

Characterisation of the Pathogenicity of Beauveria sp. and Metarhizium sp. Fungi Against the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae)


 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. Genomic DNA Extraction and Whole Genome Sequencing
2.3. Bioinformatics Analysis
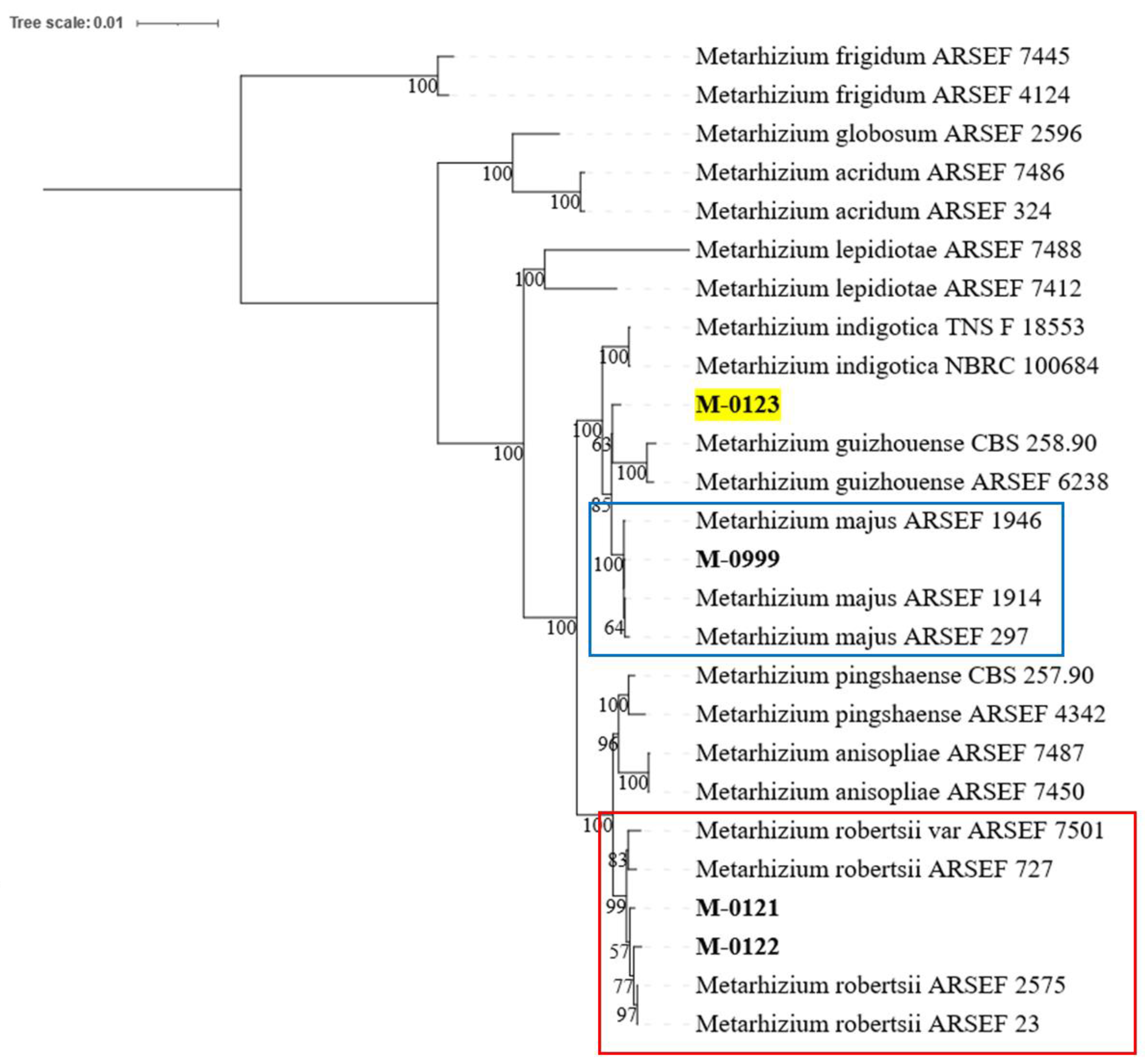
2.4. Species Identification and Phylogeny
2.5. Insect Samples and Insect Bioassay
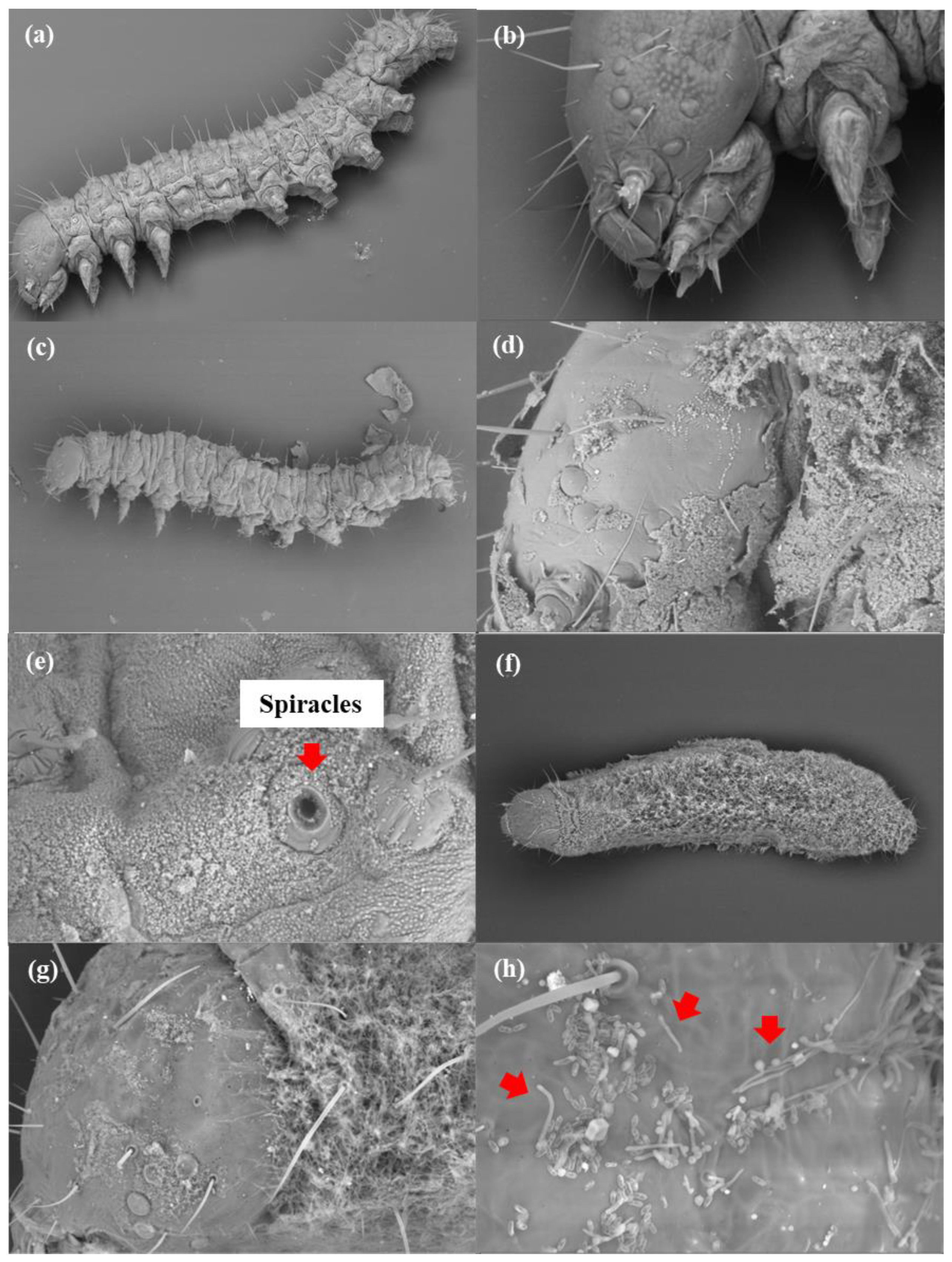
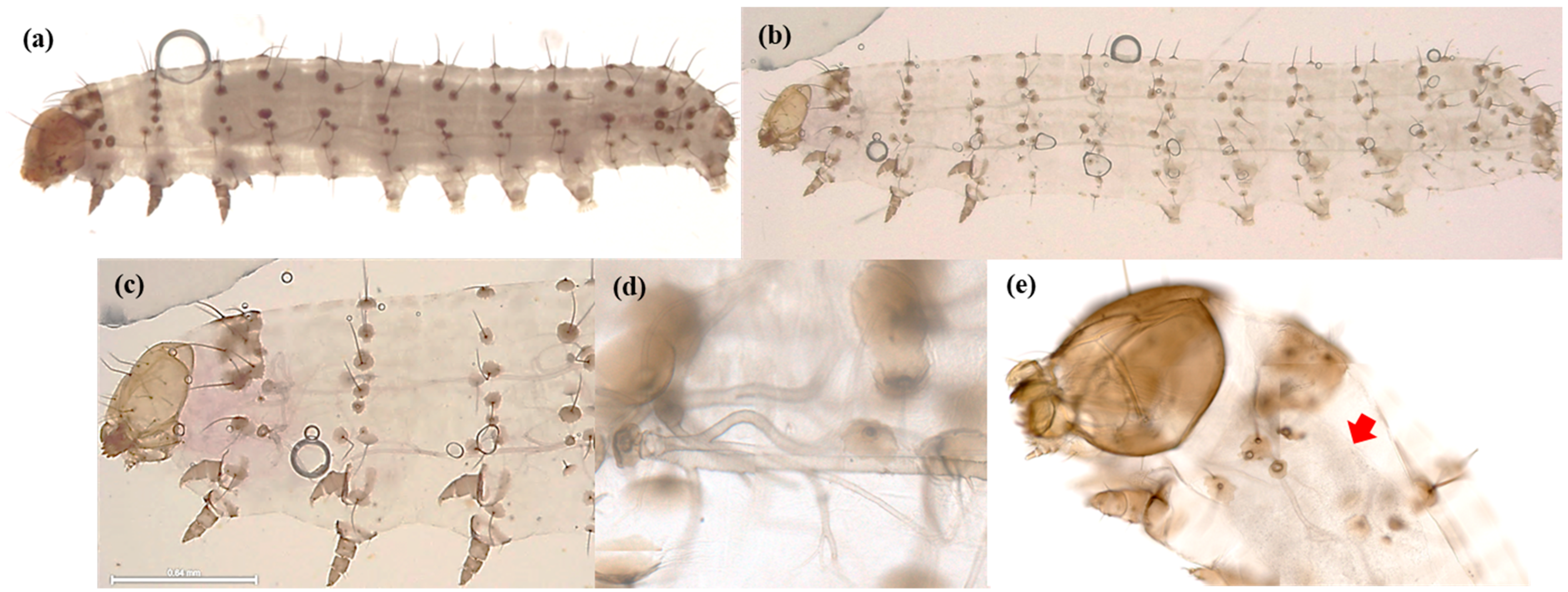
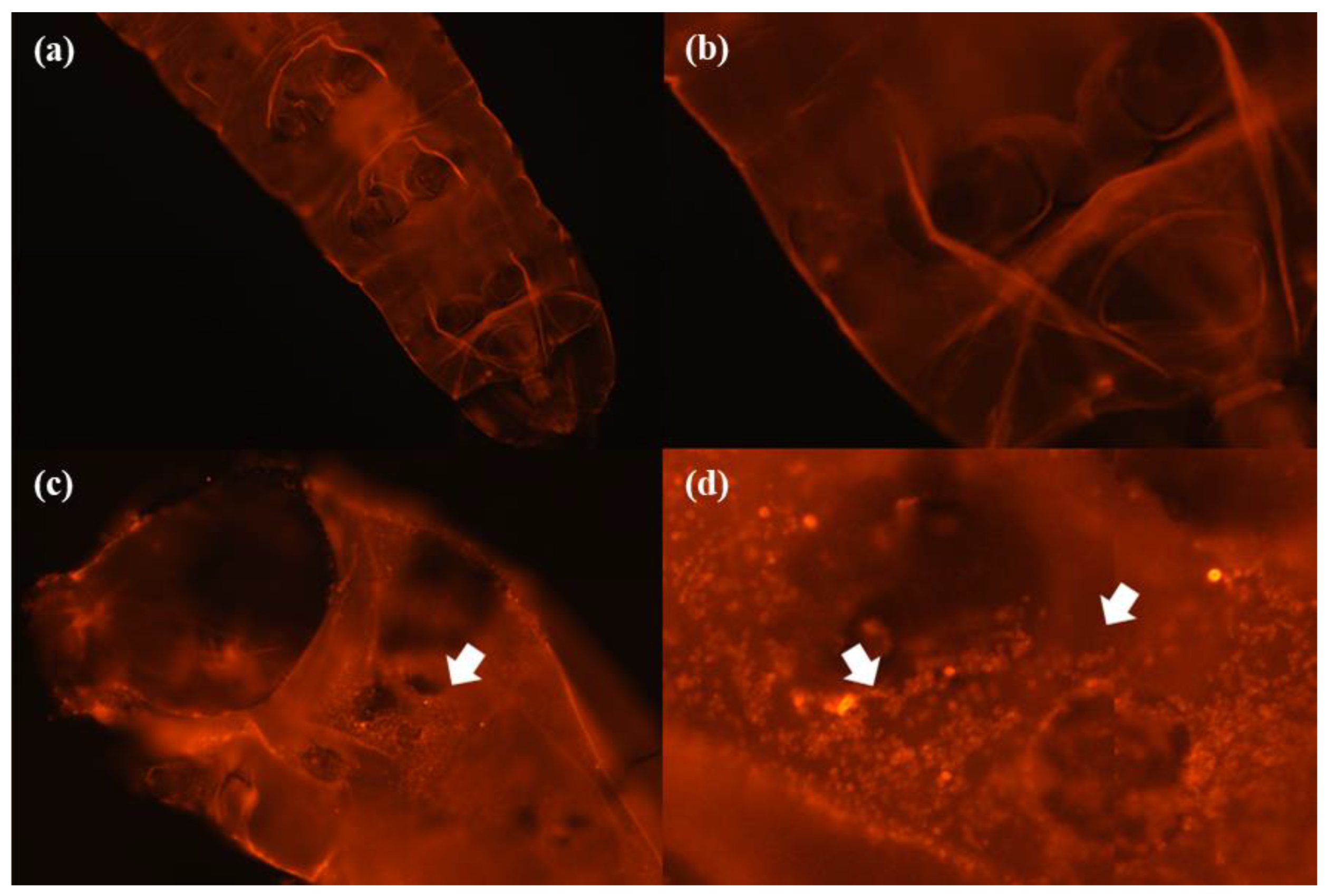
2.6. Microscopic Analysis
2.7. Biosynthesis Gene Cluster Prediction
3. Results
3.1. Species Identification
3.2. Microscope Analysis
3.2.1. External Morphology Analysis
3.2.2. Internal Morphology Analysis
3.3. Biosynthesis Gene Clusters Prediction
3.3.1. Beauveria bassiana
Bassianolide
Beauvericin
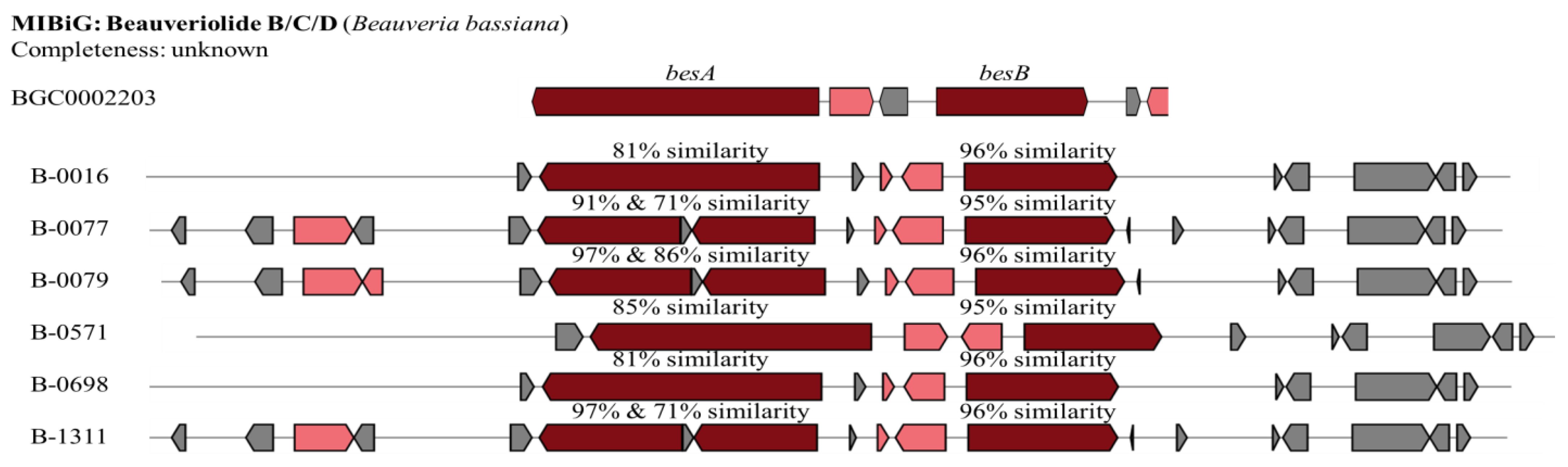
Beauveriolide B/C/D
Oosporein
3.3.2. Metarhizium Species
Destruxin
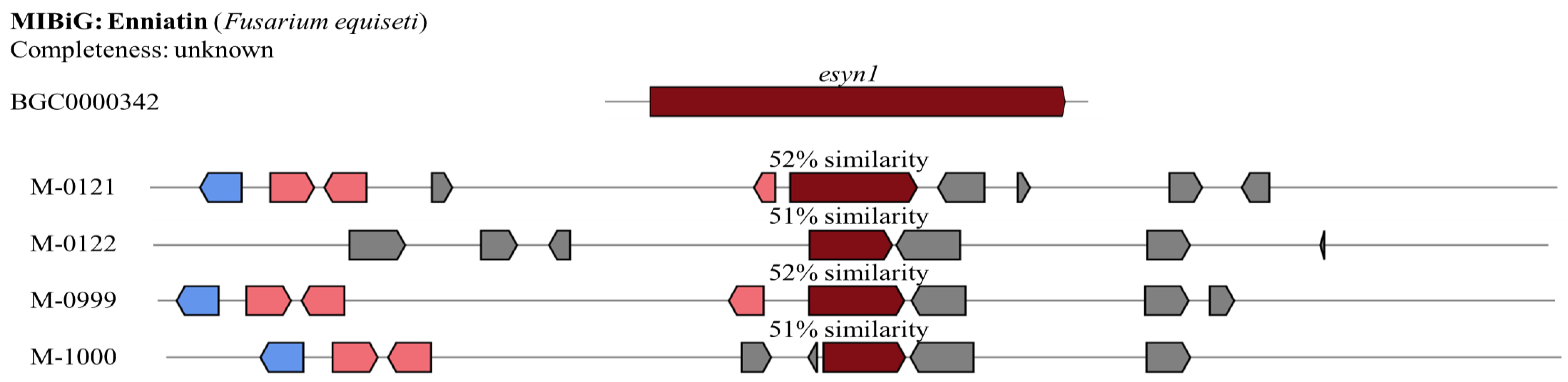
Enniatin
4. Discussion
4.1. Species Identification
4.2. Microscope Analysis
4.3. Biosynthesis Gene Cluster Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rane, R.; Walsh, T.K.; Lenancker, P.; Gock, A.; Dao, T.H.; Nguyen, V.L.; Khin, T.N.; Amalin, D.; Chittarath, K.; Faheem, M.; et al. Complex multiple introductions drive fall armyworm invasions into Asia and Australia. Sci. Rep. 2023, 12, 660. [Google Scholar] [CrossRef] [PubMed]
- Luginbill, P. The Fall ArmyWorm; U.S. Department of Agriculture: Washington, DC, USA, 1928; pp. 1–91. [Google Scholar]
- Sparks, A.N. A Review of the Biology of the Fall Armyworm. Fla. Entomol. 1979, 62, 82. [Google Scholar] [CrossRef]
- Sisay, B.; Sevgan, S.; Weldon, C.W.; Krüger, K.; Torto, B.; Tamiru, A. Responses of the Fall Armyworm (Spodoptera frugiperda) to Different Host Plants: Implications for Its Management Strategy. Pest Manag. Sci. 2023, 79, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Agriculture and Fisheries. Fall Armyworm. Available online: https://www.business.qld.gov.au/industries/farms-fishing-forestry/agriculture/biosecurity/plants/insects/field-crop/fall-armyworm (accessed on 26 June 2024).
- Department of Energy, Environment and Climate Action, Agriculture Victoria. Fall Armyworm—Agriculture. Available online: https://agriculture.vic.gov.au/biosecurity/pest-insects-and-mites/priority-pest-insects-and-mites/fall-armyworm (accessed on 1 July 2024).
- Department of Primary Industries and Regional Development, Government of Western Australia. Fall Armyworm in Western Australia. Available online: https://www.agric.wa.gov.au/fall-armyworm-western-australia?nopaging=1 (accessed on 29 June 2024).
- Shah, P.A.; Pell, J.K. Entomopathogenic Fungi as Biological Control Agents. Appl. Microbiol. Biotechnol. 2003, 61, 413–423. [Google Scholar] [CrossRef]
- Araújo, J.P.M.; Hughes, D.P. Diversity of Entomopathogenic Fungi: Which Groups Conquered the Insect Body? Adv. Genet. 2016, 94, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of Phytopathogenic Fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [PubMed]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic Fungi: Unconventional Applications. Rev. Environ. Sci. Biotechnol. 2020, 19, 23–42. [Google Scholar] [CrossRef]
- Chandler, D. Chapter 5—Basic and Applied Research on Entomopathogenic Fungi. In Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 69–89. ISBN 978-0-12-803527-6. [Google Scholar]
- Altinok, H.H.; Altinok, M.A.; Koca, A.S. Modes of action of entomopathogenic fungi. Curr. Trends Nat. Sci. 2019, 8, 117–124. [Google Scholar]
- Dar, S.A.; Rather, B.A.; Kandoo, A.A. Insect Pest Management by Entomopathogenic Fungi. J. Entomol. Zool. Stud. 2017, 5, 1185–1190. [Google Scholar]
- Trakimas, G.; Krams, R.; Krama, T.; Kortet, R.; Haque, S.; Luoto, S.; Eichler Inwood, S.; Butler, D.M.; Jõers, P.; Hawlena, D.; et al. Ecological Stoichiometry: A Link Between Developmental Speed and Physiological Stress in an Omnivorous Insect. Front. Behav. Neurosci. 2019, 13, 42. [Google Scholar] [CrossRef]
- Mondal, S.; Baksi, S.; Koris, A.; Vatai, G. Journey of Enzymes in Entomopathogenic Fungi. Pac. Sci. Rev. A Nat. Sci. Eng. 2016, 18, 85–99. [Google Scholar] [CrossRef]
- Caloni, F.; Fossati, P.; Anadón, A.; Bertero, A. Beauvericin: The Beauty and the Beast. Environ. Toxicol. Pharmacol. 2020, 75, 103349. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Moraga, E.; Vey, A. Bassiacridin, a Protein Toxic for Locusts Secreted by the Entomopathogenic Fungus Beauveria bassiana. Mycol. Res. 2004, 108, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Rosas-García, N.M.R.; Martínez, M.M.; Villegas-Mendoza, J.M. Detección de bassianolida y beauvericina en cepas de Beauveria bassiana y su participación en la actividad patogénica hacia Spodoptera sp. Biotecnia 2020, 22, 93–99. [Google Scholar] [CrossRef]
- Cheong, P. Bioactive Metabolites of an Isolate of the Entomopathogenic Fungus Beauveria bassiana. Ph.D. Thesis, Lincoln University, Lincoln, New Zealand, 2015. [Google Scholar]
- Kim, J.-C.; Hwang, I.M.; Kim, H.M.; Kim, S.; Shin, T.S.; Woo, S.-D.; Park, H.W. Rapid Analysis of Insecticidal Metabolites from the Entomopathogenic Fungus Beauveria bassiana 331R Using UPLC-Q-Orbitrap MS. Mycotoxin Res. 2024, 40, 123–132. [Google Scholar] [CrossRef]
- Fornelli, F.; Minervini, F.; Logrieco, A. Cytotoxicity of Fungal Metabolites to Lepidopteran (Spodoptera frugiperda) Cell Line (SF-9). J. Invertebr. Pathol. 2004, 85, 74–79. [Google Scholar] [CrossRef]
- Kanaoka, M.; Isogai, A.; Murakoshi, S.; Ichinoe, M.; Suzuki, A.; Tamura, S. Bassianolide, a New Insecticidal Cyclodepsipeptide from Beauveria bassiana and Verticillium lecanii. Agric. Biol. Chem. 1978, 42, 629–635. [Google Scholar] [CrossRef]
- Feng, P.; Shang, Y.; Cen, K.; Wang, C. Fungal Biosynthesis of the Bibenzoquinone Oosporein to Evade Insect Immunity. Proc. Natl. Acad. Sci. USA 2015, 112, 11365–11370. [Google Scholar] [CrossRef] [PubMed]
- Altimira, F.; Arias-Aravena, M.; Jian, L.; Real, N.; Correa, P.; González, C.; Godoy, S.; Castro, J.F.; Zamora, O.; Vergara, C.; et al. Genomic and Experimental Analysis of the Insecticidal Factors Secreted by the Entomopathogenic Fungus Beauveria pseudobassiana RGM 2184. J. Fungi 2022, 8, 253. [Google Scholar] [CrossRef] [PubMed]
- Abendstein, D.; Schweigkofler, W.; Strasser, H. Study on Insecticidal, Antifeedant and Growth Inhibitory Properties of Oosporein on Selected Pest Organisms. Insect Pathog. Insect Parasit. Nematodes 2003, 26, 103–106. [Google Scholar]
- Apirajkamol, N.; Hogarty, T.M.; Mainali, B.; Taylor, P.W.; Walsh, T.K.; Tay, W.T. Virulence of Beauveria Sp. and Metarhizium Sp. Fungi towards Fall Armyworm (Spodoptera frugiperda). Arch. Microbiol. 2023, 205, 328. [Google Scholar] [CrossRef] [PubMed]
- Apirajkamol, N.; Tay, W.T.; Mainali, B.; Taylor, P.; Walsh, T.K. High Molecular Weight DNA Extraction from Fungal Spores for Long Read Sequencing. Protocol.io. 2024. Available online: https://doi.org/10.17504/protocols.io.n92ld8ybnv5b/v1 (accessed on 11 April 2024).
- Nurk, S.; Walenz, B.P.; Rhie, A.; Vollger, M.R.; Logsdon, G.A.; Grothe, R.; Miga, K.H.; Eichler, E.E.; Phillippy, A.M.; Koren, S. HiCanu: Accurate Assembly of Segmental Duplications, Satellites, and Allelic Variants from High-Fidelity Long Reads. Genome Res. 2020, 30, 1291–1305. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness with Single-Copy Orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Minnis, A.M.; Sung, G.-H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and Systematics of the Anamorphic, Entomopathogenic Genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Humber, R.A.; Bischoff, J.F.; Rehner, S.A. Clarification of Generic and Species Boundaries for Metarhizium and Related Fungi through Multigene Phylogenetics. Mycologia 2014, 106, 811–829. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Tay, W.T.; Rane, R.V.; James, W.; Gordon, K.H.J.; Downes, S.; Kim, J.; Kuniata, L.; Walsh, T.K. Resistance Bioassays and Allele Characterization Inform Analysis of Spodoptera frugiperda (Lepidoptera: Noctuidae) Introduction Pathways in Asia and Australia. J. Econ. Entomol. 2022, 115, 1790–1805. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: New and Improved Predictions for Detection, Regulation, Chemical Structures and Visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Chen, B.; Song, S.; Li, B.; Yang, X.; Wang, C. Production of Diverse Beauveriolide Analogs in Closely Related Fungi: A Rare Case of Fungal Chemodiversity. mSphere 2020, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kang, Q.; Lu, Y.; Bai, L.; Wang, C. Unveiling the Biosynthetic Puzzle of Destruxins in Metarhizium Species. Proc. Natl. Acad. Sci. USA 2012, 109, 1287–1292. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biosynthesis Gene Clusters | CSIRO Isolates | Overall Similarity (%) | Core Biosynthesis Gene Similarity (%) | ||
|---|---|---|---|---|---|
| 1 | 2 | 3 | |||
| Beauveria bassiana | |||||
| Bassianolide | B-0016 | 53 | 85 | ||
| B-0077 | 60 | 97 | |||
| B-0079 | 53 | 97 | |||
| B-0571 | 66 | 97 | |||
| B-0698 | 60 | 88 | |||
| B-1311 | 53 | 97 | |||
| Beauvericin | B-0016 | 70 | 97 | ||
| B-0077 | 80 | 93 | |||
| B-0079 | 70 | 93 | |||
| B-0571 | 70 | 88 | |||
| B-0698 | 70 | 97 | |||
| B-1311 | 80 | 93 | |||
| Beauveriolide B/C/D | B-0016 | 66 | 81 | 96 | |
| B-0077 | 83 | 91 | 71 | 95 | |
| B-0079 | 66 | 97 | 86 | 96 | |
| B-0571 | 83 | 85 | 95 | ||
| B-0698 | 66 | 81 | 96 | ||
| B-1311 | 83 | 97 | 71 | 96 | |
| Oosporein | B-0016 | 85 | 89 | ||
| B-0077 | 85 | 91 | |||
| B-0079 | 85 | 89 | |||
| B-0571 | 85 | 92 | |||
| B-0698 | 85 | 89 | |||
| B-1311 | 85 | 91 | |||
| Metarhizium species | |||||
| Destruxin | M-0121 | 76 | - | 98 | - |
| Enniatin | M-0121 | 100 | 52 | ||
| M-0122 | 100 | 52 | |||
| M-0999 | 100 | 51 | |||
| M-1000 | 100 | 52 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apirajkamol, N.; Mainali, B.; Taylor, P.W.; Walsh, T.K.; Tay, W.T. Characterisation of the Pathogenicity of Beauveria sp. and Metarhizium sp. Fungi Against the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Agriculture 2025, 15, 170. https://doi.org/10.3390/agriculture15020170
Apirajkamol N, Mainali B, Taylor PW, Walsh TK, Tay WT. Characterisation of the Pathogenicity of Beauveria sp. and Metarhizium sp. Fungi Against the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Agriculture. 2025; 15(2):170. https://doi.org/10.3390/agriculture15020170
Chicago/Turabian StyleApirajkamol, Nonthakorn (Beatrice), Bishwo Mainali, Phillip Warren Taylor, Thomas Kieran Walsh, and Wee Tek Tay. 2025. "Characterisation of the Pathogenicity of Beauveria sp. and Metarhizium sp. Fungi Against the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae)" Agriculture 15, no. 2: 170. https://doi.org/10.3390/agriculture15020170
APA StyleApirajkamol, N., Mainali, B., Taylor, P. W., Walsh, T. K., & Tay, W. T. (2025). Characterisation of the Pathogenicity of Beauveria sp. and Metarhizium sp. Fungi Against the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Agriculture, 15(2), 170. https://doi.org/10.3390/agriculture15020170