Abstract
Light quality affects the visual and morphological traits of ornamental species, and its effects on plant quality can be quantitatively assessed using image analysis combined with machine-learning classifiers. The objective was to characterize the morphological, reproductive, and color-related traits of Viola cornuta L. cv. Sorbeth® Coconut® Swirl® grown under red, black, green, and blue shade nets in open-field conditions in Montecillo, Mexico, based on the combined use of traditional measurements and image-based analysis. Measurements were taken 69 days after transplanting. Image analysis with a multiclass Naive Bayes classifier (98.9% accuracy) quantified flower area and the three color classes (yellow, cream, and purple). Leaf area and ground cover were measured through color-based segmentation and adaptative thresholding. Open-field plants showed the largest ground cover, with flowers (19.4%), compact canopy (37% smaller than under the black net), and the highest number of flowers (33 flowers/plant). The yellow floral area was also the largest (0.3 cm2/flower). Black, green, and blue nets promoted larger leaf areas (10 to 11 cm2/leaf), while the black net produced the largest flowers (18.6 cm2). Blue and red nets reduced cream (4.3 cm2) and purple (7.3 cm2) areas, respectively. Photoselective nets differentially modulated viola morphology and pigmentation, while open-field conditions yielded compact plants with large flower areas of the highest visual quality.
1. Introduction
Violas and pansies are ornamental [1] and edible species [2,3] that have been cultivated in England since the 19th century [4]. Violas can survive freezing temperatures as low as −10 to −15 °C [5] and grow best at 18 °C [6]. Greenhouse production is often carried out under black shade nets that transmit 20% to 50% of solar radiation [7]. In the United States, their commercial value is up to 32 million USD [8]. Violas can be grown both in pots and in ornamental beds in landscape design, making them a profitable cultivation option.
The use of photoselective shade nets in plant production is based on their ability to regulate both light intensity and spectral quality, thereby modifying plant morphology and physiology, which in turn affects quality, yield, and harvest dates [9]. Light influences growth and development, affecting flowering and ornamental appearance, thus affecting their marketability [10]. These effects occur through photomorphogenic and physiological processes, whereby variations in light intensity, duration, and incidence modifies both the quality and quantity of production [11]. Plants can sense light through photoreceptors such as chlorophyll, phytochromes, cryptochromes, and phototropins, which are sensitive to different bands of the solar spectrum [9]. Photoselective nets contain chromatic additives that reflect and scatter specific wavelengths then modify the light spectrum that reaches the plants, particularly in the UV (200–400 nm), visible (400–700 nm), and far-red (705–740 nm) bands. These spectral shifts influence the accumulation of phytochemicals and aromatic compounds, as well as plant growth, yield, and crop quality [12]. Shade nets also modify microclimatic conditions [9].
It has been shown that these nets affect crop yield [13] and flower induction in Rosa hybrida [14] and plant height, leaf area, and shoot/root ratio in Solanum lycopersicum Mill [15]. In Paeonia lactiflora, netting shortened plant height, leaf number, stem diameter, branch number, node number, and crown width, associated with a decreased photosynthetic capacity; in addition, the nets reduced soluble sugars, proteins, and malondialdehyde contents. These reductions delayed flowering induction, decreased flower weight, and resulted in larger but light-colored flowers. These symptoms are associated with a lesser anthocyanin content, a subclass of flavonoid, due to downregulation of genes encoding phenylalanine, ammonia-lyase, and chalcone synthase [16].
In ornamental plant species, commercial and aesthetic value largely depends on color, form, and size. Coloration in plant structures is mainly determined by pigments such as anthocyanins, carotenoids, betalains, and chlorophyls, which play essential roles in photosynthesis, photoprotection, and defense against environmental stress. The accumulation of these pigments is regulated by both genetic and environmental factors. Pigment content has usually been quantified by means of laboratory-based analytical techniques; however, in recent years, alternative approaches, such as image analysis and colorimetric devices, are increasingly applied [17,18].
Commercial companies often use standard quality norms to classify and assess agricultural products, but evaluation also relies on human experience in specific situations. Nowadays, to evaluate the effect of photoselective shade nets on the appearance and quality of ornamental plants, the changes can be measured through image analysis that can produce data on plant morphology traits and on flower area, size, and color.
In this study, image analysis was performed by means of semantic image segmentation performed by a multiclass Naive Bayes pixel classifier (NBC) implemented in the open-source software PlantCV, programmed to quantify several phenotypic traits from Viola cornuta L. plants cultivated under different photoselective shade nets. The NBC is a probabilistic model that assumes feature independence and is one of the most efficient supervised learning algorithms for pixel-level classification [19]. We postulated that image-derived phenotypic traits could effectively be used to detect and quantify the changes produced in V. cornuta when grown under contrasting light conditions. Another objective was to measure morphological and color traits from the obtained images and compare them with a combination of conventional measurements. Then, the innovation of this study lies in integrating/combining digital phenotyping with image analysis to quantitatively assess ornamental quality, which is a methodological approach not previously applied to V. cornuta L.
2. Materials and Methods
Plants of the hybrid Sorbeth® Coconut® Swirl® (PanAmerican Seed Company®, Chicago, IL, USA) of Viola cornuta L. form many white flowers with a purple picotee pattern (Figure 1A), with short stems and good ground cover [20]. The seeds were sown in Texcoco, Mexico on 9 January 2018 in plastic germination trays filled with a 2:1 peat/perlite mixture. Four weeks later, when half of the seedlings had fully expanded cotyledons, thy were nourished with Ultrasol® (SQM, Tlajomulco de Zúñiga, Jalisco, Mexico) Inicial fertilizer (15N–30P–15K), which was applied at 0.5 g L−1.

Figure 1.
(A) Viola cornuta L. cv. Sorbeth® Coconut® Swirl® plant, and (B) original flower image on the left and the same flower semantically segmented on the right into four classes: (a) yellow, (b) gray, (c) cream, and (d) purple. The gray area was excluded in this study.
At 49 days after sowing, when seedlings had 4 to 7 leaves, they were transplanted into pots of 15 cm in diameter and 20 cm in height, filled with the same substrate as previously described. Pots were arranged in rows, with pots in a row and the space between rows each separated by 10 cm. From transplanting to the onset of flowering, plants were fertilized with Steiner [21] nutrient solution at 25% and an osmotic potential of −0.018 MPa, provided by a dripping irrigation system. Afterwards, the Steiner solution was increased to 50% with −0.036 MPa of osmotic potential. The first flower was removed from each plant, as local Viola producers usually do.
The experiment had 5 treatments (as described below) and 45 pots (replicates) per treatment, with one plant per pot. Treatments were the 4 shade-net treatments each assigned to a tunnel that was 4 m long, 1.5 m wide, and 1.90 m high, plus a control in the open. The five treatments were as follows: open field (no net, T1); red shade net (T2); black shade net (T3); blue shade net (T4); and green shade net (T5). The photosynthetically active radiation (PAR) transmission of all nets was 60%, except for the black net, which had 50%. The differences between the five nets were considered negligible, since variations in temperature and relative humidity were below 0.9 °C and 6.4%, respectively; the irradiance levels remained sufficiently high throughout the spring-summer experimental period, since cloudy days were infrequent. The PAR was measured with a LI-1500 quantum sensor (LI-COR®, Lincoln, NE, USA).
Air temperature (°C) was recorded for each treatment with H08-003-002 thermometers (HOBO®, ONSET Inc., Bourne, MA, USA) located 1 m above the ground inside each tunnel and shielded with white polyurethane radiation screens to prevent direct solar heating. Mean air temperature (Tmean) was then calculated for each treatment (Table 1). The global solar radiation (MJ m−2 d−1) data was registered at a weather station located 50 m away from the experimental site.

Table 1.
Means of air temperature and relative humidity in each environment.
2.1. Morphological Variables and Number of Reproductive Structures
The size of morphological variables and the number of reproductive structures were measured in five plants, randomly sampled, through destructive samples performed in each treatment (n = 5) at 69 days after transplanting (DAT). The variables registered were as follows: leaf area per plant (LA, cm2/plant), measured with a leaf area meter LI-3100 (LI-COR Inc., Lincoln, NE, USA); leaf dry biomass (LDB, g/plant), measured after drying the leaves in a forced-air oven (FELISA®, Zapopan, Jalisco, Mexico) at 75 °C for 72 h until samples reached constant weight, with a analytical balance model 1474 (Sartorius®, Göttingen, Germany); and the specific leaf area (SLA) was calculated by the LA/LDB ratio (cm2/g). In addition, the following variables were recorded per plant: number of leaves (NL), number of open flowers (NOF), number of senescent flowers (NSF), number of floral buds (NFB), and number of capsules (NC): Viola fruits are botanically named capsules, which are defined as dry fruits that open upon maturity. Plant height (cm) was measured from the soil to the stem apex, and the number of stems per plant was manually counted.
2.2. Percentage of Plant Ground Cover
Plant cover (PC), leaf cover (LC), and flower cover (FC) were obtained from digital plant photos taken in a 1 m3 wooden box lined with black paper as a background. For illumination, the box had two General Electric 55 W, 60 Hz fluorescent lamps mounted on top, providing 3690 lux. Digital images from five plants per treatment (n = 5), randomly selected, were captured at a 70 cm distance above the plant canopy with a digital camera programmed with the following settings: fluorescent white balance, 1 s shutter speed, and F/18 aperture. All images were captured in a single session during the morning to minimize the potential variation caused by differences in phenological stage or light exposure. Additionally, the wooden box was positioned adjacent to the experimental area to facilitate rapid image acquisition and to reduce plant handling time. Images were stored in JPEG format at 720 × 480 resolution. All the measured plants exhibited floral buds, open flowers, senescent flowers, and fruits. Software PlantCV version 4.6 in Python version 3.12 was used for image analysis according to the following workflow: RGB images were converted to grayscale using the CIE-a channel to segment leaves through simple thresholding. For flower images, grayscale conversion was performed using the K channel from the CMYK color model, followed by simple thresholding to remove the background. The amount of pixels corresponding to flower and leaf surfaces was counted in each plant, and from these data the percentage of total, leaf, and flower ground cover was calculated.
2.3. Flower and Leaf Area and Color Measurements
At 69 DAT, during flowering and fruiting, five fully expanded leaves and five newly opened flowers in five randomly selected plants per treatment (n = 25) were collected and scanned with a ScanJet® scanner (Hewlett Packard®, Zhongshan, China) at 300 dpi on a white background in JPEG format.
For semantic segmentation, a multiclass Naive Bayes classifier was trained using flower pixel samples. Input variables were the three channels of the RGB color model whereas the output variable was the target color class in three categories: purple, cream, and yellow (Figure 1B). For each color category, 13 samples of 20 × 20 pixels were extracted from independent flower images, encompassing distinct hues within the same category. After removing duplicated pixel values and randomly standardizing the sample size to match the category with the lowest number of samples, a total of 750 pixels per category were retained and randomly partitioned into two subsets: for training (80%) and for testing (20%). This process was repeated ten times using Monte Carlo cross-validation to obtain the confusion matrices. From each matrix, Precision, Recall, F1, and Specificity metrics were calculated, and the averages were used to evaluate the classifier performance. The flower segmentation workflow was as follows: (1) White balance correction using a ColorChecker Classic (Calibrite®, Wilmington, DE, USA). Parameters from the scanned color card were applied to correct all flower and leaf images; (2) RGB image conversion to grayscale by using the hue channel of the HSV color model and a simple thresholding for image binarization; (3) application of morphological and noise-removal filters (erosion-dilation, removal of small objects), and creation of a final mask; (4) background removal using the previous mask; (5) semantic segmentation of the floral surface with the Naive Bayes classifier, grouping pixels into the three color classes. Pixel counts were attained for each color class and for the total floral area, and thereafter the flower surface area (cm2/flower) was calculated. Calibration was performed using the previously described color card with a known chip length (12 mm × 12 mm). The corresponding chip length and area in pixels, obtained through the plantcv.analize.size function, were used to calculate a proportional conversion factor that transformed pixel-based measurements into metric units. Mean values of CIE-Lab channels were also computed for each class.
For leaves, a similar workflow was used: white balance correction of RGB image, followed by grayscale conversion using the CIE-L channel, and then a simple thresholding for image binarization. After noise-removal in each image, the final mask was used to remove background in the corrected RGB image. Then, leaf area and average CIE-Lab color values were calculated.
2.4. Statistical Analysis
Statistical analyses were performed using open-source Python™ modules (version 3.12.1). The normality test was assessed using the Shapiro–Wilk test (SciPy version 1.7.3) and homogeneity of variances with the Levene test. When assumptions of normality and homoscedasticity were met, analysis of variance (ANOVA) was performed using the statsmodels module version 0.13.1, followed by the Tukey mean test (α = 0.05) with the statsmodels.stats.multicomp submodule. When the above assumptions were not satisfied, a non-parametric analysis was applied using the Kruskal–Wallis test (SciPy) and the Conover median test with the Holm correction (scikit-posthocs version 0.9.0). Data for all response variables were independent, as all samples were randomly selected. Variables such as specific leaf area, flower and leaf surface area, and the color channels of both leaves and flowers did not meet the assumptions of normality and homogeneity of variances; therefore, they were analyzed with a non-parametric method.
3. Results
Significant differences between treatments were observed for physiological and morphological variables (p ≤ 0.05), except for the number of capsules and plant height. Plants grown under the red net (Figure 2B) produced the lowest number of leaves (NL), 47% to 58% less leaves compared to the other treatments. In contrast, plants grown under the blue and green nets showed larger leaf area (LA), by 65% and 34%, respectively, compared with those under the red net or in the open field (Figure 2D). The median specific leaf area (SLA) under the green net was up to 35% higher than under the red net or the open field conditions (Figure 2C).
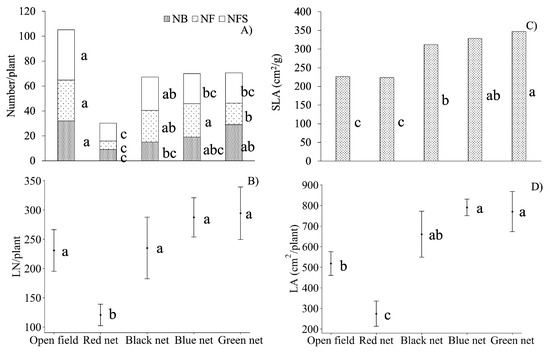
Figure 2.
(A) Number of floral buds (NFB), number of flowers (NF), and number of senescent flowers (NSF); (B) number of leaves (LN); (C) specific leaf area (SLA); and (D) leaf area (LA) per plant of Viola cornuta L. at 69 days after transplanting (DAT). n = 5. Means with different letters within each graph and for each floral structure indicate significant differences according to the Tukey mean test (α = 0.05), except for SLA, which was analyzed with the Conover median test (α = 0.05).
Open field plants showed higher values in several traits: 71% more floral buds per plant than those under the red net, and 53% more than under the black one (Figure 2A). They also produced 47 to 79% more flowers per plant than plants grown under the green or red nets, although they also had 40 to 64% more senescent flowers than those under the blue, green, or red nets.
The blue net caused a significantly higher number of stems (NS) compared with those under the other shade nets (Table 2). Both blue net and open field plants had nearly twice the number of stems than those grown under the red net.
Table 2.
Number of stems (NS) and plant height (PH) of Viola cornuta L. at 69 DAT (n = 5).
One plant grown under the black net exhibited the largest canopy cover and thus it was used as the reference value (100%) since the remaining plants were smaller. The canopy cover values were proportionally adjusted in all the treatments (Figure 3A).
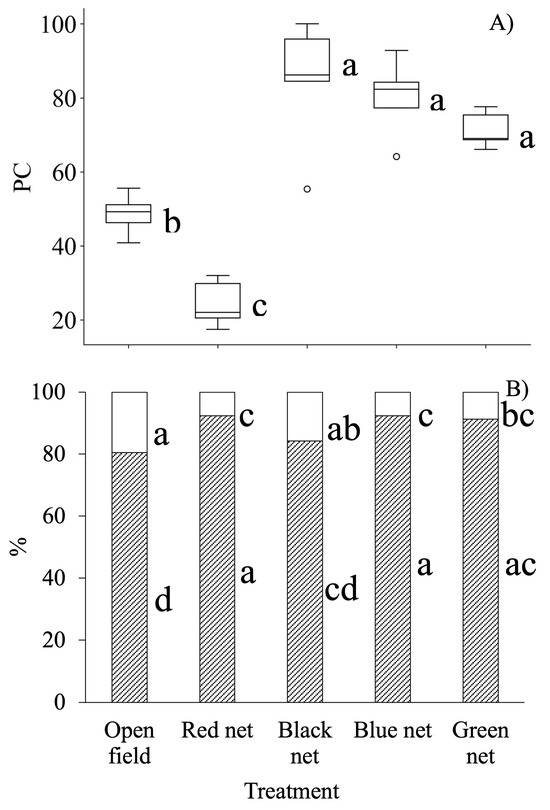
Figure 3.
(A). Mean percentage of plant cover (PC) in Viola cornuta L., as compared to the largest canopy cover under the black shade net at 85 DAT. (B). Mean percentage of flower cover (FC,  ), and leaf cover (LC,
), and leaf cover (LC,  ). n = 5. Values with different letters within each cover type indicate significant differences according to Tukey mean test (α = 0.05). DAT = days after transplanting.
). n = 5. Values with different letters within each cover type indicate significant differences according to Tukey mean test (α = 0.05). DAT = days after transplanting.
), and leaf cover (LC, ). n = 5. Values with different letters within each cover type indicate significant differences according to Tukey mean test (α = 0.05). DAT = days after transplanting.
Mean PC under open field and red net treatments (Figure 3A) were 44% and 73% lower, respectively, compared with plants under the black net (α = 0.05). The lowest mean LC occurred in the open field, up to 13% less than under the blue, red, or green nets, resulting in up to 2.5 times greater FC (Tukey, α = 0.05) (Figure 3B). Plants grown under the black net exhibited up to twice the FC compared with those under the blue or green nets (Figure 4).
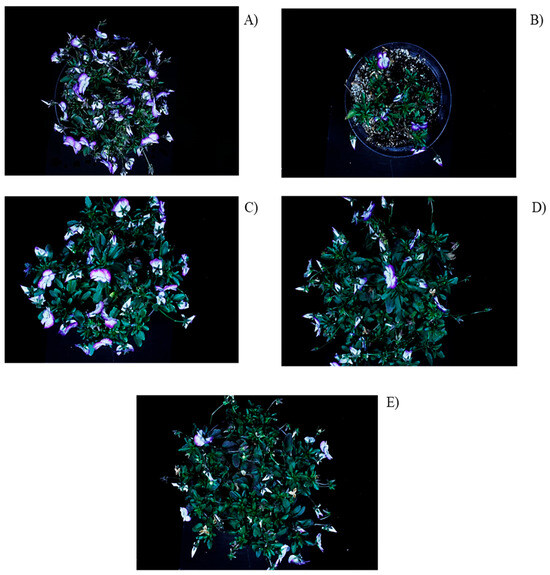
Figure 4.
Viola cornuta L. plants at 85 DAT grown under (A) open field, and shade nets: (B) red, (C) black, (D) blue, and (E) green.
The Naive Bayes classifier achieved an overall accuracy of 98.9% when classifying Viola flowers into three color categories: purple, cream, and yellow (Table 3). If noise-removal functions are not implemented and illumination conditions differ, the classifier is likely to exhibit reduced accuracy in its predictions. Consequently, future research should aim to develop more robust models capable of considering variations in light intensity and image noise. In this context, deep learning represents an appropriate and potentially better alternative for improving classification performance under variable image conditions.
Table 3.
Mean values of metrics from a Naive Bayes classifier of pixels in the test.
Yellow area pixels were classified with 100% precision and specificity. Cream area pixels achieved 100% recall and 99% specificity, while purple area pixels were classified with 98% recall and 99% specificity.
In the scanned images, the yellow-class floral area (Figure 5) was larger under open field conditions than under red, blue, or green nets, while under the black net it only exceeded blue and green nets (α = 0.05). For the cream floral class, the smallest area was recorded under the blue net. In contrast, the area of this class was greater under the red net compared with the open field and the blue and green nets (α = 0.05), whereas the purple class area was the lowest under the red net. Total flower size was greater under the black net compared with the open field and red net.
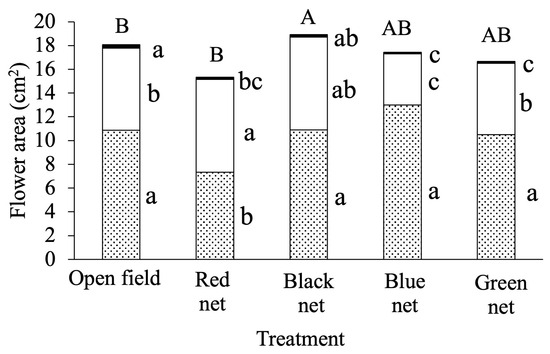
Figure 5.
Total flower surface of Viola cornuta L. and its classification into yellow ( ), cream (), and purple (
), cream (), and purple ( ) areas at 69 days after transplanting (DAT). Different lowercase letters within each surface type (color class) and different uppercase letters for the total flower surface indicate differences among medians (n = 25) according to the Conover median test with Holm adjustment (α = 0.05).
) areas at 69 days after transplanting (DAT). Different lowercase letters within each surface type (color class) and different uppercase letters for the total flower surface indicate differences among medians (n = 25) according to the Conover median test with Holm adjustment (α = 0.05).
), cream (), and purple () areas at 69 days after transplanting (DAT). Different lowercase letters within each surface type (color class) and different uppercase letters for the total flower surface indicate differences among medians (n = 25) according to the Conover median test with Holm adjustment (α = 0.05).
Leaf area was smaller in plants grown in the open field and under the red net. Regarding leaf color (Table 4), the highest brightness was observed in plants grown under the red net and in the open field (α = 0.05). Moreover, leaves under the red net showed the highest level of yellow (CIE-b) and a more intense green color compared to those grown under the black and green nets.
Table 4.
Brightness (CIE-L), green level (negative values, CIE-a), and yellow level (positive values, CIE-b) of leaves of Viola cornuta L.
The purple area of flowers (Table 5) grown under the red net exhibited higher brightness, though with lower red and blue levels compared with the other treatments (α = 0.05). Under the same net, the cream area also showed higher brightness than flowers grown in the open field or under the blue and green nets. For the yellow area, a consistent pattern was observed across the three CIE-Lab color channels: brightness, red, and blue levels were all lower under the blue and green nets.
Table 5.
Brightness (CIE-L), green level (negative values, CIE-a), and yellow level (positive values, CIE-b) of flowers of Viola cornuta L. by color class (purple, cream, yellow). n = 25.
4. Discussion
Viola cornuta L. plants cultivated under open field conditions (8.2 MJ m−2 d−1 in the photosynthetically active radiation band, 19.9 °C, and 42% of relative humidity) exhibited a more compact growth habit compared with those grown under black, blue, and green shade nets. Similar responses were reported in Codiaeum variegatum var. “Bush on Fire” [22] grown in open field conditions where they developed a more compact canopy and thicker leaves. The photosynthetic irradiance in the open field (37.8 mol m−2 d−1) exceeded the threshold of 26 mol m−2 d−1, above which flower quality in Viola declines if plants are not grown under cooling conditions [23]. In addition, plants were exposed to temperature above the optimum, which is ≤18 °C [6]. Despite these two drawbacks, the plants in the open field produced a greater number of flowers than those grown under red or blue nets and thus had a higher proportion of flower coverage as compared to plants grown under red, blue, or green nets.
This response contrasts with the findings reported for Begonias, where more flowers were produced under the black shade net than in the control [24]. In open field conditions, the high red/far-red ratio likely contributed to create a more compact plant architecture, whereas the greater daily light integral accelerated flowering and improved the overall floral quality of Viola, as also happened in Petunia [25]. The daily light integral was a more reliable index for flowering performance and quality that the far-red light levels, emphasizing the importance of total light energy rather than the spectral composition by itself [25]. Moreover, although responses may differ among species and cultivars, higher temperatures often stimulate floral induction in many spring-flowering plants [26]. This may partly explain the greater blooming expression achieved in the open field, where the higher temperatures might have accelerated developmental processes.
Plants grown under shade nets experienced 40 to 50% lower irradiance in the photosynthetic band, lower temperatures by 0.85 °C, and had 13% higher relative humidity (RH). Similarly to our experiment, in Viola wittrockiana Gams, the greatest temperature and lowest relative humidity from the environment under shade nets were registered under red nets [27].
Morphological changes associated with shade nets involved increases in stem elongation (also observed in Capsicum annuum, Glycine max L. Merr., and Ranunculus), higher leaf area (as in Ocimum basilicum and C. annuum), and thinner leaves (as in O. basilicum) [28,29,30,31]. However, in our study no differences in Viola plant height were observed such as those reported for Begonias under shade nets [24]. None of the treatments reached the recommended height of 15 to 20 cm for V. cornuta reported by [20]. Stem elongation is regulated by the phytochrome, which is activated by red light to produce its active form (Pfr) and inactivated by far-red light, resulting in longer stems. Temperature may also influence this process through thermal reversion, in which Pfr is converted back to the inactive form (Pr), and both processes occur simultaneously [32]. These interactions add complexity to the phenotypic outcome of stem length in different species.
Plants grown under green or blue nets developed greater leaf area than those in the open field and under the red net, but their leaves were thinner. Pansy “Clear Yellow” plants were observed to have greater leaf areas under blue nets than under black nets [27]; however, this difference was not observed in V. cornuta. For the blue net, this response may be attributed to a lower red/far-red light ratio [33] which triggers the shade-avoidance response, whereas the green net has been associated with increases in the hormones GA3 and IAA, both of which promote growth. It is known that green light penetrates deeper into the canopy, thus enhancing plant photosynthesis [34].
Regarding the area coverage as a quality index, plants under the black net presented a larger cover area than in the open field plants. However, the percentages of leaf and flower coverage were similar between these two environments. Plant cover area under the black net was similar to the plants grown under the green and blue nets, although in the latter two treatments, the proportion of floral coverage was smaller than under the black net.
Compared with the other nets, plants under the red net exhibited thicker but fewer leaves; thus, a reduced leaf area, and also fewer flowers per plant. These results contrast with reports of higher leaf area under red nets in Calendula officinalis L., Viola tricolor [35], Cucumis sativus L. [13], and Solanum lycopersicum [15]. Evidently, these responses to shade are diverse among genotypes [36].
Regarding color quality, leaf brightness (CIE-L) was higher in the open field and in the red net plants than under the black, blue, or green nets. Brightness is linked to leaf reflectance, so it depends on structural components, morphological adaptations, pigment content, and cuticular wax deposition. Wax accumulation is affected by temperature, light intensity, and plant water status [37]. For flower color, the red net produced higher brightnesses in all three color classes, whereas blue and green nets induced smaller values in all the CIE-Lab channels compared to other treatments. In Begonia plants, a lower luminosity value occurred in the open field compared with plants grown under a black net, because tepals in the open field had a more intense color [24].
Shade nets also influenced flower size, since larger flowers were observed under the black net as compared to the open field or the red net. Flower diameters in Viola are inversely related to temperature within the 9 to 21 °C range [38], a relationship that can be described by a linear model for Viola × wittrockiana Gams. This response has been attributed to the lesser time available for floral development as temperature increases [39]. Consistently, in our study, the lowest temperatures occurred under the black net, where plants produced the largest flowers. Conversely, plants grown under the blue net showed the smallest flower areas in the cream color class, a class that lacks anthocyanins [40], whereas the yellow area was larger in the open field than under the blue or green nets.
Although the proportion of yellow surface in the studied Viola cultivar was smaller than the other colors (1.0 to 2.8% of the flower area in open field conditions and 0.33 to 0.85% under shade nets), yellow provides a contrasting hue of high ornamental value. Furthermore, yellow intensity (positive CIE-b values) in the open field environment exhibited its highest levels. In Viola × wittrockiana the yellow and orange petal colors are associated with carotenoid pigments such as lycopene and violaxanthin [41], whose accumulation is stimulated by elevated UV-B radiation [2]. V. tricolor plants exposed to open field conditions accumulate more carotenoids than those under green or blue shade nets [35]. Therefore, open-grown Viola cornuta not only increased carotenoid concentrations but also increased the floral petal area containing these pigments.
The smallest areas of dark colors, such as purple and blue, which have low CIE-b values, were registered in Viola plants grown under the red net. This result contrasts with reports on lycianthus, where the flowers in both the control treatment and under the red net exhibited a bluer hue [10]. In Viola, blue and violet hues are mainly attributed to anthocyanins such as nasunin, rutin (flavonol glycoside) [40], luteolin-6-C-rhamnosyl-8-C-glucoside (co-pigment), violantine, violanin [42], cianidin, and pelargonidin derivatives [3]. These pigments are water soluble, unstable, and prone to degradation [42]. An increase in malvidin content over time in Viola cornuta cv. Yesterday, Today and Tomorrow (YTT) suggests that anthocyianin synthesis can be stimulated by pollination and light [43], including UV radiation and red light [44]. However, higher pH, temperature and prolongated light exposure accelerate anthocyanin degradation [45]. These findings highlight the need for further research to tackle the complex relationships among genotype, light quantity and quality, and temperature, for determining morphology, physiology and coloration of ornamental species.
Image analysis applied to detect variations in size, distribution of plant structures, and coloration of Viola plants represents a valuable tool for the genetic improvement of this species’ [4] ornamental quality, as well as for assessing the influence of environmental conditions on their development. This approach not only enables the identification of optimal conditions for production and cultivation in terms of visual quality but also in relation to the content of bioactive compounds with potential benefits for human health [3] such as carotenoids, anthocyanins, and chlorophyl. Image analysis enables detection of subtle variations in color that might be associated with these pigments. Machine learning classifiers have been used in several studies for evaluating ornamental species based on these morphological and color traits. For example, different ornamental species were identified according to these features with 93.9% accuracy [46]. Similarly, image analysis combined with neural networks achieved 70 to 88% accuracy in assessing ornamental quality scores for Viola, Dianthus and Cyclamen, supporting applications in crop management and market grading [47]. For morphological measurements, an XGBoost-based classifier obtained over 92% accuracy in identifying desirable quality traits in ornamental pepper plants [48]. In addition, indices derived from RGB images were used to build predictive models of anthocyanin content in Rosa chinensis petals, where both back-propagation neural network and random forest algorithms achieved a coefficient of determination above 0.75 [49].
Although the use of shade nets is a convenient strategy to regulate the microclimatic environment for ornamental plant production, in Viola cornuta, a species sensitive to water stress, these conditions did not result in improved plant quality. Plants grown in the open field were more compact and exhibited a larger flowered area and a greater number of flowers. In the open the leaves were smaller but brighter, and the flowers were smaller than those produced by plants grown under the black net, but in the open the flowers displayed a higher proportion of yellow coloration, a very attractive color in ornamental plants. Despite being a minor color component, this yellow hue contributes to enhanced ornamental appeal.
5. Conclusions
The Viola cornuta L. plant color and morphology, as well as its flower structures, were affected by the type of photoselective shade net under which they were grown, thus pointing out their potential to modulate ornamental quality in accordance with consumer preferences. Image analysis combined with semantic segmentation using a multiclass Naive Bayes pixel classifier proved effective in detecting and quantifying subtle morphological and color variations induced by differences in light quality and intensity, achieving an overall accuracy of 98.9%. This approach offers practical potential for rapid, low-cost phenotyping of ornamental plants under different growing conditions. Unlike deep learning architectures, which require extensive datasets and computational resources, the Naive Bayes classifier provides an accessible and efficient alternative for routine color and morphology assessment in both research and commercial production.
Plants grown in the open field conditions were more compact and exhibited greater floral coverage, representing the best visual quality among treatments. Thus, cultivation under shade nets does not necessarily enhance commercial appearance, although these structures may still be beneficial for reducing water stress, to which Viola cornuta is particularly sensitive.
Ornamental species are characterized by subtle variations in appearance that, although seemingly minor, can significantly influence consumer perception and market value. Traits such as variegation, leaf reflectance, and color of leaves and flowers determine aesthetic quality, yet their evaluation commonly requires expert panels for consumer-oriented assessment. Further research should integrate expert-based evaluations with image-derived traits to strengthen quality assessment frameworks. By developing deep learning classifiers capable of analyzing whole images through instance segmentation could further advance automatic recognition of ornamental quality. Such models may effectively overcome image noise and variations in illumination, thereby enhancing the robustness and accuracy of the classification process.
Finally, it is important to recognize that this study might be limited by the moderate sample size and the number of pixels used for classifier training, as well as by the specific environmental conditions in which the experiment was conducted. Broader validation across cultivars and environmental settings would help to confirm the general applicability of the proposed approach.
Author Contributions
Conceptualization: J.A.C.-S. and F.A.H.-T.; Methodology: J.A.C.-S. and F.A.H.-T.; Software: J.A.C.-S.; Validation: J.M.G.-C. and V.A.G.-H.; Formal analysis: J.A.C.-S. and V.A.G.-H.; Investigation: F.A.H.-T.; Data curation: F.A.H.-T.; Writing—original draft preparation: J.A.C.-S.; Writing–review and editing: J.M.G.-C. and V.A.G.-H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
Wish to express our appreciation to the former CONACYT (SECITHI) in Mexico for the scholarship granted to the first author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sharma, R.; Dilta, B.S.; Baweja, H.S.; Kumari, P.; Thaneshwari, T.; Sahare, H.A. Effect of seed priming and planting date on growth, flowering and seed production of pansy (Viola × wittrockiana Gams.). Ann. Biol. 2018, 34, 176–180. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/20183334327 (accessed on 8 October 2025).
- Park, J.W.; Kim, W.S. Effect of UV-B radiation on antioxidation in petals of edible flower pansy. Flower Res. J. 2017, 25, 24–28. [Google Scholar] [CrossRef]
- González-Barrio, R.; Periago, M.J.; Luna-Recio, C.; Garcia-Alonso, F.J.; Navarro-González, I. Chemical composition of the edible flowers, pansy (Viola wittrockiana) and snapdragon (Antirrhinum majus) as new sources of bioactive compounds. Food Chem. 2018, 252, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, Y.; Iwata, H.; Hase, N.; Matsuura, S. Genetic combining ability of petal shape in garden pansy (Viola × wittrockiana Gams) based on image analysis. Euphytica 2006, 151, 311–319. [Google Scholar] [CrossRef]
- Buczacki, S. Plantas de Invierno; Octopus Publishing Group Limited: Madrid, Spain, 1997; pp. 84–85. [Google Scholar]
- Warner, R.M.; Erwin, J.E. Prolonged high-temperature exposure differentially reduces growth and flowering of 12 Viola × wittrockiana Gams. Sci. Hortic. 2006, 108, 295–302. [Google Scholar] [CrossRef]
- Collado, C.E.; Hernández, R. Effects of light intensity, spectral composition, and paclobutrazol on the morphology, physiology, and growth of petunia, geranium, pansy, and dianthus ornamental transplants. J. Plant Growth Regul. 2022, 41, 461–478. [Google Scholar] [CrossRef]
- USDA (United States Department of Agriculture). Floriculture Crops 2019. Summary; USDA: Washington, DC, USA, 2019. Available online: https://www.nass.usda.gov/Publications/Todays_Reports/reports/floran20.pdf (accessed on 8 October 2025).
- Stamps, R.H. Use of colored shade netting in horticulture. HortScience 2009, 44, 239–241. [Google Scholar] [CrossRef]
- Almeida, J.M.; Calaboni, C.; Rodrigues, P.H.V. Pigments in flower stems of lisianthus under different photoselective shade nets. Ornam. Hortic. 2021, 27, 535–543. [Google Scholar] [CrossRef]
- Mills-Ibibofori, T.; Dunn, B.L.; Maness, N.; Payton, M. Effect of LED lighting and gibberellic acid supplementation on potted ornamentals. Horticulturae 2019, 5, 51. [Google Scholar] [CrossRef]
- Sivakumar, D.; Jifon, J.; Soundy, P. Spectral quality of photo-selective shade nettings improves antioxidants and overall quality in selected fresh produce after postharvest storage. Food Rev. Int. 2018, 34, 290–307. [Google Scholar] [CrossRef]
- Ayala-Tafoya, F.; Yáñez-Juárez, M.G.; López-Orona, C.A.; Medina-López, R.; Velázquez-Alcaraz, T.J.; Díaz-Valdés, T. Sunlight transmitted by colored shade nets on photosynthesis and yield of cucumber. Ciência Rural 2018, 48, e20170829. [Google Scholar] [CrossRef]
- Hou, W.; Luo, Y.; Wang, X.; Chen, Q.; Sun, B.; Wang, Y.; Liu, Z.; Tang, H.; Zhang, Y. Effects of shading on plant growth, flower quality and photosynthetic capacity of Rosa hybrida. AIP Conf. Proc. 2018, 1956, 020005. [Google Scholar] [CrossRef]
- Thwe, A.A.; Kasemsap, P.; Vercambre, G.; Gay, F.; Phattaralerphong, J.; Gautier, H. Impact of red and blue nets on physiological and morphological traits, fruit yield and quality of tomato (Solanum lycopersicum Mill.). Sci. Hortic. 2020, 264, 109185. [Google Scholar] [CrossRef]
- Zhao, D.; Hao, Z.; Tao, J. Effects of shade on plant growth and flower quality in the herbaceous peony (Paeonia lactiflora Pall.). Plant Physiol. Biochem. 2012, 61, 187–196. [Google Scholar] [CrossRef]
- Zlatev, Z.; Stoykova, V.; Shivacheva, G.; Vasilev, M. Design and implementation of a measuring device to determine the content of pigments in plant leaves. Appl. Syst. Innov. 2023, 6, 64. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Y.; Long, T.; Wang, S.; Yang, J. Regulation mechanism of plant pigments biosynthesis: Anthocyanins, carotenoids, and betalains. Metabolites 2022, 12, 871. [Google Scholar] [CrossRef]
- Peretz, O.; Koren, M.; Koren, O. Naive Bayes classifier—An ensemble procedure for recall and precision enrichment. Eng. Appl. Artif. Intell. 2024, 136, 108972. [Google Scholar] [CrossRef]
- PanAmerican Seed. Guía de Información de Productos 2005; PanAmerican Seed Company: West Chicago, IL, USA, 2005; 96p. [Google Scholar]
- Steiner, A.A. The universal nutrient solution. In Proceedings of the 6th International Congress on Soilless Culture, Lunteren, The Netherlands, 29 April–5 May 1984; International Society for Soilless Culture: Wageningen, The Netherlands, 1984; pp. 633–649. [Google Scholar]
- Krishnamoorthy, N.; Srikrishnah, S.; Sutharsan, S. Influence of different shade levels on the growth and quality of Codiaeum variegatum var. ‘Bush on fire’ in the Batticaloa District. Int. J. Res. Publ. 2020, 6, 1–22. Available online: https://www.ijrp.org/paper-detail/217 (accessed on 8 October 2025).
- Torres, A.P.; Lopez, R.G. Measuring Daily Light Integral (DLI). Purdue Extension Publication HO-238-W. 2009. Available online: https://www.extension.purdue.edu/extmedia/HO/HO-238-B-W.pdf (accessed on 8 October 2025).
- Oliveira, J.E.D.; Sabino, J.H.F.; Sillmann, T.A.; Mattiuz, C.F.M. Cultivation under photoselective shade nets alters the morphology and physiology of Begonia Megawatt varieties. Ciência Agrotecnologia 2024, 48, e015924. [Google Scholar] [CrossRef]
- Xia, J.; Mattson, N. Daily light integral and far-red radiation influence morphology and quality of liners and subsequent flowering and development of petunia in controlled greenhouses. Horticulturae 2024, 10, 1106. [Google Scholar] [CrossRef]
- Tun, W.; Yoon, J.; Jeon, J.S.; An, G. Influence of climate change on flowering time. J. Plant Biol. 2021, 64, 193–203. [Google Scholar] [CrossRef]
- Austerman, P.; Dunn, B.L.; Singh, H.; Fontanier, C.; Stanphill, S. Height control of greenhouse-grown pansy using colored shade nets. HortTechnology 2023, 33, 36–43. [Google Scholar] [CrossRef]
- Stagnari, F.; Mattia, C.; Galieni, A.; Santarelli, V.; D’Egidio, S.; Pagnani, G.; Pisante, M. Light quantity and quality supplies sharply affect growth, morphological, physiological and quality traits of basil. Ind. Crops Prod. 2018, 122, 277–289. [Google Scholar] [CrossRef]
- Kabir, M.Y.; Díaz-Pérez, J.C.; Nambeesan, S.U. Effect of shade levels on plant growth, physiology, and fruit yield in bell pepper (Capsicum annuum L.). Acta Hortic. 2020, 1268, 311–318. [Google Scholar] [CrossRef]
- Khalid, M.H.B.; Raza, M.A.; Yu, H.Q.; Sun, F.A.; Zhang, Y.Y.; Lu, F.Z.; Si, L.; Iqbal, N.; Khan, I.; Fu, F.L.; et al. Effect of shade treatments on morphology, photosynthetic and chlorophyll fluorescence characteristics of soybeans (Glycine max L. Merr.). Appl. Ecol. Environ. Res. 2019, 17, 2551–2569. [Google Scholar] [CrossRef]
- Mayoli, R.N.; Isutsa, D.K.; Tunya, G.O. Growth of ranunculus cutflower under tropical high altitude conditions. 1: Effects of GA3 and shade. Afr. J. Hortic. Sci. 2009, 2, 13–28. Available online: https://www.academia.edu/48584182/Growth_of_ranunculus_cutflower_under_tropical_high_altitude_conditions_1_Effects_of_GA3_and_shade (accessed on 8 October 2025).
- Jung, J.; Domijan, M.; Klose, C.; Biswas, S.; Ezer, D.; Gao, M.; Khattak, A.; Box, M.; Charoensawan, V.; Cortijo, S.; et al. Phytochromes function as thermosensors in Arabidopsis. Science 2016, 354, 886–889. [Google Scholar] [CrossRef]
- Arthurs, S.P.; Stamps, R.H.; Giglia, F.F. Environmental modification inside photoselective shadehouses. HortScience 2013, 48, 975–979. [Google Scholar] [CrossRef]
- Roh, Y.S.; Yoo, Y.K. Light quality of light emitting diodes affects growth, chlorophyll fluorescence and phytohormones of Tulip ‘Lasergame’. Hortic. Environ. Biotechnol. 2023, 64, 245–255. [Google Scholar] [CrossRef]
- Zare, S.K.A.; Sedaghathoor, S.; Dahkaei, M.P.; Hashemabadi, D. The effect of light variations by photoselective shade nets on pigments, antioxidant capacity, and growth of two ornamental plant species: Marigold (Calendula officinalis L.) and violet (Viola tricolor). Cogent Food Agric. 2019, 5, 1650415. [Google Scholar] [CrossRef]
- Singh, H.; Dunn, B.L.; Fontanier, C.; Singh, H. Colored shade nets affect growth but not flowering of four greenhouse-grown potted ornamental species. HortScience 2023, 58, 1075–1076. [Google Scholar] [CrossRef]
- Naveena, N.; Thamaraiselvi, S.P.; Rajadurai, K.R.; Sivakumar, R. Effect of coloured shade nets on physiology and quality of cut foliage plants. J. Pharmacogn. Phytochem. 2019, 8, 1141–1144. Available online: https://www.phytojournal.com/archives/2019/vol8issue4/PartS/8-3-641-857.pdf (accessed on 8 October 2025).
- Antoniazzi, L. El cultivo de la viola. Rev. Hortic. 2007, 199, 44–47. Available online: http://www.horticom.com/revistasonline/horticultura/rh199/44_47.pdf (accessed on 8 October 2025).
- Pearson, S.; Parker, A.; Adams, S.R.; Hadley, P.; May, D.R. The effects of temperature on the flower size of pansy (Viola × wittrockiana Gams.). J. Hortic. Sci. 1995, 70, 183–190. [Google Scholar] [CrossRef]
- Hase, N.; Matsuura, S.; Yamaguchi, M. HPLC evaluation of anthocyanins and flavonols in relation to the flower color of pansy (Viola × wittrockiana Gams). Hortic. Res. 2005, 4, 125–129. [Google Scholar] [CrossRef]
- Gamsjaeger, S.; Baranska, M.; Schulz, H.; Heiselmayer, P.; Musso, M. Discrimination of carotenoid and flavonoid content in petals of pansy cultivars (Viola × wittrockiana) by FT-Raman spectroscopy. J. Raman Spectrosc. 2011, 42, 1240–1247. [Google Scholar] [CrossRef]
- Skowyra, M.; Calvo, M.; Gallego, M.; Azman, N.; Almajano, M. Characterization of phytochemicals in petals of different colours from Viola × wittrockiana Gams. and their correlation with antioxidant activity. J. Agric. Sci. 2014, 6, 93–105. [Google Scholar] [CrossRef]
- Farzad, M.; Griesbach, R.; Weiss, M.R. Floral color change in Viola cornuta L. (Violaceae): A model system to study regulation of anthocyanin production. Plant Sci. 2002, 162, 225–231. [Google Scholar] [CrossRef]
- Bastías, R.M.; Corelli-Grappadelli, L. Light quality management in fruit orchards: Physiological and technological aspects. Chil. J. Agric. Res. 2012, 72, 574–581. [Google Scholar] [CrossRef]
- Jenshi-Roobha, J.; Saravanakumar, M.; Aravindhan, K.M.; Suganya-Devi, P. The effect of light, temperature, and pH on stability of anthocyanin pigments in Musa acuminata bract. Res. Plant Biol. 2011, 1, 5–12. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/20123239922 (accessed on 8 October 2025).
- Ojha, A.; Kumar, V. Image classification of ornamental plants leaf using machine learning algorithms. In Proceedings of the 2022 4th International Conference on Inventive Research in Computing Applications (ICIRCA), Coimbatore, India, 21–23 September 2022; IEEE: Piscataway, NJ, USA, 2022; pp. 834–840. Available online: https://ieeexplore.ieee.org/abstract/document/9985604 (accessed on 8 October 2025).
- Parsons, N.R.; Edmondson, R.N.; Song, Y. Image analysis and statistical modelling for measurement and quality assessment of ornamental horticulture crops in glasshouses. Biosyst. Eng. 2009, 104, 161–168. [Google Scholar] [CrossRef]
- do Nascimento, A.M.M.; Ruiz-Gonzalez, R.; Martínez-Martínez, V.; Medeiros, A.M.; Santos, F.S.; Rêgo, E.R.; Pimenta, S.; Sudré, C.P.; Bento, C.d.S.; Cambra, C.; et al. Ornamental potential classification and prediction for pepper plants (Capsicum spp.): A comparison using morphological measurements and RGB images as data source. Appl. Sci. 2025, 15, 7801. [Google Scholar] [CrossRef]
- Liu, X.Y.; Yu, J.R.; Deng, H.N. Non-destructive prediction of anthocyanin content of Rosa chinensis petals using digital images and machine learning algorithms. Horticulturae 2024, 10, 503. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).