Abstract
Heat stress (HS) in dairy cattle triggers systemic physiological disruptions, including milk yield decline, immune suppression, and reproductive dysfunction, jeopardizing sustainable livestock production. While conventional single-omics or phenotypic studies have provided fragmented insights, they fail to decipher the multi-layered regulatory networks and gene–environment interactions underlying HS. This review integrates current knowledge on HS-induced physiological responses and molecular adaptations in dairy cattle from a multi-omics perspective, highlighting integrative approaches that combine IoT-enabled monitoring and AI-driven genetic improvement strategies. However, key challenges persist, such as complexities in bioinformatic data integration, CRISPR off-target effects, and ethical controversies. Future directions will emphasize the development and application of AI-aided predictive models to enable precision breeding, thereby advancing climate-resilient genetic improvement in dairy cattle.
1. Introduction
Amid global climate change, the increasing frequency of extreme heat events has pushed dairy cattle—temperate-origin ruminants with a thermoneutral zone (TNZ) threshold of 5–25 °C—beyond their thermal comfort limits, leading to severe heat stress (HS) [1,2]. Prolonged HS has become a major constraint on the sustainable development of livestock production [3]. This condition causes multidimensional damage to dairy cows, including reduced dry matter intake [4,5], negative energy balance, immune dysfunction [6], and reproductive endocrine disruption [7]. Although conventional studies have identified key physiological responses to HS, the molecular regulatory networks underlying these adaptations—including their tissue- and cell-specific dynamics, stage-dependent variations, and gene–environment interactions—remain poorly characterized.
Early research primarily used single-omics approaches to investigate HS mechanisms, such as metabolomics-based tracking of energy metabolism markers like β-hydroxybutyrate [8]. Genomic studies have identified genetic loci associated with HS, revealing, for example, that miRNA-541 modulates the expression of Heat Shock Protein 27 (HSP27) under thermal stress [9]. However, HS induces systemic physiological dysregulation through dynamic, multi-layered biological processes (e.g., signal transduction, metabolic adaptation) that cannot be comprehensively characterized by single-omics approaches.
Recent advancements in multi-omics (e.g., genomics, transcriptomics, proteomics, metabolomics) integration have established a new framework for systematically analyzing complex traits by correlating genomic variations, epigenetic modifications, metabolic pathways, and phenotypic data [10]. These innovations emphasize the crucial role of multi-omics technologies in unraveling the interconnected “genotype-environment-phenotype” relationship. Simultaneously, the merging of Internet of Things (IoT) and artificial intelligence (AI) technologies has opened new possibilities in HS research. Sensors allow for the real-time monitoring of microenvironmental parameters (e.g., temperature, humidity) and behavioral traits (e.g., feed intake, activity levels) in individual cows [11], providing high-resolution spatiotemporal datasets. AI-based data fusion models additionally optimize multi-omics input integration to allow heterogeneous data to be synthesized across temporal as well as spatial coordinates. These enable key thresholds of environmental interactions to be identified at the level of genes, such as thermal humidity index thresholds. In addition to enhancing phenotypic predictability, these technological advances further enable precision breeding approaches. At the level of genetic intervention, advances at the level of gene-editing technologies like CRISPR-Cas9 have made it possible to implement resilience breeding. Initial research indicates that the technologies are able to introduce modifications to key nodes in HS regulatory networks to increase dairy cattle’s thermotolerance [12]. Off-target effects, epigenetic instability, as well as some concerns regarding ethics, however, present impediments to the large-scale use of technologies [13].
This review condenses existing understanding of the physiological adaptation and resilience responses of dairy cattle subjected to HS at the multi-omics level. The combined uses of genomics, transcriptomics, metabolomics, and epigenomics are first laid out to unravel pathways of HS. Molecular networks underlying temperature perception, signal transduction, and metabolic adaptation of heat-stressed cows are next addressed. The potential and limitations of gene-editing technologies and smart selection methods to enhance thermotolerance are finally critically evaluated.
2. Multidimensional Analysis of Physiological Responses to HS in Dairy Cows
2.1. Impact on Production Performance
Under current climatic projections for 2.7 °C annual temperature rises, the incidence of extreme heat events has doubled every decade since 1979, significantly increasing dairy cattle exposure to hyperthermal stress [14]. Heat perception in bovines is mediated by hypothalamic integration of external stimuli, and optimal production performance is achieved within the thermal neutral zone (TNZ), where physical and behavioral heat tolerance reach thermal equilibrium [15]. When homeostasis is lost, cattle develop heat stress (HS) [15]. Substantial evidence attests to HS-generated production losses such as decreased milk production, changes to the nature of the milk produced, decreased estrus persistence, and compromised fertility [16]. Notably, metabolic heat generated by lactation adds to thermal loading and energy burdens [17]. In this chapter, the relationship between thermoregulatory demand and adaptations to this demand under HS is explored, with alternative factors of production performance reduction discussed.
Extensive evidence confirms that HS typically reduces dry matter intake (DMI) by more than 30% in lactating cows, a response more pronounced than in most other livestock species, thereby intensifying metabolic challenges during periods of thermal stress [5]. Initially, this reduced feed intake was interpreted as an adaptive response aimed at minimizing heat production from metabolism and digestion [15]. However, more recent insights suggest that the mechanism is far more complex, involving various metabolic trade-offs. This aligns with research by Chen et al. [18] and Sguizzato et al. [19], which indicates that pregnant dairy cows experience higher digestive and metabolic demands compared to non-pregnant cows, and that heat production during lactation is significantly greater than during the dry period. Notably, recent findings from Wheelock et al. [5] show that the reduction in DMI accounts for only 50% of the HS-induced milk yield depression, with the remaining 50% of the production loss being linked to altered carbohydrate metabolism. These findings compel a paradigm shift from viewing DMI reduction as purely adaptive to recognizing it as part of broader metabolic disruption.
Rumen fermentation have dual roles in dairy production: supplying milk precursors while generating metabolic heat that exacerbates thermal stress [20]. Under conditions of hyperthermia, defective carbohydrate metabolism induces an underlying energy deficit, manifesting as negative energy balance (NEB) revealed by elevated levels of β-hydroxybutyrate (BHB)—an established biomarker of energy deficiency. The effect was quantitatively confirmed by GC-MS metabolomics by Guo et al. [8], noting marked increases in BHB levels among HS-treated cattle. In addition, the study of metagenomics identifies that HS changes the microbial composition of the rumen, specifically suppressing the growth of Clostridium butyricum and interfering with pathways for the production of butyrate [21], thereby impairing an important source of energy for metabolism by the mammary gland. In combination, these data identify NEB as an underlying mechanism for changes in production performance with HS.
Beyond thermal stress and NEB, HS impairs dairy productivity through multifaceted mechanisms, with recent studies elucidating the molecular basis of these production declines. Not only does HS disrupt metabolic homeostasis and energy balance by oxidative stress and NF-κB-mediated inflammation [22], HS strongly upregulates Heat Shock Proteins (HSPs) [23] that serve dual protective and reparative functions. The upregulation of Heat Shock Protein 70 (HSP70) is an adaptive mechanism to thermal stress, counteracting protein misfolding and apoptosis [24]. We propose that HS-induced production losses are caused by a complex interplay between metabolic energy redistribution and intracellular stress responses, where the key residing in balancing self-preservation mechanisms versus metabolic efficiency. Future research should unraveling the crosstalk between cellular signaling networks under various HS conditions and explore genetic editing and molecular interventions to enhance thermotolerance and productivity in dairy cattle.
2.2. Immune Barrier Collapse
In the initial stage of HS, physiological responses are characterized by high rectal temperature (RT), higher respiratory rate (RR), and higher drooling scores (DS) [25]. As HS develops to the exhaustion stage, immune function is more seriously impacted by it, where it presents as immune cell dysregulation both in quantity and function [26]. These ultimately impair innate immunity, exacerbate oxidative stress, and elevate inflammatory risks. We examine the mechanisms driving these HS-induced shifts in immune cell populations and function as follows. Previous studies have correlated changes in the immune levels of heat-stressed dairy cows with blood cortisol concentrations [27], suggesting that their immune cells are influenced by cortisol. During HS, hypothalamic secretes corticotropin-releasing hormone (CRH) stimulates pituitary adrenocorticotropic hormone (ACTH) release, activating adrenal glucocorticoid and stress hormone secretion [28]. Increased cortisol levels of dairy cattle undergoing HS were identified by analysis of blood GC-MS [8], thus verifying activation of the hypothalamo–pituitary–adrenal (HPA) axis under thermal stress. As an important glucocorticoid, cortisol directly influences immune cells with several studies reporting significant immunomodulatory actions of cortisol. These effects include promoting neutrophil apoptosis, altering macrophage polarization (M1/M2 ratio), reducing lymphocyte counts, and suppressing antigen-presenting cell concentrations [29]. Flow cytometry shows cortisol upregulates Annexin1 (A1) expression [30] while downregulating L-selectin on neutrophils, facilitating their bone marrow mobilization [31]. Cortisol levels peak early during HS but decline as stress progresses [28]. Blood profiling of thermotolerant Zebu cattle further reveals a correlation between higher neutrophil counts and lower cortisol concentrations [32], suggesting that high cortisol levels suppress neutrophil populations, while reduced cortisol levels facilitate their mobilization from bone marrow.
Notably, compensatory increases in immune cell numbers may not fully restore immune function, as HS impairs immune cell functionality. For instance, although neutrophils typically executed antimicrobial functions via the Mitogen-Activated Protein Kinase (MAPK) signaling pathway, research by Ricci et al. [33] implicitly indicated that this pathway becomes inhibited under HS. Macrophage polarization is also affected: M1 macrophages, which rely on NF-κB signaling for pro-inflammatory and antimicrobial responses, and M2 macrophages, which exert anti-inflammatory effects through STAT6 phosphorylation-mediated gene transcription [34], exhibit altered polarization during HS. Reports from Lacetera et al. [35] further demonstrated that HS induces a Th1/Th2 imbalance, with reduced Th1 cell numbers and a Th2-dominant response that typically favors humoral immunity. However, this Th2 bias does not enhance humoral immunity, as CD21+ B lymphocyte populations decline, impairing antigen presentation to T cells and subsequent Th2 differentiation [36]. As a result, both innate and adaptive immune functions are compromised under HS, increasing disease susceptibility.
While hormonal changes have traditionally explained HS-induced immune alterations in dairy cattle, the multi-omics technologies provide novel insights. HS induces oxidative stress, causing the over-production of reactive oxygen species (ROS) and intracellular redox imbalance [37]. Not only does ROS directly compromise immune cell function, but it also modulates immune-related gene expression through epigenetics and transcriptional regulation [38]. For instance, the transcriptome analyses reveal significantly upregulation of pro-inflammatory cytokines in peripheral blood mononuclear cells, while simultaneously suppressing the activity of the antioxidant enzyme superoxide dismutase (SOD) and glutathione peroxidase (GPx) [39], demonstrating the pivotal role of oxidative stress in immune malfunctioning caused by HS. Proteomics studies demonstrate that oxidative stress significantly changes the expression pattern of immune-related proteins. Increased levels of acute-phase proteins in serum from heat-stressed cows indicate activation of systemic inflammation [40]. Metabolomics reveals that oxidative stress profoundly affects immune cell energy metabolism. HS triggers major metabolic alterations, particularly in energy and antioxidant pathways, including elevated lactate and pyruvate coupled with depleted glutathione and ascorbate (reflecting antioxidant exhaustion) [41]. These changes impair immune cell energy supply while exacerbating oxidative damage to immune function.
Immune dysfunction in heat-stressed dairy cattle primarily results from hormone-mediated changes in immune cell populations and oxidative stress impacting cellular function through diverse signaling pathways. Most evidence linking gut and immune changes under HS highlights increased intestinal permeability and LPS translocation [42]. Studies in murine models suggest microbial SCFAs modulate immune responses by recruiting neutrophils [43] and promoting Th1 differentiation [44], which direct studies in cattle are scarce. Notably, stress-induced microbiota shifts may reduce SCFA output, indirectly affecting immune regulation [45].
2.3. Endocrine Alterations
The endocrine system is an overall regulator of homeostasis, and it is responsible for the response to HS. Elevated insulin levels alter energy metabolism pathways [5], which may contribute to negative energy balance (NEB) in dairy cows under HS. Concurrently, plasma prolactin (PRL) levels increase significantly during HS [7], possibly serving as an oxidative stress defense mechanism in mammary cells. This section discusses endocrine alterations in HS-exposed cows, focusing on nutrient metabolism and reproductive physiology.
Existing research indicates that there are higher levels of basal insulin along with an increase in biological activity [46]. Under HS, skeletal muscle and adipose tissue rely more on insulin-stimulated GLUT4-mediated glucose uptake [47], indicating compensatory insulin hypersecretion to enhance peripheral glucose storage. HS induces an increase in circulating norepinephrine and epinephrine [48], which act as catabolic signals promoting lipolysis. Although this response should theoretically shift glucose toward mammary gland use [49], heat-stressed cows show paradoxical metabolic adaptations. Despite elevated catecholamines, increased insulin activity coincides with reduced fat mobilization [5,50], indicating insulin-mediated glucose partitioning overrides lipolytic signals. Heat-stressed cows show marked decreases in circulating nonesterified fatty acids (NEFA) [51,52], consistent with insulin-mediated suppression of lipolysis. This dual metabolic disruption—suppressed lipid mobilization coupled with declining blood glucose levels—worsens NEB in heat-stressed cows. Interestingly, metabolic profiles differ fundamentally between heat-stressed and thermoneutral cows under NEB. Under thermoneutral conditions, early-lactation cows maintain energy balance via catecholamine-driven NEFA mobilization while downregulating insulin signaling [53].
The mechanisms underlying insulin regulation in heat-stressed dairy cattle remain controversial. Two divergent perspectives are evident in the literature: (1) the duration of hyperinsulinemia appears stressor-dependent [52], and (2) elevated PRL and blood calcium levels may induce hyperinsulinemia in these animals [54]. While increased PRL levels in heat-stressed cows are well-documented [54], direct evidence linking PRL fluctuations to insulin dynamics is lacking. Based on current findings, we propose that HS induces insulin dysregulation through complex endocrine interactions rather than a single pathway. Insulin suppresses catecholamine-driven lipolysis, shifting energy dependence toward glucose catabolism and worsening NEB. This metabolic shift starkly contrasts with thermoneutral adaptive strategies, revealing unique regulatory patterns imposed by HS.
HS also has significant effects on the reproductive system. HS was reported to significantly reduce the preovulatory luteinizing hormone (LH) surge [55]. Additional evidence revealed that elevated cortisol levels indirectly disrupt LH surge development by suppressing the release of the gonadotropin-releasing hormone (GnRH) [56], reflecting the multi-hormonal interference of HS on reproductive signaling. LH suppression is developmentally regulated: heifers show pronounced attenuation, whereas mature cows are less affected [57], suggesting a link between LH dynamics and reproductive maturity. Additionally, transcriptomic and proteomic analyses reveal that HS downregulates key steroidogenic genes (e.g., STAR, CYP19A1) in ovarian tissues [10], ultimately reducing estradiol production. Although FSH levels increase under HS [58], the estrogen synthesis requires the synergy of both LH and follicle-stimulating hormone (FSH) [59]. Thus, elevated FSH alone cannot compensate for the lack of estradiol. Conflicting data exist regarding progesterone dynamics: some studies report no significant change, while others observe increases or decreases [60,61]. These discrepancies may be attributable to variations in HS intensity, DMI, and luteal function. Metabolomics data confirm this relationship, showing energy depletion disrupts hormonal synthesis [62]. Heat-stressed cows exhibit reduced progesterone levels, though folate supplementation have the potential to mitigate this effect [63]. In conclusion, dairy cattle under HS exhibit a characteristic reproductive endocrine profile: suppressed GnRH, LH, progesterone, and estradiol alongside elevated FSH. While thermotolerant breeds possibly preserve fertility by sustaining estradiol to offset GnRH inhibition, the underlying metabolic and molecular mechanisms require further investigation.
3. Mechanisms Underlying HS Generation from a Multi-Omics Perspective
3.1. Investigating Cellular Physiological Alterations Through Thermosensation Mechanisms
Thermal stress induction requires specialized temperature sensing. Initial detection occurs via TRP-expressing afferent neurons, triggering regulatory responses to maintain homeostasis [64]. The following is an analysis of the physiological changes at the cellular level of heat-stressed dairy cows from the standpoint of TRP channel-mediated temperature perception (Figure 1).
Figure 1.
Diagram of the physiological response mechanisms to HS in dairy cows. Dotted lines indicate inhibitory effects, while solid lines represent promotive effects. The red × indicates that the pathway is inhibited. Transient Receptor Potential Vanilloid (TRPV), Transient Receptor Potential Melastatin (TRPM), Permeability Transition Pore (PTP), AMP-activated Protein Kinase (AMPK), Ca2+/calmodulin-dependent Protein Kinase Kinase (CaMKK), Nicotinamide Adenine Dinucleotide Phosphate (NADPH), NADPH Oxidase (NOX), Inhibitor of Nuclear Factor Kappa B Alpha (IκBα), Sirtuin 1 (SIRT1), NF-κB p65 Subunit (p65), Heat Shock Factor (HSF), Heat Shock Protein (HSP), Bcl-2-Interacting Mediator of Cell Death (Bim), cellular Jun Proto-oncogene (c-Jun), c-Jun N-terminal Kinase (JNK), p38 Mitogen-Activated Protein Kinase (p38), Activator Protein-1 (AP-1), Inhibitor of Nuclear Factor Kappa-B Kinase (IKK), Myeloid differentiation primary response 88 (MyD88), Toll-Like Receptor 4 (TLR4), Histone Deacetylase (HDAC), Cluster of Differentiation 14 (CD14), Lipopolysaccharide (LPS).
TRP channels work by regulating Ca2+ levels to affect membrane potential and transduce signals [65]. Transcriptome and genome analysis has divided mammalian TRP channels into nine families with the TRPV (Vanilloid) and TRPM (Melastatin) subfamilies being thermosensitive [66]. Dairy cows also possess TRPV and TRPM channels across skin, oral mucosa, and visceral organs that detect temperature changes, regulate Ca2+ levels, reshape cellular signaling, and contribute to thermal stress adaptation [67]. Furthermore, TRPV3 and TRPV4 are activated over intermediate thermal values (22–40 °C) and transmit cutaneous thermoadaptation [68]. Notably, research indicated that the thermal stress threshold in dairy cattle begins at 26 °C (50% relative humidity, temperature humidity index (THI) = 72) [69], suggesting that the early stages of HS primarily involve the activation of TRPV3 and TRPV4 channels. Among these, TRPV4 critically regulates hypothalamic thermoregulation by promoting sweating and vasodilation to enhance heat dissipation [68]. Conversely, TRPV3 is predominantly expressed in keratinocytes, suggesting its involvement in epidermal thermal adaptation [70]. As temperatures continue to rise, TRPV1 activation triggers an influx of intracellular Ca2+, leading to the expression of HSPs that help mitigate cellular damage [71]. Intriguingly, heterologous expression studies in oocytes reveal that TRPV4-mediated Ca2+ influx can lead to mitochondrial overload [68], disrupting oxidative phosphorylation and causing electron leakage from the respiratory chain [72,73]. This cascade triggers an ROS burst. Proteomic and metabolomic analyses in heat-stressed poultry show that excessive ROS production results in oxidative modifications of mitochondrial proteins [74], thereby disrupting cellular energy metabolism. Consistent with this, Rhoads et al. [75] report that environmental stressors trigger ROS-mediated denaturation/inactivation of ATP synthase and TCA cycle enzymes, impairing bioenergetics. Excessive Ca2+ and ROS synergistically promote prolonged mitochondrial PTP opening [76], causing swelling and outer membrane rupture [77,78]. Ca2+ overload exacerbates mitochondrial outer membrane permeabilization [79], releasing cytochrome c (Cyt c) and triggering apoptosis.
It remains unclear whether mitochondrial Ca2+ influx in heat-exposed animals is primarily mediated by TRP channels. Vais et al. [80] suggested that mitochondrial Ca2+ uptake was largely regulated by the Mitochondrial Calcium Uniporter (MCU), whereas other studies proposed that TRP channels modulate Ca2+ influx under oxidative stress conditions [81]. RNA-seq comparison of TRP channels and MCU genes in heat-stressed versus thermoneutral cow mitochondria could reveal key insights. If TRP genes are upregulated with stable or reduced MCU expression they would indicate TRP-mediated Ca2+ regulation. Notably, the TRPM2 gene variant rs527146862 (Met269Val), associated with thermosensation, exhibits geographic adaptation in cattle populations (e.g., Dehong, Hainan, and Wenshan cattle), with higher gene missense mutation frequencies in southern regions exposed to chronic HS—potentially enhancing thermal tolerance [82]. These underscore the potential of modulating TRP gene expression to breed heat-resistant dairy cattle and mitigate HS.
3.2. The Impact of Physiological Changes on Signal Transduction
Temperature perception triggers a cascade of physiological changes, corresponding closely to intracellular signal transduction mechanisms. The following discusses the signal transduction dynamics in dairy cattle under HS from the viewpoint of cellular physiology changes.
ROS can oxidize the active site (Cys258) of MAPK phosphatases-1 (MKP-1) [83], blocking its dephosphorylation function, prolonging p38 and JNK phosphorylation, causing cytotoxicity via cell abnormalities. To combat heat-induced cytoanomalies, the body triggers transient p38 phosphorylation, activating downstream kinases and promoting HSP27 nuclear translocation to stabilize the cytoskeleton [84]. Additionally, persistent phosphorylation of JNK activates c-Jun, decreasing pro-apoptotic Bim and inducing apoptosis via the mitochondrial pathway [85]. Beyond the MAPK pathway, ROS generated by NADPH oxidation can initiate the caspase cascade through the intrinsic apoptotic pathway, resulting in cell death [86]. Transcriptomic data show that elevated temperatures increase the mRNA expression of NADPH oxidase (NOX), thereby promoting NADPH oxidation and ROS production [87,88]. Intriguingly, some studies suggest that NOX may exhibit Ca2+-dependent characteristics [89]. Moreover, murine studies have shown that Ca2+/calmodulin-dependent kinase kinase β (CaMKKβ) directly phosphorylates AMPKα (Thr172), activating AMPK and enhancing lipid metabolism [90]. Simultaneously, the AMPK pathway promotes PGC-1α-driven mitochondrial biogenesis [91]. Notably, if confirmed in heat-stressed dairy cattle, this pathway suggests Ca2+ triggers both mitochondrial damage and biogenesis, potentially reflecting the attempt of cell to maintain homeostasis amid energy metabolic dysregulation. NF-κB signaling alterations further support this homeostatic adaptation. For instance, during inflammation, ROS oxidize the Cys179 residue of IκBα, leading to its ubiquitination and degradation [92,93], releasing NF-κB into the nucleus and inducing the expression of IL-6 and TNF-α [92]. Conversely, HSP70 binds to the IKK complex, inhibiting NF-κB signaling, while HSP90 stabilizes the HSF1 trimer, enhancing HSP transcription [94,95], thereby suppressing inflammatory effects. Also, in the NFκB signaling pathway, transcriptome data suggest that LPS activation of NF-κB upregulates expression of IL-6 mRNA [96]. These findings indicate that while Ca2+ influx primarily activates MAPK, AMPK and NF-κB under HS, and ROS and epigenetics trigger overlapping adaptive responses. Concurrently, HS-induced gut barrier dysfunction promotes LPS-TLR4/MyD88 signaling, activating IKK to amplify inflammation [97]. Metagenomic studies show that HS alters the Firmicutes/Bacteroidetes ratio in the gastrointestinal tract of dairy cattle, reducing the levels of short-chain fatty acids [98,99]. The reduction in butyrate leads to decreased histone deacetylase (HDAC) activity [100,101], which in turn promotes the NF-κB signaling pathway [102]. Beyond inflammation, HS activates crosstalking signaling pathways in dairy cattle, exacerbating inflammatory responses. Excessive ROS stimulates MAPK (JNK/p38) and enhances AP-1 through c-Jun phosphorylation [103], which synergizes with NF-κB to further amplify the inflammatory response. During inflammation, dairy cattle activate AMPK, which antagonizes NF-κB via SIRT1-mediated deacetylation (Lys310) of p65, suppressing its pro-inflammatory activity [104]. Thus, the MAPK, AMPK, and NF-κB pathways in heat-stressed dairy cattle interact through both synergistic and antagonistic mechanisms to regulate their effects. Thus, HS triggers crosstalk among MAPK, AMPK, and NF-κB pathways in cattle. Notably, p38-mediated HSF1 phosphorylation modulates HSP70 activity, releasing JNK/p38 from inhibition and driving apoptosis [105].
NF-κB signaling disrupts dairy cattle physiology by promoting inflammation and oxidative stress, which MAPK exacerbates while AMPK mitigates. Apart from causing inflammation, the AMPK signaling pathway induces FOXO3a to increase the expression of anti-apoptotic factors including Bcl-2 and MnSOD [106,107], finally suppressing apoptosis as well as boosting cellular antioxidant capability. These findings suggest that HS triggers Ca2+/ROS-dependent MAPK activation, which cooperates with NF-κB to drive inflammation and apoptosis in cattle. AMPK opposes these effects and restores energy balance. While genomic analyses highlight MAPK’s role [108], AMPK’s regulatory potential, particularly its crosstalk with MAPK/NF-κB, needs deeper investigation.
4. Multi-Omics-Driven Stress Resistance Strategies
4.1. Modifying HSP Gene Expression in Heat-Stressed Dairy Cows Using CRISPR-Cas Technology
Gene-editing technologies enable the specific management of target traits in livestock by altering the sequence of genes or epigenetic elements [109]. CRISPR-Cas, due to ease of use and simplicity, has been applied to manage traits in various types of biology. For instance, Monochamus alternatus adults with MaltHSP70-2 overexpression via gene editing exhibit enhanced thermotolerance [110]. CRISPRa-mediated upregulation of Heat Shock Proteins (HSPs) confers improved heat resistance in rice [111]. The subsequent section examines CRISPR-Cas editing for multi-locus genetic management in heat-stressed dairy cattle (Figure 2).
Figure 2.
Illustration of strategies for alleviating heat stress in dairy cows. The red × indicates that the improved cattle will not exhibit the heat stress-driven changes shown on the left side.
CRISPR-mediated introduction of Senepol’s slick-hair mutation (PRLR) into dairy cattle has shown improved heat tolerance via hair modification [112], proving genetic editing’s potential. Moreover, GWAS reveals the direct role of HSF1 in mediating HS responses in lactating cows [113]. HSP70 was found to protect the cell by promoting the removal of denatured protein from the protein pool and inhibiting apoptosis [114], as well as enhancing survival of the cell by upregulating expression [115]. The expression of HSPs was confirmed to be controlled by the status of methylation of the genomic DNA. Kisliouk et al. [116] demonstrated that POU2F1 binding activity is regulated by methylation of distal CpG sites in the HSP70 promoter, epigenetically modulating HSP70 expression in response to HS. To regulate HSP70 expression, CRISPRa can be implemented through two approaches: (1) restoring transcriptional activity by targeting CRISPR effector proteins to the HSPA1A promoter via sgRNA; (2) establishing an epigenetic regulatory network by using distal CpG island of the HSPA1A promoter with sgRNA-guided demethylases, reducing methylation to enhance transcription factor binding and sustain HSP70 expression. Additionally, inhibiting epigenetic modifiers (e.g., DNMT3B or HDAC) can suppress their pro-inflammatory effects on NF-κB.
While single-gene editing can distinguish specific phenotypes, the complexity of HS regulatory networks in dairy cattle suggests that multi-gene synergistic regulation may be essential for enhancing thermotolerance. For instance, simultaneous editing of HSP70 (HSPA1A) and superoxide dismutase 2 (SOD2) in bovine mammary cells could synergistically mitigate protein denaturation and oxidative damage. The efficacy of this approach can be assessed by quantifying milk protein synthesis capacity and measuring malondialdehyde reduction via GC-MS. Furthermore, bioreductive liposomes have been employed to deliver Cas9/sgRNA ribonucleoproteins, selectively targeting the critical gene IL1RAP in human leukemia stem cells to reduce leukemic burden via IL1RAP knockout [117]. Similarly, lipid nanoparticles encapsulating Cas9/sgRNA ribonucleoproteins (RNPs) could enable targeted overexpression of thermotolerance-related genes in heat-stressed dairy cattle. Moreover, heat shock elements (HSEs) are special DNA sequences that are bound by HSFs, which trigger the activation of downstream genes under HS conditions [118]. By inserting HSEs into the promoter of the CRISPRa system, target gene expression can be selectively induced under HS, potentially enabling sustained and stable transcriptional regulation.
In summary, integrating GWAS-identified thermotolerance-related genes in dairy cattle with CRISPR/Cas9-mediated gene editing, combined with multi-omics analyses (transcriptomics, proteomics, and metabolomics) of thermal stress responses, establishes a molecular framework for precision breeding. These advanced approaches will significantly accelerate the development of novel heat-tolerant dairy cattle breeds through targeted genetic improvement.
4.2. Modulating Heat Stress-Induced Gastrointestinal Microbiota Shifts via Quorum Sensing Molecules
Gastrointestinal microbiota modulate host physiological responses to HS through intestinal barrier function, microbial metabolites, and immune regulation. For instance, stress-induced proliferation of Clostridium butyricum suppresses butyrate production [21], subsequently activating the NF-κB signaling pathway and triggering inflammatory effects. Shifts in the Firmicutes/Bacteroidetes ratio reduce short-chain fatty acid (SCFA) production [98,99], further disrupting immune cell function [44]. Quorum sensing (QS), a microbial communication system, offers a promising approach to modulate microbial communities. This section explores the potential of QS mechanisms and multi-omics technologies for regulating heat-stressed dairy cow microbiota.
QS influences inter-microbiota gene expression and population behaviors through signaling molecules, thus maintaining microbial homeostasis and modifying host immune functions under stress. For example, the QS molecule N-3-oxo-dodecanoyl-L-homoserine lactone (3O-C12-HSL) interferes with TLR-mediated immune signaling through the inhibition of NF-κB activation and enhancement of Th2 immune responses [119], possibly relieving intestinal pro-inflammatory reactions. Hu et al. [120] demonstrated that AI-2 supplementation elevated butyrate levels in the mouse intestine and improved gut barrier integrity. More recently, investigations into the application of a quorum sensing modulator, 4-hydroxy-2,5-dimethyl-3(2H)-furanone (HDMF), in Hu sheep have revealed that the incorporation of HDMF enhanced serum antioxidant capacity and mitigated oxidative stress [121]. These beneficial effects appear to be linked to the AI-2 signaling molecule and the formation of biofilms. Additionally, Liu et al. [122] demonstrated that elevating the temperature of drinking water during cold winter can significantly boost the growth performance, serum immune function, and antioxidant capabilities of Hu sheep, with an accompanying increase in microbial protein production within the rumen. These improvements are also found to be positively correlated with microbial density, AI-2 signaling molecule, and biofilm formation. Therefore, AI-2 signaling molecule may mitigate HS-induced intestinal dysfunction by modulating probiotic metabolism, serum antioxidant capability, and rumen fermentation. This implies that either 3O-C12-HSL or AI-2 analogs would be potential candidates to overcome immune dysregulation and energy imbalance in heat-stressed dairy cows. However, considering the complexity and wide-ranging cross-species signaling effects of QS networks, such approaches could also probably activate undesired microbes, raising the risk of increased infection susceptibility pathway [123]. The following may enhance that specificity in the future: (1) using peptide or amino acid polymers to copy natural QS signals [124]; (2) liposome or microencapsulated slow-release QS molecule packaging [125]. Those delivery forms would be effective only at specific pH levels in the gut or when particular bacterial enzymes are present, thus limiting their distribution throughout the body. Additionally, multi-omics integration could dynamically track QS molecule expression, metabolite flux, and their interactions with host factors, elucidating the QS-mediated gut–immune axis.
4.3. Intelligent Breeding and Digitalized Management
Traditional breeding selects animals based on visible traits with limitations like low precision in finding genes, slow progress, and difficulty improving multiple traits together [126]. Environmental factors (like temperature and humidity) affect trait measurements, while interactions between many genes control complex features (e.g., heat tolerance). Additionally, bad genes can persist due to genetic linkages, reducing breeding efficiency. While precision breeding using multi-omics data is feasible, it generates vast amounts of phenotypic data, posing challenges for storage and analysis. Therefore, this section explores how integrating multi-omics with IoT and AI can enable smart breeding and digitalized management.
Previous research has identified seven novel dairy cattle HS-associated genes by employing RT, RS, and DS methods such as WssGWAS and RNA-Seq [25]. By utilizing IoT sensors, WssGWAS, whole-genome sequencing to identify regulatory elements of candidate genes, along with isotope labeling, GC-MS technology, and KEGG pathway analysis to find key metabolites and related signaling pathways, this gene–metabolome research framework has been used to investigate the hierarchical effect of genetic variations on metabolite levels [127]. Meanwhile, omic and non-omic effects can be integrated into individual breeding value estimation to identify candidate genes [128], enabling the effective combination of multi-omics data with breeding programs. Furthermore, pre-trained multimodal models using omics data can reconstruct interaction networks among cells, genes, and tissues [129]. Wu et al. [130] have successfully applied integrated omics and artificial intelligence for multi-scale predictive modeling of genotype-environment-phenotype relationships. In addition, by monitoring phenotypic variations in dairy cattle over various environments via IoT and combining omics data, deep learning is able to be used to establish a digital twin model for the cattle under HS to predict regulatory loci. A digital twin model serves as a virtual representation that transforms environmental and physiological changes into digital inputs for computational analysis [131]. This model enables the prediction of physiological responses through the regulation of specific target sites. While previous studies have successfully developed behavioral digital twin models for healthy cattle [132], these models have not yet incorporated multi-omics data, highlighting the untapped potential and feasibility of the proposed approach. Thus, leveraging extensive datasets, including phenotypic data, associated genes, metabolic data, and pathway analysis, AI can be employed to screen and analyze these datasets. This would allow the construction of a novel multi-gene weighted scoring model for heat tolerance traits, facilitating digitalized management and intelligent breeding strategies.
5. Conclusions and Future Perspectives
Under HS, dairy cattle exhibit reduced dry matter intake, endocrine disruption, and immune dysfunction, ultimately decreasing milk yield, quality, and reproductive performance. We analyze key regulatory networks, focusing on thermosensitive ion channels and ROS in HS responses. We discuss interactions among MAPK, AMPK, and NF-κB signaling pathways, particularly the relationship between MAPK and Heat Shock Proteins (HSPs). Furthermore, using multi-omics approaches, we explore potential strategies for developing innovative breeding models to enhance heat tolerance.
Heat tolerance development involves complex gene–environment interactions, requiring multidimensional technologies for comprehensive analysis. By integrating multi-omics data, environmental parameters, and AI, we can systematically decode and target HS response mechanisms in dairy cattle. While the integration of multi-omics, IoT, and AI technologies is theoretically promising, practical implementation faces significant challenges. The primary obstacle lies in integrating multi-source data, particularly addressing the spatiotemporal mismatches between GWAS population data, multi-omics datasets, and continuous IoT-generated environmental monitoring data. This requires developing novel algorithms. Furthermore, current causal inference methods limit our mechanistic understanding. Although machine learning approaches can model complex gene–environment interactions, their “black-box” nature obscures causal pathway identification and complicates the assessment of confounding factors, such as gut microbiome-mediated metabolic interference. Simultaneously, the environmental intelligence and precision breeding can boost agricultural productivity and climate resilience, poor farmers still face major infrastructural, economic, and institutional barriers. Other challenges include (1) digital and infrastructure gaps. For example, less than 20% of African smallholders have stable mobile internet access [133]; (2) genotyping, AI tools, and laboratory infrastructure-related high costs; and (3) weak institutional support. Public–private partnerships remain thin, funding is erratic, and seed certification processes are slow. Additionally, ethical and industrial challenges also arise. Strict regulation of gene-edited farm animals, highlighted by the Regulation Tracker website [134], has slowed breeding progress, while phenotype differences between lab and farm conditions hinder practical application of research findings.
In conclusion, the integration of multi-omics, IoT, and AI technologies with CRISPR-Cas-mediated genome editing offers a transformative approach to systematically decode and enhance heat tolerance in dairy cattle. While challenges persist, including data integration complexities, editing precision, and ethical considerations, this innovative framework demonstrates significant scientific promise and practical applicability for advancing livestock thermotolerance.
Author Contributions
Conceptualization, Z.X. and Q.Q.; methodology, L.L., K.O. and M.Q.; software, Z.X.; validation, Z.X. and L.L.; resources, Q.Q.; data curation, Z.X., L.L. and Q.Q.; writing—original draft preparation, Z.X.; writing—review and editing, Q.Q. and L.L.; visualization, Z.X.; supervision, Q.Q.; project administration, Q.Q.; funding acquisition, Q.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 32260861; the Major Discipline Academic and Technical Leaders Training Program of Jiangxi Province, grant number 20243BCE51165; and the Jiangxi Provincial Natural Science Foundation, grant number 20232BAB215051.
Acknowledgments
During the preparation of this review, we used Adobe Illustrator 2021 and the following open-source websites: https://www.iconfont.cn/ (accessed on 1 March 2025), https://smart.servier.com/ (accessed on 1 March 2025), and https://commons.wikimedia.org/wiki/Main_Page (accessed on 1 March 2025) for the purposes of making illustrations.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| ACTH | Adrenocorticotropic Hormone |
| AI | Artificial Intelligence |
| A1 | Annexin 1 |
| ATAC-seq | Assay for Transposase-Accessible Chromatin using sequencing |
| BHB | β-hydroxybutyrate |
| BLG | β-lactoglobulin |
| CaMKKβ | Ca2+/Calmodulin-Dependent Kinase Kinase β |
| CD21+ | Cluster of Differentiation 21 positive |
| CRH | Corticotropin-Releasing Hormone |
| ChIP-seq | Chromatin ImmunoPrecipitation sequencing |
| CRISPR-Cas9 | Clustered Regularly Interspaced Short Palindromic Repeats-CRISPR-associated protein 9 |
| CYP19A1 | Cytochrome P450 Family 19 Subfamily A Member 1 |
| Cytc | Cytochrome c |
| DIA | Data-Independent Acquisition |
| DMI | Dry Matter Intake |
| DNA | Deoxyribonucleic Acid |
| DNMT3B | DNA (Cytosine-5)-Methyltransferase 3 Beta |
| DS | Drooling Scores |
| EI-MN-PB | Environmental Intelligence–Molecular Network–Precision Breeding |
| FSH | Follicle Stimulating Hormone |
| FOXO3a | Forkhead Box O3a |
| GC-MS | Gas Chromatography–Mass Spectrometry |
| GO | Gene Ontology |
| GLUT4 | Glucose Transporter Type 4 |
| GPx | Glutathione Peroxidase |
| GnRH | Gonadotropin-Releasing Hormone |
| GWAS | Genome-Wide Association Study |
| gRNA | Guide RNA |
| HPA | Hypothalamo–Pituitary–Adrenal |
| HPO | Hypothalamic–Pituitary–Ovarian |
| HDAC | Histone Deacetylase |
| HSEs | Heat Shock Elements |
| HSF1 | Heat Shock Factor 1 |
| HSP | Heat Shock Protein |
| HSP27 | Heat Shock Protein 27 |
| HSP70 | Heat Shock Protein 70 |
| HSPA1A | Heat Shock Protein Family A (Hsp70) Member 1A |
| HS | Heat Stress |
| IKK | IκB Kinase |
| IL-6 | Interleukin-6 |
| IMS | Ion Mobility Spectrometry |
| IoT | Internet of Things |
| IP3 | Inositol Trisphosphate |
| IκBα | Inhibitor of Nuclear Factor Kappa B Alpha |
| JNK | c-Jun N-terminal Kinase |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LH | Luteinizing Hormone |
| LPS | Lipopolysaccharides |
| MAPK | Mitogen-Activated Protein Kinase |
| MCU | Mitochondrial Calcium Un |
| MDA | Malondialdehyde |
| MKP-1 | MAPK Phosphatases-1 |
| MnSOD | Manganese Superoxide Dismutase |
| MS | Mass Spectrometry |
| MyD88 | Myeloid differentiation primary response 88 |
| NADPH | Nicotinamide Adenine Dinucleotide Phosphate |
| NEB | Negative Energy Balance |
| NGS | Next-Generation Sequencing |
| NMR | Nuclear Magnetic Resonance |
| NOX | NADPH Oxidase |
| Npc1/1 | Niemann-Pick Disease, Type C1 |
| NF-κB | Nuclear Factor-Kappa B |
| QS | Quorum Sensing |
| P38 | Mitogen-Activated Protein Kinase p38 |
| PGC-1α | Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-Alpha |
| ROS | Reactive Oxygen Species |
| RNA | Ribonucleic Acid |
| RNA-seq | RNA Sequencing |
| RR | Respiratory Rate |
| RT | Rectal Temperature |
| SCFAs | Short-Chain Fatty Acids |
| scRNA-seq | Single-Cell RNA Sequencing |
| SOD | Superoxide Dismutase |
| STAT6 | Signal Transducer and Activator of Transcription 6 |
| STEM | Short Time-series Expression Miner |
| TCA | Tricarboxylic Acid Cycle |
| THI | Temperature Humidity Index |
| Th | T Helper Cell |
| THZ | Thermal Hazard Zone |
| TLR4 | Toll-Like Receptor 4 |
| TNF-α | Tumor Necrosis Factor-Alpha |
| TRP | Transient Receptor Potential |
| TRPM | Transient Receptor Potential Melastatin |
| TRPV | Transient Receptor Potential Vanilloid |
| WES | Whole Exome Sequencing |
| WGS | Whole Genome Sequencing |
| WssGWAS | Weighted Single-Step Genome-Wide Association Study |
References
- Rounce, D.R.; Hock, R.; Maussion, F.; Hugonnet, R.; Kochtitzky, W.; Huss, M.; Berthier, E.; Brinkerhoff, D.; Compagno, L.; Copland, L.; et al. Global glacier change in the 21st century: Every increase in temperature matters. Science 2023, 379, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.A.; Collier, R.J.; Stone, A.E. Invited review: Physiological and behavioral effects of heat stress in dairy cows. J. Dairy Sci. 2020, 103, 6751–6770. [Google Scholar] [CrossRef] [PubMed]
- Slayi, M.; Zhou, L.; Jaja, I.F. Strategies, challenges, and outcomes of heat stress resilience in sub-saharan african community-based cattle feedlots: A systematic review. Front. Vet. Sci. 2024, 11, 1455917. [Google Scholar] [CrossRef] [PubMed]
- West, J.W. Nutritional strategies for managing the heat-stressed dairy cow. J. Dairy Sci. 1997, 77, 21. [Google Scholar] [CrossRef]
- Wheelock, J.B.; Rhoads, R.P.; VanBaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef]
- Bagath, M.; Krishnan, G.; Devaraj, C.; Rashamol, V.P.; Pragna, P.; Lees, A.M.; Sejian, V. The impact of heat stress on the immune system in dairy cattle: A review. Res. Vet. Sci. 2019, 126, 94–102. [Google Scholar] [CrossRef]
- Dovolou, E.; Giannoulis, T.; Nanas, I.; Amiridis, G.S. Heat stress: A serious disruptor of the reproductive physiology of dairy cows. Animals 2023, 13, 1846. [Google Scholar] [CrossRef]
- Guo, Y.; Peng, Q.; Tao, J. Blood metabolomic studies of heat stress cow with GC-MS. Acta Vet. Zootech. Sin. 2015, 46, 1356–1362. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, H.; Luo, H.; Mi, S.; Liu, W.; Chu, Q.; Wang, Y. Knowledge map ping analysis on heat stress genetic mechanisms research of dairy cattle. Acta Vet. Zootech. Sin. 2021, 52, 1141–1153. [Google Scholar] [CrossRef]
- Min, L.; Zhao, S.; Tian, H.; Zhou, X.; Zhang, Y.; Li, S.; Yang, H.; Zheng, N.; Wang, J. Metabolic responses and “omics” technologies for elucidating the effects of heat stress in dairy cows. Int. J. Biometeorol. 2016, 61, 1149–1158. [Google Scholar] [CrossRef]
- Magana, J.; Gavojdian, D.; Menachem, Y.; Lazebnik, T.; Zamansky, A.; Adams-Progar, A. Machine learning approaches to predict and detect early-onset of digital dermatitis in dairy cows using sensor data. Front. Vet. Sci. 2023, 10, 1295430. [Google Scholar] [CrossRef] [PubMed]
- Worku, D.; Hussen, J.; De Matteis, G.; Schusser, B.; Alhussien, M.N. Candidate genes associated with heat stress and breeding strategies to relieve its effects in dairy cattle: A deeper insight into the genetic architecture and immune response to heat stress. Front. Vet. Sci. 2023, 10, 1151241. [Google Scholar] [CrossRef] [PubMed]
- Nuffield Council on Bioethics. Genome editing—An ethical review. Jahrb. Wiss. Eth. 2017, 22, 293–340. [Google Scholar] [CrossRef]
- Vautard, R.; Barnes, C.R.; Philip, S.; Kew, S.; Pinto, I.; Otto, F.E.L. Heat extremes linearly shift with global warming, with frequency doubling per decade since 1979. Environ. Res. Lett. 2024, 19, 094033. [Google Scholar] [CrossRef]
- Oliveira, C.P.; Sousa, F.C.d.; da Silva, A.L.; Schultz, É.B.; Valderrama Londoño, R.I.; de Souza, P.A.R. Heat stress in dairy cows: Impacts, identification, and mitigation strategies—A review. Animals 2025, 15, 249. [Google Scholar] [CrossRef]
- Țogoe, D.; Mincă, N.A. The impact of heat stress on the physiological, productive, and reproductive status of dairy cows. Agriculture 2024, 14, 1241. [Google Scholar] [CrossRef]
- Hao, L.Y.; Wang, J.; Sun, P.; Bu, D.P. The effect of heat stress on the metabolism of dairy cows: Updates & review. Austin J. Nutr. Metab. 2016, 3, 1036. [Google Scholar]
- Chen, X.; Li, C.; Fang, T.; Yao, J.; Gu, X. Effects of heat stress on endocrine, thermoregulatory, and lactation capacity in heat-tolerant and -sensitive dry cows. Front. Vet. Sci. 2024, 11, 1405263. [Google Scholar] [CrossRef]
- Sguizzato, A.L.L.; Marcondes, M.I.; Dijkstra, J.; Valadares Filho, S.d.C.; Campos, M.M.; Machado, F.S.; Silva, B.C.; Rotta, P.P. Energy requirements for pregnant dairy cows. PLoS ONE 2020, 15, e0235619. [Google Scholar] [CrossRef]
- West, J.W. Effects of heat-stress on production in dairy cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Gao, J.; Qi, Z. Effects of heat stress on rumen microbiota and its relationship with performance of dairy cows. Chin. J. Anim. Nutr. 2019, 31, 4458–4463. [Google Scholar]
- Marquez-Acevedo, A.S.; Hood, W.; Collier, R.; Skibiel, A. Mitochondrial response to heat stress and its implications on dairy cattle bioenergetics, metabolism, and production. J. Dairy Sci. 2023, 106, 7295–7309. [Google Scholar] [CrossRef] [PubMed]
- Bharati, J.; Dangi, S.S.; Chouhan, V.S.; Mishra, S.R.; Bharti, M.K.; Verma, V.; Shankar, O.; Yadav, V.P.; Das, K.; Paul, A.; et al. Expression dynamics of HSP70 during chronic heat stress in Tharparkar cattle. Int. J. Biometeorol. 2017, 61, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Roufayel, R.; Johnston, D.S.; Mosser, D.D. The elimination of miR-23a in heat-stressed cells promotes NOXA-Induced cell death and is prevented by HSP70. Cell Death Dis. 2014, 5, e1546. [Google Scholar] [CrossRef]
- Luo, H.; Hu, L.; Brito, L.F.; Dou, J.; Sammad, A.; Chang, Y.; Ma, L.; Guo, G.; Liu, L.; Zhai, L.; et al. Weighted single-step GWAS and RNA sequencing reveals key candidate genes associated with physiological indicators of heat stress in Holstein cattle. J. Anim. Sci. Biotechnol. 2022, 13, 108. [Google Scholar] [CrossRef]
- Joo, S.; Lee, S.J.; Park, D.S.; Kim, D.H.; Gu, B.; Park, Y.; Rim, C.; Kim, M.; Kim, E. Changes in blood metabolites and immune cells in Holstein and Jersey dairy cows by heat stress. Animals 2021, 11, 974. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Li, R.; Wu, Y.; Zhang, D.; Xu, H.; Zhang, Y.; Qi, Z. Effect of Seasonal Thermal stress on oxidative status, immune response and stress hormones of lactating dairy cows. Anim. Nutr. 2021, 7, 216–223. [Google Scholar] [CrossRef]
- Mishra, S.R. Behavioural, physiological, neuro-endocrine and molecular responses of cattle against heat stress: An updated review. Trop. Anim. Health Prod. 2021, 53, 400. [Google Scholar] [CrossRef]
- Catozzi, C.; Ávila, G.; Zamarian, V.; Pravettoni, D.; Sala, G.; Ceciliani, F.; Lacetera, N.; Lecchi, C. In-vitro effect of heat stress on bovine monocytes lifespan and polarization. Immunobiology 2020, 225, 151888. [Google Scholar] [CrossRef]
- Feher, J. The Adrenal Cortex; Feher, J., Ed.; Academic Press: Boston, MA, USA, 2012; pp. 810–819. [Google Scholar]
- Cavalcanti, D.M.H.; Lotufo, C.M.C.; Borelli, P.; Ferreira, Z.S.; Markus, R.P.; Farsky, S.H.P. Endogenous glucocorticoids control neutrophil mobilization from bone marrow to blood and tissues in non-inflammatory conditions. Br. J. Pharmacol. 2007, 152, 1291–1300. [Google Scholar] [CrossRef]
- Tejaswi, V.; Balachander, B.; Samad, H.A.; Sarkar, M.; Maurya, V.P.; Singh, G. Assessment of heat stress induced alterations in polymorphonuclear (PMN) cell activity in native and crossbred cows. J. Appl. Anim. Res. 2020, 48, 549–552. [Google Scholar] [CrossRef]
- Ricci, E.; Ronchetti, S.; Gabrielli, E.; Pericolini, E.; Gentili, M.; Roselletti, E.; Vecchiarelli, A.; Riccardi, C. GILZ restrains neutrophil activation by inhibiting the MAPK pathway. J. Leukoc. Biol. 2019, 105, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Maciuszek, M.; Klak, K.; Rydz, L.; Verburg-van Kemenade, B.M.L.; Chadzinska, M. Cortisol metabolism in carp macrophages: A role for macrophage-derived cortisol in M1/M2 polarization. Int. J. Mol. Sci. 2020, 21, 8954. [Google Scholar] [CrossRef] [PubMed]
- Lacetera, N.; Bernabucci, U.; Scalia, D.; Ronchi, B.; Kuzminsky, G.; Nardone, A. Lymphocyte functions in dairy cows in hot environment. Int. J. Biometeorol. 2005, 50, 105–110. [Google Scholar] [CrossRef]
- Park, D.S.; Gu, B.-H.; Park, Y.J.; Joo, S.S.; Lee, S.-S.; Kim, S.-H.; Kim, E.T.; Kim, D.H.; Lee, S.S.; Lee, S.J.; et al. Dynamic changes in blood immune cell composition and function in Holstein and jersey steers in response to heat stress. Cell Stress Chaperon. 2021, 26, 705–720. [Google Scholar] [CrossRef]
- Murata, H.; Kunii, H.; Kusama, K.; Sakurai, T.; Bai, H.; Kawahara, M.; Takahashi, M. Heat stress induces oxidative stress and activates the KEAP1-NFE2L2-ARE pathway in bovine endometrial epithelial cells. Biol. Reprod. 2021, 105, 1114–1125. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, H.; Zhao, M.; Chang, C.; Lu, Q. The Bach family of transcription factors: A comprehensive review. Clin. Rev. Allergy Immunol. 2016, 50, 345–356. [Google Scholar] [CrossRef]
- Marin, D.; Fernandez, G.J.; Hernandez, J.C.; Taborda, N. A systems biology approach unveils different gene expression control mechanisms governing the immune response genetic program in peripheral blood mononuclear cells exposed to SARS-CoV-2. PLoS ONE 2024, 19, e0314754. [Google Scholar] [CrossRef]
- Ceciliani, F.; Ceron, J.J.; Eckersall, P.D.; Sauerwein, H. Acute phase proteins in ruminants. J. Proteom. 2012, 75, 4207–4231. [Google Scholar] [CrossRef]
- Tian, H.; Wang, W.; Zheng, N.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Identification of diagnostic biomarkers and metabolic pathway shifts of heat-stressed lactating dairy cows. J. Proteom. 2015, 125, 17–28. [Google Scholar] [CrossRef]
- Koch, F.; Thom, U.; Albrecht, E.; Weikard, R.; Nolte, W.; Kuhla, B.; Kuehn, C. Heat stress directly impairs gut integrity and recruits distinct immune cell populations into the bovine intestine. Proc. Natl. Acad. Sci. USA 2019, 116, 10333–10338. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.R.; Ferguson, G.J.; Kulkarni, S.; Damoulakis, G.; Anderson, K.; Bohlooly-Y, M.; Stephens, L.; Hawkins, P.T.; Curi, R. SCFAs induce mouse neutrophil chemotaxis through the GPR43 receptor. PLoS ONE 2011, 6, e21205. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-Chain Fatty Acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Karl, J.P.; Margolis, L.M.; Madslien, E.H.; Murphy, N.E.; Castellani, J.W.; Gundersen, Y.; Hoke, A.V.; Levangie, M.W.; Kumar, R.; Chakraborty, N.; et al. Changes in intestinal microbiota composition and metabolism coincide with increased intestinal permeability in young adults under prolonged physiological stress. Am. J. Physiol. Liver Physiol. 2017, 4, G559–G571. [Google Scholar] [CrossRef]
- Min, L.; Cheng, J.; Shi, B.; Yang, H.; Zheng, N.; Wang, J. Effects of heat stress on serum insulin, adipokines, AMP-activated protein kinase, and heat shock signal molecules in dairy cows. J. Zhejiang Univ. Sci. B 2015, 16, 541–548. [Google Scholar] [CrossRef]
- Collier, R.J.; Collier, J.L.; Rhoads, R.P.; Baumgard, L.H. Invited review: Genes involved in the bovine heat stress response. J. Dairy Sci. 2008, 91, 445–454. [Google Scholar] [CrossRef]
- Starkie, R.L.; Hargreaves, M.; Rolland, J.; Febbraio, M.A. Heat stress, cytokines, and the immune response to exercise. Brain Behav. Immun. 2005, 19, 404–412. [Google Scholar] [CrossRef]
- McFadden, J.W.; Rico, J.E. Invited review: Sphingolipid biology in the dairy cow: The emerging role of ceramide. J. Dairy Sci. 2019, 102, 7619–7639. [Google Scholar] [CrossRef]
- Rhoads, M.; Rhoads, R.; Vanbaale, M.; Collier, R.; Sanders, S.; Weber, W.; Crooker, B.; Baumgard, L. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef]
- Abbas, Z.; Sammad, A.; Hu, L.; Fang, H.; Xu, Q.; Wang, Y. Glucose metabolism and dynamics of facilitative glucose transporters (GLUTs) under the influence of heat stress in dairy cattle. Metabolites 2020, 10, 312. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Bruce Currie, W. Partitioning of nutrients during pregnancy and lactation: A review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci. 1980, 63, 1514–1529. [Google Scholar] [CrossRef] [PubMed]
- Blond, B.; Majkić, M.; Spasojević, J.; Hristov, S.; Radinović, M.; Nikolić, S.; Anđušić, L.; Čukić, A.; Došenović Marinković, M.; Vujanović, B.D.; et al. Influence of heat stress on body surface temperature and blood metabolic, endocrine, and inflammatory parameters and their correlation in cows. Metabolites 2024, 14, 104. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, M.Z.; Umer, S.; Khan, I.M.; Xu, H.; Zhu, H.; Wang, Y. Cellular and molecular adaptation of bovine granulosa cells and oocytes under heat stress. Animals 2020, 10, 110. [Google Scholar] [CrossRef]
- Oakley, A.E.; Breen, K.M.; Clarke, I.J.; Karsch, F.J.; Wagenmaker, E.R.; Tilbrook, A.J. Cortisol reduces gonadotropin-releasing hormone pulse frequency in follicular phase ewes: Influence of ovarian steroids. Endocrinology 2009, 150, 341–349. [Google Scholar] [CrossRef]
- Gauthier, D. The influence of season and shade on oestrous behaviour, timing of preovulatory LH surge and the pattern of progesterone secretion in FFPN and creole heifers in a tropical climate. Reprod. Nutr. Dev. 1986, 26, 767–775. [Google Scholar] [CrossRef]
- Roth, Z. Influence of heat stress on reproduction in dairy cows—Physiological and practical aspects. J. Anim. Sci. 2020, 98, S80–S87. [Google Scholar] [CrossRef]
- Armstrong, D.T.; Dorrington, J.H. Estrogen biosynthesis in the ovaries and testes. Adv. Sex Horm. Res. 1977, 3, 217–258. [Google Scholar]
- Guzeloglu, A.; Ambrose, J.D.; Kassa, T.; Diaz, T.; Thatcher, M.J.; Thatcher, W.W. Long-term follicular dynamics and biochemical characteristics of dominant follicles in dairy cows subjected to acute heat stress. Anim. Reprod. Sci. 2001, 66, 15–34. [Google Scholar] [CrossRef]
- Wolfenson, D.; Roth, Z. Impact of heat stress on cow reproduction and fertility. Anim. Front. 2018, 9, 32–38. [Google Scholar] [CrossRef]
- Turk, R.; Rošić, N.; Vince, S.; Perkov, S.; Samardžija, M.; Beer-Ljubić, B.; Belić, M.; Robić, M. The influence of heat stress on energy metabolism in simmental dairy cows during the periparturient period. Vet. Arh. 2020, 90, 1–10. [Google Scholar] [CrossRef]
- Kilany, A.A.; El-Darawany, A.-H.A.; El-Tarabany, A.A.; Al-Marakby, K.M. Effect of folic acid supplements on progesterone profile and blood metabolites of heat-stressed Holstein cows during the early stage of pregnancy. Animals 2022, 12, 1872. [Google Scholar] [CrossRef] [PubMed]
- Schepers, R.J.; Ringkamp, M. Thermoreceptors and thermosensitive afferents. Neurosci. Biobehav. Rev. 2010, 34, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Subcellular Biochemistry in Transient Receptor Potential (TRP) Channels; Harris, J.R., Boekema, E.J., Eds.; Springer: Singapore, 2018; pp. 141–165. [Google Scholar]
- Himmel, N.J.; Cox, D.N. Transient receptor potential channels: Current perspectives on evolution, structure, function and nomenclature. Proc. R. Soc. B 2020, 287, 20201309. [Google Scholar] [CrossRef]
- Saito, S.; Fukuta, N.; Shingai, R.; Tominaga, M. Evolution of vertebrate transient receptor potential vanilloid 3 channels: Opposite temperature sensitivity between mammals and western clawed frogs. PLoS Genet. 2011, 7, e1002041. [Google Scholar] [CrossRef]
- Güler, A.D.; Lee, H.; Iida, T.; Shimizu, I.; Tominaga, M.; Caterina, M. Heat-evoked activation of the ion channel, TRPV4. J. Neurosci. 2002, 22, 6408–6414. [Google Scholar] [CrossRef]
- Dikmen, S.; Hansen, P.J. Is the temperature-humidity index the best indicator of heat stress in lactating cows in a subtropical environment? J. Dairy Sci. 2009, 92, 109–116. [Google Scholar] [CrossRef]
- Peier, A.; Reeve, A.; Andersson, D.; Moqrich, A.; Earley, T.J.; Hergarden, A.C.; Story, G.M.; Colley, S.; Hogenesch, J.; McIntyre, P.; et al. A heat-sensitive TRP channel expressed in keratinocytes. Science 2002, 296, 2046–2049. [Google Scholar] [CrossRef]
- Hsu, W.-L.; Yoshioka, T. Role of TRP channels in the induction of heat shock proteins (HSPs) by heating skin. Biophysics 2015, 11, 25–32. [Google Scholar] [CrossRef]
- Lee, S.H.; Duron, H.E.; Chaudhuri, D. Beyond the TCA cycle: New insights into mitochondrial calcium regulation of oxidative phosphorylation. Biochem. Soc. Trans. 2023, 51, 1661–1673. [Google Scholar] [CrossRef]
- Walkon, L.L.; Strubbe-Rivera, J.O.; Bazil, J. Calcium overload and mitochondrial metabolism. Biomolecules 2022, 12, 1891. [Google Scholar] [CrossRef] [PubMed]
- Mujahid, A.; Pumford, N.R.; Bottje, W.; Nakagawa, K.; Miyazawa, T.; Akiba, Y.; Toyomizu, M. Mitochondrial oxidative damage in chicken skeletal muscle induced by acute heat stress. J. Poult. Sci. 2007, 44, 439–445. [Google Scholar] [CrossRef]
- Rhoads, D.M.; Umbach, A.L.; Subbaiah, C.C.; Siedow, J.N. Mitochondrial reactive oxygen species. contribution to oxidative stress and interorganellar signaling. Plant Physiol. 2006, 141, 357–366. [Google Scholar] [CrossRef]
- Zhao, Q.; Fujiwara, Y.; Kondo, T. Mechanism of cell death induction by nitroxide and hyperthermia. Free Radic. Biol. Med. 2006, 40, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Lemasters, J.J. Regulated and unregulated mitochondrial permeability transition pores: A new paradigm of pore structure and function? FEBS Lett. 2002, 512, 1–7. [Google Scholar] [CrossRef]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef]
- Green, D.R.; Kroemer, G. The pathophysiology of mitochondrial cell death. Science 2004, 305, 626–629. [Google Scholar] [CrossRef]
- Vais, H.; Payne, R.; Paudel, U.; Li, C.; Foskett, J.K. Coupled transmembrane mechanisms control MCU-mediated mitochondrial Ca2+ uptake. Proc. Natl. Acad. Sci. USA 2020, 117, 21731–21739. [Google Scholar] [CrossRef]
- Kumar, A.; Majhi, R.; Acharya, T.; Smalla, K.; Gundelfinger, E.; Goswami, C. TRPV4 interacts with mitochondrial proteins and acts as a mitochondrial structure-function regulator. bioRxiv 2018, 330993. [Google Scholar] [CrossRef]
- Liu, D.; Yang, Y.; Chen, Z.; Fan, Y.; Liu, J.; Xu, Y.; Ahmed, Z.; Zhang, J.; Li, F.; Qi, X.; et al. Temperature adaptation patterns in chinese cattle revealed by TRPM2 gene mutation analysis. Anim. Biotechnol. 2024, 35, 2299944. [Google Scholar] [CrossRef]
- Kim, H.S.; Ullevig, S.L.; Zamora, D.; Lee, C.F.; Asmis, R. Redox Regulation of MAPK phosphatase 1 controls monocyte migration and macrophage recruitment. Proc. Natl. Acad. Sci. USA 2012, 109, E2803–E2812. [Google Scholar] [CrossRef] [PubMed]
- Maik-Rachline, G.; Lifshits, L.; Seger, R. Nuclear p38: Roles in physiological and pathological processes and regulation of nuclear translocation. Int. J. Mol. Sci. 2020, 21, 6102. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Shi, L.Z.; Chi, H. Regulation of JNK and P38 MAPK in the immune system: Signal integration, propagation and termination. Cytokine 2009, 48, 161–169. [Google Scholar] [CrossRef]
- D’Angelo, D.; Vecellio Reane, D.; Raffaello, A. Neither too much nor too little: Mitochondrial calcium concentration as a balance between physiological and pathological conditions. Front. Mol. Biosci. 2023, 10, 1336416. [Google Scholar] [CrossRef] [PubMed]
- Moon, E.J.; Sonveaux, P.; Porporato, P.E.; Danhier, P.; Gallez, B.; Batinic-Haberle, I.; Nien, Y.-C.; Schroeder, T.; Dewhirst, M.W. NADPH oxidase-mediated reactive oxygen species production activates hypoxia-inducible factor-1 (HIF-1) via the ERK pathway after hyperthermia treatment. Proc. Natl. Acad. Sci. USA 2010, 107, 20477–20482. [Google Scholar] [CrossRef]
- Segal, A.W.; Abo, A. The biochemical basis of the NADPH oxidase of phagocytes. Trends Biochem. Sci. 1993, 18, 43–47. [Google Scholar] [CrossRef]
- Evans, M.J.; Choi, W.-G.; Gilroy, S.; Morris, R.J. A ROS-assisted calcium wave dependent on the AtRBOHD NADPH oxidase and TPC1 cation channel propagates the systemic response to salt stress. Plant Physiol. 2016, 171, 1771–1784. [Google Scholar] [CrossRef]
- Tokumitsu, H.; Sakagami, H. Molecular mechanisms underlying Ca2+/calmodulin-dependent protein kinase kinase signal transduction. Int. J. Mol. Sci. 2022, 23, 11025. [Google Scholar] [CrossRef]
- Jäger, S.; Handschin, C.; St-Pierre, J.; Spiegelman, B.M. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1α. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar] [CrossRef]
- Kanarek, N.; Ben-Neriah, Y. Regulation of NF-κB by ubiquitination and degradation of the IκBs. Immunol. Rev. 2012, 246, 77–94. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Alberti, G.; Paladino, L.; Vitale, A.M.; Caruso Bavisotto, C.; Conway de Macario, E.; Campanella, C.; Macario, A.J.L.; Marino Gammazza, A. Functions and therapeutic potential of extracellular Hsp60, Hsp70, and Hsp90 in neuroinflammatory disorders. Appl. Sci. 2021, 11, 736. [Google Scholar] [CrossRef]
- Lazaro, I.; Oguiza, A.; Recio, C.; Mallavia, B.; Madrigal-Matute, J.; Blanco, J.; Egido, J.; Martin-Ventura, J.-L.; Gomez-Guerrero, C. Targeting HSP90 ameliorates nephropathy and atherosclerosis through suppression of NF-κB and STAT signaling pathways in diabetic mice. Diabetes 2015, 64, 3600–3613. [Google Scholar] [CrossRef] [PubMed]
- Nyati, K.K.; Masuda, K.; Zaman, M.M.-U.; Dubey, P.K.; Millrine, D.; Chalise, J.P.; Higa, M.; Li, S.; Standley, D.M.; Saito, K.; et al. TLR4-induced NF-κB and MAPK signaling regulate the IL-6 mRNA stabilizing protein Arid5a. Nucleic Acids Res. 2017, 45, 2687–2703. [Google Scholar] [CrossRef]
- Andreakos, E.; Sacre, S.M.; Smith, C.; Lundberg, A.; Kiriakidis, S.; Stonehouse, T.; Monaco, C.; Feldmann, M.; Foxwell, B.M. Distinct pathways of LPS-induced NF-κB activation and cytokine production in human myeloid and nonmyeloid cells defined by selective utilization of MyD88 and Mal/TIRAP. Blood 2004, 103, 2229–2237. [Google Scholar] [CrossRef]
- Patra, A.K.; Kar, I. Heat stress on microbiota composition, barrier integrity, and nutrient transport in gut, production performance, and its amelioration in farm animals. J. Anim. Sci. Technol. 2021, 63, 211–247. [Google Scholar] [CrossRef]
- Wen, C.; Li, S.; Wang, J.; Zhu, Y.; Zong, X.; Wang, Y.; Jin, M. Heat stress alters the intestinal microbiota and metabolomic profiles in mice. Front. Microbiol. 2021, 12, 706772. [Google Scholar] [CrossRef]
- Bridgeman, S.; Ellison, G.; Newsholme, P.; Mamotte, C. The HDAC inhibitor butyrate impairs β cell function and activates the disallowed gene hexokinase I. Int. J. Mol. Sci. 2021, 22, 13330. [Google Scholar] [CrossRef]
- Rahman, M.M.; Kukita, A.; Kukita, T.; Shobuike, T.; Nakamura, T.; Kohashi, O. Two histone deacetylase inhibitors, trichostatin a and sodium butyrate, suppress differentiation into osteoclasts but not into macrophages. Blood 2003, 101, 3451–3459. [Google Scholar] [CrossRef]
- Zhou, R.; Gong, A.-Y.; Chen, D.; Miller, R.E.; Eischeid, A.N.; Chen, X.-M. Histone deacetylases and NF-kB signaling coordinate expression of CX3CL1 in epithelial cells in response to microbial challenge by suppressing miR-424 and miR-503. PLoS ONE 2013, 8, e65153. [Google Scholar] [CrossRef]
- Bosch, R.; Philips, N.; Suárez-Pérez, J.A.; Juarranz, A.; Devmurari, A.; Chalensouk-Khaosaat, J.; González, S. Mechanisms of photoaging and cutaneous photocarcinogenesis, and photoprotective strategies with phytochemicals. Antioxidants 2015, 4, 248–268. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, W.; Pan, H.; Feldser, H.G.; Lainez, E.; Miller, C.; Leung, S.; Zhong, Z.; Zhao, H.; Sweitzer, S.; et al. SIRT1 activators suppress inflammatory responses through promotion of p65 deacetylation and inhibition of NF-κB activity. PLoS ONE 2012, 7, e46364. [Google Scholar] [CrossRef] [PubMed]
- Chin, Y.; Gumilar, K.E.; Li, X.-G.; Tjokroprawiro, B.A.; Lu, C.-H.; Lu, J.; Zhou, M.; Sobol, R.W.; Tan, M. Targeting HSF1 for cancer treatment: Mechanisms and inhibitor development. Theranostics 2023, 13, 2281–2300. [Google Scholar] [CrossRef] [PubMed]
- Fasano, C.; Disciglio, V.; Bertora, S.; Lepore Signorile, M.; Simone, C. FOXO3a from the nucleus to the mitochondria: A round trip in cellular stress response. Cells 2019, 8, 1110. [Google Scholar] [CrossRef]
- Guan, G.; Chen, Y.; Dong, Y. Unraveling the AMPK-SIRT1-FOXO pathway: The in-depth analysis and breakthrough prospects of oxidative stress-induced diseases. Antioxidants 2025, 14, 70. [Google Scholar] [CrossRef]
- Zeng, H.; Xia, H.; Wang, X.; Wang, Y.; Fang, J.; Li, S.; Zhai, Y.; Han, Z. Comprehensive profiling of ceRNA (circRNA-miRNA-mRNA) networks in hypothalamic-pituitary-mammary gland axis of dairy cows under heat stress. Int. J. Mol. Sci. 2023, 24, 888. [Google Scholar] [CrossRef]
- Song, M.; Koo, T. Recent advances in CRISPR technologies for genome editing. Arch. Pharmacal Res. 2021, 44, 537–552. [Google Scholar] [CrossRef]
- Li, H.; Li, S.; Chen, J.; Dai, L.; Chen, R.; Ye, J.; Hao, D. A heat shock 70kDa protein MaltHSP70-2 contributes to thermal resistance in Monochamus alternatus (Coleoptera: Cerambycidae): Quantification, localization, and functional analysis. BMC Genom. 2022, 23, 646. [Google Scholar] [CrossRef]
- Nandy, S.; Pathak, B.; Zhao, S.; Srivastava, V. Heat-shock-inducible CRISPR/Cas9 system generates heritable mutations in rice. Plant Direct 2019, 3, e00145. [Google Scholar] [CrossRef]
- Dikmen, S.; Khan, F.A.; Huson, H.J.; Sonstegard, T.S.; Moss, J.I.; Dahl, G.E.; Hansen, P.J. The SLICK hair locus derived from senepol cattle confers thermotolerance to intensively managed lactating Holstein cows. J. Dairy Sci. 2014, 97, 5508–5520. [Google Scholar] [CrossRef]
- Sigdel, A.; Abdollahi-Arpanahi, R.; Aguilar, I.; Peñagaricano, F. Whole genome mapping reveals novel genes and pathways involved in milk production under heat stress in US Holstein cows. Front. Genet. 2019, 10, 928. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Chai, J.; Xiong, H.; Li, W.; Huang, T.; Liu, Y.; Suo, X.; Zhang, N.; Li, X.; Jiang, S.; et al. Association analysis of HSP70A1A haplotypes with heat tolerance in Chinese Holstein cattle. Cell Stress Chaperon. 2013, 18, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Turman, M.A.; Rosenfeld, S.L. Heat shock protein 70 overexpression protects LLC-PK1 tubular cells from heat shock but not hypoxia. Kidney Int. 1999, 55, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Kisliouk, T.; Cramer, T.; Meiri, N. Methyl CpG level at distal part of heat-shock protein promoter HSP70 exhibits epigenetic memory for heat stress by modulating recruitment of POU2F1-associated nucleosome remodeling deacetylase (NuRD) complex. J. Neurochem. 2017, 141, 358–372. [Google Scholar] [CrossRef]
- Ho, T.-C.; Kim, H.S.; Chen, Y.; Li, Y.; LaMere, M.W.; Chen, C.; Wang, H.; Gong, J.; Palumbo, C.D.; Ashton, J.M.; et al. Scaffold-mediated CRISPR-Cas9 delivery system for acute myeloid leukemia therapy. Sci. Adv. 2021, 7, eabg3217. [Google Scholar] [CrossRef]
- Wu, C. Heat shock transcription factors: Structure and regulation. Annu. Rev. Cell Dev. Biol. 1995, 11, 441–469. [Google Scholar] [CrossRef]
- Wu, R.; Li, X.; Ma, N.; Jin, X.; Yuan, X.; Qu, C.; Tang, H.; Liu, Z.; Zhang, Z. Bacterial quorum sensing molecules promote allergic airway inflammation by activating the retinoic acid response. iScience 2020, 23, 101288. [Google Scholar] [CrossRef]
- Hu, R.; Yang, T.; Ai, Q.; Shi, Y.; Ji, Y.; Sun, Q.; Tong, B.; Chen, J.; Wang, Z. Autoinducer-2 promotes the colonization of Lactobacillus Rhamnosus GG to improve the intestinal barrier function in a neonatal mouse model of antibiotic-induced intestinal dysbiosis. J. Transl. Med. 2024, 22, 177. [Google Scholar] [CrossRef]
- Fu, C.; Ge, J.; Qu, M.; Ouyang, K.; Qiu, Q. Effects of 4-hydroxy-2,5-dimethyl-3(2H)-furanone supplementation on growth performance, serum antioxidant capacity, rumen fermentation characteristics, rumen bacterial quorum sensing, and microbial community in Hu sheep. Anim. Biosci. 2025, 38, 1422–1434. [Google Scholar] [CrossRef]
- Liu, C.; Li, L.; Dai, J.; Qu, M.; Ouyang, K.; Qiu, Q. Heated drinking water in winter improves growth performance of male Hu sheep by modulating rumen quorum sensing and metabolites, and enhancing serum antioxidant capacity. Anim. Biosci. 2025. ahead of print. Available online: https://www.animbiosci.org/journal/view.php?number=25456 (accessed on 6 July 2025). [CrossRef]
- Moura-Alves, P.; Faé, K.; Houthuys, E.; Dorhoi, A.; Kreuchwig, A.; Furkert, J.; Barison, N.; Diehl, A.; Munder, A.; Constant, P.; et al. AhR sensing of bacterial pigments regulates antibacterial defence. Nature 2014, 512, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Meng, F.; Ge, Z.; Zhou, Y.; Fan, Z.; Du, J. Bioinspired peptide/polyamino acid assemblies as quorum sensing inhibitors for the treatment of bacterial infections. J. Mater. Chem. B 2024, 12, 11596–11610. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; He, G.; Yang, Y.; Wang, N.; Zhang, Y.; Su, Y.; Zhao, F.; Wu, J.; Wang, L.; Lin, Y.; et al. Nanomaterials regulate bacterial quorum sensing: Applications, mechanisms, and optimization strategies. Adv. Sci. 2024, 11, 2306070. [Google Scholar] [CrossRef] [PubMed]
- Zenger, K.R.; Khatkar, M.S.; Jones, D.B.; Khalilisamani, N.; Jerry, D.R.; Raadsma, H.W. Genomic selection in aquaculture: Application, limitations and opportunities with special reference to marine shrimp and pearl oysters. Front. Genet. 2019, 9, 693. [Google Scholar] [CrossRef]
- Scherer, N.; Fässler, D.; Borisov, O.; Cheng, Y.; Schlosser, P.; Wuttke, M.; Haug, S.; Li, Y.; Telkämper, F.; Patil, S.; et al. Coupling metabolomics and exome sequencing reveals graded effects of rare damaging heterozygous variants on gene function and human traits. Nat. Genet. 2025, 57, 193–205. [Google Scholar] [CrossRef]
- Christensen, O.F.; Börner, V.; Varona, L.; Legarra, A. Genetic evaluation including intermediate omics features. Genetics 2021, 219, iyab130. [Google Scholar] [CrossRef]
- Cui, H.; Tejada-Lapuerta, A.; Brbić, M.; Saez-Rodriguez, J.; Cristea, S.; Goodarzi, H.; Lotfollahi, M.; Theis, F.J.; Wang, B. Towards multimodal foundation models in molecular cell biology. Nature 2025, 640, 623–633. [Google Scholar] [CrossRef]
- Wu, Y.; Xie, L. AI-driven multi-omics integration for multi-scale predictive modeling of causal genotype-environment-phenotype relationships. Comput. Struct. Biotechnol. J. 2025, 27, 265–277. [Google Scholar] [CrossRef]
- Sharma, A.; Kosasih, E.; Zhang, J.; Brintrup, A.; Calinescu, A. Digital twins: State of the art theory and practice, challenges, and open research questions. J. Ind. Inf. Integr. 2022, 30, 100383. [Google Scholar] [CrossRef]
- Han, X.; Lin, Z.; Clark, C.; Vucetic, B.; Lomax, S. AI based digital twin model for cattle caring. Sensors 2022, 22, 7118. [Google Scholar] [CrossRef]
- Abdulai, A.-R.; Tetteh Quarshie, P.; Duncan, E.; Fraser, E. Is agricultural digitization a reality among smallholder farmers in Africa? Unpacking farmers’ lived realities of engagement with digital tools and services in rural Northern Ghana. Agric. Food Secur. 2023, 12, 11. [Google Scholar] [CrossRef]
- Global Gene Editing Regulation Tracker. Available online: https://crispr-gene-editing-regs-tracker.geneticliteracyproject.org/ (accessed on 11 March 2025).
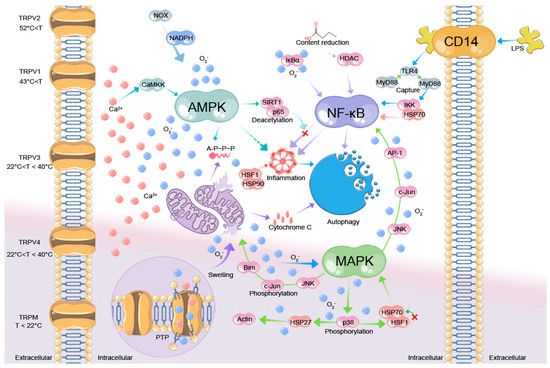
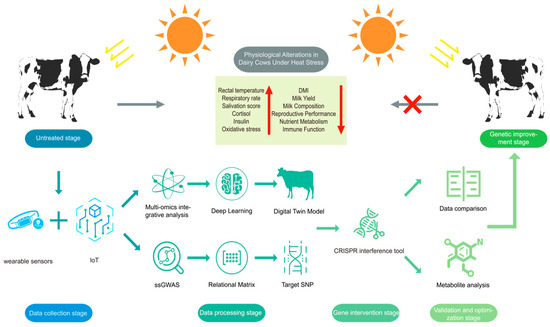
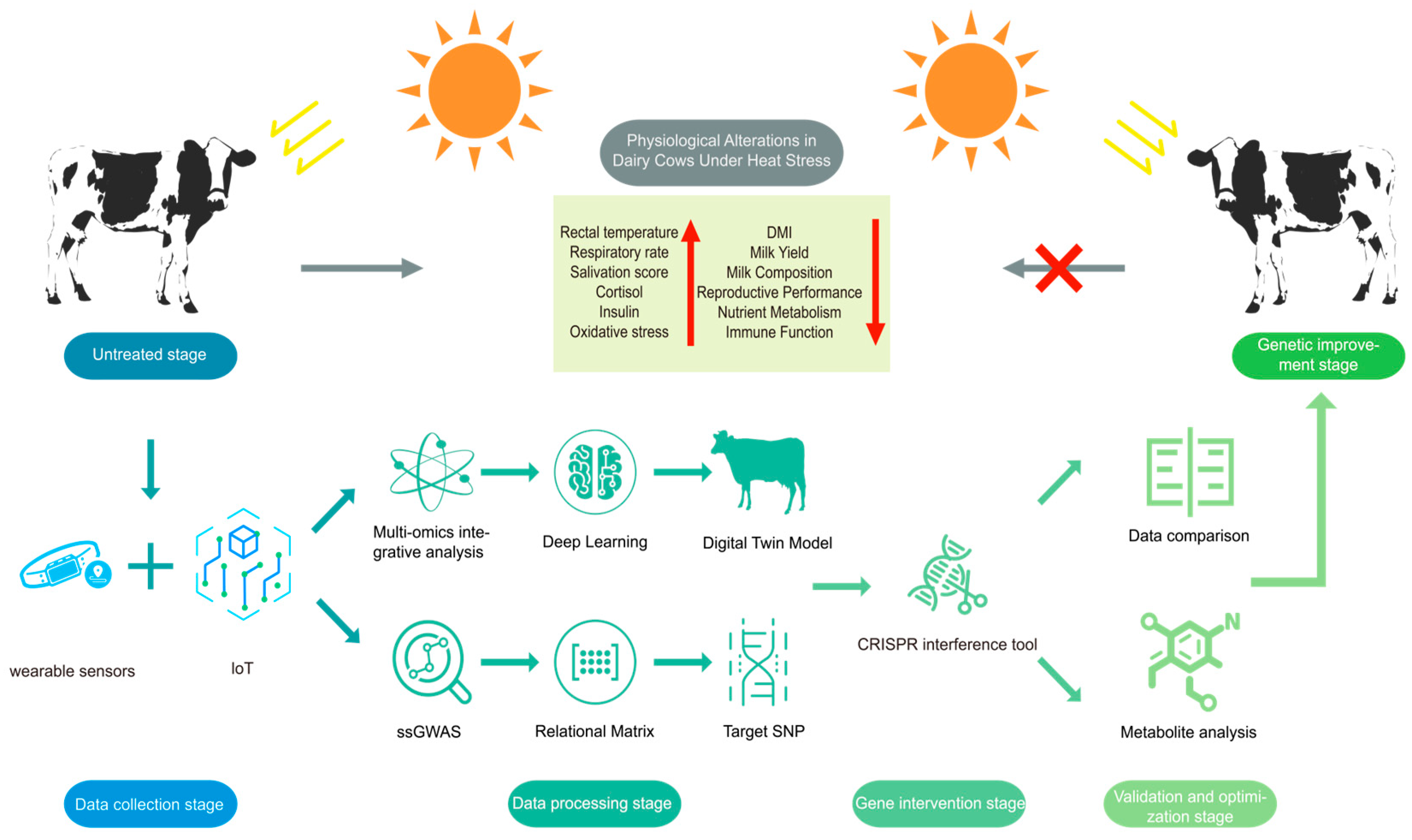
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).