Study on Toxic Substances in Astragalus adsurgens Infected with Alternaria gansuense


Abstract
1. Introduction
2. Materials and Methods
2.1. Overview and Management of Experimental Site
2.2. Collection and Preservation of Plant Materials
2.3. Digestion of Grass Samples
2.4. Isolation and Cultivation of A. gansuense
2.5. Determination of Selenium Content
2.6. Determination of Mineral Elements
2.7. Preparation for Determination of Swainsonine
2.8. Sample Processing
2.9. Drawing and Measurement of the Standard Curve for Swainsonine
2.10. Statistical Analysis
3. Results
3.1. Determination Results of Selenium Content
3.2. Standard Curves for Other Mineral Elements
3.3. Element Content
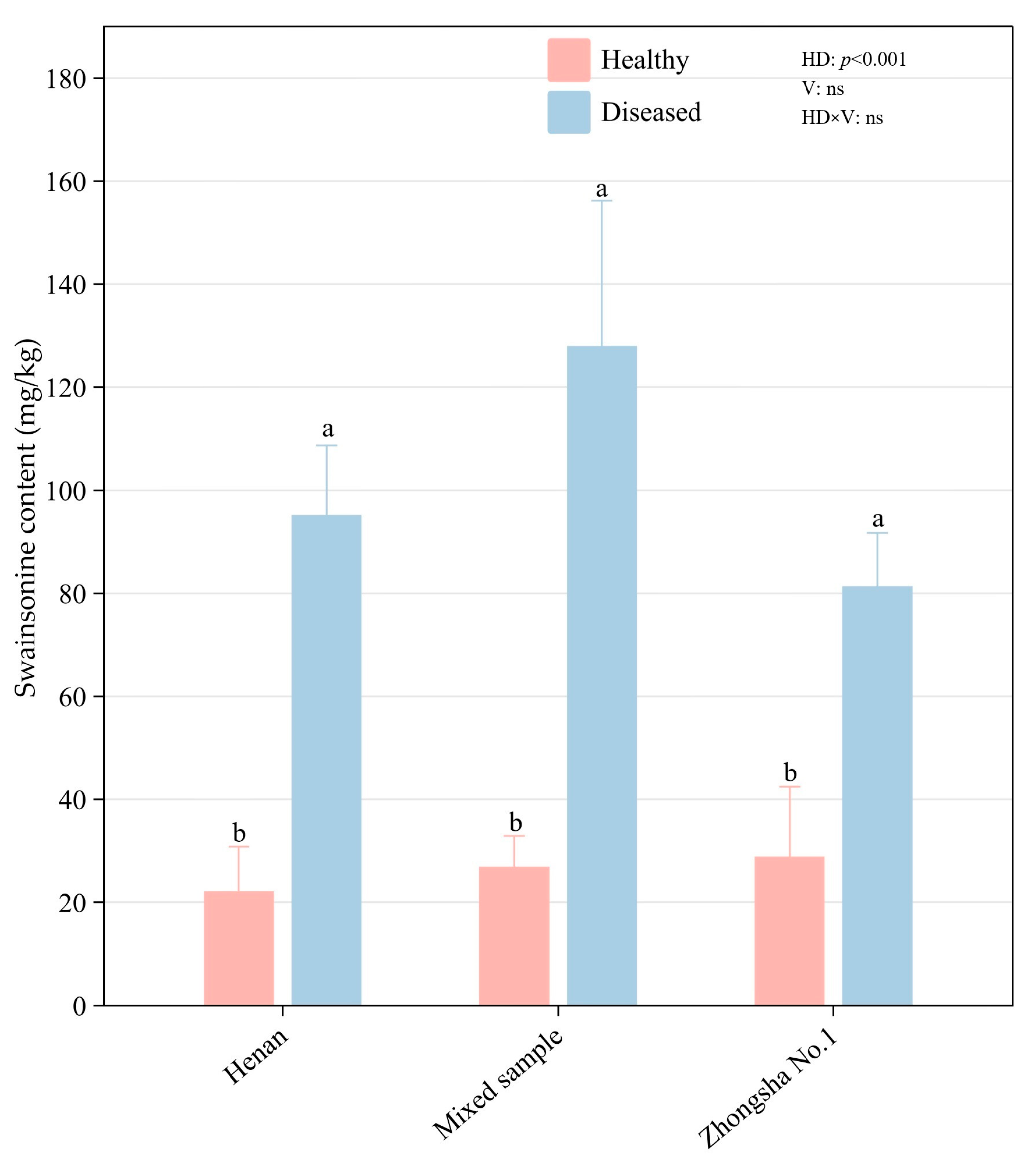
3.4. Results of Swainsonine Test
3.5. The Correlation Between Swainsonineand Elements
4. Discussion
4.1. Changes in Selenium Content of Diseased Plants and Its Impact on Livestock Feeding
4.2. Alterations in the Concentrations of Other Mineral Elements in Diseased A. adsurgens
4.3. The Influence of Changes in Swainsonine Content of Diseased Astragalus adsurgens on Livestock and Its Correlation with Mineral Elements
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, Q.Z.; Gui, R.; Na, R.S.; Nada, M.D.; Zhai, Z.H. Study on productivity of Astragalus adsurgens Pall. with different growth periods. Grassl. China 1999, 5, 30–35. [Google Scholar]
- Wang, J.; Lei, Z.Y.; Feng, X.Q. Gas Chromatographic Determination of Trace 3-Nitropropionic Acid in Astragalus adsurgens. Grassl. China 1992, 3, 69–72. [Google Scholar]
- Su, S.F. Revisiting the Forage Utilization Issues of Astragalus adsurgens. Pratacultural Sci. 1987, 2, 3–9. [Google Scholar]
- Cook, D.; Ralphs, M.; Welch, K.; Stegelmeier, B. Locoweed poisoning in livestock. Rangelands 2009, 31, 16–21. [Google Scholar] [CrossRef]
- Zhang, G.G.; Dong, S.T.; Yang, Z.B.; Yang, W.R.; Jiang, S.Z. Experiment on Feeding Small-tailed Han Sheep with Astragalus adsurgens. Pratacultural Sci. 2008, 3, 74–77. [Google Scholar]
- Li, Y.Z.; Nan, Z.B. Symptomology and etiology of a new disease, yellow stunt, and root rot of standing milkvetch caused by Embellisia sp. in Northern China. Mycopathologia 2007, 163, 327–334. [Google Scholar] [CrossRef]
- Li, Y.Z.; Nan, Z.B.; Hou, F.J. The roles of an Embellisia sp. causing yellow stunt and root rot of Astragalus adsurgens and other fungi in the decline of legume pastures in northern China. Australas. Plant Pathol. 2007, 36, 397–402. [Google Scholar] [CrossRef]
- Liu, J.L. Study on Molecular Biology of Embellisia astragali. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2016. [Google Scholar]
- Li, Y.Z.; Nan, Z.B. A new species, Embellisia astragali sp. nov., causing standing milk-vetch disease in China. Mycologia 2007, 99, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Liu, J.L.; Li, Y.Z.; Creamer, R. A re-examination of the taxonomic status of Embellisia astragali. Curr. Microbiol. 2016, 72, 404–409. [Google Scholar] [CrossRef]
- Li, Y.Z. Study of yellow stunt and root rot (Embellisia astragali sp. nov. Li & Nan) of Astragalus adsurgens. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2007. [Google Scholar]
- Li, Y.Z.; Creamer, R.; Baucom, D.; Nan, Z.B. Pathogenic Embellisia astragali on Astragalus adsurgens is very closely related to locoweed endophyte. Phytopathology 2011, 101, S102–S103. [Google Scholar]
- James, L.F.; Hartley, W.J.; Kampen, K.R.V. Syndromes of astragalus poisoning in livestock. J. Am. Vet. Med. Assoc. 1981, 178, 146–150. [Google Scholar] [CrossRef]
- Stegelmeier, B.L.; James, L.F.; Panter, K.E.; Gardner, D.R.; Pfister, J.A.; Ralphs, M.H.; Molyneux, R.J. Dose response of sheep poisoned with locoweed (Oxytropis sericea). J. Vet. Diagn. Investig. 1999, 11, 448–456. [Google Scholar] [CrossRef]
- Gardner, D.; Molyneux, R.; Ralphs, M. Analysis of swainsonine: Extraction methods, detection, and measurement in populations of locoweeds (Oxytropis spp.). J. Agric. Food Chem. 2001, 49, 4573–4580. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, R.; James, L. Loco intoxication: Indolizidine alkaloids of spotted locoweed (Astragalus lentiginosus). Science 1982, 216, 190–191. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, J.; Nakagawa, H.; Takahashi, M.; Kudo, T.; Kamiyama, N.; Sun, B.; Oshima, T.; Sato, Y.; Deguchi, K.; Todo, S. Swainsonine reduces 5-fluorouracil tolerance in the multistage resistance of colorectal cancer cell lines. Mol. Cancer 2007, 6, 58. [Google Scholar] [CrossRef]
- Cook, D.; Donzelli, B.G.G.; Creamer, R.; Baucom, D.L.; Gardner, D.R.; Pan, J.; Moore, N.; Jaromczyk, J.W.; Schardl, C.L. Swainsonine Biosynthesis Genes in Diverse Symbiotic and Pathogenic Fungi. G3-Genes Genomes Genet. 2017, 7, 1791–1797. [Google Scholar] [CrossRef]
- Smith, G.S.; Allred, K.W.; Kiehl, D.E. Swainsonine content of New Mexican locoweeds. J. Anim. Sci. 1992, 70, 405–407. [Google Scholar]
- O”Donnell, J.; Dickinson, C.H. Pathogenicity of Alternaria and Cladosporium isolates on Phaseolus. Trans. Br. Mycol. Soc. 1980, 74, 335–342. [Google Scholar] [CrossRef]
- Cui, Z.H.; Zhai, X.H.; Li, Q.F.; Wang, J.H.; Geng, G.X. Determination of Swainsonine in Astragalus locoweed by internal standard gas chromatography. J. Northwest Sci-Tech Univ. Agric. For. 2008, 36, 17–21. [Google Scholar] [CrossRef]
- Noor, A.I.; Neyaz, M.; Cook, D.; Creamer, R. Molecular Characterization of a Fungal Ketide Synthase Gene Among Swainsonine-Producing Alternaria Species in the USA. Curr. Microbiol. 2020, 77, 2554–2563. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.C.; James, L.F. Toxicity of nitro-containing Astragalus to sheep and chicks. J. Range Manag. 1975, 28, 260–263. [Google Scholar] [CrossRef]
- Wu, W.H. Plant Biology, 2nd ed.; Science Press: Beijing, China, 2008. [Google Scholar]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Lazar, T.; Zeiger, E. Plant Physiology, 5th ed.; Science Press: Beijing, China, 2015. [Google Scholar]
- Guo, Y.Y.; Xu, K.; Wang, X.F.; Zhu, Y.H. Research progress on mineral nutrition and plant disease mechanism. J. Gansu Agric. Univ. 2003, 4, 385–393. [Google Scholar]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Science Press: Beijing, China, 2013. [Google Scholar]
- Tian, Y.Q.; Huang, L.P.; Zhang, Z. Research Progress of the Relationship Between Mineral Nutrients Deficiency or Imbalance and Plant Disease. Chin. Agric. Sci. Bull. 2016, 32, 174–176. [Google Scholar]
- Fidanza, M.A.; Dernoeden, P.H. Interaction of nitrogen source, application timing, and fungicide on Rhizoctonia blight in ryegrass. HortScience 1996, 31, 389–392. [Google Scholar] [CrossRef]
- Bhaskar, C.V.; Rao, G.R.; Reddy, K.B. Effect of nitrogen and potassium nutrition on sheath rot incidence and phenol content in rice (Oryz asativa L.). Indian J. Plant Physiol. 2001, 16, 254–257. [Google Scholar]
- Sharma, S.R.; Kolte, S.J. Effect of soil-applied NPK fertilizers on severity of black spot disease (Alternaria brassicae) and yield of oilseed rape. Plant Soil 1994, 167, 313–320. [Google Scholar] [CrossRef]
- Liu, X.Y.; He, P.; Jin, J.Y. Advances in effect of potassium nutrition on plant disease resistance and its mechanism. Plant Nutr. Fertil. Sci. 2006, 12, 445–450. [Google Scholar] [CrossRef]
- Mu, K.G.; Zhao, X.Q.; Li, J.Q.; Liu, X.L. Progressing on the Relation between Mineral Nutrients and Plant Disease. J. China Agric. Univ. 2000, 5, 84–90. [Google Scholar]
- Conway, W.S.; Sams, C.E.; Kelman, A. Enhancing the natural resistance of plant tissues to postharvest diseases through calcium applications. HortScience 1994, 29, 66–69. [Google Scholar] [CrossRef]
- Huber, D.M.; Jones, J.B. The role of magnesium in plant disease. Plant Soil 2013, 368, 73–85. [Google Scholar] [CrossRef]
- Bhargava, A.K.; Singh, R.D. Effect of Nitrogenous Fertilizers and Trace Elements on the Severity of Alternaria Blight of Bottle Gound. Ann. Arid. Zone 1992, 31, 63–64. [Google Scholar]
- Gadi, B.; Jeffrey, G. Copper as a biocidal tool. Curr. Med. Chem. 2005, 12, 2163–2175. [Google Scholar] [CrossRef]
- Sharma, A.K.; Sawant, I.S.; Sawant, S.D.; Saha, S.; Kadam, P.; Somkuwar, R.G. Aqueous chlorine dioxide for the management of powdery mildew vis-a-vis maintaining quality of grapes and raisins. J. Eco-Friendly Agric. 2017, 12, 59–64. [Google Scholar]
- Ehret, D.L.; Utkhede, R.S.; Frey, B.; Menzies, J.G.; Bogdanoff, C. Foliar applications of fertilizer salts inhibit powdery mildew on tomato. Can. J. Plant Pathol. 2002, 24, 437–444. [Google Scholar] [CrossRef]
- Zeng, C.Y. Study on the Resistance Mechanism of Nine Astragalus adsurgens Varieties to Yellow Stunt Root and Rot and Comprehensive Evaluation for Germplasm Characteristics. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2016. [Google Scholar]
- Sa, F.; Vong, W.T.; Chan, T.M.; Lam, C.W.K. Determination of heavy metals in proprietary Chinese medicines by inductively coupled plasma–mass spectrometry. Clin. Chim. Acta 2010, 411, 909. [Google Scholar] [CrossRef]
- Chamkouri, N.; Torabpour, M. Microwave assisted digestion coupled with inductively coupled plasma optical emission spectrometry for determining element concentrations in halophytes. J. Basic Res. Med. Sci. 2015, 2, 49–56. [Google Scholar]
- Shi, M. The Effects of Yellow Stunt and Root Rot of Astragalus adsurgens on Swainsonine Content in Plants and Evaluation on New Disease Resistant Germplasms. Master’s Thesis, Lanzhou University, Lanzhou, China, 2019. [Google Scholar]
- Xu, X.L. Determination of Trace Heavy Metals in Porphyra by Microwave Digestion-ICP-MS Method. Zhejiang Chem. Ind. 2013, 44, 37–39. [Google Scholar]
- Leite, C.C.; De, J.A.; Potes, M.L.; Vieira, M.A.; Samios, D.; Silva, M.M. Direct Determination of Cd, Co, Cu, Fe, Mn, Na, Ni, Pb, and Zn in Ethanol Fuel by High-Resolution Continuum Source Flame Atomic Absorption Spectrometry. Energy Fuels 2015, 29, 7358–7363. [Google Scholar] [CrossRef]
- Stendig, L.G.; Penciner, J.; Rudy, N.; Wacker, W.E. Comparison of diluents for serum magnesium estimation by atomic absorption spectrophotometry. Magnesium 1984, 3, 50–56. [Google Scholar]
- Cui, Z.H. The study of Enzymatic Method and Gas chromatography for Determination of Swainsonine. Master’s Thesis, Northwest A&F University, Xi’an, China, 2008. [Google Scholar]
- Krasnodêbska, O.B.; Kowalska, J. Ultrasound-assisted acetic acid extraction of metals from soils. Chem. Anal. 2003, 48, 967–974. [Google Scholar]
- Huang, C.H.; Han, Q.; Guo, D.S. Extraction and Content Determination of Crude Polysaccharides from Wild Portulaca oleracea Grass Powder. Jiangsu Agric. Sci. 2011, 39, 398–400. [Google Scholar] [CrossRef]
- Wang, S.; Wu, S.Q.; Chen, G.Y.; Hu, J.J.; Zhang, L.; Ma, C.H. Determination of Swainsonine Content in Rabbit Serum by α-Mannosidase Method. China Herbiv. Sci. 2012, 32, 42–45. [Google Scholar]
- Panigati, M.; Falciola, L.; Mussini, P.; Beretta, G.; Facino, R.M. Determination of selenium in Italian rices by differential pulse cathodic stripping voltammetry. Food Chem. 2007, 105, 1091–1098. [Google Scholar] [CrossRef]
- Schwarz, K.; Bieri, J.G.; Briggs, G.M.; Scott, M.L. Prevention of Exudative Diathesis in Chicks by Factor 3 and Selenium. Proc. Soc. Exp. Biol. Med. 1957, 95, 621. [Google Scholar] [CrossRef]
- Zhao, G.Q.; Yin, Z.F.; Wei, L.H.; Wang, Y.J.; Yang, X.Y. The duality of selenium and its related health issues. J. Chengde Med. Coll. 1999, 1, 92–93. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Peng, Z.K.; Luo, Z.M. The multiple biological functions of selenium and its impact on human and animal health. J. Hunan Agric. Univ. 1997, 23, 294–300. [Google Scholar] [CrossRef]
- Rayman, M.P.; Margaret, P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- O’Toole, D.; Raisbeck, M.F. Pathology of experimentally induced chronic selenosis (alkali disease) in yearling cattle. J. Vet. Diagn. Investig. 1995, 7, 364–373. [Google Scholar] [CrossRef]
- Zhu, J.X.; Zhou, W.S.; Guo, S.H. Advances of Studies on Microelement Selenium in Plants. J. Anhui Agric. Sci. 2009, 37, 5844–5845. [Google Scholar] [CrossRef]
- Brown, T.A.; Shrift, A. Selenium: Toxicity and tolerance in higher plants. Biol. Rev. 1982, 57, 59–84. [Google Scholar] [CrossRef]
- Wu, J.; Liu, X.F.; Xu, H.S. Functions of Selenium in Plants. Plant Physiol. J. 1999, 35, 417–423. [Google Scholar] [CrossRef]
- Wang, J.H. Selenosis. Chin. J. Vet. Sci. 1993, 13, 401–408. [Google Scholar] [CrossRef]
- Shao, S.X.; Zheng, B.S.; Zhao, C.Z.; Yin, C.Q.; Lang, Y.B.; Zhang, A.L. Study on Selenium in Poisonous Oxytropis Plants (Locoweed) from the Hexi Corridor and Its Association with Livestock Poisoning. Adv. Earth Sci. 2004, 19, 502–505. [Google Scholar]
- Shao, S.X.; Zheng, B.S.; Wang, M.S.; Li, X.Y.; Liu, X.J.; Ling, H.W.; Luo, C. Environmental Geochemistry of Selenium in the Hexi Corridor Region and Investigation into the Causes of Livestock Poisoning by Toxic Plants. Acta Mineral. Sin. 2006, 26, 448–452. [Google Scholar] [CrossRef]
- Li, W.; Li, F.; Bi, D.; Wang, H.K.; Li, H.Y.; Zhou, S.B.; Zhao, Q.G.; Yin, X.B. Distribution and Speciation of Selenium in Alkaline Soils and Agricultural Products of Lanzhou. Soils 2012, 44, 632–638. [Google Scholar] [CrossRef]
- Xie, C.J.; Li, H.Y.; Wang, G.Y.; Wang, P. Analysis of Selenium Speciation in Soils and Key Agricultural Products from Lanzhou City. Gansu Sci. Technol. Inf. 2016, 45, 89–91. [Google Scholar]
- Shi, M.; Li, Y.Z. Alternaria gansuense, a Plant Systematic Fungal Pathogen Producing Swainsonine in Vivo and in Vitro. Curr. Microbiol. 2023, 80, 232. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Pfister, J.A. Swainsonine-containing plants and their relationship to endophytic fungi. J. Agric. Food Chem. 2014, 62, 7326–7334. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Lee, S.T.; Pfister, J.A.; Stonecipher, C.A.; Welsh, S.L. A swainsonine survey of North American Astragalus and Oxytropis taxa implicated as locoweeds. Toxicon 2016, 118, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.; Gardner, D.R.; Ralphs, M.H.; Pfister, J.A.; Welch, K.D.; Green, B.T. Swainsoninine concentrations and endophyte amounts of Undifilum oxytropis in different plant parts of Oxytropis sericea. J. Chem. Ecol. 2009, 35, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; He, B.X. A study on the toxicity of Oxytropis kansuonisis in sheep. Chin. J. Vet. Med. 1995, 21, 5–6. [Google Scholar]
- Wang, K.; Mo, C.H.; Zhao, B.Y.; Cao, G.R. Study on the Toxicity of Intermittent Feeding of Oxytropis kansuensis to Sheep. Anim. Husb. Vet. Med. 1998, 30, 29–30. [Google Scholar]
- Gu, B.Q.; Xue, D.M.; Cao, G.R.; Duan, D.X. Pathological Observations of Oxytropis kansuensis Poisoning in Goats. J. Anim. Sci. Vet. Med. 1991, 3, 9–11. [Google Scholar]
- Li, Q.F.; Wang, J.H.; Yuan, Y.; Qi, X.R.; Zhao, Y. Pathology of Oxytropis glacialis Poisoning in Goats. Chin. J. Vet. Sci. 2001, 21, 75–78. [Google Scholar] [CrossRef]
- Yang, B.; Nan, Z.B.; Li, Y.Z. Toxicity of standing milkvetch infected with Alternaria gansuense in white mice. Front. Vet. Sci. 2025, 11, 1477970. [Google Scholar] [CrossRef]
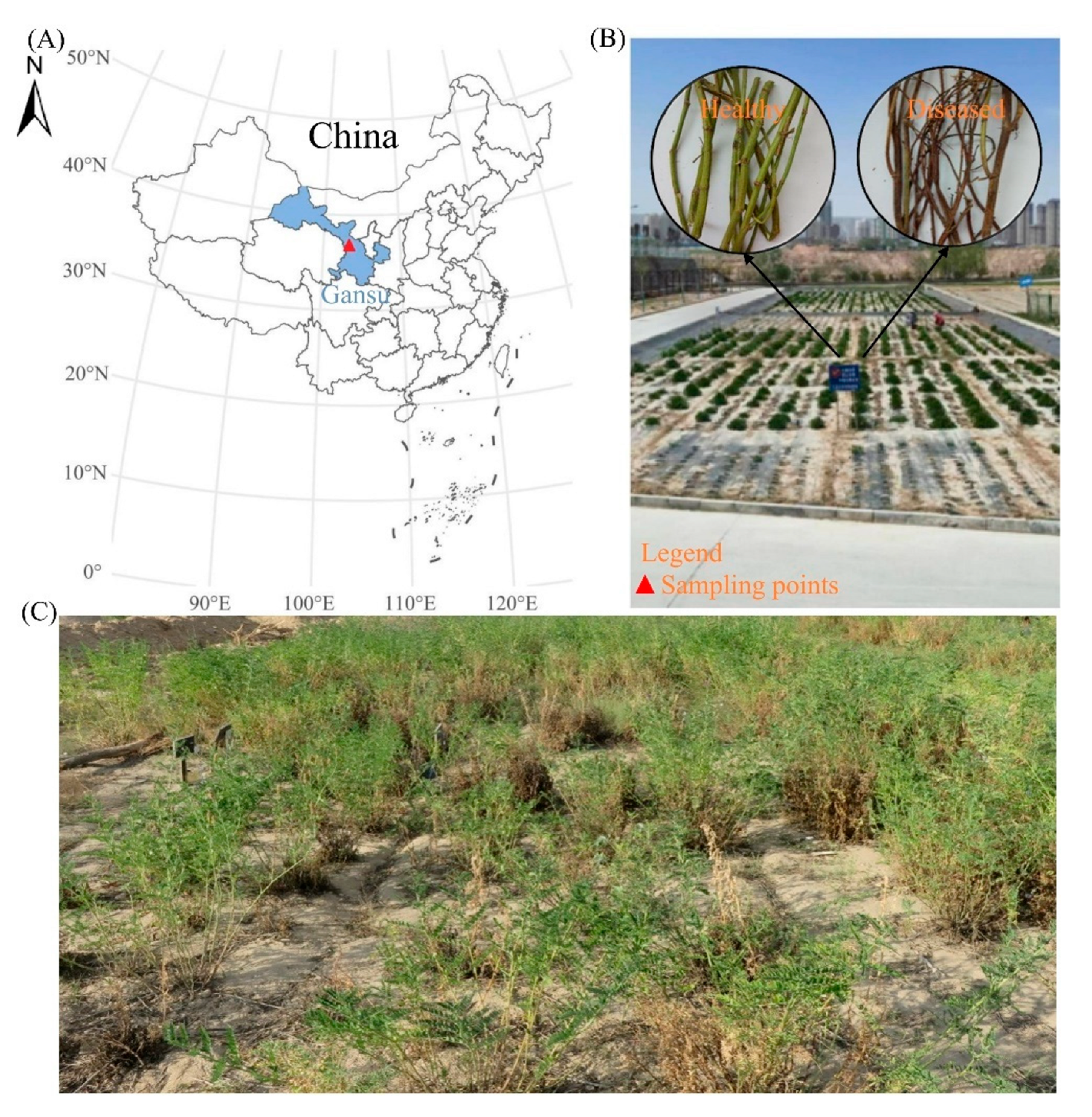




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Healthy Plants | Diseased Plant |
|---|---|---|
| Henan | 0.217 ± 0.038 ab | 1.109 ± 0.110 a |
| Liaoning Fuxin | 0.254 ± 0.068 ab | 0.960 ± 0.034 ab |
| Zahua | 0.367 ± 0.030 a | 0.798 ± 0.050 b |
| Zhongsha No.1 | 0.192 ± 0.022 b | 0.748 ± 0.055 b |
| Element | Concentration Gradient of Standard Solution (mg/L) | Regression Equation | Correlation Coefficient R2 |
|---|---|---|---|
| Cu | 0, 0.5, 1, 2, 3 | y = 21.629x − 0.12 | 0.9958 |
| Mn | 0, 1, 2, 3, 5 | y = 7.6736x − 0.0982 | 0.9976 |
| Fe | 0, 0.5, 1, 2, 3 | y = 15.498x − 0.0272 | 0.9992 |
| Zn | 0, 0.1, 0.2, 0.4, 0.5 | y = 2.9954x ± 0.0036 | 0.9991 |
| Ca | 0, 0.5, 1, 2, 4 | y = 25.905x − 0.0731 | 0.9975 |
| Na | 0, 0.1, 0.2, 0.6 | y = 2.2244x ± 0.0016 | 0.9991 |
| Mg | 0, 0.1, 0.2, 0.4, 0.6 | y = 1.2207x − 0.0182 | 0.9967 |
| Elements/Varieties | Henan | Zahua | Zhongsha No.1 | Liaoning Fuxin | |
|---|---|---|---|---|---|
| Mn | Healthy | 17.24 ± 2.85 a | 23.73 ± 2.70 a | 19.01 ± 0.61 a | 21.17 ± 5.02 a |
| Diseased | 23.64 ± 0.45 a | 15.00 ± 0.20 b | 15.39 ± 0.37 b | 24.51 ± 0.89 a | |
| Fe | Healthy | 63.57 ± 4.07 c | 139.81 ± 15.18 ab | 198.88 ± 27.76 a | 107.52 ± 21.69 bc |
| Diseased | 53.75 ± 6.70 b | 65.23 ± 8.23 b | 65.99 ± 6.39 b | 135.26 ± 13.00 a | |
| Zn | Healthy | 13.60 ± 0.28 a | 12.94 ± 0.82 a | 7.58 ± 0.65 b | 11.72 ± 2.97 ab |
| Diseased | 11.41 ± 0.17 a | 9.83 ± 0.54 b | 8.16 ± 0.20 c | 8.99 ± 0.33 bc | |
| Ca | Healthy | 3366.83 ± 267.13 a | 4925.83 ± 420.20 a | 4681.69 ± 695.35 a | 3773.57 ± 757.41 a |
| Diseased | 4213.55 ± 184.85 a | 3363.16 ± 453.80 a | 3619.22 ± 198.31 a | 4149.22 ± 460.40 a | |
| Na | Healthy | 734.87 ± 14.37 ab | 716.83 ± 59.66 ab | 624.14 ± 51.77 b | 846.43 ± 43.19 a |
| Diseased | 741.06 ± 24.99 a | 397.72 ± 10.42 c | 416.98 ± 17.83 c | 623.45 ± 57.23 b | |
| Mg | Healthy | 2397.03 ± 121.52 a | 2601.19 ± 35.95 a | 1902.82 ± 286.89 a | 2521.81 ± 346.38 a |
| Diseased | 2288.30 ± 43.06 a | 1841.96 ± 66.55 c | 1523.38 ± 61.35 d | 2097.05 ± 59.23 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Li, Y. Study on Toxic Substances in Astragalus adsurgens Infected with Alternaria gansuense. Agriculture 2025, 15, 1401. https://doi.org/10.3390/agriculture15131401
Liu H, Li Y. Study on Toxic Substances in Astragalus adsurgens Infected with Alternaria gansuense. Agriculture. 2025; 15(13):1401. https://doi.org/10.3390/agriculture15131401
Chicago/Turabian StyleLiu, Huaqi, and Yanzhong Li. 2025. "Study on Toxic Substances in Astragalus adsurgens Infected with Alternaria gansuense" Agriculture 15, no. 13: 1401. https://doi.org/10.3390/agriculture15131401
APA StyleLiu, H., & Li, Y. (2025). Study on Toxic Substances in Astragalus adsurgens Infected with Alternaria gansuense. Agriculture, 15(13), 1401. https://doi.org/10.3390/agriculture15131401