
Potential for Enhancing Forage Sorghum Yield and Yield Components in a Changing Pannonian Climate

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
- LA—leaf area (cm2);
- LL—leaf length (cm);
- LW—leaf width (cm);
3. Results
3.1. Sorghum Genotypes Showed Variability in Biomass Yield and Yield Components Responses to Climatic Factors in the Pannonian Region
3.2. Key Agronomic Traits Associated with Biomass Yield Improvement
3.3. Promising Hybrid Combinations for Biomass Production Under Challenging Climatic Conditions Are Identified Based on Combining Ability Effects
4. Discussion
4.1. Genotypic Variation in Biomass Yield and Its Climatic Adaptability in the Pannonian Region: Implications and Interpretation
4.2. Relevance of Key Agronomic Traits in Enhancing Biomass Yield
4.3. Combining Ability as a Tool for Identifying Climate-Resilient Biomass Hybrids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GFY | Green fodder yield |
| DMY | Dry matter yield |
| PH | Plant height |
| SD | Stem diameter |
| SDCP | Stem diameter at cutting point |
| IN | Internodes number |
| LN | Leaves number |
| LA | Leaf area |
| PL | Panicle length |
| NL | Neck length |
| DH | Number of days to heading (BBCH 51–55) |
| DSD | Number of days to soft-dough grain maturity stage (BBCH 85) |
| LA | Leaf area |
| LL | Leaf length |
| LW | Leaf width |
| GCA | General combining ability |
| SCA | Specific combining ability |
| SD | Standard deviation |
| CV | Coefficient of variation |
| SE | Standard error |
| df | Degrees of freedom |
| SCA_LA | Special combining ability for leaf area |
| SCA_PH | Special combining ability for plant height |
| SCA_LN | Special combining ability for leaf number |
| SCA_SD | Special combining ability for stem diameter |
| SCA_DMY | Special combining ability for dry matter yield |
| SCA_GFY | Special combining ability for green fodder yield |
References
- Bazié, D.; Dibala, C.I.; Kondombo, C.P.; Diao, M.; Konaté, K.; Sama, H.; Kayodé, A.P.P.; Dicko, M.H. Physicochemical and Nutritional Potential of Fifteen Sorghum Cultivars from Burkina Faso. Agriculture 2023, 13, 675. [Google Scholar] [CrossRef]
- Ananda, G.K.S.; Myrans, H.; Norton, S.L.; Gleadow, R.; Furtado, A.; Henry, R.J. Wild Sorghum as a Promising Resource for Crop Improvement. Front. Plant Sci. 2020, 11, 1108. [Google Scholar] [CrossRef] [PubMed]
- Faostat Statistics. Crop and Livestock Products—Statistics on Area Harvested of Sorghum and Average Yield of Grain in World, European Region and Serbia. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 1 April 2025).
- Soni, P.G.; Yadav, R.K.; Kumar, A.; Kumar, R.; Gajender, G.; Yadav, T.; Sharma, A.; Kushwaha, M. Effect of water quality regimes, irrigation schedules, RSC levels and neutralizer on physiological and biochemical behaviour of fodder sorghum (Sorghum bicolor). Indian J. Agric. Sci. 2020, 90, 1702–1707. [Google Scholar] [CrossRef]
- Rakshit, S.; Aruna, C.; Yadav, P.; Patroti, P.; Girish, G.; Ganapathy, K.N.; Ratnavathi, C.V.; Padmaja, P.G.; Bahadure, D.M. Stability for grain oil content in sorghum (Sorghum bicolor). Indian J. Agric. Sci. 2021, 91, 1636–1639. [Google Scholar] [CrossRef]
- Mahmood, A.; Ullah, H.; Ijaz, M.; Javaid, M.M.; Shahzad, A.N.; Honermeier, B. Evaluation of Sorghum Hybrids for Biomass and Biogas Production. Aust. J. Crop Sci. 2013, 7, 1456–1462. [Google Scholar]
- Rocha, J.; Oliveira, S.; Viana, C.M.; Ribeiro, A.I. Chapter 8—Climate change and its impacts on health, environment and economy. In One Health: Integrated Approach to 21st Century Challenges to Health; Prata, J.C., Ribeiro, A.I., Rocha-Santos, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 253–271. [Google Scholar] [CrossRef]
- Hossain, M.S.; Islam, M.N.; Rahman, M.M.; Mostofa, M.G.; Khan, M.A.R. Sorghum: A Prospective Crop for Climatic Vulnera-bility, Food and Nutritional Security. J. Agric. Food Res. 2022, 8, 100300. [Google Scholar] [CrossRef]
- Mohammed, A.; Misganaw, A. Modeling future climate change impacts on sorghum (Sorghum bicolor) production with best management options in Amhara Region, Ethiopia. CABI Agric. Biosci. 2022, 3, 22. [Google Scholar] [CrossRef]
- He, L.; Rosa, L. Solutions to agricultural green water scarcity under climate change. PNAS nexus 2023, 2, 117. [Google Scholar] [CrossRef]
- Mwamahonje, A.; Mdindikasi, Z.; Mchau, D.; Mwenda, E.; Sanga, D.; Garcia-Oliveira, A.L.; Ojiewo, C.O. Advances in Sorghum Improvement for Climate Resilience in the Global Arid and Semi-Arid Tropics: A Review. Agronomy 2024, 14, 3025. [Google Scholar] [CrossRef]
- Pataki, I.; Katić, S.; Mihailović, V.; Milić, D.; Vasiljević, S.; Mikić, A. Use of hybridization (F1) in forage sorghum (Sorghum bicolor (L.) Moench) breeding. Ratar. Povrt./Field Veg. Crops Res. 2010, 47, 225–230. [Google Scholar]
- Bhat, B.V. Chapter 11—Breeding Forage Sorghum. In Woodhead Publishing Series in Food Science, Technology and Nutrition; Aruna, C., Visarada, K.B.R.S., Venkatesh Bhat, B., Tonapi, V.A., Eds.; Woodhead Publishing: Delhi, India, 2019; pp. 175–191. [Google Scholar] [CrossRef]
- Jain, S.K.; Patel, P.R. Principal component and cluster analysis in sorghum (Sorghum bicolor (L.) Moench). Forage Res. 2016, 42, 90–95. [Google Scholar]
- Kavithamani, D.; Yuvaraja, A.; Selvi, B. Principal component analysis and grouping of sorghum (Sorghum bicolor L. Moench) gene pool for genetic diversity. Electron. J. Plant Breed. 2019, 10, 1426–1434. [Google Scholar] [CrossRef]
- Lee, S.; Choi, Y.-M.; Shin, M.-J.; Yoon, H.; Wang, X.; Lee, Y.; Yi, J.; Desta, K.T. Agro-Morphological and Biochemical Characterization of Korean Sorghum (Sorghum bicolor (L.) Moench) Landraces. Agronomy 2022, 12, 2898. [Google Scholar] [CrossRef]
- Navyashree, R.; Mummigatti, U.V.; Nethra, P.; Basavaraj, B.; Hanamaratti, N.G. Correlation and principal component analysis of morphophysiological and biochemical traits in sorghum under drought stress. Int. J. Res. Agron. 2024, 7, 15–23. [Google Scholar] [CrossRef]
- Gebre, W.; Mekbib, F.; Tirfessa, A.; Bekele, A. Phenotypic diversity of sorghum [Sorghum bicolor (L.) Moench] genotypes based on qualitative and quantitative traits. Heliyon 2025, 11, 42378. [Google Scholar] [CrossRef]
- Pataki, I. Combining Abilities and Mode of Inheritance of Yield Components of F1 Forage Sorghum (Sorghum bicolor L. Moench) Hybrids Created by Crossing Grain Sorghum and Sudan Grass. Ph.D. Thesis, Faculty of Agriculture, University of Novi Sad, Novi Sad, Serbia, 2010. [Google Scholar]
- Bangarwa, K.S.; Grewal, R.P.S.; Lodhi, G.P. A single recessive gene for stem sweetness in sorghum. Euphytica 1987, 36, 225–226. [Google Scholar] [CrossRef]
- Jain, S.K.; Elangovan, M.; Patel, N.V. Genetic and environmental variability in dual sorghum [Sorghum bicolor (L.) Moench] for yield and related traits. Forage Res. 2009, 34, 201–204. [Google Scholar]
- House, L.R. A Guide to Sorghum Breeding, 2nd ed.; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, India, 1985. [Google Scholar]
- Pataki, I. Sorghum and Sudan grass as fodder and energy plants. In Proceedings of the 45th Conference of Agronomists of Serbia, Zlatibor, Serbia, 30 January–5 February 2011. [Google Scholar]
- Kikindonov, T.; Kikindonov, G. Initial development and growth of biomass and dry matter in Sudan grass and Sorghum × Sudan grass hybrids. Discourse J. Agric. Food Sci. 2014, 2, 149–151. [Google Scholar]
- Soil Texture Calculator. Natural Resources Conservation Service. U.S. Department of Agriculture. Available online: https://www.nrcs.usda.gov/resources/education-and-teaching-materials/soil-texture-calculator (accessed on 22 June 2025).
- Republic Hydrometeorological Institute of Serbia. Available online: http://www.hidmet.gov.rs/index_eng.php (accessed on 2 April 2025).
- UPOV. International Union for the Protection of New Varieties of Plants, Geneva, Sorghum Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability. Available online: https://www.upov.int/edocs/tgdocs/en/tg122.pdf (accessed on 15 October 2018).
- Krishnamurthy, K.; Jagannath, M.K.; Rajashekara, B.G.; Raghunatha, G. Estimation of Leaf Area in Grain Sorghum from Single Leaf Measurements. Agron. J. 1974, 66, 544–545. [Google Scholar] [CrossRef]
- Atumo, T.T.; Mengesha, G.G. Growth and yield performance of sorghum (Sorghum bicolor L.) crop under anthracnose stress in dryland crop-livestock farming system. Exp. Results 2022, 3, e14. [Google Scholar] [CrossRef]
- Begna, T.; Birhan, T.; Tadesse, T. Gene Action and Combining Ability Estimation in Elite Sorghum [Sorghum bicolor (L.) Moench] Inbred Lines Under Dry Lowland Areas of Ethiopia. PREPRINT (Version 1). Research Square 2023. [Google Scholar] [CrossRef]
- Lancashire, P.D.; Bleiholder, H.; Boom, T.v.d.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Ingle, K.P.; Gahukar, S.J.; Khelurkar, V.C.; Ghorade, R.B.; Kalpande, V.V.; Jadhav, P.V.; Moharil, M.P. Heterosis and Combining Ability for Grain Yield Trait in Rabi Sorghum [Sorghum bicolor (L.) Moench] Using Line × Tester Mating Design. Int. J. Curr. Microbiol. Appl. Sci. 2018, 32, 1925–1934. [Google Scholar]
- XLSTAT Statistical and Data Analysis Solution. 2022. Available online: https://www.xlstat.com (accessed on 17 March 2025).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 24 March 2025).
- Abbass, K.; Qasim, M.Z.; Song, H.; Murshed, M.; Mahmood, H.; Younis, I. A Review of the Global Climate Change Impacts, Adaptation, and Sustainable Mitigation Measures. Environ. Sci. Pollut. Res. 2022, 29, 42539–42559. [Google Scholar] [CrossRef] [PubMed]
- Lana, M.A.; Eulenstein, F.; Schlindwein, S.L.; Graef, F.; Sieber, S.; Hertwig Bittencourt, H. Yield stability and lower susceptibility to abiotic stresses of improved open-pollinated and hybrid maize cultivars. Agron. Sustain. Dev. 2017, 37, 30. [Google Scholar] [CrossRef]
- Farhadi, A.; Paknejad, F.; Golzardi, F.; Ilkaee, M.N.; Aghayari, F. Effects of Limited Irrigation and Nitrogen Rate on the Herbage Yield, Water Productivity, and Nutritive Value of Sorghum Silage. Commun. Soil Sci. Plant Anal. 2022, 53, 576–589. [Google Scholar] [CrossRef]
- Ghalkhani, A.; Golzardi, F.; Khazaei, A.; Mahrokh, A.; Illés, A.; Bojtor, C.; Mousavi, S.M.N.; Széles, A. Irrigation Management Strategies to Enhance Forage Yield, Feed Value, and Water-Use Efficiency of Sorghum Cultivars. Plants 2023, 12, 2154. [Google Scholar] [CrossRef]
- Pourali, S.; Aghayari, F.; Ardakani, M.R.; Paknejad, F.; Golzardi, F. Benefits from Intercropped Forage Sorghum–Red Clover Under Drought Stress Conditions. Gesunde Pflanz. 2023, 75, 1769–1780. [Google Scholar] [CrossRef]
- Ngidi, A.; Shimelis, H.; Abady, S.; Figlan, S.; Chaplot, V. Response of Sorghum Bicolor Genotypes for Yield and Yield Components and Organic Carbon Storage in the Shoot and Root Systems. Sci. Rep. 2024, 14, 9499. [Google Scholar] [CrossRef]
- Rosenow, D.T.; Clark, L.E. Drought and Lodging Resistance for Quality Sorghum Crop. In Proceedings of the 50th Annual Corn and Sorghum Industry Research Conference, Chicago, IL, USA, 6–7 December 1995; pp. 82–97. [Google Scholar]
- Rosenow, D.T.; Ejeta, G.; Clark, L.E.; Gilbert, M.L.; Henzell, R.G.; Borrell, A.K.; Muchow, R.C. Breeding for Pre- and Post-flowering Drought Stress Resistance in Sorghum. In Proceedings of the International Conference on Genetic Improvement of Sorghum and Pearl Millet, Lubbock, TX, USA, 22–27 September 1996; pp. 400–411. [Google Scholar]
- Abraha, T.; Githiri, S.M.; Kasili, R.; Araia, W.; Nyende, A.B. Genetic Variation among Sorghum (Sorghum bicolor L. Moench) Landraces from Eritrea under Post-Flowering Drought Stress Conditions. Am. J. Plant Sci. 2015, 6, 1410–1424. [Google Scholar] [CrossRef]
- Maiti, R.K. Sorghum Science; Science Publishers: Beirut, Lebanon, 1996. [Google Scholar]
- Takanashi, H. Genetic Control of Morphological Traits Useful for Improving Sorghum. Breed. Sci. 2023, 73, 57–69. [Google Scholar] [CrossRef]
- Araus, J.L.; Slafer, G.A.; Reynolds, M.P.; Royo, C. Plant breeding and drought in C3 cereals: What should we breed for? Ann. Bot. 2002, 89, 925–940. [Google Scholar] [CrossRef]
- Mutava, R.N.; Prasad, P.V.V.; Tuinstra, M.R.; Kofoid, K.D.; Yu, J. Characterization of sorghum genotypes for traits related to drought tolerance. Field Crops Res. 2011, 123, 10–18. [Google Scholar] [CrossRef]
- Bazaluk, O.; Havrysh, V.; Fedorchuk, M.; Nitsenko, V. Energy Assessment of Sorghum Cultivation in Southern Ukraine. Agriculture 2021, 11, 695. [Google Scholar] [CrossRef]
- George, T.T.; Obilana, A.O.; Oyenihi, A.B.; Obilana, A.B.; Akamo, D.O.; Awika, J.M. Trends and Progress in Sorghum Research over Two Decades, and Implications for Global Food Security. S. Afr. J. Bot. 2022, 151, 960–969. [Google Scholar] [CrossRef]
- Đukanović, L.; Đuričić, M.; Tošković, S.; Marić, V.; Đukanović, D.; Poštić, D.; Štrbanović, R. Effect of Forage Sorghum Hybrids on Green Fodder Yeld. J. Process. Energy Agric. 2016, 20, 176–178. [Google Scholar]
- Peng, J.; Kim, M.; Sung, K. Yield Prediction Modeling for Sorghum–Sudangrass Hybrid Based on Climatic, Soil, and Cultivar Data in the Republic of Korea. Agriculture 2020, 10, 137. [Google Scholar] [CrossRef]
- Prażak, R. Prospects for Sorghum Cultivation in Poland. Acta Agrobot. 2016, 69, 1661. [Google Scholar] [CrossRef]
- Negri, L.; Bosi, S.; Fakaros, A.; Ventura, F.; Magagnoli, S.; Masetti, A.; Lami, F.; Oliveti, G.; Poggi, G.M.; Dinelli, G. Millets and Sorghum as Promising Alternatives to Maize for Enhancing Climate Change Adaptation Strategies in the Mediterranean Basin. Field Crops Res. 2024, 318, 109563. [Google Scholar] [CrossRef]
- Hajjarpoor, A.; Eltigani, A.; Seiler, C.; Matros, A.; Kottmann, L.; Balko, C.; Windpassinger, S.; Goldbach, J.; Eder, J.; Rahman, A.S.; et al. Germany-wide suitability analysis of sorghum cultivation for climate change mitigation. In Proceedings of the Digital Tools, Big Data, Modeling and Sensing Methods for Sustainable and Climate Smart Crop and Grassland Systems, Göttingen, Germany, 4–6 October 2023; Volume 33, pp. 95–96. [Google Scholar]
- Schaffasz, A.; Windpassinger, S.; Friedt, W.; Snowdon, R.; Wittkop, B. Sorghum as a Novel Crop for Central Europe: Using a Broad Diversity Set to Dissect Temperate-Adaptation. Agronomy 2019, 9, 535. [Google Scholar] [CrossRef]
- Eniola, O.A.; Odiyi, A.C.; Fayeun, L.S.; Mogaji, B.O.; Obilana, A.B. Yield and Yield Components of Grain Sorghum (Sorghum Bicolor (L.) Moench) Hybrid in Akure Agro-Ecology. In Proceedings of the 4th Annual Conference of Association of Seed Scientists of Nigeria (Assn), Akure, Nigeria, 24–28 June 2018; pp. 134–145. [Google Scholar]
- Osman, A.; Abd El-Wahab, A.; Ahmed, M.F.E.; Buschmann, M.; Visscher, C.; Hartung, C.B.; Lingens, J.B. Nutrient Composition and In Vitro Fermentation Characteristics of Sorghum Depending on Variety and Year of Cultivation in Northern Italy. Foods 2022, 11, 3255. [Google Scholar] [CrossRef] [PubMed]
- Atique-ur-Rehman; Qamar, R.; Altaf, M.M.; Alwahibi, M.S.; Al-Yahyai, R.; Hussain, M. Phosphorus and Potassium Application Improves Fodder Yield and Quality of Sorghum in Aridisol under Diverse Climatic Conditions. Agriculture 2022, 12, 593. [Google Scholar] [CrossRef]
- Elangovan, M.; Kiran Babu, P.; Seetharama, N.; Patil, J.V. Genetic Diversity and Heritability Characters Associated in Sweet Sorghum [Sorghum bicolor (L.) Moench]. Sugar Tech 2014, 16, 200–210. [Google Scholar] [CrossRef]
- Lekgari, A.; Dweikat, I. Assessment of Genetic Variability of 142 Sweet Sorghum Germplasm of Diverse Origin with Molecular and Morphological Markers. Open J. Ecol. 2014, 4, 371–393. [Google Scholar] [CrossRef]
- Regassa, T.H.; Wortmann, C.S. Sweet sorghum as a bioenergy crop: Literature review. Biomass Bioenergy 2014, 64, 348–355. [Google Scholar] [CrossRef]
- Chaudhary, D.P.; Saini, R.K.; Maurya, B.K.; Sharma, M.; Kumar, R.; Sen, R.; Singh, S.K. Study of Genetic Variability and Fodder Yield Components in Forage Sorghum (Sorghum bicolor L. Moench). Bull. Environ. Pharmacol. Life Sci. 2018, 7, 5–9. [Google Scholar]
- Mangena, P.; Shimelis, H.; Laing, M. Characterisation of Sweet Stem Sorghum Genotypes for Bio-Ethanol Production. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2018, 68, 323–333. [Google Scholar] [CrossRef]
- Gratani, L. Plant Phenotypic Plasticity in Response to Environmental Factors. Adv. Bot. 2014, 2014, 208747. [Google Scholar] [CrossRef]
- Sinha, S.; Kumaravadivel, N. Understanding Genetic Diversity of Sorghum Using Quantitative Traits. Scientifica 2016, 2016, 3075023. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.; Are, A.K.; Bhavanasi, R.; Munghate, R.S.; Kavi Kishor, P.B.; Sharma, H.C. Quantitative genetic analysis of agronomic and morphological traits in sorghum, Sorghum bicolor. Front. Plant Sci. 2015, 6, 945. [Google Scholar] [CrossRef]
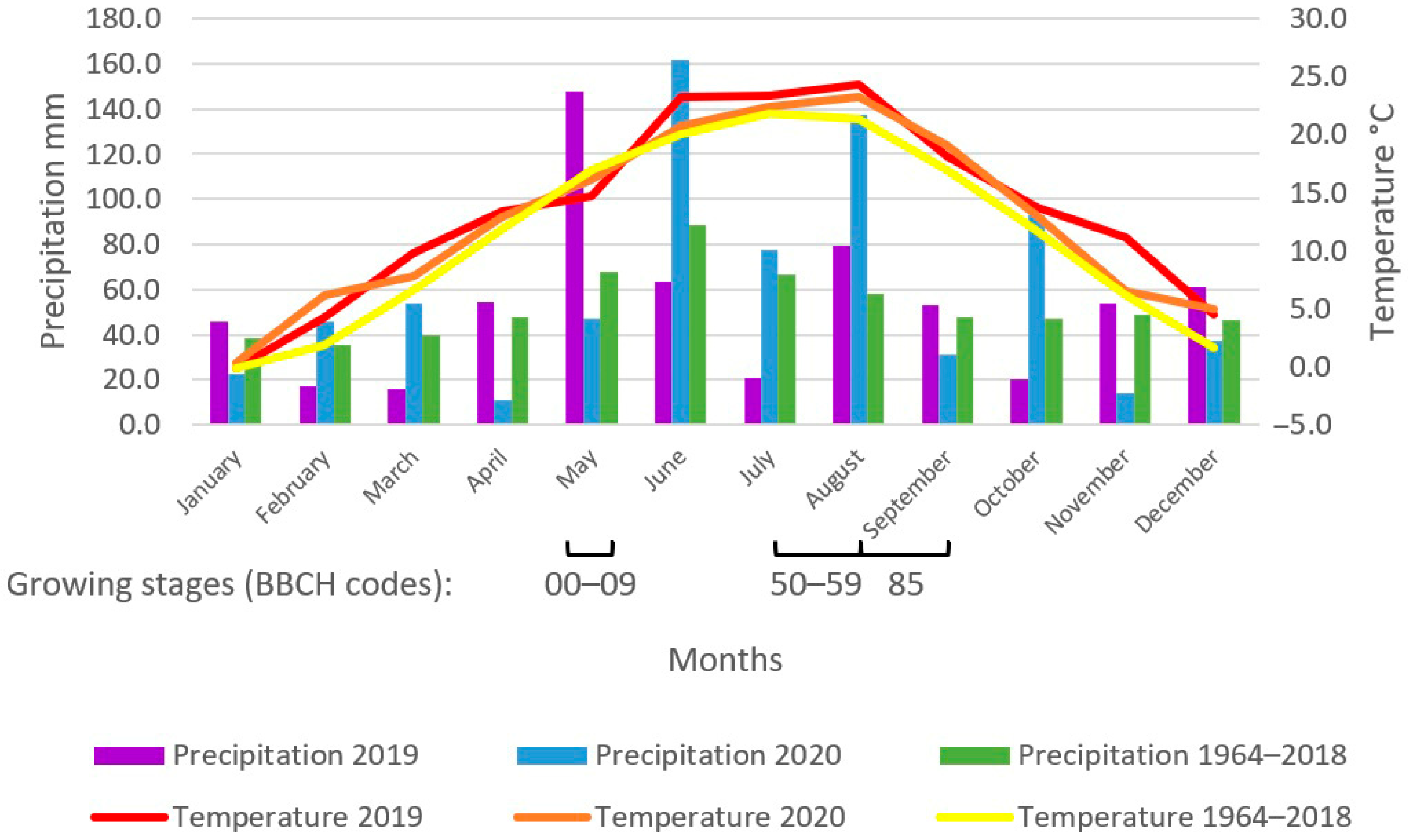

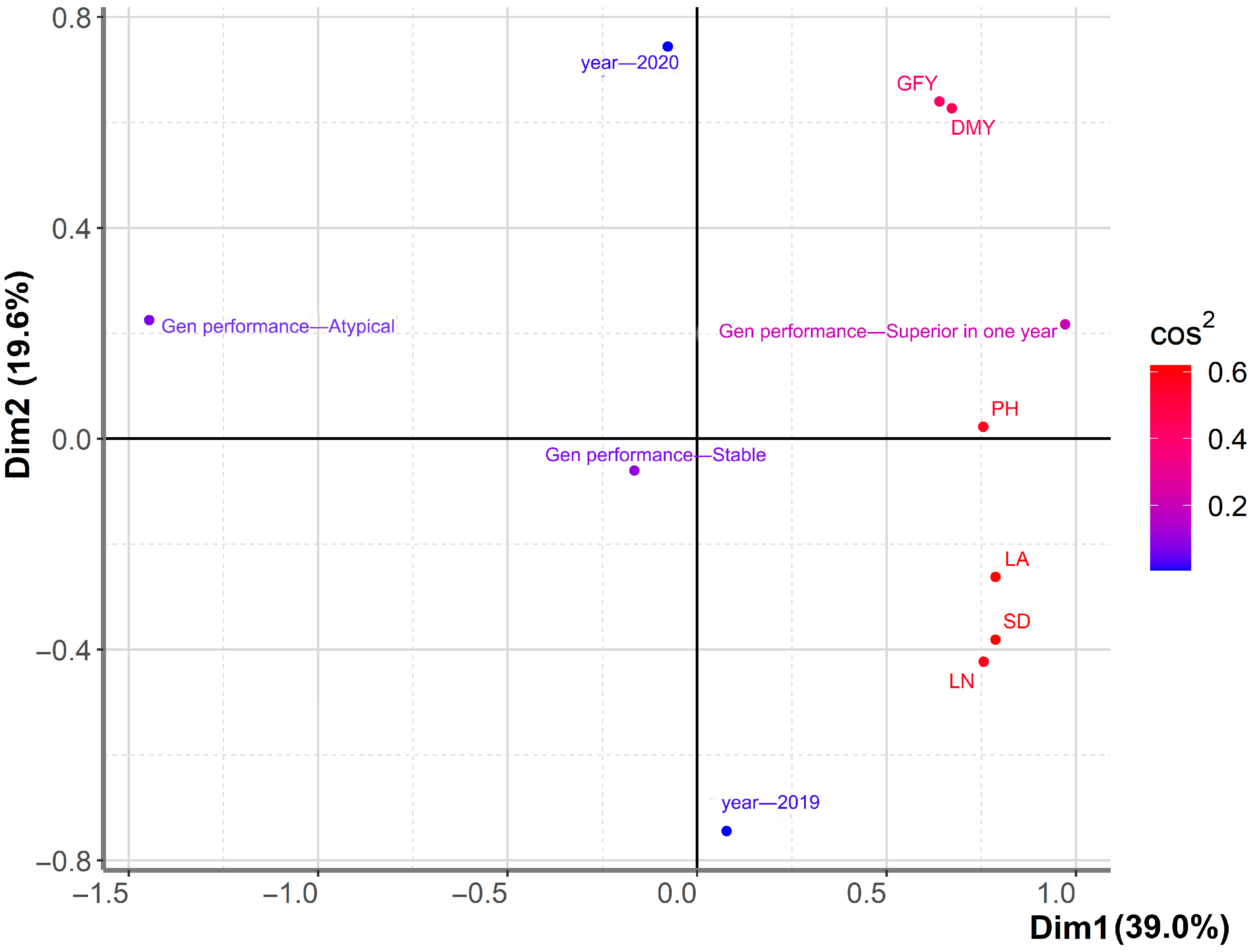


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2019 | 2020 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | SD | CV (%) | SE | Min | Max | Mean | SD | CV (%) | SE | |
| DH | 47.00 | 79.00 | 56.72 | 6.30 | 11.11 | 0.47 | 57.00 | 94.00 | 70.39 | 7.34 | 10.43 | 0.55 |
| DSD | 15.00 | 33.00 | 25.51 | 3.40 | 13.33 | 0.25 | 28.00 | 36.00 | 32.78 | 1.24 | 3.77 | 0.09 |
| PH | 66.23 | 372.30 | 237.16 | 55.75 | 23.51 | 4.16 | 82.80 | 342.50 | 241.21 | 45.56 | 18.89 | 3.40 |
| IN | 2.50 | 12.80 | 8.23 | 1.57 | 19.04 | 0.12 | 4.70 | 13.00 | 8.65 | 1.41 | 16.28 | 0.11 |
| SDCP | 4.70 | 21.93 | 13.26 | 2.72 | 20.48 | 0.20 | 6.27 | 25.15 | 13.86 | 2.79 | 20.15 | 0.21 |
| SD | 4.79 | 17.56 | 10.43 | 2.17 | 20.77 | 0.16 | 4.22 | 15.74 | 9.41 | 1.83 | 19.44 | 0.14 |
| LN | 4.90 | 14.10 | 8.15 | 1.83 | 22.46 | 0.14 | 3.20 | 11.90 | 6.99 | 1.28 | 18.36 | 0.10 |
| LA | 145.26 | 522.94 | 291.47 | 66.05 | 22.66 | 4.92 | 138.63 | 500.03 | 278.44 | 64.55 | 23.18 | 4.81 |
| PL | 16.71 | 46.16 | 28.41 | 6.12 | 21.54 | 0.46 | 12.40 | 42.00 | 27.13 | 4.98 | 18.35 | 0.37 |
| NL | 0.00 | 22.87 | 9.08 | 5.31 | 58.48 | 0.40 | 0.00 | 13.50 | 5.22 | 2.93 | 56.02 | 0.22 |
| GFY | 24.08 | 114.54 | 51.95 | 16.23 | 31.25 | 1.21 | 12.05 | 137.70 | 61.33 | 23.38 | 38.11 | 1.74 |
| DMY | 6.17 | 47.26 | 18.78 | 7.71 | 41.06 | 0.58 | 2.78 | 79.00 | 23.97 | 12.42 | 51.81 | 0.93 |
| Source of Variation | df | PH | SD | LN | LA | GFY | DMY |
|---|---|---|---|---|---|---|---|
| Genotype group | 2 | ** | ** | ** | ** | ** | ** |
| Year | 1 | n.s. | ** | ** | n.s. | n.s. | n.s. |
| Genotype group × Year | 2 | n.s. | n.s. | n.s. | n.s. | ** | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolapčev Rakić, A.; Prodanović, S.; Sikora, V.; Vasiljević, S.; Župunski, V.; Jevtić, R.; Uhlarik, A. Potential for Enhancing Forage Sorghum Yield and Yield Components in a Changing Pannonian Climate. Agriculture 2025, 15, 1439. https://doi.org/10.3390/agriculture15131439
Dolapčev Rakić A, Prodanović S, Sikora V, Vasiljević S, Župunski V, Jevtić R, Uhlarik A. Potential for Enhancing Forage Sorghum Yield and Yield Components in a Changing Pannonian Climate. Agriculture. 2025; 15(13):1439. https://doi.org/10.3390/agriculture15131439
Chicago/Turabian StyleDolapčev Rakić, Anja, Slaven Prodanović, Vladimir Sikora, Sanja Vasiljević, Vesna Župunski, Radivoje Jevtić, and Ana Uhlarik. 2025. "Potential for Enhancing Forage Sorghum Yield and Yield Components in a Changing Pannonian Climate" Agriculture 15, no. 13: 1439. https://doi.org/10.3390/agriculture15131439
APA StyleDolapčev Rakić, A., Prodanović, S., Sikora, V., Vasiljević, S., Župunski, V., Jevtić, R., & Uhlarik, A. (2025). Potential for Enhancing Forage Sorghum Yield and Yield Components in a Changing Pannonian Climate. Agriculture, 15(13), 1439. https://doi.org/10.3390/agriculture15131439