379. [Google Scholar]
- Marasas, W.F.O.; Leistner, L.; Hofmann, G.; Eckardt, C. Occurrence of toxigenic strains of Fusarium in maize and barley in Germany. European J. Appl. Microbiol. Biotechnol. 1979, 7, 289–305. [Google Scholar] [CrossRef]
- Burka, L.T.; Doran, J.; Wilson, B.J. Enzyme inhibition and the toxic action of moniliformin and other vinylogous α-ketoacids. Biochem. Pharmacol. 1982, 31, 79–84. [Google Scholar] [CrossRef]
- Chen, L.Y.; Tian, X.L.; Yang, B. A study on the inhibition of rat myocardium glutathione peroxidase and glutathione reductase by moniliformin. Mycopathologia 1990, 110, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Pirrung, M.C.; Nauhaus, S.K.; Singh, B. Cofactor-directed, time-dependent inhibition of thiamine enzymes by the fungal toxin moniliformin. J. Org. Chem. 1996, 61, 2592–2593. [Google Scholar] [CrossRef] [PubMed]
- Deruiter, J.; Jacyno, J.M.; Cutler, H.G.; Davis, R.A. Studies on aldose reductase inhibitors from fungi. II. Moniliformin and small ring analogues. J. Enzyme Inhib. 1993, 7, 249–256. [Google Scholar] [CrossRef]
- Reams, R.; Thacker, H.L.; Novilla, M.; Laska, D.; Horn, J.; Harrington, D.; Greenlee, W.; Vesonder, R. Development of an L6 myoblast in vitro model of moniliformin toxicosis. Mycopathologia 1996, 133, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Kriek, N.P.; Marasas, W.F.; Steyn, P.S.; van Rensburg, S.J.; Steyn, M. Toxicity of a moniliformin-producing strain of Fusarium moniliforme var. subglutinans isolated from maize. Food Cosmet. Toxicol. 1977, 15, 579–587. [Google Scholar] [CrossRef]
- Uhlig, S.; Jestoi, M.; Parikka, P. Fusarium avenaceum—The north European situation. Int. J. Food Microbiol. 2007, 119, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Thiel, P.G.; Marasas, W.E.O.; Sydenham, E.W.; Shephard, G.S.; Gelderblom, W.C.A.; Nieuwenhuis, J.J. Survey of fumonisin production by Fusarium species. Appl. Environ. Microbiol. 1991, 57, 1089–1093. [Google Scholar] [PubMed]
- Rheeder, J.P.; Marasas, W.E.O.; Vismer, H.E. Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, W.C.A.; Jaskiewicz, K.; Marasas, W.F.; Thiel, P.G.; Horak, R.M.; Vleggaar, R.; Kreik, N.P.J. Fumonisins-novel mycotoxins with cancer-promoting activity produced by Fusarium moniliforme. Appl. Environ. Microbiol. 1988, 54, 1806–1811. [Google Scholar] [PubMed]
- Kellerman, T.S.; Marasas, W.F.O.; Thiel, P.G.; Gelderblom, W.C.A.; Cawood, M.; Coetser, J.A.W. Leukoencepahalomalacia in two horses induced by oral dosing of fumonisin B1, Ondersterpoort. J. Vet. Res. 1990, 57, 319–324. [Google Scholar]
- Harrison, L.R.; Colvin, B.M.; Greene, J.T.; Newman, L.E.; Cole, J.R. Pulmonary edema and hydrothorax in swine produced by fumonisin B1, a toxic metabolite of Fusarium moniliforme. J. Vet. Diagn. Investig. 1990, 2, 217–221. [Google Scholar] [CrossRef]
- Rheeder, J.P.; Marasas, W.F.O.; Thiel, P.G.; Sydenham, E.W.; Shephard, G.S.; van Schalkwyk, D.J. Fusarium moniliforme and fumonisins in corn in relation to human esophageal cancer in Transkei. Phytopathology 1992, 82, 353–357. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Yamashita, A.; Luo, Y. Fumonisin occurrence in corn from high- and low-risk areas for human oesophageal cancer in China. Appl. Enivron. Toxicol. 1994, 60, 1626–1629. [Google Scholar]
- Abbas, H.K.; Boyette, C.D.; Hoagland, R.E.; Vesonder, R.E. Bioherbicidal potential of Fusarium moniliforme and its phytotoxin, fumonisin B1. Weed Sci. 1991, 139, 673–677. [Google Scholar]
- Danielsen, S.; Jensen, D.E. Relationships between seed germination, fumonisin content, and Fusarium verticillioides infection in selected maize samples from different regions of Costa Rica. Plant Pathol. 1998, 47, 609–614. [Google Scholar] [CrossRef]
- Lamprecht, S.C.; Marasas, W.E.O.; Alberts, J.F.; Cawood, M.E.; Gelderblom, W.C.A.; Shephard, G.S. Phytotoxicity of fumonisins and TA-toxin to corn and tomato. Phytopathology 1994, 84, 383–391. [Google Scholar] [CrossRef]
- Abbas, H.K.; Boyette, C.D. Phytotoxicity of fumonisin B1 on weed and crop species. Weed Technol. 1992, 6, 548–552. [Google Scholar]
- Doehlert, D.C.; Knutson, C.A.; Vesonder, R.E. Phytotoxic effects of fumonisin B1 on maize seedling growth. Mycopathologia 1994, 127, 117–121. [Google Scholar] [CrossRef]
- Abbas, H.K.; Ocamb, C.M. First report of production of fumonisin B1 by Fusarium polyphialidicum collected from seeds of Pinus strobus. Plant Dis. 1995, 79, 642. [Google Scholar] [CrossRef]
- Jackowiak, H.; Packa, D.; Wiwart, M.; Perkowski, J.; Busko, M.; Borusiewicz, A. Scanning electron microscopy of mature wheat kernels infected with Fusarium culmorum. J. Appl. Genet. 2002, 43, 167–176. [Google Scholar]
- Pekkarinen, A.I.; Mannonen, L.; Jones, B.L.; Niku-Paavola, M.L. Production of proteases by Fusarium species grown on barley grains and in media containing cereal proteins. J. Cereal Sci. 2000, 31, 253–261. [Google Scholar] [CrossRef]
- Janardhan, A.; Subramanyam, D.; Praveen, A.; Reddi, M.; Narasimha, G. Aflatoxin impacts on germinating seeds. Ann. Biol. Res. 2011, 2, 180–188. [Google Scholar]
- Van Asch, M.A.J. Studies on the Resistance of Wheat and Maize to Fungal Pathogenesis. Ph.D. Thesis, University of Natal, Pietermaritzburg, South Africa, 1990. [Google Scholar]
- Abbas, H.K.; Gelderblom, W.C.A.; Cawood, M.E.; Shier, W.T. Biological activities of fumonisins, mycotoxins from Fusarium moniliforme, in jimsonweed (Datura stramonium L.) and mammalian cell cultures. Toxicon 1993, 31, 345–353. [Google Scholar] [CrossRef]
- Abbas, H.K.; Duke, S.O.; Tanaka, T. Phytotoxicity of fumonisins and related compounds. J. Toxicol. Toxin Rev. 1993, 12, 225–251. [Google Scholar] [CrossRef]
- Krasnoff, S.B.; Sommers, C.H.; Moon, Y.-S.; Donzelli, B.G.G.; Vandenberg, J.D.; Churchill, A.C.L. Production of mutagenic metabolites by Metarhizium anisopliae. J. Agric. Food Chem. 2006, 54, 7083–7088. [Google Scholar] [CrossRef] [PubMed]
- Wiebe, L.A.; Bjeldanes, L.F. Fusarin C, a mutagen from Fusarium moniliforme grown on corn. J. Food Sci. 1981, 46, 1424–1426. [Google Scholar] [CrossRef]
- Gelderblom, W.C.A.; Marasas, W.F.O.; Steyn, P.S.; Thiel, P.G.; Van Der Merwe, K.J.; Van Rooyen, P.H. Structure elucidation of fusarin C, a mutagen produced by Fusarium moniliforme. J. Chem. Soc. Chem. Commun. 1984, 7, 122–124. [Google Scholar] [CrossRef]
- Savard, M.E.; Miller, D.J. Characterization of fusarin F, a new fusarin from Fusarium moniliforme. J. Nat. Prod. 1992, 55, 64–70. [Google Scholar] [CrossRef]
- Kleigrewe, K.; Aydin, F.; Hogrefe, K.; Piecuch, P.; Bergander, K.; Würthwein, E. Structure elucidation of new fusarins revealing insights in the rearrangement mechanisms of the Fusarium mycotoxin fusarin C. J. Agric. Food Chem. 2012, 60, 5497–5505. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; Porter, J.K.; Norred, W.P.; Leslie, J.F. Production of fusaric acid by Fusarium species. Appl. Environ. Microbiol. 1996, 62, 4039–4043. [Google Scholar] [PubMed]
- Sieber, C.M.K.; von Bargen, K.W.; Studt, L.; Niehaus, E.-M.; Espino, J.J. Deciphering the cryptic genome: Genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLoS Pathol. 2013, 9, e1003475. [Google Scholar]
- Brown, D.W.; Butchko, R.A.E.; Busman, M.; Proctor, R.H. Identification of gene clusters associated with fusaric acid, fusarin, and perithecial pigment production in Fusarium verticillioides. Fungal Genet. Biol. 2012, 49, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.K.; Mirocha, C.J.; Kommedahl, T.; Vesonder, R.F.; Golinski, P. Production of trichothecene and non-trichothecene mycotoxins by Fusarium species isolated from maize in Minnesota. Mycopathologia 1989, 108, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; Marijanovic, D.R.; Norred, W.P.; Hinton, D.M. Production of fusarin C on cereal and soybean by Fusarium moniliforme. Appl. Environ. Microbiol. 1989, 55, 2745–2748. [Google Scholar] [PubMed]
- Tseng, T. Mycotoxins produced by Fusarium spp. of Taiwan. Bot. Bull. Acad. Sin. 1993, 34, 261–269. [Google Scholar]
- Gäumann, E. Fusaric acid as a wilt toxin. Phytopathology 1957, 47, 342–357. [Google Scholar]
- Stipanovic, R.D.; Wheeler, M.H.; Puckhaber, L.S.; Liu, J.; Bell, A.A.; Williams, H.J. Nuclear magnetic resonance (NMR) studies on the biosynthesis of fusaric acid from Fusarium oxysporum f. sp. vasinfectum. J. Agric. Food Chem. 2011, 59, 5351–5356. [Google Scholar] [CrossRef] [PubMed]
- Shilina, S.G.; Bekker, Z.E.; Goshaev, M.G. Isolation and comparative characterization of vivotoxin from wilt-infected cotton plants and of fusaric acid. Ekologo-Fiziologlcheskle Metody Bor’be Fuzarioznym Viltom Khlop 1973, 2, 219–230. [Google Scholar]
- D’Alton, A.; Etherton, B. Effects of fusaric acid on tomato root hair membrane potentials and ATP levels. Plant Physiol. 1984, 74, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Marré, M.T.; Vergani, P.; Albergoni, F.G. Relationship between fusaric acid uptake and its binding to cell structures by leaves of Egeria densa and its toxic effects on membrane permeability and respiration. Physiol. Mol. Plant Pathol. 1993, 42, 141–157. [Google Scholar] [CrossRef]
- Bouizgarne, B.; El-Maarouf-Bouteau, H.; Frankart, C.; Reboutier, D.; Madiona, K.; Pennarun, A.M. Early physiological responses of Arabidopsis thaliana cells to fusaric acid: Toxic and signalling effects. New Phytol. 2006, 169, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.; Guo, S. Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Physiol. Biochem. 2012, 60, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Cheeke, P.R.; Shull, L.R. Mycotoxins. In Natural Toxicants in Feeds and Poisonous Plants; AVI Publishing Company Inc.: Westport, CT, USA, 1985; pp. 393–477. [Google Scholar]
- Mirocha, C.J.; Christensen, C.M. Oestrogenic mycotoxins synthesized by Fusarium. In Mycotoxins; Purchase, I.F.H., Ed.; Elsevier: Amesterdam, The Nerthlands, 1974; pp. 129–148. [Google Scholar]
- McNutt, S.H.; Purwin, P.; Murray, C. Vulvovaginitis in swine. J. Am. Vet. Med. Assoc. 1928, 73, 484–492. [Google Scholar]
- Graham, R.; Tunnicliff, E.A.; McCullock, E.C. Spoiled feed shown to be the cause of pig trouble. Agric. Exp. Sta. Annu. Rep. 1928, 41, 144–145. [Google Scholar]
- Koen, J.S.; Smith, H.C. An unusual case of genital involvement in swine associated with eating moldy corn. Vet. Med. 1945, 40, 131–133. [Google Scholar]
- Christensen, C.M.; Nelson, G.H.; Mirocha, C.J. Effect on the white rat uterus of a toxic substance isolated from Fusarium. Appl. Microbiol. 1965, 13, 653–659. [Google Scholar] [PubMed]
- Pullar, M.E.; Lerew, W.M. Vulvovaginitis of swine. Aust. Vet. J. 1937, 13, 28–31. [Google Scholar] [CrossRef]
- McErlean, B.A. Vulvovaginitis of swine. Vet. Rec. 1952, 64, 539–540. [Google Scholar]
- Stob, M.; Baldwin, R.S.; Tuite, J.; Andrews, F.N.; Gillette, K.G. Isolation of an anabolic, uterotrophic compound from corn infected with Gibberella zeae. Nature 1962, 196, 13–18. [Google Scholar] [CrossRef]
- Urry, W.H.; Wehrmeister, H.L.; Hodge, E.B.; Hidy, P.H. The structure of zearalenone. Tetrahedron Lett. 1966, 27, 3109–3114. [Google Scholar] [CrossRef]
- Mirocha, C.J.; Christensen, C.M.; Nelson, G.H. Estrogenic metabolite produced by Fusarium graminiearuin in stored corn. Appl. Microbiol. 1967, 15, 497–503. [Google Scholar] [PubMed]
- Codex Committee on Food Additives and Contaminants (CCFAC). Posting Date. Joint FAO/WHO Expert Committee on Food Additives: Position Paper on Zearalenone; Publication CCFAC 00/19; Codex Alimentarius Commission: Rome, Italy, 2000. [Google Scholar]
- Zill, G.; Engelhardt, G.; Wohner, B.; Wallnoefer, P.R. The fate of Fusarium mycotoxin zearalenone in maize cell suspension cultures. Mycotox. Res. 1990, 6, 31–40. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismaiel, A.A.; Papenbrock, J. Mycotoxins: Producing Fungi and Mechanisms of Phytotoxicity. Agriculture 2015, 5, 492-537. https://doi.org/10.3390/agriculture5030492
Ismaiel AA, Papenbrock J. Mycotoxins: Producing Fungi and Mechanisms of Phytotoxicity. Agriculture. 2015; 5(3):492-537. https://doi.org/10.3390/agriculture5030492
Chicago/Turabian StyleIsmaiel, Ahmed A., and Jutta Papenbrock. 2015. "Mycotoxins: Producing Fungi and Mechanisms of Phytotoxicity" Agriculture 5, no. 3: 492-537. https://doi.org/10.3390/agriculture5030492
APA StyleIsmaiel, A. A., & Papenbrock, J. (2015). Mycotoxins: Producing Fungi and Mechanisms of Phytotoxicity. Agriculture, 5(3), 492-537. https://doi.org/10.3390/agriculture5030492