Harnessing Bacillus subtilis–Moss Synergy: Carbon–Structure Optimization for Erosion-Resistant Barrier Formation in Cold Mollisols

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Test Materials
2.1.1. Soil Sample Collection
2.1.2. Preparation of Bacillus subtilis Inoculant
2.1.3. Moss Source Preparation and Application Methods
2.2. Experimental Design
2.3. Measurement Indicators and Methods
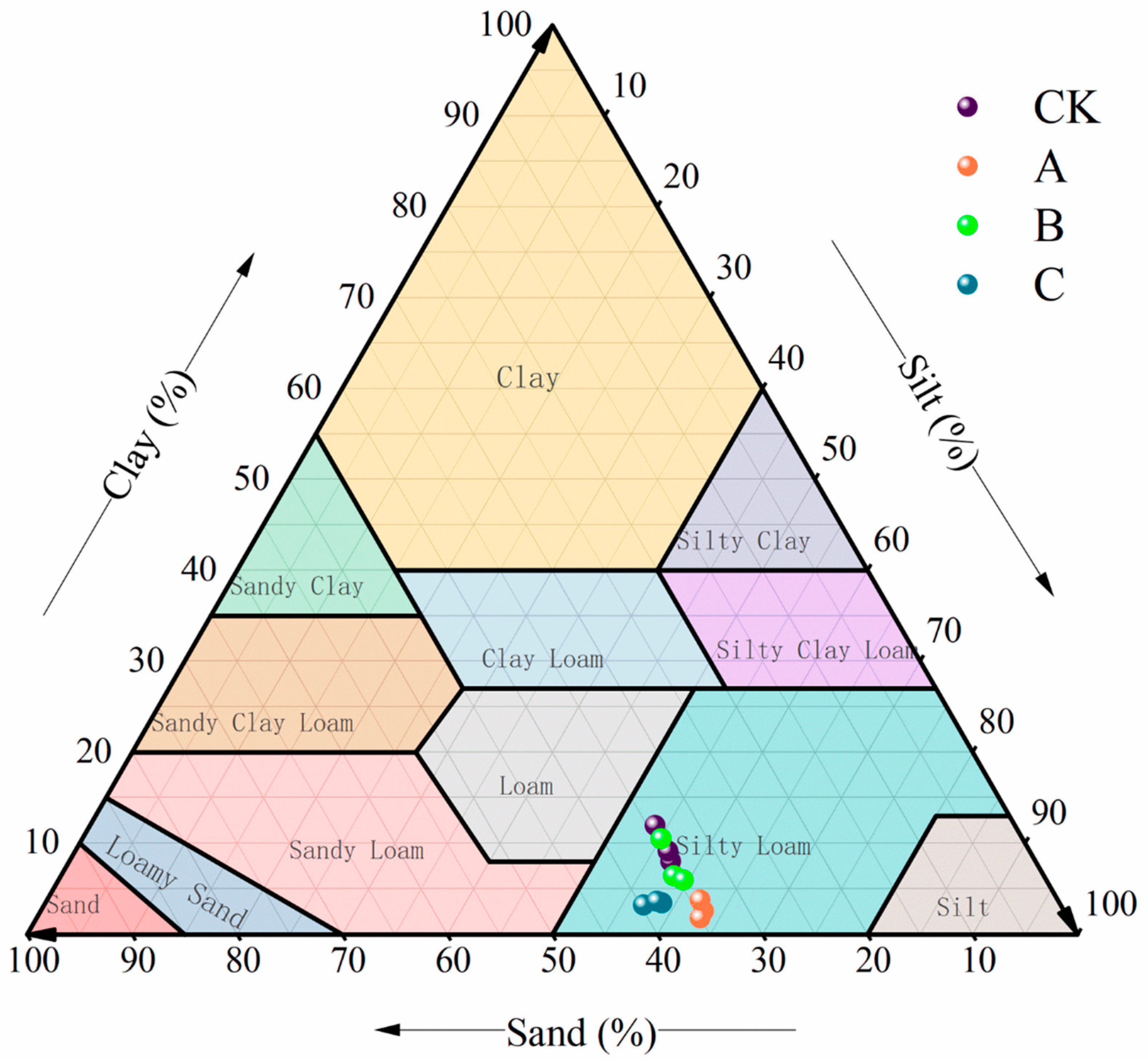
2.3.1. Soil Particle Size Analysis
2.3.2. Soil Moisture Characterization Curve
2.3.3. Water-Stable Soil Aggregates
2.3.4. Runoff and Sediment Yield
2.3.5. Remaining Soil Indicators
2.4. Statistical Analysis
3. Results
3.1. Differences in Soil Particle Size Distribution and Soil Aggregate Stability in Different Treatment Groups
3.1.1. Soil Particle Size Distribution
3.1.2. Water-Stable Soil Aggregates
3.2. Effects of Bacterial and Moss Inoculation on Soil Moisture Characteristics
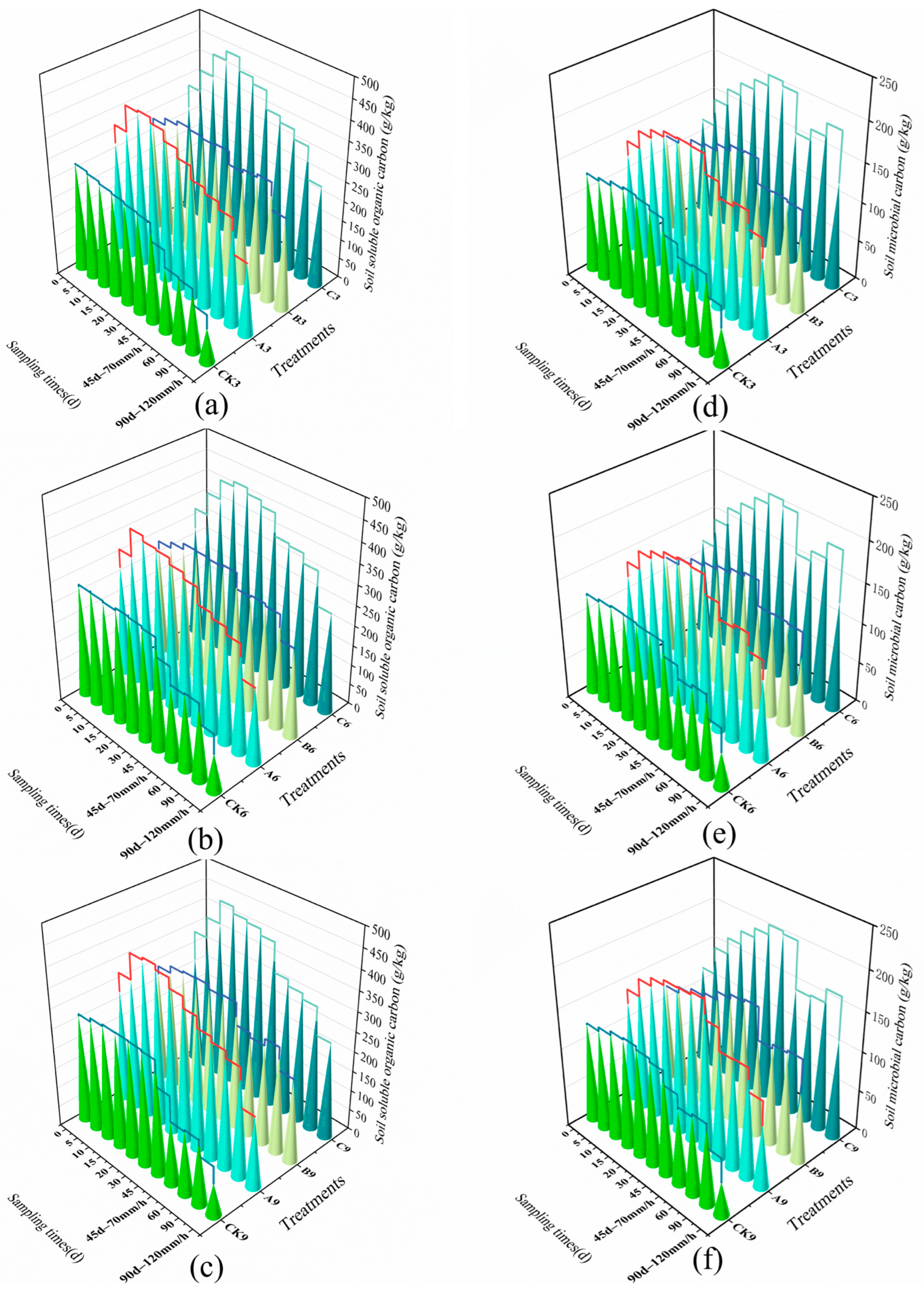
3.3. Characteristics of the Effects of Bacterial and Moss Inoculation on Soil Organic Carbon
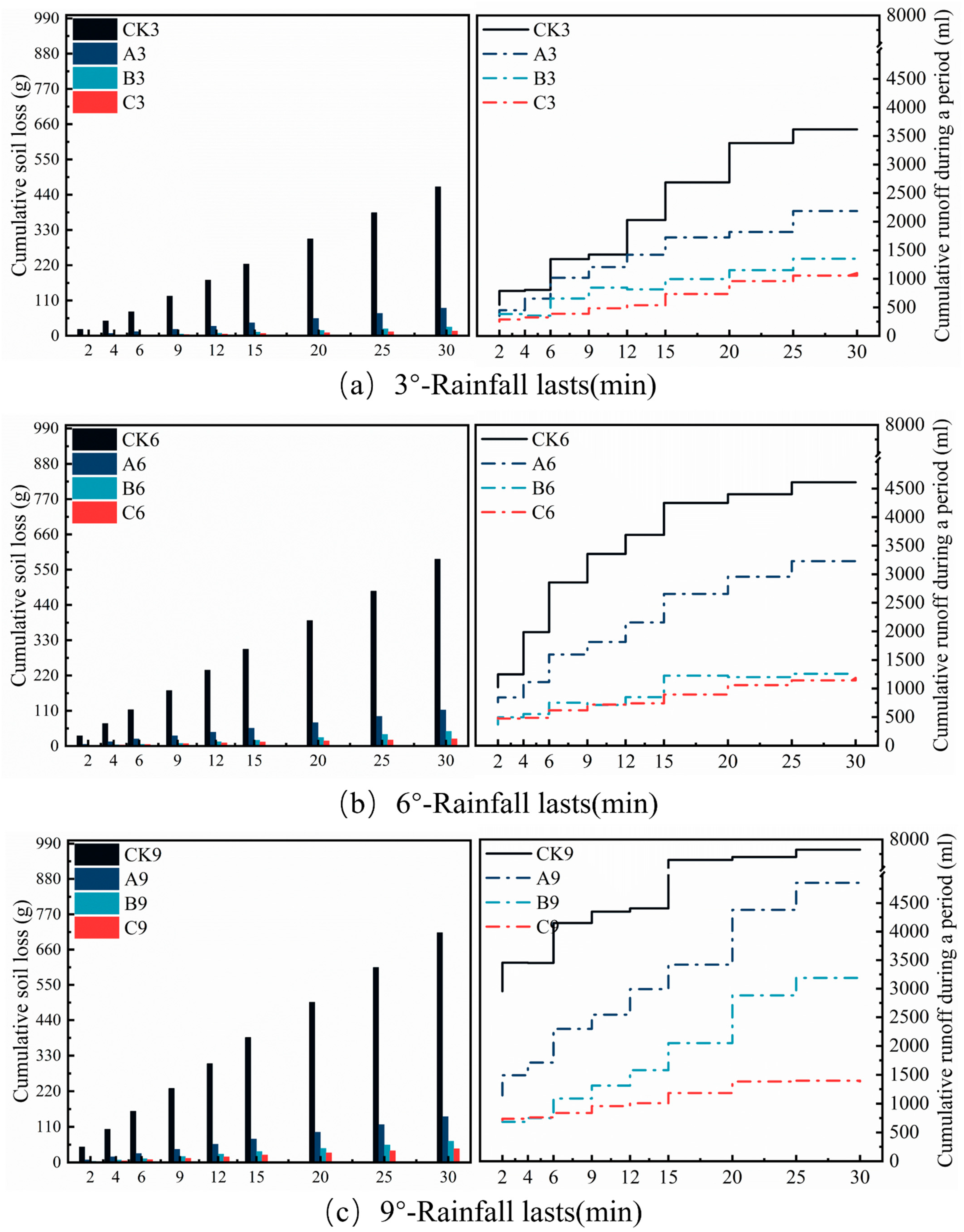
3.4. Effects of Bacterial and Moss Inoculation on Rainfall-Induced Soil Erosion
3.5. Correlation Between Soil Physicochemical Properties and Rainfall Erosion Indicators
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sáez-Sandino, T.; Delgado-Baquerizo, M. Soil carbon worldwide is reduced by an increasing number of global change stressors. Nat. Clim. Change 2024, 14, 683–684. [Google Scholar] [CrossRef]
- Borrelli, P.; Robinson, D.A.; Fleischer, L.R.; Lugato, E.; Ballabio, C.; Alewell, C.; Meusburger, K.; Modugno, S.; Schutt, B.; Ferro, V.; et al. An assessment of the global impact of 21st century land use change on soil erosion. Nat. Commun. 2017, 8, 2013. [Google Scholar] [CrossRef]
- Li, J.; Pei, J.; Fang, C.; Li, B.; Nie, M. Drought may exacerbate dryland soil inorganic carbon loss under warming climate conditions. Nat. Commun. 2024, 15, 617. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, S.; Guo, X.; Xiao, L.; Yang, Y.; Luo, Y.; Mishra, U.; Luo, Z. Responses of soil organic carbon to climate extremes under warming across global biomes. Nat. Clim. Change 2023, 14, 98–105. [Google Scholar] [CrossRef]
- Wuepper, D.; Borrelli, P.; Finger, R. Countries and the global rate of soil erosion. Nat. Sustain. 2019, 3, 51–55. [Google Scholar] [CrossRef]
- Fang, H.; Zhai, Y.; Li, C. Evaluating the impact of soil erosion on soil quality in an agricultural land, northeastern China. Sci. Rep. 2024, 14, 15629. [Google Scholar] [CrossRef]
- Ma, R.; Tian, Z.; Zhao, Y.; Wu, Y.; Liang, Y. Response of soil quality degradation to cultivation and soil erosion: A case study in a Mollisol region of Northeast China. Soil Tillage Res. 2024, 242, 106159. [Google Scholar] [CrossRef]
- Wang, H.; Yang, S.; Wang, Y.; Gu, Z.; Xiong, S.; Huang, X.; Sun, M.; Zhang, S.; Guo, L.; Cui, J.; et al. Rates and causes of black soil erosion in Northeast China. Catena 2022, 214, 106250. [Google Scholar] [CrossRef]
- Yuan, C.; Fan, H. Response mechanism of black soil structure to compound erosion forces in sloping farmland, Northeast China. Soil Tillage Res. 2024, 240, 106103. [Google Scholar] [CrossRef]
- Anderson, R.L.; Rowntree, K.M.; Le Roux, J.J. An interrogation of research on the influence of rainfall on gully erosion. Catena 2021, 206, 105482. [Google Scholar] [CrossRef]
- Zhao, X.; Song, X.; Li, L.; Wang, D.; Meng, P.; Li, H. Effect of microrelief features of tillage methods under different rainfall intensities on runoff and soil erosion in slopes. Int. Soil Water Conserv. Res. 2024, 12, 351–364. [Google Scholar] [CrossRef]
- Chen, P.; Xie, Y.; Ren, X.; Cheng, C.; Wei, X. Spatial variation of soil organic carbon density in the black soil region of Northeast China under the influence of erosion and deposition. J. Clean. Prod. 2024, 475, 143616. [Google Scholar] [CrossRef]
- Li, X.; Fan, H.; Wang, P.; Zhang, X.; Li, A.; Yang, X.; Zhang, G. Interactive effect of soil dispersity and rainfall intensity on splash erosion: Insights from laboratory tests. Catena 2024, 238, 107843. [Google Scholar] [CrossRef]
- Wang, N.; Zheng, Z.; Li, T.; He, S.; Zhang, X.; Wang, Y.; Yu, H.; Huang, H.; Ye, D. Soil surface roughness impacts erosion behavior through selective regulation of flow properties in rainfall-seepage scenarios. Soil Tillage Res. 2025, 246, 106350. [Google Scholar] [CrossRef]
- Wu, X.; Cai, C.; Li, D.; Zhou, J.; Zhang, W. Non-linear response of sediment size characteristics and associated transport patterns to soil structural stability in sheet erosion under field rainfall simulation. Catena 2023, 228, 107120. [Google Scholar] [CrossRef]
- Coban, O.; De Deyn, G.B.; van der Ploeg, M. Soil microbiota as game-changers in restoration of degraded lands. Science 2022, 375, abe0725. [Google Scholar] [CrossRef]
- Phillips, M.L.; McNellis, B.E.; Howell, A.; Lauria, C.M.; Belnap, J.; Reed, S.C. Biocrusts mediate a new mechanism for land degradation under a changing climate. Nat. Clim. Change 2022, 12, 71–76. [Google Scholar] [CrossRef]
- Chamizo, S.; Rodríguez-Caballero, E.; Román, J.R.; Cantón, Y. Effects of biocrust on soil erosion and organic carbon losses under natural rainfall. Catena 2017, 148, 117–125. [Google Scholar] [CrossRef]
- Zeng, Z.; Liu, D.D.; Dai, Q.H.; Zeng, J.; Jiang, J. Effects of moss patches on the sediment loss, flow hydraulics and surface microtopography of soil slopes in karst mountainous areas. Catena 2025, 249, 108672. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, Z.L.; Shen, N. Plot-based study to evaluate raindrop detachment capacity on moss-dominated biocrusted slope under simulated rainfall with different intensities. Catena 2023, 226, 107084. [Google Scholar] [CrossRef]
- Reed, S.C.; Coe, K.K.; Sparks, J.P.; Housman, D.C.; Zelikova, T.J.; Belnap, J. Changes to dryland rainfall result in rapid moss mortality and altered soil fertility. Nat. Clim. Change 2012, 2, 752–755. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Zhang, S.; Huang, Y.; Ouyang, Z.; Mai, Z. Biological soil crust elicits microbial community and extracellular polymeric substances restructuring to reduce the soil erosion on tropical island, South China Sea. Mar. Environ. Res. 2024, 197, 106449. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.; Yu, X.; Jia, G. Soil microorganism regulated aggregate stability and rill erosion resistance under different land uses. Catena 2023, 228, 107176. [Google Scholar] [CrossRef]
- Li, C.; Chen, X.; Jia, Z.; Zhai, L.; Zhang, B.; Gruters, U.; Ma, S.; Qian, J.; Liu, X.; Zhang, J.; et al. Meta-analysis reveals the effects of microbial inoculants on the biomass and diversity of soil microbial communities. Nat. Ecol. Evol. 2024, 8, 1270–1284. [Google Scholar] [CrossRef]
- Sáez-Sandino, T.; García-Palacios, P.; Maestre, F.T.; Plaza, C.; Guirado, E.; Singh, B.K.; Wang, J.; Cano-Díaz, C.; Eisenhauer, N.; Gallardo, A.; et al. The soil microbiome governs the response of microbial respiration to warming across the globe. Nat. Clim. Change 2023, 13, 1382–1387. [Google Scholar] [CrossRef]
- Deng, Z.; Wang, J.; Yan, Y.; Wang, J.; Shao, W.; Wu, Z. Biochar-based Bacillus subtilis inoculants promote plant growth: Regulating microbial community to improve soil properties. J. Environ. Manag. 2024, 373, 123534. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.K.; Morris, D.J.P.; Vogt, S.; Gleber, S.C.; Bigalke, M.; Wilcke, W.; Rillig, M.C. Visualizing the dynamics of soil aggregation as affected by arbuscular mycorrhizal fungi. ISME J. 2019, 13, 1639–1646. [Google Scholar] [CrossRef]
- Li, Z.; Kravchenko, A.N.; Cupples, A.; Guber, A.K.; Kuzyakov, Y.; Philip Robertson, G.; Blagodatskaya, E. Composition and metabolism of microbial communities in soil pores. Nat. Commun. 2024, 15, 3578. [Google Scholar] [CrossRef]
- Ruan, Z.; Chen, K.; Cao, W.; Meng, L.; Yang, B.; Xu, M.; Xing, Y.; Li, P.; Freilich, S.; Chen, C.; et al. Engineering natural microbiomes toward enhanced bioremediation by microbiome modeling. Nat. Commun. 2024, 15, 4694. [Google Scholar] [CrossRef]
- Liao, K.J.; Tao, Y.; Zeng, Y.Y.; Tu, J.W.; She, S.J.; Fu, Y.J.; Hou, L.H.; Chen, L.Z. A feasible method of induced biological soil crust propagation through the inoculation of moss and addition of soil amendments in a Pb-Zn tailing pond. Sci. Total Environ. 2024, 910, 168569. [Google Scholar] [CrossRef]
- Bittelli, M.; Andrenelli, M.C.; Simonetti, G.; Pellegrini, S.; Artioli, G.; Piccoli, I.; Morari, F. Shall we abandon sedimentation methods for particle size analysis in soils? Soil Tillage Res. 2019, 185, 36–46. [Google Scholar] [CrossRef]
- Karup, D.; Moldrup, P.; Tuller, M.; Arthur, E.; de Jonge, L.W. Prediction of the soil water retention curve for structured soil from saturation to oven-dryness. Eur. J. Soil Sci. 2017, 68, 57–65. [Google Scholar] [CrossRef]
- Jones, D.L.; Willett, V.B. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Sadeghi, S.H.; Najafinejad, A.; Gharemahmudli, S.; Darki, B.Z.; Behbahani, A.M.; Kheirfam, H. Reduction in soil loss caused by a freeze-thaw cycle through inoculation of endemic soil microorganisms. Appl. Soil Ecol. 2021, 157, 103770. [Google Scholar] [CrossRef]
- Xiao, Z.X.; Hou, R.J.; Li, T.X.; Meng, F.X.; Fu, Q.; Li, M.; Liu, D.; Ji, Y.; Dong, S.Q. Mechanism of microbial inhibition of rainfall erosion in black soil area, as a soil structure builder. Soil Tillage Res. 2023, 233, 105819. [Google Scholar] [CrossRef]
- Liu, Y.P.; Jia, R.L.; Wang, W.F.; Wan, Y.C.; Gao, Y.H.; Zhan, H.T.; Ren, J.; Chen, Z.; Qiu, F.; Zhu, J. Vascular plant communities and biocrusts act as controlling factors in mitigating soil erosion on the Great Wall in a semi-humid area of Northwestern China. Sci. Total Environ. 2024, 918, 170515. [Google Scholar] [CrossRef]
- Sun, F.H.; Xiao, B.; Tuller, M. Multifaceted impacts of moss-dominated biocrusts on dryland soils: From soil pore structure to aeration and water infiltration. Catena 2024, 236, 107755. [Google Scholar] [CrossRef]
- Vaezi, A.R.; Sadeghian, N.; Cerdà, A. Particle size distribution of sediment detached from rills under raindrop impact in semi-arid soils. J. Hydrol. 2020, 590, 125317. [Google Scholar] [CrossRef]
- Gharemahmudli, S.; Sadeghi, S.H.; Najafinejad, A.; Darki, B.Z.; Behbahani, A.M.; Kheirfam, H. Controlling enhanced surface runoff components as a result of a freezing-thawing cycle by inoculating soil bacteria and cyanobacteria. Soil Tillage Res. 2024, 237, 105989. [Google Scholar] [CrossRef]
- Gao, L.Q.; Bowker, M.A.; Sun, H.; Zhao, J.; Zhao, Y.G. Linkages between biocrust development and water erosion and implications for erosion model implementation. Geoderma 2020, 357, 113973. [Google Scholar] [CrossRef]
- Sun, F.H.; Xiao, B.; Kidron, G.J. Towards the influences of three types of biocrusts on soil water in drylands: Insights from horizontal infiltration and soil water retention. Geoderma 2022, 428, 116136. [Google Scholar] [CrossRef]
- Wang, W.; Li, M.Y.; Zhou, R.; Mo, F.; Wang, B.Z.; Zhu, L.; Tao, H.Y.; Zhu, Y.; Wang, W.L.; Zhao, Z.Y.; et al. Moss-dominated biocrust-based biodiversity enhances carbon sequestration via water interception and plant-soil-microbe interactions. Iscience 2023, 26, 105773. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.W.W.; Peprah-Manu, D. Pore structure effects on the water retention behaviour of a compacted silty sand soil subjected to drying-wetting cycles. Eng. Geol. 2023, 313, 106963. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Inoculation Treatment | Slope | Rainfall Intensity | Process Description | Sample Size |
|---|---|---|---|---|---|
| Bacillus subtilis treatment group (A) | 3 L m−2 Bacillus subtilis culture solution | 3° | 70 mm·h−1, 120 mm·h−1 | A3 | 3 |
| 6° | 70 mm·h−1, 120 mm·h−1 | A6 | 3 | ||
| 9° | 70 mm·h−1, 120 mm·h−1 | A9 | 3 | ||
| Moss treatment group (B) | 0.6 kg m−2 moss (fresh weight) | 3° | 70 mm·h−1, 120 mm·h−1 | B3 | 3 |
| 6° | 70 mm·h−1, 120 mm·h−1 | B6 | 3 | ||
| 9° | 70 mm·h−1, 120 mm·h−1 | B9 | 3 | ||
| Moss + Bacillus subtilis treatment group (C) | 3 L m−2 Bacillus subtilis culture solution, 0.6 kg m−2 moss (fresh weight) | 3° | 70 mm·h−1, 120 mm·h−1 | C3 | 3 |
| 6° | 70 mm·h−1, 120 mm·h−1 | C6 | 3 | ||
| 9° | 70 mm·h−1, 120 mm·h−1 | C9 | 3 | ||
| Control group (CK) | 3 L m−2 sterile water | 3° | 70 mm·h−1, 120 mm·h−1 | CK3 | 3 |
| 6° | 70 mm·h−1, 120 mm·h−1 | CK6 | 3 | ||
| 9° | 70 mm·h−1, 120 mm·h−1 | CK9 | 3 |
| Modeling Parameters | xc (μm) | w (μm) | A (%·μm) | R2 |
|---|---|---|---|---|
| CK3 | 6.812 ± 0.314 e | 6.571 ± 0.653 c | 70.55 ± 8.321 d | 0.82 |
| A3 | 14.793 ± 0.762 bc | 16.67 ± 1.748 ab | 142.723 ± 19.822 bc | 0.79 |
| B3 | 14.358 ± 0.474 bc | 12.415 ± 1.042 b | 169.776 ± 17.455 b | 0.88 |
| C3 | 11.841 ± 0.714 cd | 12.821 ± 1.578 b | 122.134 ± 19.221 c | 0.74 |
| CK6 | 8.695 ± 0.360 e | 8.913 ± 0.767 c | 83.757 ± 8.835 d | 0.85 |
| A6 | 10.528 ± 0.477 d | 10.934 ± 1.031 bc | 126.479 ± 14.972 c | 0.83 |
| B6 | 8.000 ± 0.231 e | 7.979 ± 0.488 c | 100.452 ± 7.437 cd | 0.92 |
| C6 | 19.618 ± 1.437 ab | 23.490 ± 3.560 a | 206.061 ± 43.733 a | 0.75 |
| CK9 | 11.775 ± 0.577 c | 13.475 ± 1.301 b | 114.288 ± 14.109 c | 0.82 |
| A9 | 17.175 ± 1.136 b | 19.982 ± 2.702 a | 194.228 ± 35.763 ab | 0.70 |
| B9 | 11.669 ± 0.823 c | 13.47 ± 1.862 b | 108.209 ± 19.097 cd | 0.69 |
| C9 | 26.92 ± 1.797 a | 31.978 ± 4.971 a | 260.702 ± 60.691 a | 0.71 |
| Average value of the same treatment | ||||
| CK | 9.094 ± 1.447 | 9.653 ± 2.053 | 89.531 ± 13.121 | 0.83 |
| A | 14.165 ± 1.945 | 15.862 ± 2.709 | 154.476 ± 20.876 | 0.77 |
| B | 11.342 ± 1.863 | 11.288 ± 1.652 | 126.145 ± 21.536 | 0.83 |
| C | 19.459 ± 4.376 | 22.763 ± 5.883 | 196.299 ± 42.879 | 0.78 |
| T-test | <0.01 | <0.01 | <0.01 | |
| Treatments | MWD (mm) | GWD (mm) | WR0.25 (%) | PAD (%) | D |
|---|---|---|---|---|---|
| CK3 | 0.38 ± 0.04 e | 0.27 ± 0.02 ef | 35.81 ± 2.41 a | 37.08 ± 2.64 a | 2.51 ± 0.01 a |
| A3 | 0.67 ± 0.04 b | 0.35 ± 0.02 bc | 49.41 ±1.49 bcd | 29.02 ± 2.56 bc | 2.49 ± 0.02 ab |
| B3 | 0.48 ± 0.02 c | 0.32 ± 0.03 cd | 44.34 ± 0.31 e | 29.36 ± 0.43 bc | 2.50 ± 0.01 ab |
| C3 | 0.84 ± 0.02 a | 0.47 ± 0.01 a | 52.72 ± 3.53 ab | 22.07 ± 3.92 de | 2.43 ± 0.02 c |
| CK6 | 0.34 ± 0.03 e | 0.24 ± 0.03 f | 37.68 ± 2.18 a | 33.72 ± 3.41 ab | 2.51 ± 0.01 a |
| A6 | 0.65 ± 0.04 b | 0.36 ± 0.02 bc | 51.15 ± 1.32 ab | 26.51 ± 2.93 cd | 2.49 ± 0.03 ab |
| B6 | 0.46 ± 0.06 cd | 0.30 ± 0.01 de | 46.74 ±1.39 cde | 25.55 ±1.63 cde | 2.50 ± 0.01 ab |
| C6 | 0.83 ± 0.03 a | 0.47 ± 0.01 a | 53.92 ± 2.27 a | 20.24 ± 2.78 e | 2.43 ± 0.02 c |
| CK9 | 0.39 ± 0.05 de | 0.27 ± 0.02 ef | 36.15 ± 1.14 a | 36.42 ± 0.88 a | 2.50 ± 0.03 ab |
| A9 | 0.66 ± 0.02 b | 0.39 ± 0.02 b | 50.21 ±1.29 abc | 27.85 ±2.89 bcd | 2.48 ± 0.02 ab |
| B9 | 0.47 ± 0.04 c | 0.31 ± 0.01 de | 46.01 ± 0.90 de | 26.69 ± 2.12 cd | 2.50 ± 0.03 ab |
| C9 | 0.80 ± 0.02 a | 0.46 ± 0.01 a | 51.72 ± 1.91 ab | 23.45 ± 0.40 cde | 2.45 ± 0.07 bc |
| Average value of the same treatment | |||||
| CK | 0.37 ± 0.04 | 0.26 ± 0.02 | 36.55 ± 1.91 | 35.74 ± 2.31 | 2.51 ± 0.02 |
| A | 0.66 ± 0.03 | 0.37 ± 0.02 | 50.26 ± 1.37 | 27.79 ± 2.80 | 2.49 ± 0.02 |
| B | 0.47 ± 0.04 | 0.31 ± 0.01 | 45.70 ± 0.87 | 27.20 ± 1.40 | 2.50 ± 0.01 |
| C | 0.82 ± 0.03 | 0.46 ± 0.01 | 52.79 ± 2.57 | 21.92 ± 3.37 | 2.43 ± 0.04 |
| T-test | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Treatments | θs (cm3·cm−3) | θr (cm3·cm−3) | FC (cm3·cm−3) | PWP (cm3·cm−3) | PAWC (cm3·cm−3) | a (kPa−1) |
|---|---|---|---|---|---|---|
| CK3 | 0.519 ± 0.012 a | 0.113 ± 0.007 de | 0.312 ± 0.016 ab | 0.261 ± 0.005 ab | 0.051 ± 0.004 bc | 0.100 ± 0.023 a |
| A3 | 0.431 ± 0.006 bd | 0.152 ± 0.006 a | 0.337 ± 0.008 a | 0.285 ± 0.001 a | 0.052 ± 0.009 bc | 0.028 ± 0.006 c |
| B3 | 0.473 ± 0.009 b | 0.103 ± 0.004 ef | 0.29 ± 0.008 bc | 0.238 ± 0.004 bc | 0.052 ± 0.007 bc | 0.100 ± 0.017 a |
| C3 | 0.431 ± 0.010 cd | 0.131 ± 0.007 bc | 0.312 ± 0.008 ab | 0.256 ± 0.002 b | 0.056 ± 0.005 bc | 0.054 ± 0.015 bc |
| CK6 | 0.517 ± 0.014 a | 0.102 ± 0.007 ef | 0.313 ± 0.008 ab | 0.253 ± 0.007 b | 0.060 ± 0.005 a | 0.100 ± 0.026 a |
| A6 | 0.427 ± 0.006 cd | 0.142 ± 0.006 ab | 0.336 ± 0.003 ac | 0.235 ± 0.004 bc | 0.101 ± 0.006 ab | 0.024 ± 0.005 c |
| B6 | 0.477 ± 0.014 bc | 0.096 ± 0.006 f | 0.288 ± 0.007 b | 0.209 ± 0.005 c | 0.079 ± 0.010 bc | 0.100 ± 0.026 a |
| C6 | 0.431 ± 0.008 cd | 0.129 ± 0.006 cd | 0.317 ± 0.004 ab | 0.258 ± 0.005 b | 0.059 ± 0.001 bc | 0.045 ± 0.011 bc |
| CK9 | 0.508 ± 0.017 a | 0.090 ± 0.009 f | 0.296 ± 0.006 bc | 0.244 ± 0.005 bc | 0.052 ± 0.002 bc | 0.100 ± 0.031 a |
| A9 | 0.426 ± 0.006 cd | 0.167 ± 0.005 a | 0.322 ± 0.014 ab | 0.275 ± 0.006 a | 0.047 ± 0.005 c | 0.051 ± 0.011 bc |
| B9 | 0.455 ± 0.015 bc | 0.098 ± 0.007 f | 0.269 ± 0.021 c | 0.227 ± 0.007 c | 0.042 ± 0.007 c | 0.100 ± 0.031 a |
| C9 | 0.421 ± 0.006 d | 0.121 ± 0.005 de | 0.307 ± 0.016 ab | 0.250 ± 0.008 b | 0.057 ± 0.004 bc | 0.042 ± 0.008 c |
| Average value of the same treatment | ||||||
| CK | 0.515 ± 0.005 a | 0.102 ± 0.011 ab | 0.307 ± 0.008 b | 0.253 ± 0.008 ab | 0.054 ± 0.004 b | 0.100 ± 0.002 a |
| A | 0.428 ± 0.003 c | 0.154 ± 0.012 a | 0.332 ± 0.007 a | 0.265 ± 0.024 a | 0.067 ± 0.027 a | 0.034 ± 0.014 b |
| B | 0.468 ± 0.011 b | 0.099 ± 0.003 ab | 0.282 ± 0.010 c | 0.225 ± 0.013 c | 0.058 ± 0.017 b | 0.100 ± 0.011 a |
| C | 0.428 ± 0.005 c | 0.127 ± 0.005 ab | 0.312 ± 0.005 b | 0.255 ± 0.004 a | 0.057 ± 0.001 b | 0.047 ± 0.006 b |
| Treatment | Soil Loss (g) | Runoff Volume (L) | Infiltration Volume (L) | Sediment Concentration (g·L−1) |
|---|---|---|---|---|
| CK3 | 121.17 ± 3.15 c | 7.44 ± 1.44 b | 17.24 ± 1.98 bc | 16.29 ± 1.51 a |
| A3 | 23.83 ± 1.48 f | 5.05 ± 0.86 c | 19.11 ± 2.24 b | 4.72 ± 0.25 b |
| B3 | 8.28 ± 0.77 i | 4.01 ± 0.65 c | 21.10 ± 1.85 a | 2.07 ± 0.25 c |
| C3 | 5.21 ± 0.70 j | 4.59 ± 0.19 c | 19.15 ± 1.91 b | 1.13 ± 0.11 d |
| CK6 | 195.84 ± 5.19 b | 13.11 ± 1.12 a | 14.18 ± 1.87 c | 14.94 ± 1.01 a |
| A6 | 38.39 ± 3.45 e | 9.86 ± 1.01 b | 17.45 ± 1.49 bc | 3.90 ± 0.55 b |
| B6 | 19.27 ± 1.10 g | 5.95 ± 0.25 c | 19.15 ± 3.44 b | 3.24 ± 0.07 c |
| C6 | 14.92 ± 0.85 h | 5.91 ± 0.99 c | 17.76 ± 2.74 bc | 2.52 ± 0.20 d |
| CK9 | 380.88 ± 5.95 a | 30.99 ± 2.55 a | 11.42 ± 1.64 c | 12.29 ± 1.05 a |
| A9 | 54.23 ± 3.96 d | 18.16 ± 1.31 b | 9.15 ± 1.29 d | 2.99 ± 0.33 c |
| B9 | 30.84 ± 1.48 g | 11.50 ± 1.01 b | 12.98 ± 1.45 c | 2.68 ± 0.05 c |
| C9 | 22.38 ± 2.18 h | 10.93 ± 1.54 b | 10.01 ± 1.08 c | 2.05 ± 0.11 c |
| Treatment | Soil Loss (g) | Runoff Volume (L) | Infiltration Volume (L) | Sediment Concentration (g·L−1) |
|---|---|---|---|---|
| CK3 | 465.02 ± 8.15 d | 16.62 ± 3.10 c | 24.41 ± 2.83 c | 27.98 ± 1.81 a |
| A3 | 86.41 ± 4.23 h | 10.83 ± 1.25 d | 21.78 ± 1.19 d | 7.98 ± 0.15 d |
| B3 | 27.14 ± 1.57 k | 6.94 ± 0.44 e | 29.51 ± 2.18 a | 3.91 ± 0.06 f |
| C3 | 15.13 ± 1.10 i | 5.45 ± 0.28 e | 24.70 ± 2.05 c | 2.78 ± 0.04 g |
| CK6 | 582.85 ± 9.16 b | 27.41 ± 3.19 b | 23.76 ± 1.43 c | 21.26 ± 1.05 b |
| A6 | 112.3 ± 2.08 g | 17.12 ± 2.65 c | 20.54 ± 1.65 d | 6.56 ± 0.25 e |
| B6 | 45.81 ± 2.16 j | 7.41 ± 1.10 e | 27.92 ± 4.18 ab | 6.18 ± 0.13 e |
| C6 | 22.28 ± 1.53 k | 6.97 ± 0.83 e | 22.45 ± 2.75 cd | 3.20 ± 0.03 f |
| CK9 | 712.94 ± 10.50 a | 37.46 ± 3.64 a | 23.61 ± 2.06 c | 19.03 ± 1.41 c |
| A9 | 141.51 ± 5.74 f | 24.83 ± 2.79 b | 20.27 ± 1.46 d | 5.70 ± 0.11 e |
| B9 | 65.73 ± 4.45 i | 14.22 ± 1.16 d | 25.76 ± 3.11 bc | 4.62 ± 0.09 f |
| C9 | 42.16 ± 2.77 j | 9.32 ± 0.85 de | 22.64 ± 3.66 cd | 4.53 ± 0.28 f |
| Soil Parameters | RV | SL | SC |
|---|---|---|---|
| Xc | −0.378 | −0.469 | −0.497 |
| w | −0.175 | −0.252 | −0.280 |
| A | −0.420 | −0.510 | −0.517 |
| a | 0.362 | 0.447 | 0.497 |
| us | 0.338 | 0.423 | 0.423 |
| DEI | 0.004 | 0.063 | 0.105 |
| MWD | −0.671 * | −0.741 ** | −0.769 ** |
| GMD | −0.649 * | −0.709 ** | −0.709 ** |
| WR0.25 | −0.594 * | −0.657 * | −0.643 * |
| PAD | 0.699 * | 0.720 ** | 0.657 * |
| D | 0.544 | 0.619 * | 0.623 * |
| D0C | −0.923 ** | −0.923 ** | −0.853 ** |
| MBC | −0.874 ** | −0.871 ** | −0.793 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Zheng, S.; Xiao, Z.; Fu, Q.; Meng, F.; Li, M.; Liu, D.; Liu, Q. Harnessing Bacillus subtilis–Moss Synergy: Carbon–Structure Optimization for Erosion-Resistant Barrier Formation in Cold Mollisols. Agriculture 2025, 15, 1465. https://doi.org/10.3390/agriculture15141465
Li T, Zheng S, Xiao Z, Fu Q, Meng F, Li M, Liu D, Liu Q. Harnessing Bacillus subtilis–Moss Synergy: Carbon–Structure Optimization for Erosion-Resistant Barrier Formation in Cold Mollisols. Agriculture. 2025; 15(14):1465. https://doi.org/10.3390/agriculture15141465
Chicago/Turabian StyleLi, Tianxiao, Shunli Zheng, Zhaoxing Xiao, Qiang Fu, Fanxiang Meng, Mo Li, Dong Liu, and Qingyuan Liu. 2025. "Harnessing Bacillus subtilis–Moss Synergy: Carbon–Structure Optimization for Erosion-Resistant Barrier Formation in Cold Mollisols" Agriculture 15, no. 14: 1465. https://doi.org/10.3390/agriculture15141465
APA StyleLi, T., Zheng, S., Xiao, Z., Fu, Q., Meng, F., Li, M., Liu, D., & Liu, Q. (2025). Harnessing Bacillus subtilis–Moss Synergy: Carbon–Structure Optimization for Erosion-Resistant Barrier Formation in Cold Mollisols. Agriculture, 15(14), 1465. https://doi.org/10.3390/agriculture15141465