
Nano-Titanium Dioxide Regulates the Phenylpropanoid Biosynthesis of Radish (Raphanus sativus L.) and Alleviates the Growth Inhibition Induced by Polylactic Acid Microplastics

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Cultivation of Radish
2.3. Determination of Morphological Indexes in Radish
2.4. Antioxidant Enzyme Activities in Radish
2.5. Transcriptome Sequencing and Data Analysis
2.6. Metabolomic Measurements and Data Analysis
2.7. Statistical Analyses
3. Results
3.1. Effect of Nano-TiO2 and PLA on Radish Growth
3.2. Effect of Nano-TiO2 and PLA on Radish Enzyme Activities
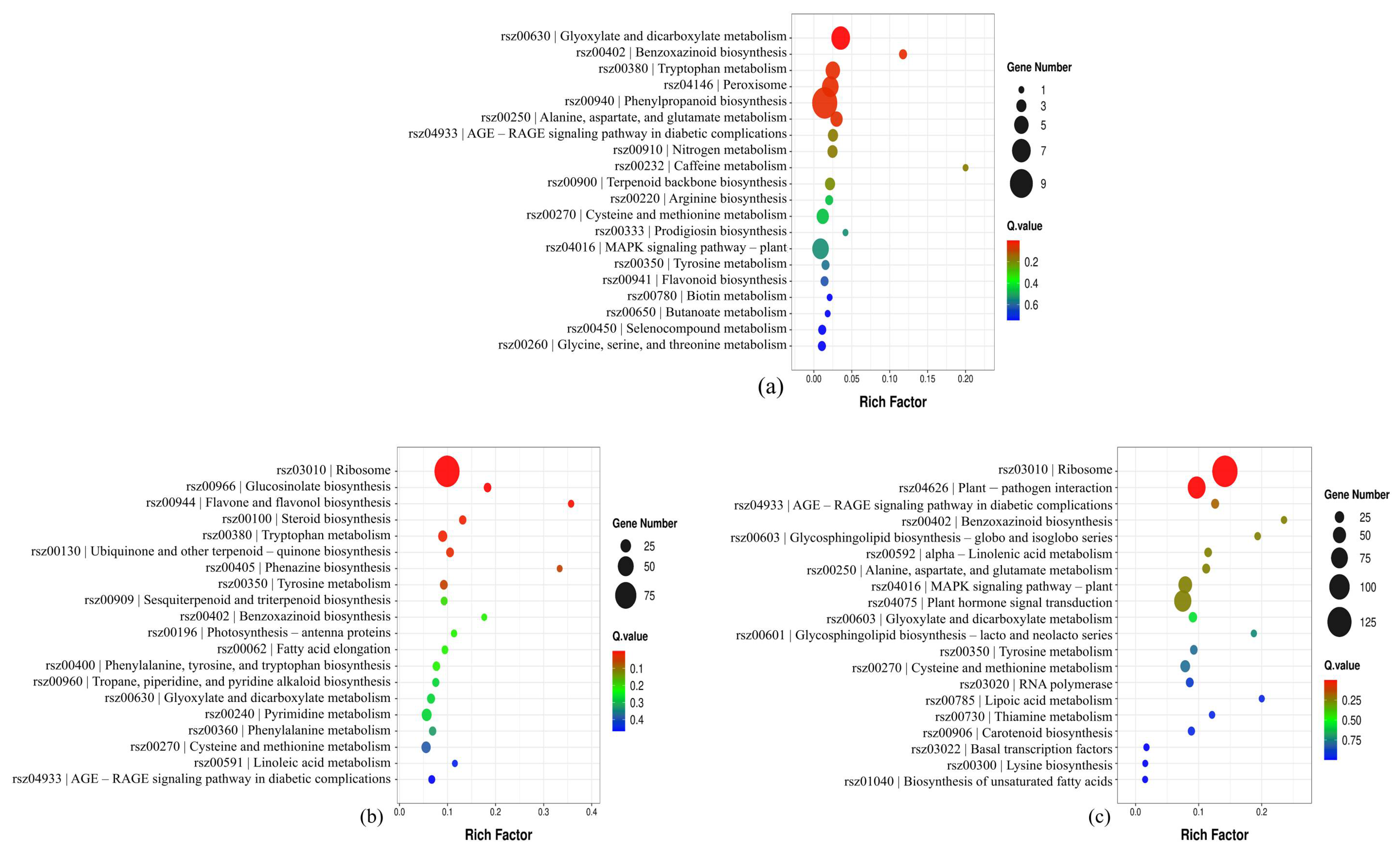
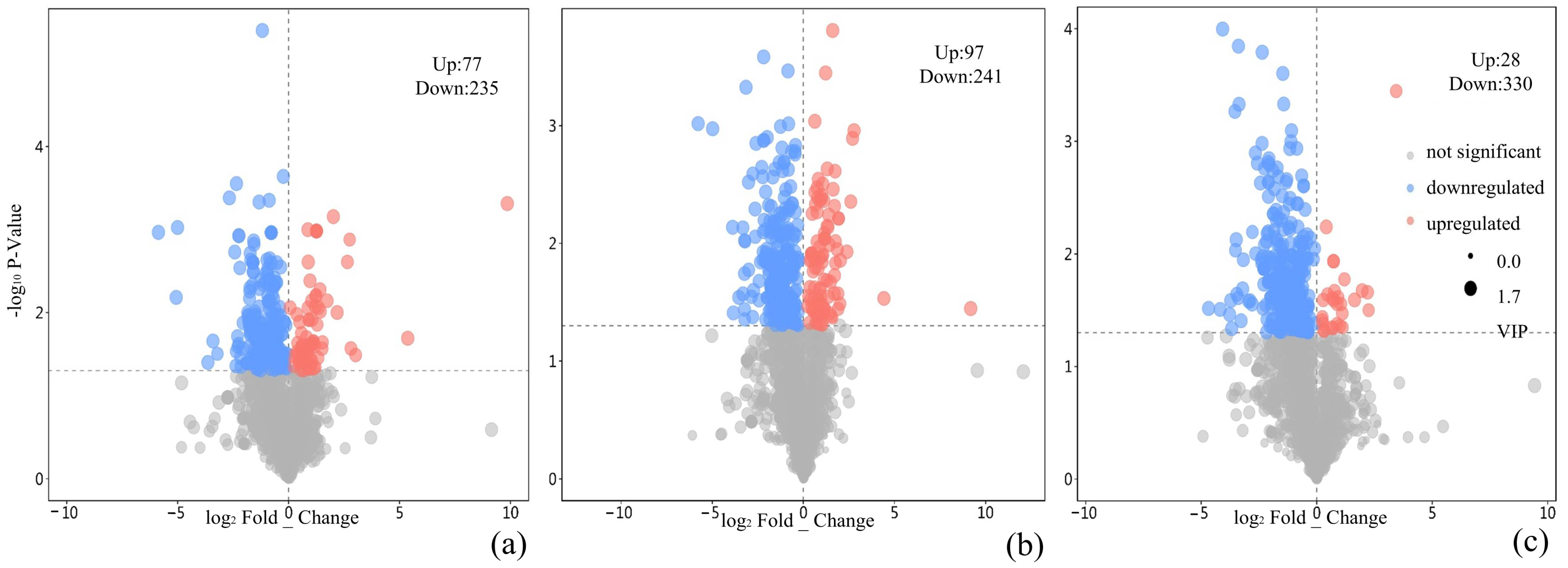
3.3. Transcriptomic Responses to PLA, Nano-TiO2, and Nano-TiO2 + PLA
3.4. Metabolomic Response After Treatment with PLA, Nano-TiO2, and Nano-TiO2 + PLA
4. Discussion
4.1. Germination and Growth Responses: Hormetic Effects and Composite Mitigation
4.2. Antioxidant Defense Dynamics: Enzyme Activation and Composite Interactions
4.3. Transcriptomic and Metabolomic Insights: Phenylpropanoid Pathway Activation Under Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Safdar, M.; Kim, W.; Park, S.; Gwon, Y.; Kim, Y.; Kim, J. Engineering plants with carbon nanotubes: A sustainable agriculture approach. J. Nanobiotechnol. 2022, 20, 275. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.; Venkatachalam, P.; Sahi, S.; Sharma, N. Reprint of: Silver and titanium dioxide nanoparticle toxicity in plants: A review of current research. Plant Physiol. 2016, 107, 147–163. [Google Scholar] [CrossRef]
- Shi, X.; Li, Z.; Chen, W. Fate of TiO2 nanoparticles entering sewage treatment plants and bioaccumulation in fish in the receiving streams. Nanoimpact 2016, 3, 96–103. [Google Scholar] [CrossRef]
- Yin, Z.F.; Wu, L.; Yang, H.G. Recent progress in biomedical applications of titanium dioxide. Phys. Chem. Chem. Phys. 2013, 15, 4844–4858. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Friot, D. Primary Microplastics in the Oceans: A Global Evaluation of Sources, 1st ed.; IUCN: Gland, Switzerland, 2017; p. 5. [Google Scholar] [CrossRef]
- Brahney, J.; Hallerud, M.; Heim, E. Plastic rain in protected areas of the United States. Science 2020, 368, 1257–1260. [Google Scholar] [CrossRef]
- Tang, K. Microplastics in agricultural soils in China: Sources, impacts and solutions. Environ. Pollut. 2023, 322, 121–235. [Google Scholar] [CrossRef]
- Tang, K.; Li, R.; Li, Z.; Wang, D. Health risk of human exposure to microplastics: A review. Environ. Chem. Lett. 2024, 22, 1155–1183. [Google Scholar] [CrossRef]
- Yang, L.; Luo, L.; Cai, W. Changes in carbohydrate metabolism and soil microorganisms under the stress of polyamide and polyethylene nanoplastics during rice (Oryza sativa L.) growth. Sci. Total Environ. 2024, 912, 169–183. [Google Scholar] [CrossRef]
- Khan, A.; Li, G. Micro/nanoplastics: Critical review of their impacts on plants, interactions with other contaminants (antibiotics, heavy metals, and polycyclic aromatic hydrocarbons), and management strategies. Sci. Total Environ. 2024, 912, 169420. [Google Scholar] [CrossRef]
- Yuan, X.; Zhang, B.; Zhang, F.; Wang, Z. Combined effects of micro/nanoplastics and ZnO nanoparticles on lactuca sativa seedlings under varied lighting. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2025, 296, 110233. [Google Scholar] [CrossRef]
- Yang, X.; Feng, K.; Wang, G.; Zhang, S.; Zhao, J.; Yuan, X.; Ren, J. Titanium dioxide nanoparticles alleviates polystyrene nanoplastics induced growth inhibition by modulating carbon and nitrogen metabolism via melatonin signaling in maize. Nanobiotechnology 2024, 22, 262. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Bai, X.; Weng, Y.; Kang, M.; Huang, Y.; Li, F.; Chen, Y. Phytotoxicity of binary nanoparticles and humic acid on (Lactuca sativa L). Environ. Sci. Process Impacts 2022, 24, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Issac, M.N.; Kandasubramanian, B. Effect of microplastics in water and aquatic systems. Environ. Sci. Pollut. 2021, 28, 19544–19562. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, X.; Ben, H.; Wang, L.; Fan, T.; Xie, X.; Shi, Y.; Li, B.; Chai, A. First Report of Radish Tuber Black Heart Rot Caused by Pectobacterium parvum in China. Plant Dis. 2025, 107, 2839. [Google Scholar] [CrossRef]
- Spitz, D.R.; Oberley, L.W. An assay for superoxide dismutase activity in mammalian tissue homogenates. Anal. Biochem. 1989, 179, 8–18. [Google Scholar] [CrossRef]
- Johansson, L.H.; Borg, L.A. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem. 1988, 174, 331–336. [Google Scholar] [CrossRef]
- Doerge, D.R.; Divi, R.L. Identification of the colored guaiacol oxidation product produced by peroxidases. Anal. Biochem. 1997, 250, 10–17. [Google Scholar] [CrossRef]
- Erofeeva, E.A. Environmental hormesis of non-specific and specific adaptive mechanisms in plants. Sci. Total Environ. 2022, 804, 150059. [Google Scholar] [CrossRef]
- Jaberzadeh, A.; Moaveni, P. Influence of bulk and nanoparticles titanium foliar application on some agronomic traits, seed gluten and starch contents of wheat subjected to water deficit stress. Not. Bot. Hort. Agrobot. Cluj. 2013, 41, 201. [Google Scholar] [CrossRef]
- Sehrish, A.K.; Ahmad, S.; Alomrani, S.O.; Ahmad, A.; Al-Ghanim, K.A.; Alshehri, M.A.; Tauqeer, A.; Ali, S.; Sarker, P.K. Nutrient strengthening and lead alleviation in (Brassica Napus L.) by foliar ZnO and TiO2-NPs modulating antioxidant system, improving photosynthetic efficiency and reducing lead uptake. Sci. Rep. 2024, 14, 19437. [Google Scholar] [CrossRef]
- Li, Q.; Yan, J.; Li, Y.; Liu, Y.; Andom, O.; Li, Z. Microplastics alter cadmium accumulation in different soil-plant systems: Revealing the crucial roles of soil bacteria and metabolism. J. Hazard. Mater. 2024, 474, 134768. [Google Scholar] [CrossRef] [PubMed]
- Bosker, T.; Bouwman, L.J.; Brun, N.R.; Behrens, P.; Vijver, M.G. Microplastics accumulate on pores in seed capsule and delay germination and root growth of the terrestrial vascular plant (Lepidium sativum). Chemosphere 2019, 226, 774–781. [Google Scholar] [CrossRef]
- Kumar, D.; Dhankher, O.P.; Tripathi, R.D.; Seth, C.S. Titanium dioxide nanoparticles potentially regulate the mechanism (s) for photosynthetic attributes, genotoxicity, antioxidants defense machinery, and phytochelatins synthesis in relation to hexavalent chromium toxicity in (Helianthus annuus L). J.Hazard. Mater. 2023, 454, 131418. [Google Scholar] [CrossRef] [PubMed]
- Shah, T.; Latif, S.; Saeed, F. Seed priming with titanium dioxide nanoparticles enhances seed vigor, leaf water status, and antioxidant enzyme activities in maize (Zea mays L.) under salinity stress. J. King Saud. Univ. Sci. 2021, 33, 101207. [Google Scholar] [CrossRef]
- Amini, S.; Maali-Amiri, R.; Mohammadi, R. cDNA-AFLP analysis of transcripts induced in chickpea plants by TiO2 nanoparticles during cold stress. Plant Physiol. Biochem. 2017, 111, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.D.; Liu, N.; Wang, W. Photosynthesis and related metabolic mechanism of promoted rice (Oryza sativa L.) growth by TiO2 nanoparticles. Front. Environ. Sci. Eng. 2020, 14, 103. [Google Scholar] [CrossRef]
- Chibani, K.; Gherli, H.; Fan, M. The role of blue light in plant stress responses: Modulation through photoreceptors and antioxidant mechanisms. Front. Plant Sci. 2025, 16, 1554281. [Google Scholar] [CrossRef]
- Ebrahimi, A.; Galavi, M.; Ramroudi, M. Effect of TiO2 nanoparticles on antioxidant enzymes activity and biochemical biomarkers in pinto bean (Phaseolus vulgaris L.). J. Mol. Biol. Res. 2015, 6, 58. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, H.; Liao, Y.; Ye, Z.; Li, M.; Klobučar, G. Ecotoxicity and genotoxicity of polystyrene microplastics on higher plant (Vicia faba). Environ. Pollut. 2019, 250, 831–838. [Google Scholar] [CrossRef]
- Larue, C.; Laurette, J.; Herlin-Boime, N. Accumulation, translocation and impact of TiO2 nanoparticles in wheat (Triticum aestivum spp.): Influence of diameter and crystal phase. Sci. Total Environ. 2012, 431, 197–208. [Google Scholar] [CrossRef]
- Liu, Z.; Qin, M.; Li, R.; Yang, L.; Liu, P.; Shi, Q. Transport Dynamics and Physiological Responses of Polystyrene Nanoplastics in Pakchoi: Implications for Food Safety and Environmental Health. J. Agric. Food Chem. 2025, 73, 10923–10933. [Google Scholar] [CrossRef] [PubMed]
- Rummel, C.D.; Jahnke, A.; Gorokhova, E.; Kühnel, D.; Schmitt-Jansen, M. Impacts of biofilm formation on the fate and potential effects of microplastic in the aquatic environment. Environ. Sci. Technol. Lett. 2017, 4, 258–267. [Google Scholar] [CrossRef]
- Cheng, X.; Yang, Z.; Ji, K.; Hu, Z.; Xi, Y.; Xiang, X. Enhanced copper adsorption by polyamide and polylactic acid microplastics: The role of biofilm development and chemical aging. Environ. Res. 2025, 282, 122040. [Google Scholar] [CrossRef]
- Gong, K.; Zhang, Q.; Shao, X.; Wu, Y.; Qiao, Z.; Qiu, L.; Zhang, W.; Peng, C. Microplastics alter Cr accumulation and fruit quality in Cr(VI) contaminated soil-cucumber system during the lifecycle: Insight from rhizosphere bacteria and root metabolism. Sci. Total Environ. 2024, 912, 168792. [Google Scholar] [CrossRef]
- Tian, X.; Weixie, L.; Wang, S.; Zhang, Y.; Xiang, Q.; Yu, X.; Zhao, K.; Zhang, L.; Penttinen, P.; Gu, Y. Effect of polylactic acid microplastics and lead on the growth and physiological characteristics of buckwheat. Chemosphere 2023, 337, 139356. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yan, C.; Shang, J.; Zeng, Z.; Huo, Y.; Li, D.; Xu, Y.; Xian, X.; Li, Y.; Gao, X.; et al. Microplastic mediated arsenic toxicity involves differential bioavailability of arsenic and modulated uptake in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2025, 300, 118448. [Google Scholar] [CrossRef]
- Arruda, S.C.; Silva, A.L.; Galazzi, R.M. Nanoparticles applied toplant science: A review. Talanta 2015, 131, 693–705. [Google Scholar] [CrossRef]
- Conway, J.R.; Beaulieu, A.L.; Beaulieu, N.L. Environmental stresses increase photosynthetic disruption by metal oxide nanomaterials ina soil-grown plant. ACS Nano 2015, 9, 11737–11749. [Google Scholar] [CrossRef]
- Yuan, X.; Zhang, F.; Wang, Z. Impacts of Micro/Nanoplastics Combined with Graphene Oxide on Lactuca sativa Seeds: Insights into Seedling Growth, Oxidative Stress, and Antioxidant Gene Expression. Plants 2024, 3, 3466. [Google Scholar] [CrossRef]
- Wan, R.; Wang, H.; Hui, T.; Yang, L.; Wang, X.; Cao, Y.; An, W.; Zhang, X.; Zhao, J.; Wang, Y.; et al. Morphological, physiological, and transcriptomic insights into response the of (Lycium barbarum L) seedlings to low-nitrogen stress. Genomics 2025, 117, 111065. [Google Scholar] [CrossRef]
- Gray, J.; Caparrós-Ruiz, D.; Grotewold, E. Grass phenylpropanoids: Regulate before using! Plant Sci. 2012, 184, 112–120. [Google Scholar] [CrossRef]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. Theevolution of phenylpropanoid metabolism in the green lineage. Crit.Rev. Biochem. Mol. Biol. 2013, 48, 123–152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Du, W.; Peralta-Videa, J. Metabolomics reveals howcucumber (Cucumis sativus) reprograms metabolites to cope with silver ions and silver nanoparticle-induced oxidative stress. Environ. Sci. Technol. 2018, 52, 8016–8026. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Doseff, A.I.; Grotewold, E. Flavones: From biosynthesisto health benefits. Plants 2016, 5, 27. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Dong, Q.; Ge, S.; He, X.; Verdier, J.; Li, D.; Zhao, J. Metabolic engineering of proanthocyanidin production by repressingthe isoflavone pathways and redirecting anthocyanidin precursor flux in legume. Plant Biotechnol. J. 2016, 14, 1604–1618. [Google Scholar] [CrossRef]
- Xu, G.; Li, X.; Zhu, T.; Wang, F.; Yin, J. When Nano and Microplastics Meet Taro (Colocasia esculenta) Roots: Their Size-Dependent Adsorption, Penetration, and Promotion on Secondary Wall Reinforcement. Environ. Sci. Technol. 2025, 59, 8345–8356. [Google Scholar] [CrossRef]
- Nakabayashi, R.; Saito, K. Integrated metabolomics forabiotic stress responses in plants. Curr. Opin. Plant Biol. 2015, 24, 10–16. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemicals | Concentrations | |
|---|---|---|
| nano-TiO2 (T) | 5 mg/L | 150 mg/L |
| PLA (P) | 5 mg/L | 150 mg/L |
| nano-TiO2 + PLA (T + P) | 5 mg/L + 5 mg/L | 5 mg/L + 150 mg/L |
| 150 mg/L + 5 mg/L | 150 mg/L + 150 mg/L | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, L.; Li, W.; Zhang, Y.; Liu, Z.; Yang, Y.; Guo, L.; Guo, C.; Yu, Z.; Fu, W. Nano-Titanium Dioxide Regulates the Phenylpropanoid Biosynthesis of Radish (Raphanus sativus L.) and Alleviates the Growth Inhibition Induced by Polylactic Acid Microplastics. Agriculture 2025, 15, 1478. https://doi.org/10.3390/agriculture15141478
Jiang L, Li W, Zhang Y, Liu Z, Yang Y, Guo L, Guo C, Yu Z, Fu W. Nano-Titanium Dioxide Regulates the Phenylpropanoid Biosynthesis of Radish (Raphanus sativus L.) and Alleviates the Growth Inhibition Induced by Polylactic Acid Microplastics. Agriculture. 2025; 15(14):1478. https://doi.org/10.3390/agriculture15141478
Chicago/Turabian StyleJiang, Lisi, Wenyuan Li, Yuqi Zhang, Zirui Liu, Yangwendi Yang, Lixin Guo, Chang Guo, Zirui Yu, and Wei Fu. 2025. "Nano-Titanium Dioxide Regulates the Phenylpropanoid Biosynthesis of Radish (Raphanus sativus L.) and Alleviates the Growth Inhibition Induced by Polylactic Acid Microplastics" Agriculture 15, no. 14: 1478. https://doi.org/10.3390/agriculture15141478
APA StyleJiang, L., Li, W., Zhang, Y., Liu, Z., Yang, Y., Guo, L., Guo, C., Yu, Z., & Fu, W. (2025). Nano-Titanium Dioxide Regulates the Phenylpropanoid Biosynthesis of Radish (Raphanus sativus L.) and Alleviates the Growth Inhibition Induced by Polylactic Acid Microplastics. Agriculture, 15(14), 1478. https://doi.org/10.3390/agriculture15141478