

Preliminary Exploration of Physiology and Genetic Basis Underlying High Yield in Indica–Japonica Hybrid Rice
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Determination of Growth in Seedling Stage, Leaf Morphology, and Chlorophyll Contents
- Chl a (mg L−1) = 12.72A663−2.59A645;
- Chl b (mg L−1) = 22.88A645−4.67A663;
- Chl a + b (mg L−1) = 8.02A663 + 20.21A645;
- Chl contents (mg g−1) = (Chlorophyll concentration × Extraction volume × Dilution factor)/Fresh weight of the sample.
2.3. Determination of Main Agronomy Characters and Yield Traits
2.4. DNA Isolation and InDel Determination for Molecular Distance and Heterosis
2.5. Statistical Analysis
3. Results
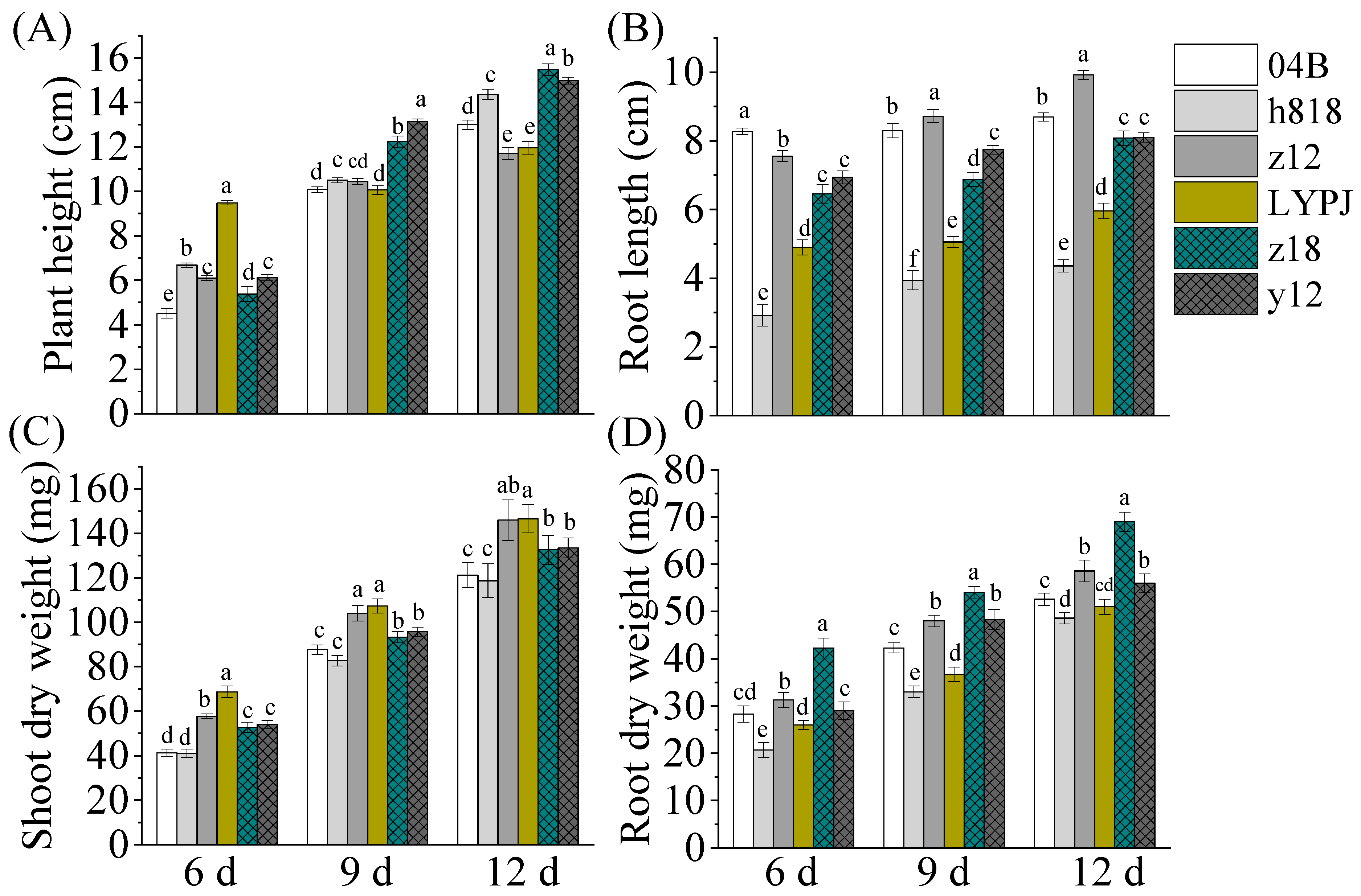
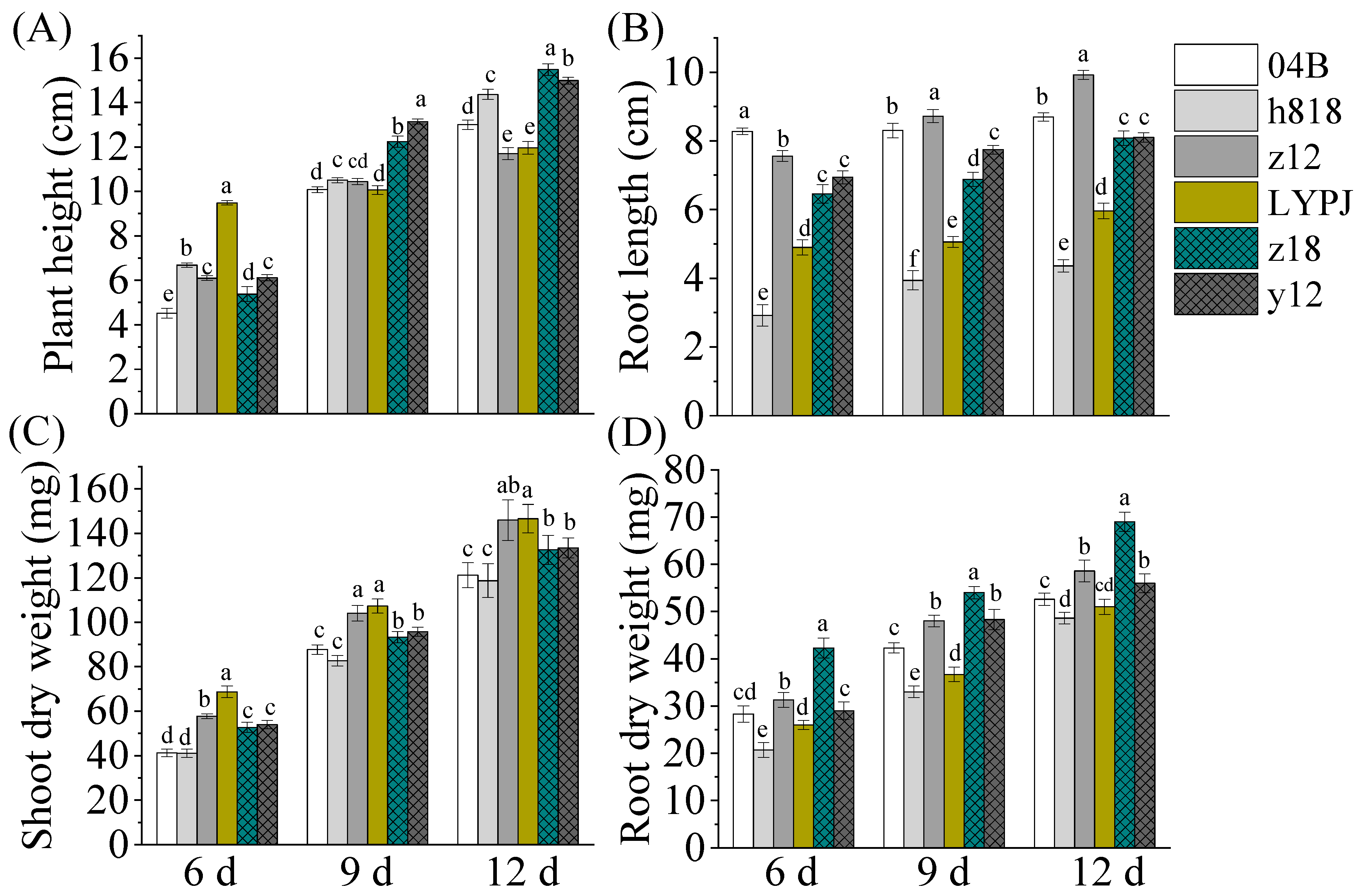
3.1. Indica-Japonica Hybrid Rice Varieties Showed Better Growth Vigor in Seedling Stage
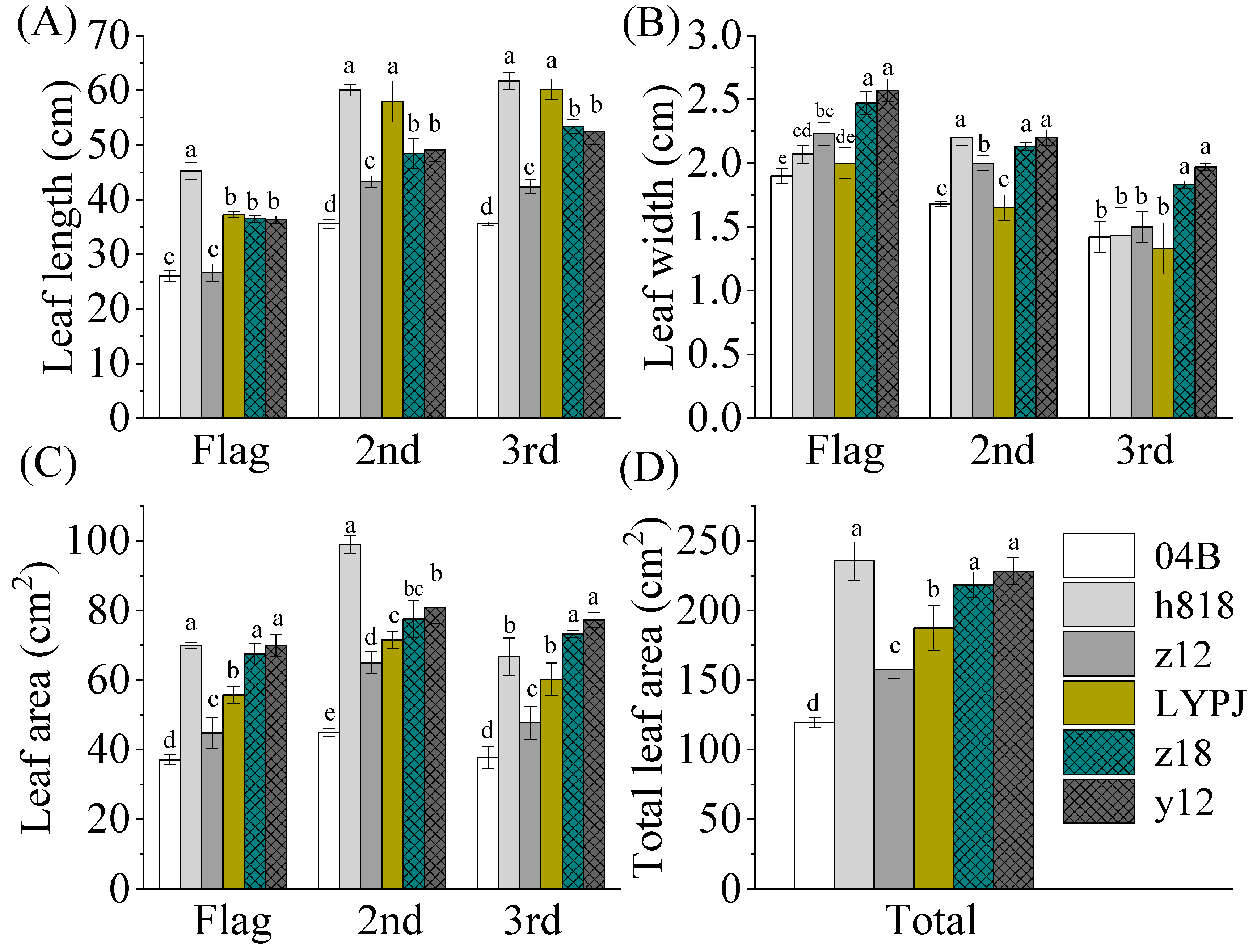
3.2. The Indica–Japonica Hybrid Rice Had Higher Leaf Area and Showed Advantages in Chlorophyll Contents in Seedling and Filling Stages
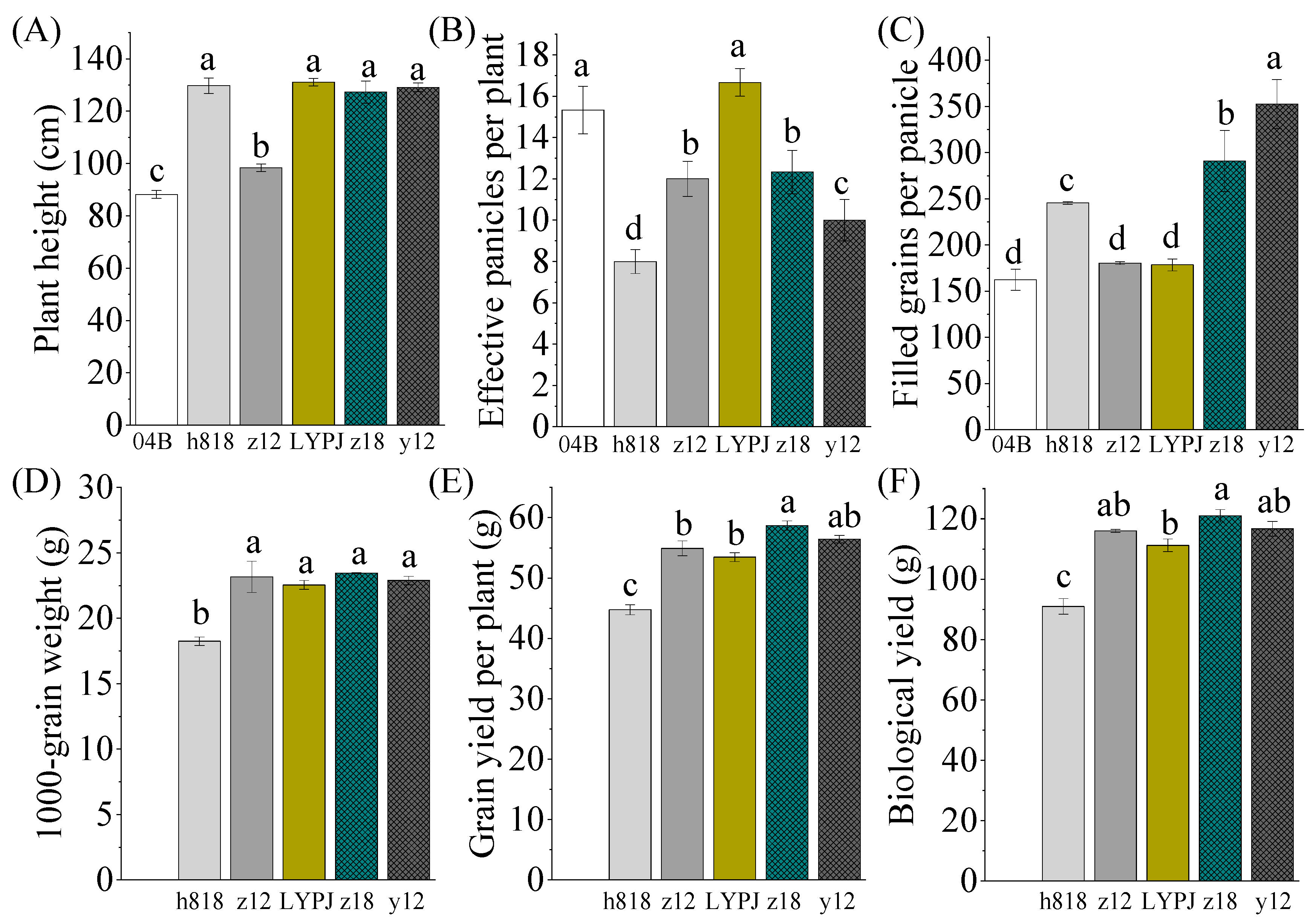
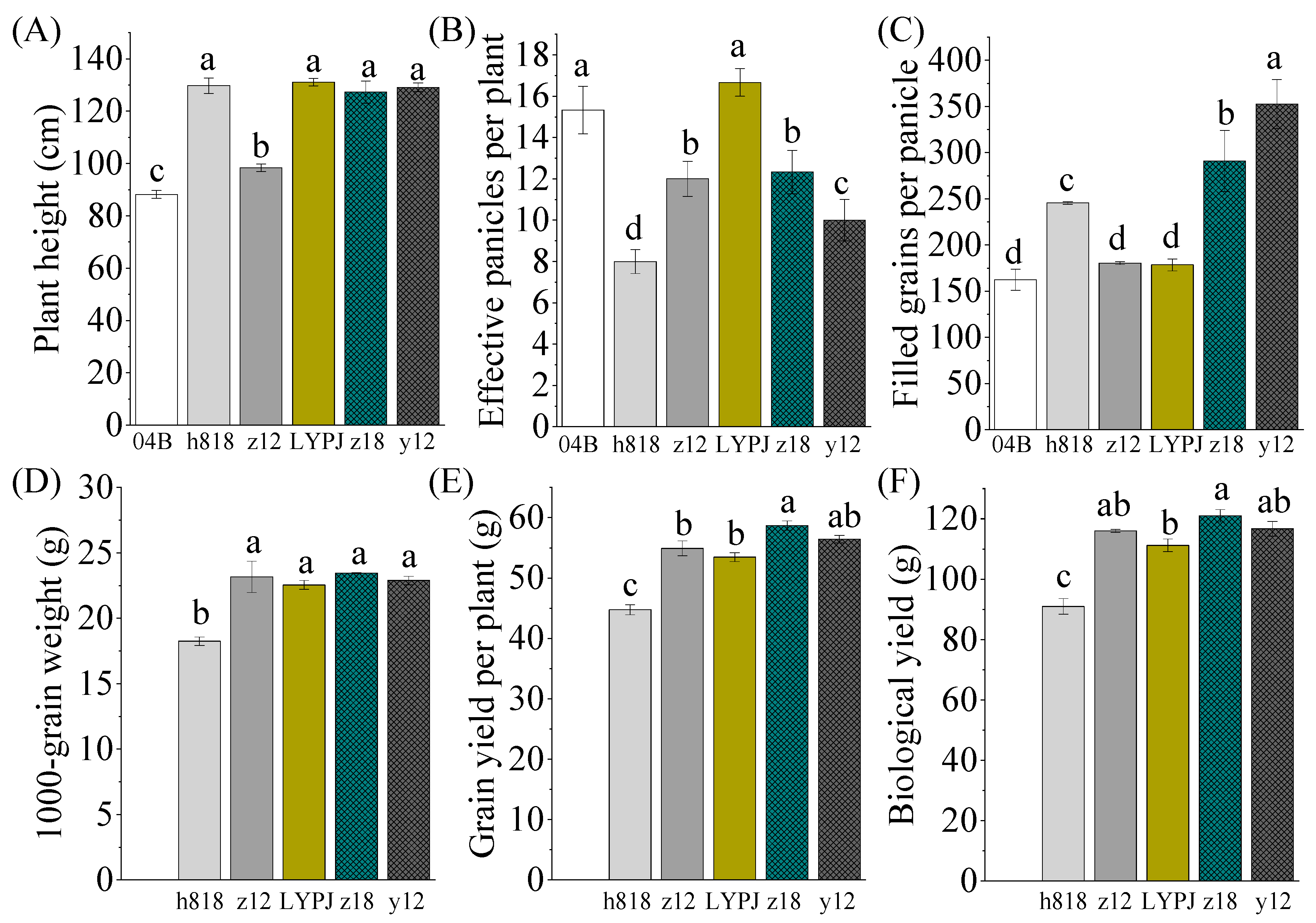
3.3. Determination of Agronomy Traits and Yield Components That Contribute to the High Yield of Indica–Japonica Hybrid Rice
3.4. The Increases in the Number of Primary Branches, Spikelets, and Grains in the Primary and Intermediate Panicles Ultimately Contribute to the Increasing of Yield in Indica–Japonica Hybrid Rice
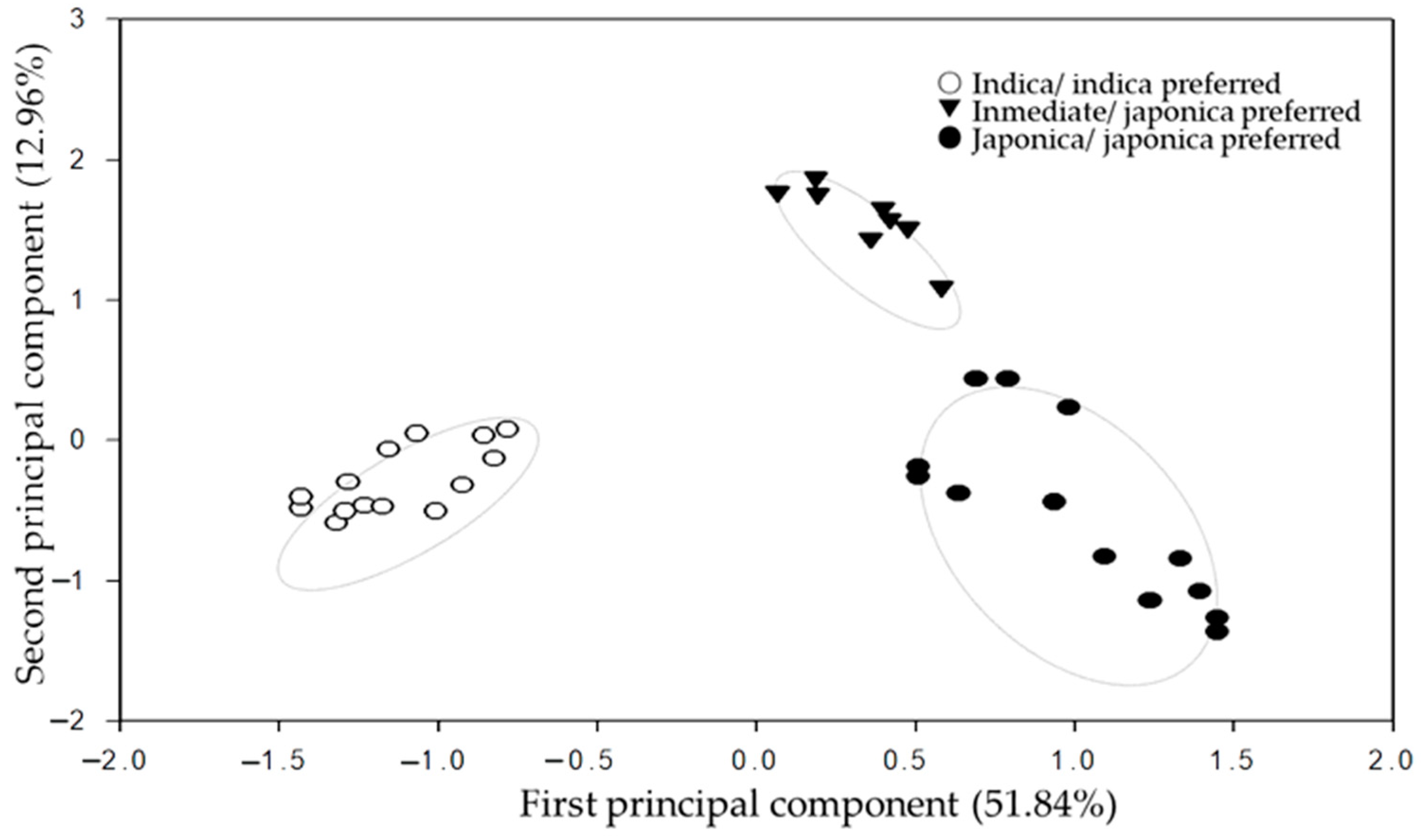
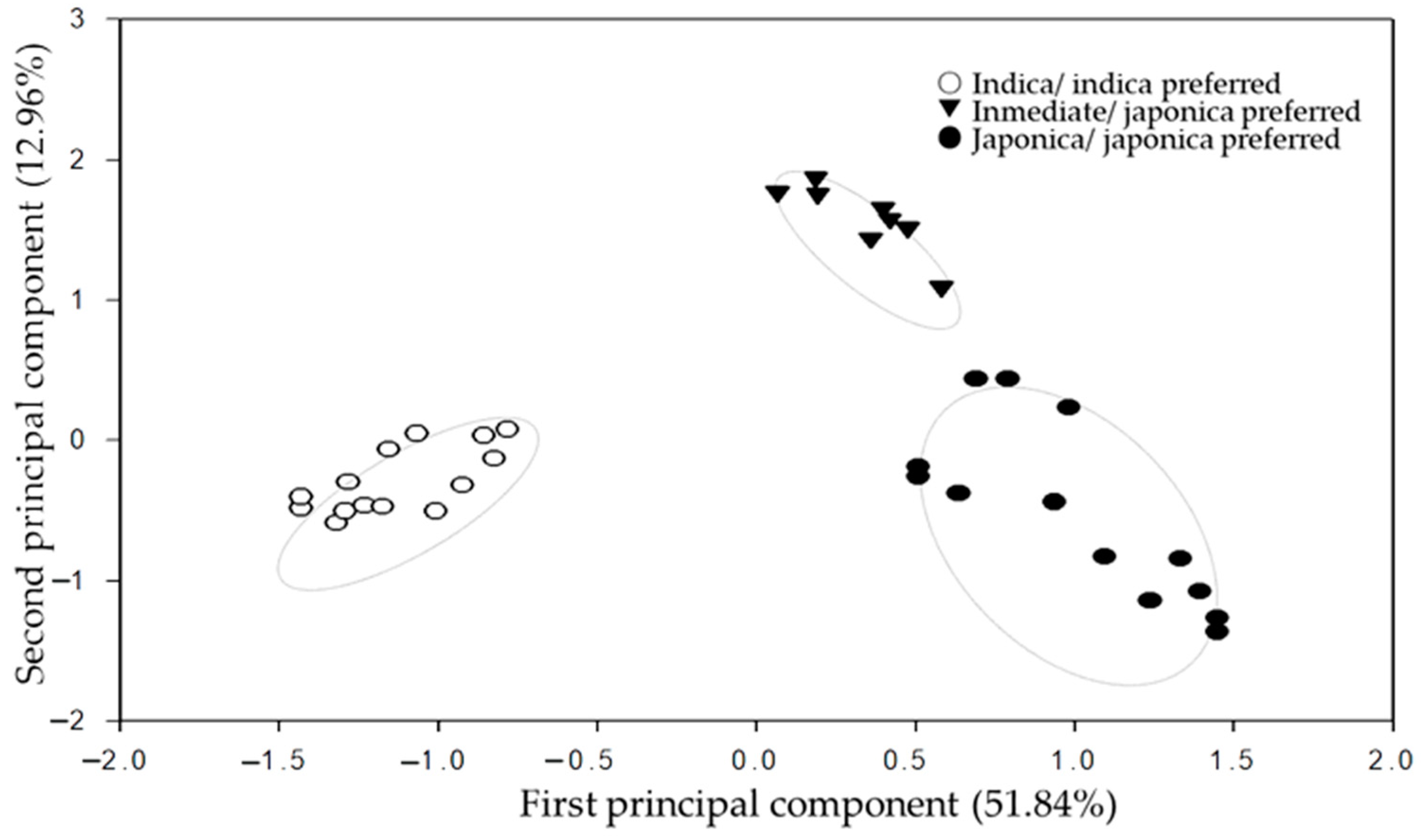
3.5. PCA Heterosis Correlation Analysis of InDel Molecular Distance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dan, Z.; Liu, P.; Huang, W.; Zhou, W.; Yao, G.; Hu, J.; Zhu, R.; Lu, B.; Zhu, Y. Balance between a Higher Degree of Heterosis and Increased Reproductive Isolation: A Strategic Design for Breeding Inter-Subspecific Hybrid Rice. PLoS ONE 2014, 9, e93122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G. The Next Generation of Rice: Inter-Subspecific Indica-Japonica Hybrid Rice. Front. Plant Sci. 2022, 13, 857896. [Google Scholar] [CrossRef] [PubMed]
- Ichihashi, Y.; Tsukaya, H. Behavior of Leaf Meristems and Their Modification. Front. Plant Sci. 2015, 6, 1060. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhang, H.; Wang, Q.; Guo, R.; Wei, L.; Song, H.; Kuang, W.; Liao, J.; Huang, Y.; Wang, Z. Genome-wide identification and characterization of long non-coding RNAs involved in flag leaf senescence of rice. Plant Mol. Biol. 2021, 105, 655–684. [Google Scholar] [CrossRef] [PubMed]
- Shang, C.; Guo, Z.; Chong, H.; Xiong, X.; Deng, J.; Harrison, M.; Liu, K.; Huang, L.; Tian, X.; Zhang, Y. Higher Radiation Use Efficiency and Photosynthetic Characteristics After Flowering Could Alleviate the Yield Loss of Indica-Japonica Hybrid Rice Under Shading Stress. Int. J. Plant Prod. 2022, 16, 105–117. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, X.; Han, B. Understanding the genetic basis of rice heterosis: Advances and prospects. Crop J. 2021, 9, 688–692. [Google Scholar] [CrossRef]
- Deng, J.; Sheng, T.; Zhong, X.; Ye, J.; Wang, C.; Huang, L.; Tian, X.; Liu, K.; Zhang, Y. Delayed Leaf Senescence Improves Radiation Use Efficiency and Explains Yield Advantage of Large Panicle-Type Hybrid Rice. Plants 2023, 12, 4063. [Google Scholar] [CrossRef]
- Chin, J.H.; Chu, S.H.; Jiang, W.; Cho, Y.I.; Basyirin, R.; Brar, D.S.; Koh, H.J. Identification of QTLs for hybrid fertility in inter-subspecific crosses of rice (Oryza sativa L.). Genes Genom. 2011, 33, 39–48. [Google Scholar] [CrossRef]
- Miura, K.; Ikeda, M.; Matsubara, A.; Song, X.J.; Ito, M.; Asano, K.; Matsuoka, M.; Kitano, H.; Ashikari, M. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nat. Genet. 2010, 42, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Yuan, X.; Wen, Y.; Yang, Y.; Ma, Y.; Yan, F.; Li, B.; Wang, Z.; Xing, M.; Zhang, R.; et al. Common population characteristics of direct-seeded hybrid indica rice for high yield. Agron. J. 2023, 115, 1606–1621. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.C.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F.; et al. Genomic variation in 3,010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Zhang, F.; Zafar, S.; Wang, J.; Lu, H.; Naveed, S.; Lou, J.; Xu, J. Genetic dissection of heterosis of indica–japonica by introgression line, recombinant inbred line and their testcross populations. Sci. Rep. 2021, 11, 10265. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.; Shi, P.J.; Royer, D.L.; Peppe, D.J.; Gallagher, R.V.; Li, Y.R.; Wang, R.; Wright, I.J. Leaf size estimation based on leaf length, width and shape. Ann. Bot. 2021, 128, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.; Lin, B.; Ren, Y.; Hu, H.; Xue, B.; Hua, S. Auxin-regulated timing of transition from vegetative to reproductive growth in rapeseed (Brassica napus L.) under different nitrogen application rates. Front. Plant. Sci. 2022, 13, 927662. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.; Qiu, C.; Wu, Y.; Liu, W.; Vincze, E.; Wu, F.; Hua, S. Secondary compost products improved tomato production via regulating primary/secondary metabolism and hormone signal interaction. Plant Growth Regul. 2024, 102, 167–178. [Google Scholar] [CrossRef]
- Jin, X.; Chen, Y.; Liu, P.; Li, C.; Cai, X.; Rong, J.; Lu, B. Introgression from cultivated rice alters genetic structures of wild relative populations: Implications for in situ conservation. AoB Plants 2018, 10, plx055. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.; Ren, Y.; Zhang, L.; Ge, R.; Wei, J. Analysis of Population Genetic Diversity and Genetic Structure of Schizothorax biddulphi Based on 20 Newly Developed SSR Markers. Front. Genet. 2022, 13, 908367. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalik, A.F.; Shishido, R.; Nomura, K.; Ikehashi, H. QTL-based analysis of leaf senescence in an indica/japonica hybrid in rice (Oryza sativa L.). Theor. Appl. Genet. 2005, 110, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Q.; Wang, H.R.; Xu, X.M.; Wang, M.; Wang, Y.H.; Chen, H.; Ping, F.; Zhong, H.; Mu, Z.; Xie, W.; et al. Two complementary genes in a presence-absence variation contribute to indica-japonica reproductive isolation in rice. Nat. Commun. 2023, 14, 4532. [Google Scholar] [CrossRef]
- Shen, G.; Hu, W.; Zhang, B.; Xing, Y. The regulatory network mediated by circadian clock genes is related to heterosis in rice. J. Integr. Plant Biol. 2015, 57, 300–312. [Google Scholar] [CrossRef]
- Guo, J.; Li, Y.; Xiong, L.; Yan, T.X.; Zuo, J.S.; Dai, Z.J.; Tang, G.; Sun, K.L.; Luan, X.; Yang, W.F.; et al. Development of Wide-Compatible Indica Lines by Pyramiding Multiple Neutral Alleles of Indica-Japonica Hybrid Sterility Loci. Front. Plant Sci. 2022, 13, 890568. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Zhang, Y.; Xiang, J.; Zhang, Y.; Zhu, D.; Chen, H. The nitrogen topdressing mode of indica-japonica and indica hybrid rice are different after side-deep fertilization with machine transplanting. Sci. Rep. 2021, 11, 1494. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Deng, X.; Yang, J.; Zhang, C.; Sun, T.; Ji, W.; Zhong, H.; Fu, X.; Li, Y. High-resolution bin-based linkage mapping uncovers the genetic architecture and heterosis-related loci of plant height in indica–japonica derived populations. Plant J. 2022, 110, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J. Transcriptome and DNA methylome analyses provide insight into the heterosis in flag leaf of inter-subspecific hybrid rice. Plant Mol. Biol. 2021, 108, 105–125. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Primary Panicle | Intermediate Panicle | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SNP | FNP | PB | SNPB | FNPB | SS (%) | SNP | FNP | PB | SNPB | FNPB | SS (%) | |
| Zhe04B | 186 d | 169 c | 16 de | 11.63 c | 10.56 bc | 90.86 a | 177 c | 148 d | 13 c | 13.62 a | 11.38 a | 83.62 b |
| Zhehui 818 | 280 b | 244 b | 21 b | 13.33 b | 11.62 ab | 87.14 a | 247 c | 230 c | 18 b | 13.72 a | 12.78 a | 93.12 a |
| Zheyou12 | 240 c | 179 c | 18 cd | 13.33 b | 9.94 c | 74.58 c | 182 d | 152 d | 16 b | 11.38 b | 9.50 a | 83.52 b |
| LYPJ | 234 c | 172 c | 14 e | 16.71 a | 12.29 a | 73.50 c | 185 d | 146 d | 13 c | 14.23 a | 11.23 a | 78.92 b |
| Zheyou18 | 398 a | 323 a | 26 a | 15.31 a | 12.42 a | 81.16 b | 324 b | 264 b | 23 a | 14.09 a | 11.48 a | 81.48 b |
| Yongyou12 | 406 a | 326 a | 26 a | 15.62 a | 12.54 a | 80.30 b | 379 a | 310 a | 26 a | 14.58 a | 11.92 a | 81.79 b |
| Yield Related Traits | Control Heterosis | Correlation Coefficient between Genetic Distance and Traits | ||||
|---|---|---|---|---|---|---|
| Range/% | Mean | No. of Combinations with Positive (Negative) Heterosis | No. of Combinations with Significance at 0.01 Level | The 12 Combinations Analyzed as a Whole | The 8 Indica–Japonica Combinations Analyzed as a Whole | |
| Grain yield | 1.89~29.97 | 20.09 | 12 (0) | 8 (−) | 0.819 ** | 0.474 |
| Panicles per plant | −0.97~−39.12 | −29.36 | 0 (12) | 1 (−) | −0.522 | −0.685 |
| Total grains per plant | 36.31~173.75 | 108.78 | 12 (0) | 1 (+) | 0.845 ** | 0.599 |
| Filled grains per plant | 24.95~126.40 | 79.08 | 12 (0) | 9 (+) | 0.834 ** | 0.633 |
| Seed setting rates | −19.24~−3.19 | −13.32 | 0 (12) | 9 (−) | −0.702 * | −0.010 |
| 1000-grain weight | −9.20~10.80 | −3.20 | 2 (10) | 3 (−) | −0.349 | 0.710 * |
| Panicle weight | 20.45~124.59 | 72.30 | 12 (0) | 11 (+) | 0.863 * | 0.771 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Cui, Y.; Song, J.; Fan, H.; Tang, L.; Wang, J. Preliminary Exploration of Physiology and Genetic Basis Underlying High Yield in Indica–Japonica Hybrid Rice. Agriculture 2024, 14, 607. https://doi.org/10.3390/agriculture14040607
Fan X, Cui Y, Song J, Fan H, Tang L, Wang J. Preliminary Exploration of Physiology and Genetic Basis Underlying High Yield in Indica–Japonica Hybrid Rice. Agriculture. 2024; 14(4):607. https://doi.org/10.3390/agriculture14040607
Chicago/Turabian StyleFan, Xiaojuan, Yongtao Cui, Jian Song, Honghuan Fan, Liqun Tang, and Jianjun Wang. 2024. "Preliminary Exploration of Physiology and Genetic Basis Underlying High Yield in Indica–Japonica Hybrid Rice" Agriculture 14, no. 4: 607. https://doi.org/10.3390/agriculture14040607
APA StyleFan, X., Cui, Y., Song, J., Fan, H., Tang, L., & Wang, J. (2024). Preliminary Exploration of Physiology and Genetic Basis Underlying High Yield in Indica–Japonica Hybrid Rice. Agriculture, 14(4), 607. https://doi.org/10.3390/agriculture14040607