
Preparation and Characterization of Liquid Fertilizers Produced by Anaerobic Fermentation

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Localization
2.2. Biodigester Design
2.3. Components of Biols
2.3.1. Common Components
2.3.2. Specific Components: Microbe Inoculums
2.4. Preparation and Fermentation Process of Biols
2.4.1. Biol from Native Forest Duff (BNF)
2.4.2. Biol from Cow Dung (BCD)
2.4.3. Fermentation Monitoring Parameters
2.5. Analytical Methods
2.5.1. Organic Matter
2.5.2. Mineral Composition
2.5.3. Amino Acids
2.5.4. Plant Hormones
2.5.5. Microbiome: Metagenomic Analysis
2.5.6. Microbiological Count
2.5.7. Determination of Volatile Compounds by HS-SBSE-GC-MS
2.6. Statistical Analysis
3. Results and Discussion
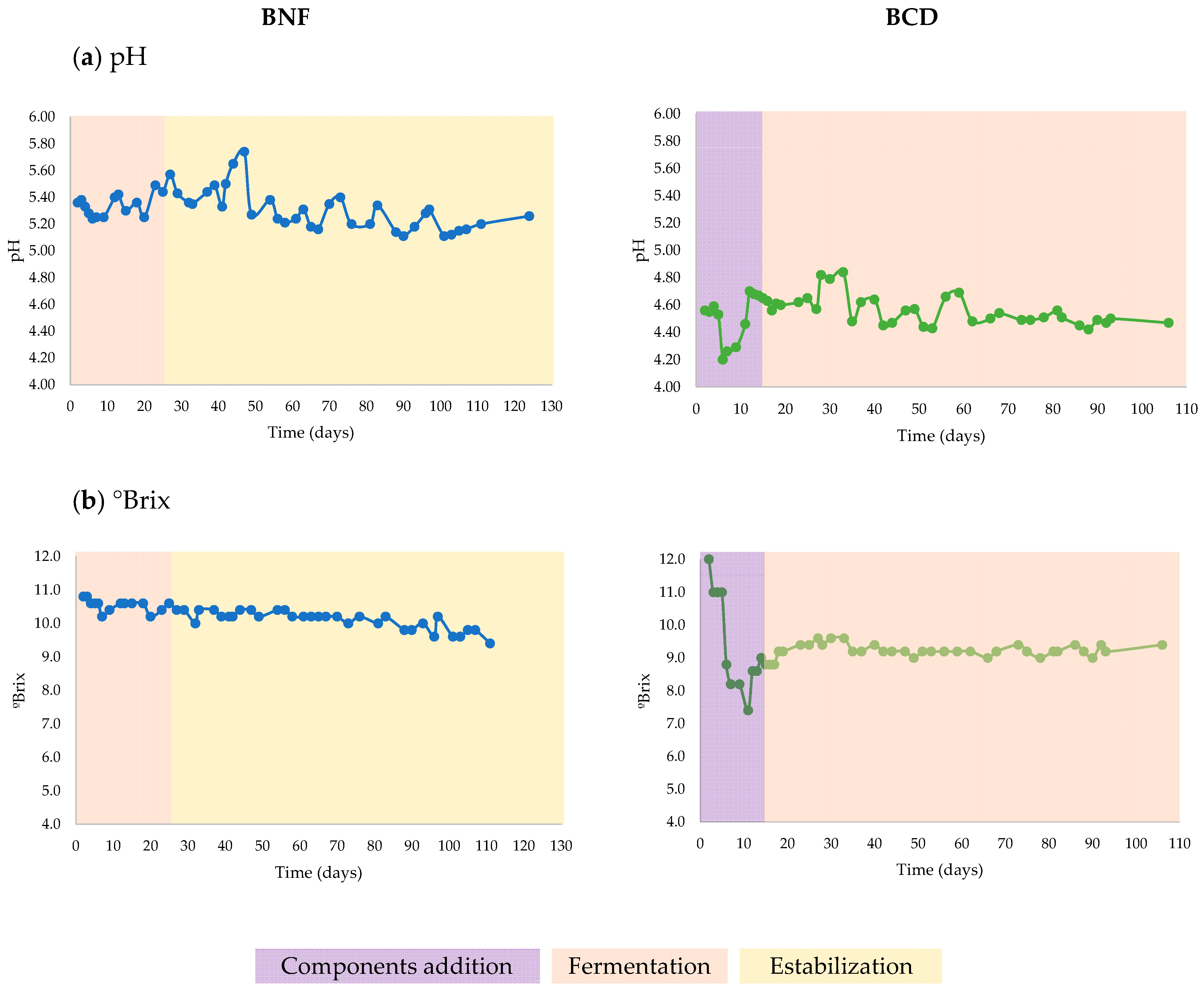
3.1. Biols Preparation and Evolution
3.2. Biols Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cataldo, E.; Fucile, M.; Mattii, G.B. Biostimulants in Viticulture: A Sustainable Approach against Biotic and Abiotic Stresses. Plants 2022, 11, 162. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Diksha; Sindhu, S.S.; Kumar, R. Biofertilizers: An Ecofriendly Technology for Nutrient Recycling and Environmental Sustainability. Curr. Res. Microb. Sci. 2022, 3, 100094. [Google Scholar] [CrossRef]
- Santoyo, G.; Orozco-Mosqueda, M.C.; Govindappa, M. Mechanisms of Biocontrol and Plant Growth-Promoting Activity in Soil Bacterial Species of Bacillus and Pseudomonas: A Review. Biocontr. Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Rathore, G.; Kaushal, R.; Sharma, V.; Sharma, G.; Chaudhary, S.; Dhaliwal, S.S.; Alsuhaibani, A.M.; Gaber, A.; Hossain, A. Evaluation of the Usefulness of Fermented Liquid Organic Formulations and Manures for Improving the Soil Fertility and Productivity of Brinjal (Solanum melongena L.). Agriculture 2023, 13, 417. [Google Scholar] [CrossRef]
- Gillespie, G.; Hardwick, D. Bioferments, Biostimulants & Biofertilizers Making Them on the Farm; Soil Land Food: Albury, Australia, 2018; Available online: https://www.gerrygillespie.net/uploads/4/5/6/5/45656863/bioferments_biostimulants_and_biofertilisers_-_making_them_on-farm_fact_sheet_final_metric_march_2019.pdf (accessed on 1 June 2025).
- O’Neill, B.; Ramos-Abensur, V. A Review of the State of Knowledge and Use of Liquid Ferments and Biol in the Andes; University of Michigan: Ann Arbor, MI, USA, 2022. [Google Scholar]
- Sarkar, S.; Kundu, S.; Ghorai Krishi Vigyan Kendra, D. Validation of Ancient Liquid Organics-Panchagavya and Kunapajala as Plant Growth Promoters; NISCAIR-CSIR: New Delhi, India, 2014; Volume 13. [Google Scholar]
- TECA; FAO. Supermagro: Abono Liquido Foliar Organico. 2008. Available online: https://www.fao.org/research-extension-systems/portals/teca/es/ (accessed on 6 January 2016).
- Leite, C.D.; Meira, A.L. Fichas Agroecologicas Preparo Do Biofertilizante Supermagro; Ministerio da Agricultura, Pecuaria e Abastecimento: Brasília, Brazil, 1991.
- Fernandes, A.L.T.; Rodrigues, G.P.; Testezlaf, R. Fertirrigação Mineral e Organomineral Na Qualidade Do Melão Cultivado Em Estufa. Sci. Agric. 2003, 60, 149–154. [Google Scholar] [CrossRef]
- Restrepo, J.; Pinheiro, S. Cromatografía Imágenes de Vida y Destrucción Del Suelo; Imágenes Gráficas S. A.: Cali, Colombia, 2015; ISBN 978-958-44-8582-3. [Google Scholar]
- Martin, D. Could a Fermented Biofertilizer Replace a Commercial Fertilizer for Glasshouse Lettuce (Lactuca sativa L.) Production? Bachelor’s Thesis, University of Nottingham, Nottingham, UK, 2018. [Google Scholar]
- Cano-Hernández, M.; Bennet-Eaton, A.; Silva-Guerrero, E.; Robles-González, S.; Sainos-Aguirre, U.; Castorena-García, H. Characterization of Bioles from the Anaerobic Fermentation of Cattle and Swine Excreta. Agrociencia 2016, 50, 471–479. [Google Scholar]
- Abo Elsoud, M.M.; Hasan, S.F.; Elhateir, M.M. Optimization of Indole-3-Acetic Acid Production by Bacillus Velezensis Isolated from Pyrus Rhizosphere and Its Effect on Plant Growth. Biocatal. Agric. Biotechnol. 2023, 50, 102714. [Google Scholar] [CrossRef]
- Marty, B. Microbiology of Anaerobic Digestion. In Anaerobic Digestion of Sewage Sludge Agricultural Wastes; Bruce, A.M., Kouzeli-Katsiri, A., Newman, P.J., Eds.; Elsevier Science Publishing Co.: New York, NY, USA, 1984; pp. 72–85. [Google Scholar]
- Lopez-Rubio, J.F. Evaluacion de La Integridad Del Suelo En Olivares Con Practicas Agricolas Contrastadas. Master’s Thesis, Universidad de Alcalá, Alcalá de Henares, Spain, 2022. [Google Scholar]
- Khemani, D.; Joseph, M.; Variganti, S. Case Based Interpretation of Soil Chromatograms. In Proceedings of the Advances in Case-Based Reasoning; Lecture Notes in Computer Science; Springer: Berlin, Heidelberg, 2008; pp. 587–599. [Google Scholar]
- Chazarra Bernabé, A.; Flórez García, E.; Peraza Sánchez, B.; Tohá Rebull, T.; Lorenzo Mariño, B.; Criado Pinto, E.; Moreno García, J.V.; Romero Fresneda, R.; Botey Fullat, R. Mapas Climáticos de España (1981-2010) y ETo (1996-2016); Agencia Estatal de Meteorología: Madrid, Spain, 2018. [Google Scholar]
- Hernández-Chontal, M.A.H.; Collado, C.J.L.; Orozco, N.R.; Velasco, J.V.; Gabriel, A.L.; Romero, G.L. Nutrient Content of Fermented Fertilizers and Its Efficacy in Combination with Hydrogel in Zea mays L. Int. J. Recycl. Org. Waste Agric. 2019, 8, 309–315. [Google Scholar] [CrossRef]
- Restrepo, J. Manual Práctico El A, B, C de La Agricultura Orgánica y Harina de Rocas, 1st ed.; SIMAS: Managua, Nicaragua, 2007; ISBN 978-99924-55-27-2. [Google Scholar]
- Dantinne, V. Manuel de La Litière Forestière Fermentée: Une Préparation Simple et Économique Pour Des Cultures Vigoureuses; Association Terre & Humanisme: Lablachère, France, 2021; ISBN 9782812621802. [Google Scholar]
- ISO/IEC 17025; General Requirements for the Competence of Testing and Calibration Laboratories. International Organization for Standardization: Geneva, Switzerland, 2017.
- UNE 77321:2003; Calidad Del Suelo. Determinación de Carbono Orgánico y Carbono Total Mediante Combustión Seca (Análisis Elemental). AENOR: Madrid, Spain, 2024.
- Amin, M.; Lim, L.W.; Takeuchi, T. Determination of Common Inorganic Anions and Cations by Non-Suppressed Ion Chromatography with Column Switching. J. Chromatogr. A 2008, 1182, 169–175. [Google Scholar] [CrossRef]
- UNE-EN ISO16963:2019; Fertilizantes. Determinación de Boro, Cobalto, Cobre, Hierro, Manganeso, Molibdeno y Zinc Utilizando ICP-AES. AENOR: Madrid, Spain, 2019.
- Han, M.; Xie, M.; Han, J.; Yuan, D.; Yang, T.; Xie, Y. Development and Validation of a Rapid, Selective, and Sensitive LC-MS/MS Method for Simultaneous Determination of D- and L-Amino Acids in Human Serum: Application to the Study of Hepatocellular Carcinoma. Anal. Bioanal. Chem. 2018, 410, 2517–2531. [Google Scholar] [CrossRef]
- Hou, S.; Zhu, J.; Ding, M.; Lv, G. Simultaneous Determination of Gibberellic Acid, Indole-3-Acetic Acid and Abscisic Acid in Wheat Extracts by Solid-Phase Extraction and Liquid Chromatography–Electrospray Tandem Mass Spectrometry. Talanta 2008, 76, 798–802. [Google Scholar] [CrossRef]
- Morrison, G.A.; Fu, J.; Lee, G.C.; Wiederhold, N.P.; Cañete-Gibas, C.F.; Bunnik, E.M.; Wickes, B.L. Nanopore Sequencing of the Fungal Intergenic Spacer Sequence as a Potential Rapid Diagnostic Assay. J. Clin. Microbiol. 2020, 58, e01972-20. [Google Scholar] [CrossRef] [PubMed]
- Benítez-Páez, A.; Sanz, Y. Multi-Locus and Long Amplicon Sequencing Approach to Study Microbial Diversity at Species Level Using the MinIONTM Portable Nanopore Sequencer. Gigascience 2017, 6, 1–12. [Google Scholar] [CrossRef]
- Sánchez-Gómez, R.; Zalacain, A.; Alonso, G.L.; Salinas, M.R. Effect of Toasting on Non-Volatile and Volatile Vine-Shoots Low Molecular Weight Phenolic Compounds. Food Chem. 2016, 204, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, S.; Kumar, S.; Baliyan, N.; Dheeman, S.; Maheshwari, D.K. Cattle Dung Manure Microbiota as a Substitute for Mineral Nutrients and Growth Management Practices in Plants. Endophytes Miner. Nutr. Manag. 2021, 3, 77–103. [Google Scholar]
- Shin, K.; van Diepen, G.; Blok, W.; van Bruggen, A.H.C. Variability of Effective Micro-Organisms (EM) in Bokashi and Soil and Effects on Soil-Borne Plant Pathogens. Crop. Prot. 2017, 99, 168–176. [Google Scholar] [CrossRef]
- Mangalanayaki, R. Biocontrol Activity of Bacillus Subtilis Isolated from Cow Dung Against Plant Pathogenic Fungi. Int. J. Pure Appl. Biosci. 2016, 4, 80–86. [Google Scholar] [CrossRef]
- Kumari, K.S.; Devi, S.P.; Ranjithkumar, R.; Djearamane, S.; Tey, L.-H.; Wong, L.S.; Kayarohanam, S.; Arumugam, N.; Almansour, A.I.; Perumal, K. Organic Remobilization of Zinc and Phosphorus Availability to Plants by Application of Mineral Solubilizing Bacteria Pseudomonas Aeruginosa. Heliyon 2023, 9, e22128. [Google Scholar] [CrossRef]
- Singh, B.; Christina, E. Indigenous Microorganisms as an Effective Tool for in Situ Bioremediation. In Relationship Between Microbes and the Environment for Sustainable Ecosystem Services, Volume 2: Microbial Mitigation of Waste for Sustainable Ecosystem Services; Elsevier: Amsterdam, The Netherlands, 2022; Volume 5, pp. 273–295. [Google Scholar] [CrossRef]
- Swain, M.R.; Ray, R.C. Biocontrol and Other Beneficial Activities of Bacillus Subtilis Isolated from Cowdung Microflora. Microbiol. Res. 2009, 164, 121–130. [Google Scholar] [CrossRef]
- Félix-Herrán, J.A.; Sañudo-Torres, R.R.; Rojo-Martínez, G.E.; Martínez-Ruiz, R.; Olalde-Portugal, V. Importance of Organic Manures. Ra Ximhai Rev. Soc. Cult. Desarrollo Sustentable 2008, 4, 57–67. [Google Scholar]
- Oliveira, D.; Schneider, S.; Marques, F.C. Contextualização e Práticas Criativas Na Agricultura Ecológica de Ipê e Antônio Prado/RS: O Biofertilizante Super Magro Como Objeto Epistêmico. Desenvolv. Meio Ambiente 2017, 41, 83–104. [Google Scholar] [CrossRef]
- de Oliveira Neto, H.T.; de Oliveira Gondim, A.R.; da Silva Sá, F.V.; Souto, L.S.; Brito, M.E.B.; da Silva, M.S.; de Lira, R.P. Growth, Gas Exchanges and Production of Beet CV. Katrina under Organo-Mineral Fertilization. Biosci. J. 2017, 33, 1126–1133. [Google Scholar] [CrossRef]
- Nekhubvi, V. The Investigation of Chemical Composition and the Specific Heat Capacity of Cow Dung and Water Mixture. In Anaerobic Digestion-Biotechnology for Environmental Sustainability; IntechOpen: London, UK, 2023. [Google Scholar]
- Ndubuisi-Nnaji, U.U.; Ofon, U.A.; Asamudo, N.U.; Ekong, V.M. Enhanced Biogas and Biofertilizer Production from Anaerobic Codigestion of Harvest Residues and Goat Manure. J. Sci. Res. Rep. 2020, 26, 1–13. [Google Scholar] [CrossRef]
- Orellana, T.; Manzano, P.; Chávez, E.; Ruiz, O.; León, R.; Manzano, A.O.; Peralta, E. Estándares de Fermentación y Maduración Artesanal de Bioles. Yachana 2013, 2, 1–7. [Google Scholar]
- Möller, K.; Müller, T. Effects of Anaerobic Digestion on Digestate Nutrient Availability and Crop Growth: A Review. Eng. Life Sci. 2012, 12, 242–257. [Google Scholar] [CrossRef]
- Criollo, H.; Lagos, T.; Piarpuezan, E.; Pérez, R. The Effect of Three Liquid Bio-Fertilizers in the Production of Lettuce (Lactuca sativa L.) and Cabbage (Brassica oleracea L. Var. Capitata). Agron. Colomb. 2011, 29, 415–421. [Google Scholar]
- Mei, R.; Narihiro, T.; Nobu, M.K.; Kuroda, K.; Liu, W.T. Evaluating Digestion Efficiency in Full-Scale Anaerobic Digesters by Identifying Active Microbial Populations through the Lens of Microbial Activity. Sci. Rep. 2016, 6, srep34090. [Google Scholar] [CrossRef]
- Zhang, W.; Werner, J.J.; Agler, M.T.; Angenent, L.T. Substrate Type Drives Variation in Reactor Microbiomes of Anaerobic Digesters. Bioresour. Technol. 2014, 151, 397–401. [Google Scholar] [CrossRef]
- Sun, L.; Pope, P.B.; Eijsink, V.G.H.; Schnürer, A. Characterization of Microbial Community Structure during Continuous Anaerobic Digestion of Straw and Cow Manure. Microb. Biotechnol. 2015, 8, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Chontal, M.A. Elaboración Artesanal de Fertilizante Líquido Fermentado “Biol”, En Dos Épocas Del Año. In Agro Veracruzano; Instituto Tecnológico Superior de San Andrés Tuxtla (ITSSAT): San Andrés Tuxtla, Mexico, 2016; pp. 270–285. ISBN 978-607-97384-0-2. [Google Scholar]
- McCord, A.I.; Stefanos, S.A.; Tumwesige, V.; Lsoto, D.; Kawala, M.; Mutebi, J.; Nansubuga, I.; Larson, R.A. Anaerobic Digestion in Uganda: Risks and Opportunities for Integration of Waste Management and Agricultural Systems. Renew. Agric. Food Syst. 2020, 35, 678–687. [Google Scholar] [CrossRef]
- Insam, H.; Gómez-Brandón, M.; Ascher, J. Manure-Based Biogas Fermentation Residues—Friend or Foe of Soil Fertility? Soil. Biol. Biochem. 2015, 84, 1–14. [Google Scholar] [CrossRef]
- Möller, K. Effects of Anaerobic Digestion on Soil Carbon and Nitrogen Turnover, N Emissions, and Soil Biological Activity. A Review. Agron. Sustain. Dev. 2015, 35, 1021–1041. [Google Scholar] [CrossRef]
- Navarro-García, G. Fertilizantes. Química y Acción, 2nd ed.; Mundiprensa: Madrid, Spain, 2023; ISBN 978-8484767633. [Google Scholar]
- European Union. Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019, Laying down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003. Off. J. Eur. Union 2019, L170, 1–114. [Google Scholar]
- Nasir, I.M.; Idaty, T.; Ghazi, M.; Omar, R.; Azlina, W.; Ab, W.; Ghani, K. Anaerobic Batch Digestion of Cattle Manure Using a Novel Oscillatory Flow Bioreactor. Int. J. Eng. Technol. 2014, 11, 65–69. [Google Scholar]
- Leblanc, H.A.; Cerrato, M.E.; Miranda, A.; Valle, G. Determinación de La Calidad de Abonos a Través de Bioensayos. Tierra Trop. 2007, 3, 97–107. [Google Scholar]
- Xu, H.; Zhou, Z.; Chen, H.; Wu, X.; Heng, X.; Li, K.; Wang, Z.; Ni, Z.; Gao, E.; Yong, Y. Complexation of Multiple Mineral Elements by Fermentation and Its Application in Laying Hens Complexation of Multiple Mineral Elements by Fermentation and Its Application in Laying Hens. TYPE Orig. Res. Publ. 2022, 29, 1001412. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, X.; Gao, T.; Zhang, M.; Min, X.; Chen, W.; Tang, C.J. Rapid Monitoring of Heavy Metal-Ammonia Complexes in Solutions by UV–Vis/ATR-FTIR Spectroscopy and Chemometric Models. Chem. Eng. J. 2024, 481, 148692. [Google Scholar] [CrossRef]
- Gurd, F.R.N.; Wilcox, P.E. Complex Formation between Metallic Cations and Proteins, Peptides, and Amino Acids. Adv. Protein Chem. 1956, 11, 311–427. [Google Scholar] [CrossRef]
- Scheper, T.H.; Lammers, F. Fermentation Monitoring and Process Control. Curr. Opin. Biotechnol. 1994, 5, 187–191. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of Gaba (γ- Aminobutyric Acid) by Microorganisms: A Review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef]
- D’Este, M.; Alvarado-Morales, M.; Angelidaki, I. Amino Acids Production Focusing on Fermentation Technologies—A Review. Biotechnol. Adv. 2018, 36, 14–25. [Google Scholar] [CrossRef]
- Sanchez, S.; Rodríguez-Sanoja, R.; Ramos, A.; Demain, A.L. Our Microbes Not Only Produce Antibiotics, They Also Overproduce Amino Acids. J. Antibiot. 2017, 71, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.; Spano, G.; Bauer, F.F.; Fairbairn, S.; Mckinnon, A.; Musarurwa, H.T.; Ferreira, A.C. The Impact of Single Amino Acids on Growth and Volatile Aroma Production by Saccharomyces Cerevisiae Strains. Front. Microbiol. 2017, 8, 2554. [Google Scholar] [CrossRef]
- Sahab, N.R.M.; Subroto, E.; Balia, R.L.; Utama, G.L. γ-Aminobutyric Acid Found in Fermented Foods and Beverages: Current Trends. Heliyon 2020, 6, e05526. [Google Scholar] [CrossRef]
- Yao, X.; Zhou, H.; Meng, H.; Ding, J.; Shen, Y.; Cheng, H.; Zhang, X.; Li, R.; Fan, S. Amino Acid Profile Characterization during the Co-Composting of a Livestock Manure and Maize Straw Mixture. J. Clean. Prod. 2021, 278, 123494. [Google Scholar] [CrossRef]
- Lee, K.-T.; Liao, H.-S.; Hsieh, M.-H. Special Issue-Review Glutamine Metabolism, Sensing and Signaling in Plants. Plant Cell Physiol. 2023, 64, 1466–1481. [Google Scholar] [CrossRef]
- Ma, J.; Jiang, H.; Li, P.; Li, C.; Liu, R.; Li, J.; Xiao, Z.; Pi, B.; Zhao, M.; Hu, W.; et al. Production of Free Amino Acid Fertilizer from Tung Meal by the Newly Isolated Pseudomonas Aeruginosa LYT-4 Strain with Simultaneous Potential Biocontrol Capacity. Renew. Energy 2020, 166, 245–252. [Google Scholar] [CrossRef]
- Van de Poll, M.C.G.; Luiking, Y.C.; Dejong, C.H.C.; Soeters, P.B. Amino Acids: Specific Functions. In Encyclopedia of Human Nutrition; Elsevier: Amsterdam, The Netherlands, 2013; pp. 79–87. [Google Scholar] [CrossRef]
- Kumari, A.; Kumar, V.; Ovadia, R.; Oren-Shamir, M. Phenylalanine in Motion: A Tale of an Essential Molecule with Many Faces. Biotechnol. Adv. 2023, 68, 108246. [Google Scholar] [CrossRef]
- Cai, G.; Li, J.; Zhou, M.; Zhu, G.; Li, Y.; Lv, N.; Wang, R.; Li, C.; Pan, X. Compost-Derived Indole-3-Acetic-Acid-Producing Bacteria and Their Effects on Enhancing the Secondary Fermentation of a Swine Manure-Corn Stalk Composting. Chemosphere 2022, 291, 132750. [Google Scholar] [CrossRef]
- Bunsangiam, S.; Thongpae, N.; Limtong, S.; Srisuk, N. Large Scale Production of Indole-3-Acetic Acid and Evaluation of the Inhibitory Effect of Indole-3-Acetic Acid on Weed Growth. Sci. Rep. 2021, 11, 13094. [Google Scholar] [CrossRef]
- Mishra, A.K.; Baek, K.H. Salicylic Acid Biosynthesis and Metabolism: A Divergent Pathway for Plants and Bacteria. Biomolecules 2021, 11, 705. [Google Scholar] [CrossRef]
- Miché, L.; Dries, A.; Ben Ammar, I.; Davidson, S.; Cagnacci, L.; Combet-Blanc, Y.; Abecassis, V.; Penton Fernandez, G.; Christen, P. Changes in Chemical Properties and Microbial Communities’ Composition of a Forest Litter–Based Biofertilizer Produced through Aerated Solid-State Culture under Different Oxygen Conditions. Environ. Sci. Pollut. Res. 2024. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Dai, L.; Guo, Z.; Wang, J.; Zhang, X.; Chen, W.; Chen, Y.; Qu, H.; Dai, L.; Guo, Z.; et al. Antifungal Activity of Dimethyl Trisulfide and Potential Biocontrol against Alternaria Alternata on Postharvest Lycium Barbarum The Half Inhibitory Concentration POD Peroxidase PDA Potato Dextrose Agar Medium MDA Malondialdehyde H2O2 Hydrogen Peroxide ROS Reactive Oxygen Species. Eur. J. Plant Pathol. 2024, 171, 645–660. [Google Scholar] [CrossRef]
- Fayek, N.M.; Xiao, J.; Farag, M.A. A Multifunctional Study of Naturally Occurring Pyrazines in Biological Systems; Formation Mechanisms, Metabolism, Food Applications and Functional Properties. Crit. Rev. Food Sci. Nutr. 2021, 63, 5322–5338. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Biological Function | Number of Species (×105) | |
|---|---|---|
| NFI | CDI | |
| Bacteria producing siderophores | 76.62 | 99.51 |
| Biocontrol | 72.44 | 29.52 |
| Exopolysaccharides | 71.13 | 43.18 |
| Nitrogen fixation | 52.37 | 72.82 |
| Pathogenic bacteria to crops | 74.07 | 12.76 |
| Phosphate solubilizing bacteria | 0.49 | 120.34 |
| Phytase production | 1.16 | 11.33 |
| Phytohormones | 74.10 | 29.73 |
| Potassium-solubilizing bacteria | 71.96 | 115.10 |
| BNF | BCD | Fp-value | |
|---|---|---|---|
| General Parameters | |||
| °Brix | 9.4 ± 0.21 a | 9.4 ± 0.50 a | 0.00 |
| Conductivity (mS/cm) | 31.54 ± 0.52 a | 32.76 ± 0.98 a | 3.63 |
| pH | 5.26 ± 0.09 b | 4.47 ± 0.08 a | 129.12 *** |
| TDS (ppt) | 22.10 ± 0.36 a | 22.94 ± 0.69 a | 3.49 |
| Organic Matter and Microorganisms | |||
| Carbon (%) | 2.82 ± 0.32 | 2.56 ± 0.29 | 1.09 |
| Organic matter (%) | 4.87 ± 0.52 | 4.41 ± 0.47 | 1.29 |
| Colony-forming units/g # | <1.0 × 101 | <1.0 × 101 | 0.00 |
| BNF | BCD | Fp-value | |
|---|---|---|---|
| Anions | |||
| Borates | 120 ± 8.0 a | 110 ± 7.4 a | 2.50 |
| Chlorides | 7114 ± 948 a | 6775 ± 903 a | 0.20 |
| Fluorides | 0.50 ± 0.07 a | 0.50 ± 0.07 a | 0.00 |
| Nitrates | 1800 ± 192 a | 2590 ± 276 b | 16.54 * |
| Nitrites | 0.10 ± 0.01 a | 0.10 ± 0.01 a | 0.00 |
| Phosphates | 75.7 ± 10.1 a | 312.0 ± 41.6 b | 91.42 *** |
| Sulfates | 7785 ± 1038 a | 8200 ± 10,913 a | 0.23 |
| Cations | |||
| Ammonium | 58.6 ± 5.23 a | 67.5 ± 6.03 a | 3.73 |
| Calcium # | 1200 ± 151 a | 1900 ± 240 b | 18.41 * |
| Cobalt | 58.0 ± 7.7 a | 58.0 ± 7.7 a | 0.00 |
| Copper | 210 ± 28 a | 210 ± 28 a | 0.00 |
| Iron | 100.0 ± 12.7 a | 240.0 ± 30.4 b | 54.21 ** |
| Magnesium # | 2500 ± 316 a | 2300 ± 291 a | 0.65 |
| Manganese | 510 ± 68 a | 490 ± 65 a | 0.13 |
| Molybdenum | 170 ± 22 a | 170 ± 22 a | 0.00 |
| Potassium # | 12,000 ± 1520 a | 13,000 ± 1646 a | 0.60 |
| Sodium # | 1800 ± 228 a | 2700 ± 342 b | 14.38 * |
| Zinc | 1800 ± 216 a | 1700 ± 204 a | 0.34 |
| Complex forms | |||
| Calcium # | 1000 ± 126 a | 1600 ± 202 b | 18.91 * |
| Cobalt | 381 ± 51 a | 359 ± 48 a | 0.30 |
| Copper | 110.0 ± 14.7 a | 130.0 ± 17.3 a | 2.33 |
| Iron | 49.0 ± 5.9 a | 110.0 ± 13.2 b | 53.46 ** |
| Magnesium # | 1500 ± 190 a | 1400 ± 177 a | 0.44 |
| Manganese | 260.0 ± 34.7 a | 240.0 ± 32.0 | 0.54 |
| Molybdenum | 130 ± 15 a | 130 ± 17 a | 0.00 |
| Potassium # | 6900 ± 874 a | 18,200 ± 2305 b | 63.02 ** |
| Sodium # | 9900 ± 1254 a | 11,700 ± 1482 a | 2.58 |
| Zinc | 880 ± 82 a | 920 ± 86 a | 0.34 |
| BNF | BCD | Fp-value | |
|---|---|---|---|
| Amino acids | |||
| Alanine | 143.0 ± 19.1 a | 203.0 ± 27.1 b | 9.85 * |
| β-Alanine | 3.08 ± 0.41 a | 3.87 ± 0.51 a | 4.27 |
| γ-Aminobutyric acid | 4.53 ± 0.60 a | 6.21 ± 0.82 b | 8.07 * |
| Glycine | 19.5 ± 2.6 a | 14.7 ± 1.9 a | 6.52 |
| Glutamic acid | 73.0 ± 9.7 a | 85.7 ± 11.4 a | 2.15 |
| Glutamine | 2.57 ± 0.34 b | 0.82 ± 0.11 a | 71.95 ** |
| Histamine | 0.22 ± 0.03 b | 0.03 ± 0.01 a | 111.64 *** |
| Histidine | 0.51 ± 0.07 b | 0.15 ± 0.02 a | 73.36 ** |
| Phenylalanine | 11.50 ± 1.53 b | 5.17 ± 0.69 a | 42.67 ** |
| Hormones | |||
| Abscisic acid | 0.20 ± 0.026 | 0.19 ± 0.025 | 0.22 |
| Indole-3-acetic acid | 0.009 ± 0.0012 | 0.050 ± 0.0066 | 109.91 *** |
| Salicylic acid | 0.036 ± 0.005 a | 0.23 ± 0.03 b | 126.95 *** |
| BNF | BCD | Fp-value | |
|---|---|---|---|
| Acids | |||
| Acetic acid | 49.6 ± 15.7 a | 055.5 ± 7.0 a | 0.23 |
| Decanoic acid | 1.9 ± 0.0 a | 5.2 ± 0.1 b | 4225.00 *** |
| Hexanoic acid | 4.7 ± 0.1 a | 61.0 ± 3.2 b | 625.80 ** |
| Octanoic acid | 3.4 ± 0.3 a | 27.3 ± 1.8 b | 354.96 ** |
| Alcohols | |||
| Acetol | 129.1 ± 41.9 a | 127.8 ± 48.8 a | 0.00 |
| Benzyl alcohol | 1.4 ± 0.1 a | 2.4 ± 0.0 b | 441.00 ** |
| Cyclopentanol | 71.0 ± 19.8 a | 79.2 ± 25.6 a | 0.13 |
| Isobutyl alcohol | 9.9 ± 0.2 a | 29.7 ± 0.4 b | 4612.24 *** |
| 3-Methyl-1-butanol | 9.6 ± 0.1 a | 353.5 ± 8.7 b | 3177.52 *** |
| 1-Octen-3-ol | 0.8 ± 0.1 a | 1.0 ± 0.0 b | 25.00 * |
| Nonanol | 0.8 ± 0.0 a | 1.4 ± 0.4 a | 4.00 |
| Phenylethyl alcohol | 6.6 ± 1.1 a | 81.9 ± 4.4 b | 553.18 ** |
| 2,4-di-tert-Butylphenol | 45.0 ± 2.1 a | 58.0 ± 1.0 b | 65.69 * |
| Aldehydes | |||
| Benzaldehyde | 15.8 ± 2.5 a | 12.9 ± 2.5 a | 1.36 |
| Decanal | 4.5 ± 1.6 a | 4.6 ± 1.1 a | 0.01 |
| Nonanal | 35.8 ± 6.0 a | 36.3 ± 0.7 a | 0.02 |
| Octanal | 3.6 ± 0.6 a | 3.2 ± 0.0 a | 0.60 |
| Esters | |||
| Ethyl acetate | 92.1 ± 2.3 a | 1021.6 ± 16.9 b | 5942.92 *** |
| Ethyl butyrate | 3.8 ± 0.0 a | 299.2 ± 3.3 b | 15,795.68 *** |
| Ethyl decanoate | 0.6 ± 0.1 a | 5.5 ± 0.0 b | 2401.00 *** |
| Ethyl dihydrocinnamate | 0.1 ± 0.0 a | 27.7 ± 1.0 b | 1552.33 *** |
| Ethyl hexanoate | 3.5 ± 2.1 a | 306.8 ± 4.8 b | 6661.18 *** |
| Ethyl lactate | 5.0 ± 0.3 a | 173.2 ± 5.3 b | 2004.92 *** |
| Ethyl octanoate | 2.8 ± 0.6 a | 65.7 ± 0.3 b | 19,782.05 *** |
| Diethyl succinate | 1.6 ± 0.7 a | 4.2 ± 0.2 b | 23.86 * |
| Isoamyl acetate | 2.3 ± 1.8 a | 36.9 ± 1.7 b | 382.48 ** |
| Isovaleric acid, ethyl ester | 21.0 ± 1.6 b | 10.6 ± 0.1 a | 81.63 * |
| Phenylacetaldehyde | 9.9 ± 0.1 a | 10.8 ± 0.1 b | 162.00 ** |
| Norisoprenoids | |||
| β-Damascenone | 2.3 ± 0.2 a | 4.0 ± 0.0 b | 115.60 ** |
| Terpenes | |||
| Elemol | 61.7 ± 42.1 a | 48.6 ± 9.7 a | 0.18 |
| D-Limonene | 3.4 ± 0.4 a | 3.9 ± 0.4 a | 1.39 |
| Linalool | 1.1 ± 0.1 a | 2.4 ± 0.0 b | 729.00 ** |
| Nerol | 1.4 ± 0.4 a | 2.3 ± 0.8 a | 1.80 |
| Pinene | 18.7 ± 1.4 a | 17.0 ± 4.8 a | 0.23 |
| α-Terpineol | 1.7 ± 0.2 a | 3.3 ± 0.1 b | 102.40 ** |
| Pyrazines | |||
| Pyrazine, 2-ethyl-6-methyl-a | 14.4 ± 0.0 a | 26.6 ± 1.3 b | 183.75 ** |
| Pyrazine, 2-ethyl-6-methyl-b | 10.3 ± 0.1 a | 17.0 ± 0.6 b | 264.06 ** |
| Pyrazine, 2-ethyl-3,5-dimethyl | 9.3 ± 0.3 a | 17.6 ± 0.5 b | 418.85 ** |
| Pyrazine, trimethyl | 87.6 ± 1.2 a | 143.1 ± 4.9 b | 243.98 ** |
| Sulfides | |||
| Dimethyl disulfide | 27.1 ± 0.4 b | 15.0 ± 0.5 a | 791.41 ** |
| Dimethyl tetrasulfide | 14.8 ± 1.2 b | 4.6 ± 0.1 a | 101.50 ** |
| Dimethyl trisulfide | 129.4 ± 9.1 b | 53.9 ± 9.4 a | 66.01 * |
| Volatile phenols | |||
| Guaiacol | 1.7 ± 0.2 a | 2.5 ± 0.4 a | 6.42 |
| Syringol | 0.3 ± 0.0 a | 0.5 ± 0.1 a | 9.00 |
| 4-Vinylguaiacol | 3.0 ± 1.2 a | 2.8 ± 0.6 a | 0.03 |
| Others | |||
| 2(5H)-Furanone | 16.0 ± 4.4 a | 17.8 ± 4.6 a | 0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Rubio, J.F.; Cebrián-Tarancón, C.; Alonso, G.L.; Salinas, M.R.; Sánchez-Gómez, R. Preparation and Characterization of Liquid Fertilizers Produced by Anaerobic Fermentation. Agriculture 2025, 15, 1225. https://doi.org/10.3390/agriculture15111225
López-Rubio JF, Cebrián-Tarancón C, Alonso GL, Salinas MR, Sánchez-Gómez R. Preparation and Characterization of Liquid Fertilizers Produced by Anaerobic Fermentation. Agriculture. 2025; 15(11):1225. https://doi.org/10.3390/agriculture15111225
Chicago/Turabian StyleLópez-Rubio, Juan Francisco, Cristina Cebrián-Tarancón, Gonzalo L. Alonso, Maria Rosario Salinas, and Rosario Sánchez-Gómez. 2025. "Preparation and Characterization of Liquid Fertilizers Produced by Anaerobic Fermentation" Agriculture 15, no. 11: 1225. https://doi.org/10.3390/agriculture15111225
APA StyleLópez-Rubio, J. F., Cebrián-Tarancón, C., Alonso, G. L., Salinas, M. R., & Sánchez-Gómez, R. (2025). Preparation and Characterization of Liquid Fertilizers Produced by Anaerobic Fermentation. Agriculture, 15(11), 1225. https://doi.org/10.3390/agriculture15111225