
Exogenous Riboflavin Application at Different Growth Stages Regulates Photosynthetic Accumulation and Grain Yield in Fragrant Rice

Abstract
1. Introduction
2. Materials and Methods
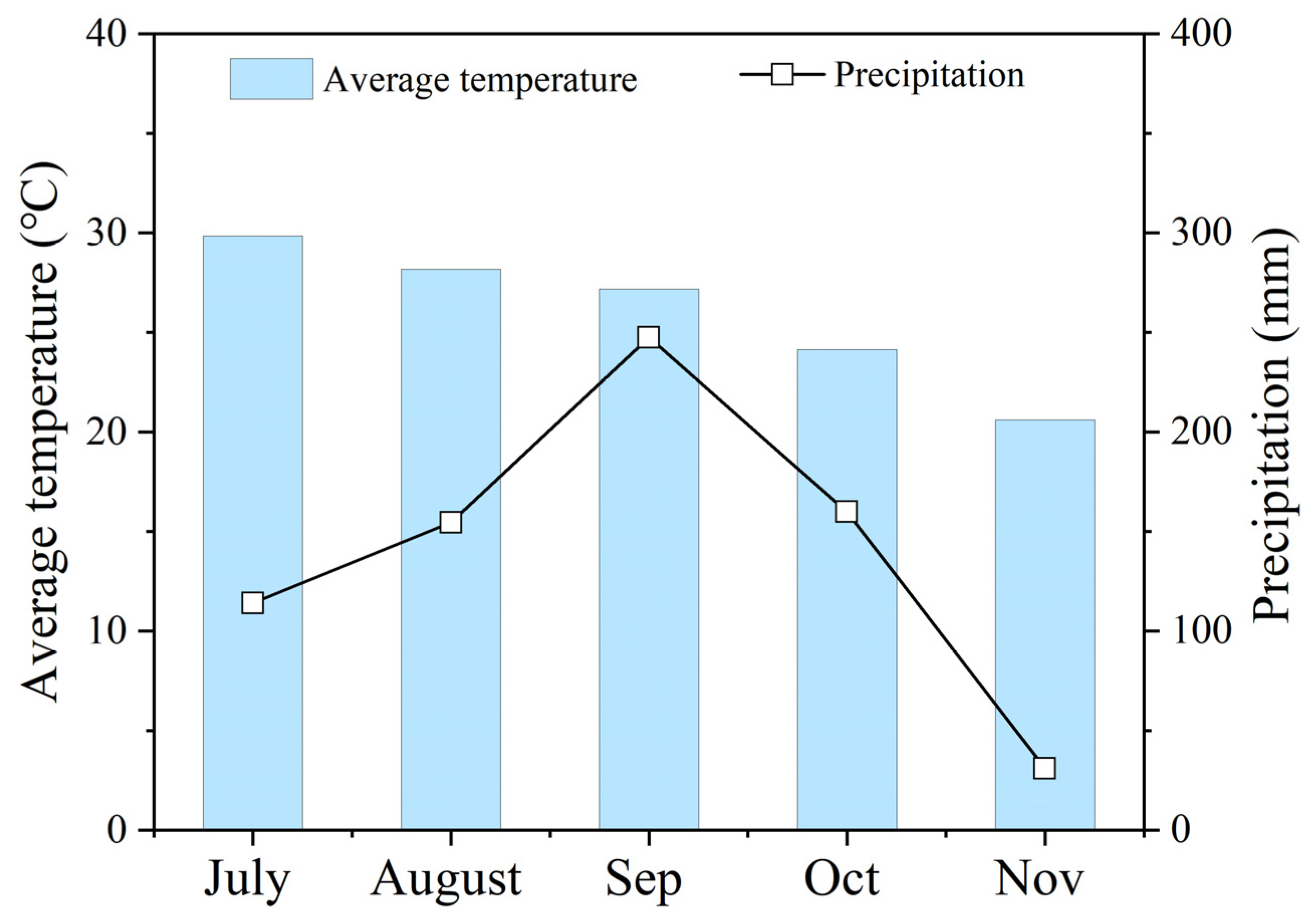
2.1. Experimental Design and Management
2.2. Sampling and Measurement
2.2.1. Determination of Dry Matter Accumulation and the Grain Filling Rate
2.2.2. Determination of Chlorophyll and Carotenoid Contents
2.2.3. Determination of Related Enzymatic Activities
2.2.4. Determination of Grain Yield and Its Components
2.2.5. Determination of Carbohydrate Contents
2.2.6. Statistical Analysis
3. Results
3.1. Yield and Its Components
3.2. Total Aboveground Biomass Accumulation
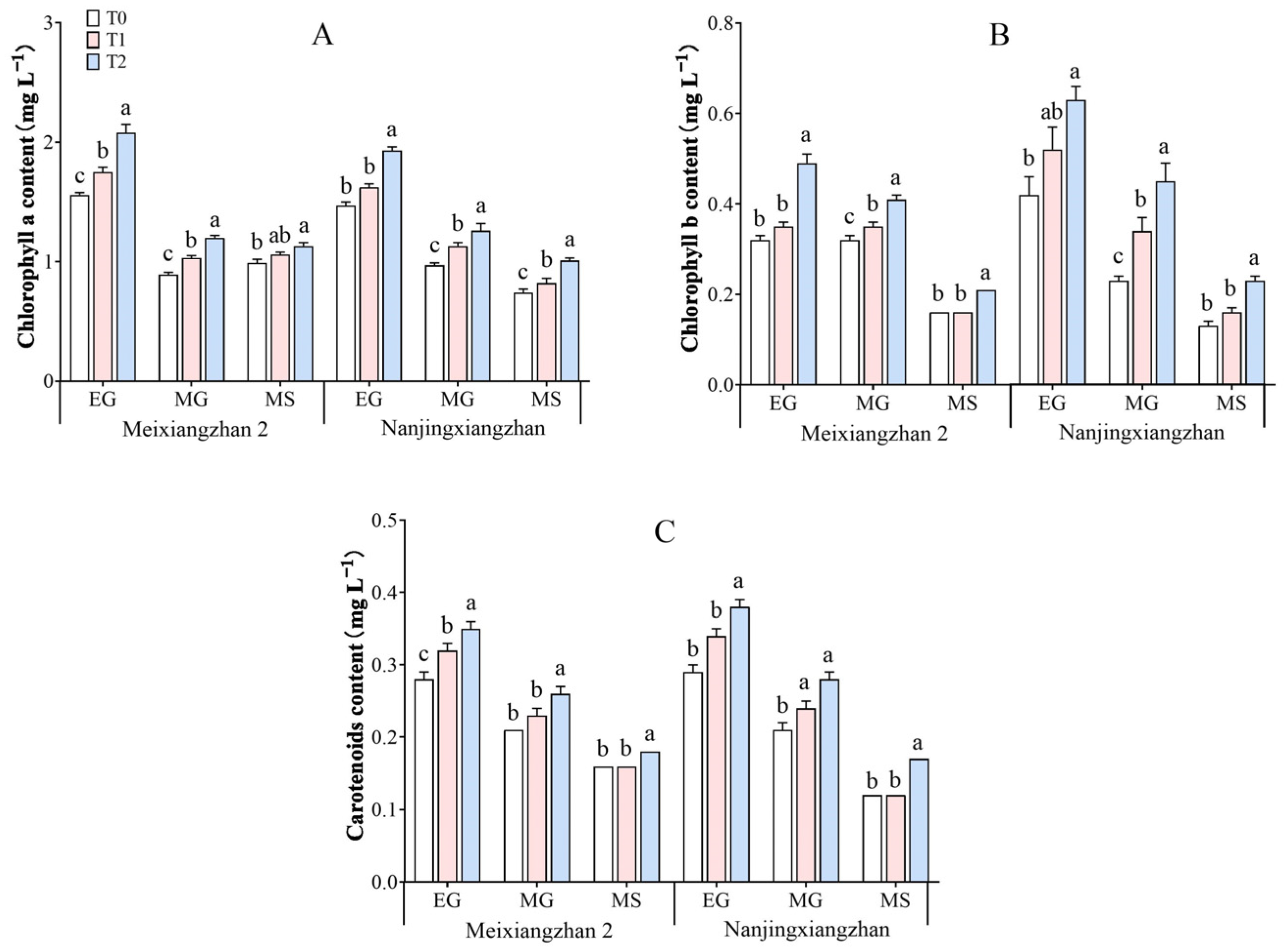
3.3. Photosynthetic Pigment Content
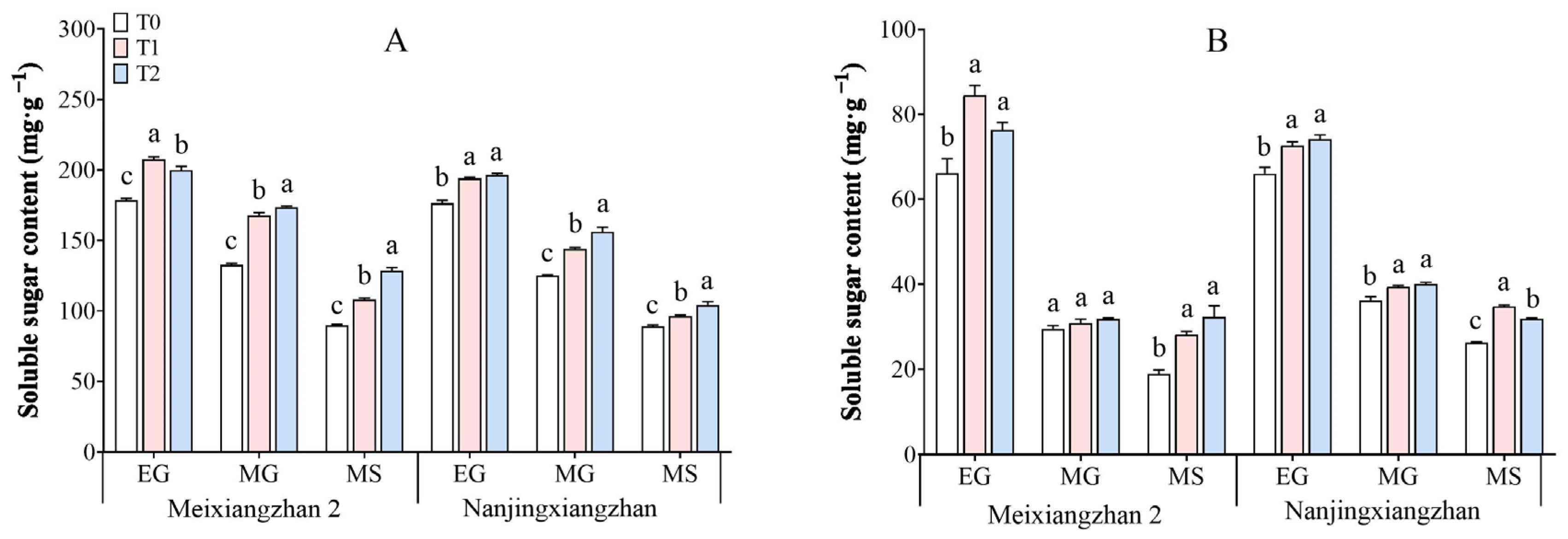
3.4. Soluble Sugar Content
3.5. Sucrose Content
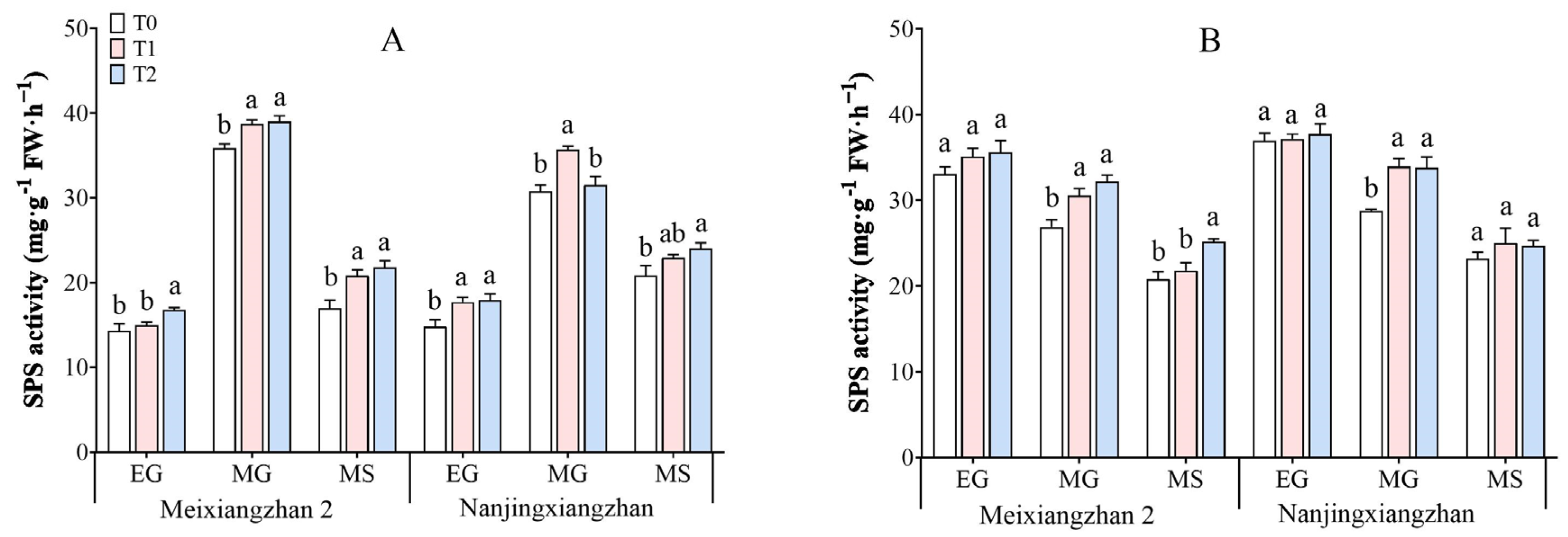
3.6. Sucrose–Phosphate Synthase (SPS) Activity
3.7. Sucrose Synthase (SS) Activity
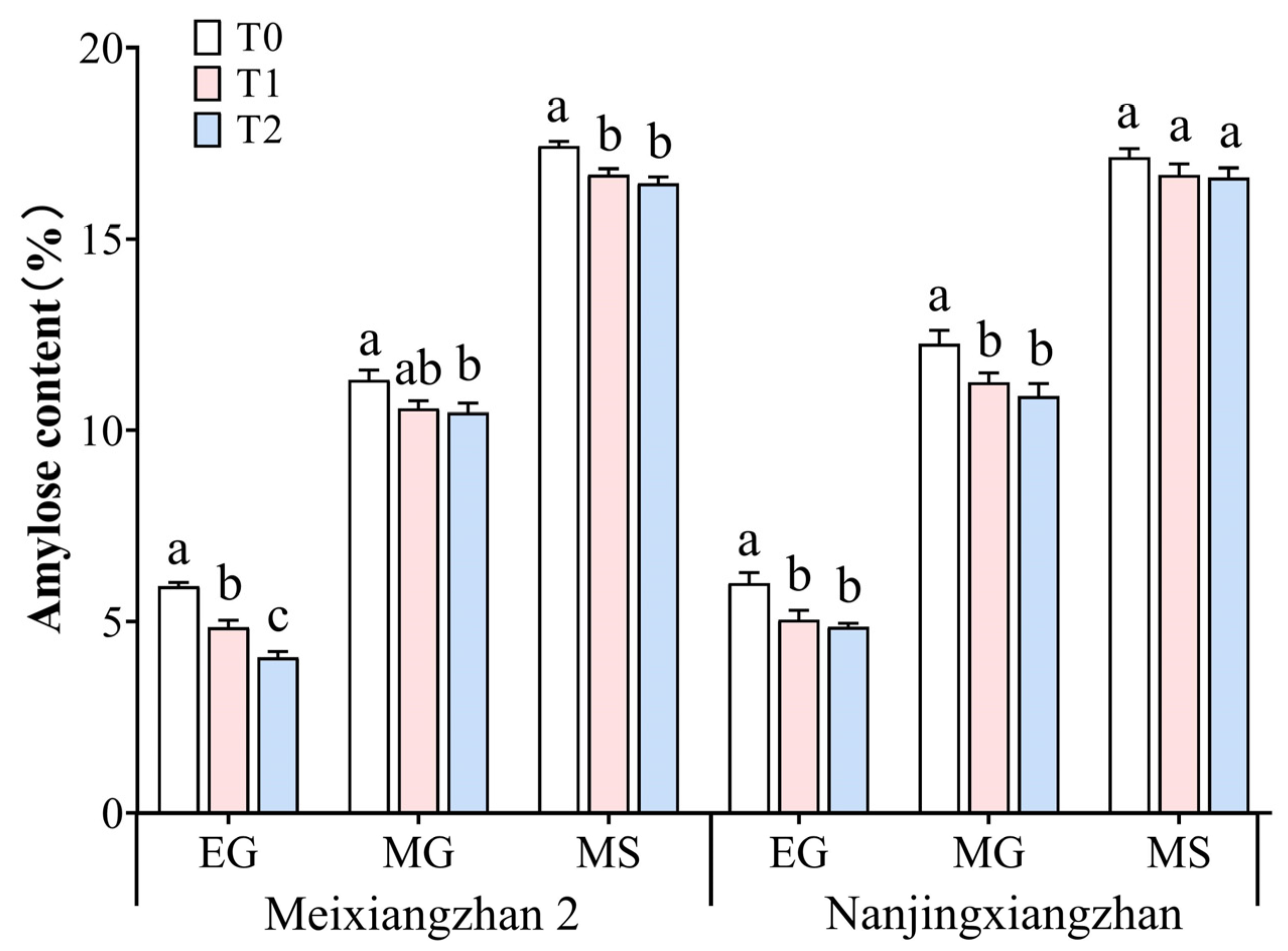
3.8. Amylose Content
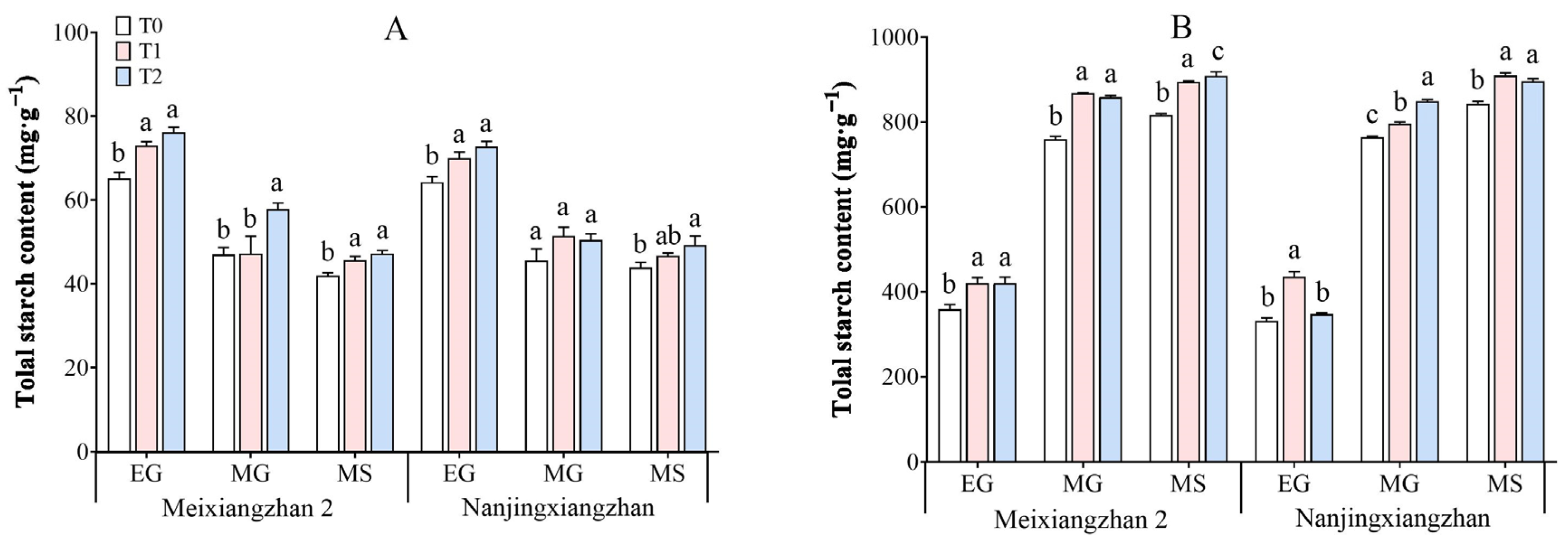
3.9. Total Starch Content
3.10. Panicle and Grain Weight Dynamics
3.11. Regression Analysis and Heatmaps for Various Indexes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sakthivel, K.; Sundaram, R.M.; Rani, N.S.; Balachandran, S.M.; Neeraja, C.N. Genetic and molecular basis of fragrance in rice. Biotechnol. Adv. 2009, 27, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Custodio, M.C.; Cuevas, R.P.; Ynion, A.G.; Velasco, M.L.; Demont, M. Rice quality: How is it defined by consumers, industry, food scientists, and geneticists? Trends Food Sci. Technol. 2019, 92, 122–137. [Google Scholar] [CrossRef] [PubMed]
- United Nations. Department of economic and social affairs, population division. In World Population Prospects; United Nations: New York, NY, USA, 2019; Volume 2017. [Google Scholar]
- You, L.Z.; Sun, Z.L. Mapping global cropping system: Challenges, opportunities, and future perspectives. Crop Environ. 2022, 1, 68–73. [Google Scholar] [CrossRef]
- Liu, Q.H.; Cai, J.; Li, T. Key starch synthase in rice grain and its relation to grain filling and rice quality. Plant Physiol. Agric. Appl. 2006, 6, 1211–1216. (In Chinese) [Google Scholar]
- Doehlert, D.C.; Kuo, T.M.; Felker, F.C. Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize. Plant Physiol. 1988, 86, 1013–1019. [Google Scholar] [CrossRef]
- Ball, S.G.; van de Wal, M.H.B.J.; Visser, R.G.F. Progress in understanding the biosynthesis of amylose. Trends Plant Sci. 1998, 3, 462–467. [Google Scholar] [CrossRef]
- Seung, D. Amylose in starch: Toward an understanding of biosynthesis, structure and function. New Phytol. 2020, 228, 1490–1504. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Zheng, F.Q.; Shen, G.Z.; Gao, J.P.; Snustad, D.P.; Li, M.G.; Zhang, J.L.; Hong, M.M. The amylose content in rice endosperm is related to the post-transcriptional regulation of the waxy gene. Plant J. 1995, 7, 613–622. [Google Scholar] [CrossRef]
- Liang, J.S.; Zhang, J.H.; Cao, X.Z. Grain sink strength may be related to the poor grain filling of indica-japonica (Oryza sativa) hybrids. Physiol. Plant. 2001, 112, 470–477. [Google Scholar] [CrossRef]
- Lisle, A.J.; Martin, M.; Fitzgerald, M.A. Chalky and translucent rice grains differ in starch composition and structure and cooking properties. Cereal Chem. 2000, 77, 627–632. [Google Scholar] [CrossRef]
- Ishimaru, T.; Horigane, A.K.; Ida, M.; Iwasawa, N.; Yoshida, M. Formation of grain chalkiness and changes in water distribution in developing rice caryopses grown under high-temperature stress. J. Cereal Sci. 2009, 50, 166–174. [Google Scholar] [CrossRef]
- Guo, C.C.; Yuan, X.J.; Yan, F.J.; Xiang, K.H.; Wu, Y.X.; Zhang, Q.; Wang, Z.L.; He, L.M.; Fan, P.; Yang, Z.Y.; et al. Nitrogen application rate affects the accumulation of carbohydrates in functional leaves and grains to improve grain filling and reduce the occurrence of chalkiness. Front. Plant Sci. 2022, 13, 921130. [Google Scholar] [CrossRef] [PubMed]
- Survase, S.A.; Bajaj, I.B.; Singhal, R.S. Biotechnological production of vitamins. Food Technol. Biotechnol. 2006, 44, 381–396. [Google Scholar]
- Asensi-Fabado, M.A.; Munné-Bosch, S. Vitamins in plants: Occurrence, biosynthesis and antioxidant function. Trends Plant Sci. 2010, 15, 582–592. [Google Scholar] [CrossRef]
- Palacios, O.A.; Bashan, Y.; De-Bashan, L.E. Proven and potential involvement of vitamins in interactions of plants with plant growth-promoting bacteria—An overview. Biol. Fertil. Soils 2014, 50, 415–432. [Google Scholar] [CrossRef]
- Cheng, S.R.; Liu, H.D.; Li, K.Q.; Zheng, L.K.; Su, M.L.; Lin, X.E.; Huang, G.B.; Ren, Y. Riboflavin improves grain yield, 2-acetyl-1-pyrroline accumulation, and antioxidative properties of fragrant rice. J. Sci. Food Agric. 2024, 104, 1178–1189. [Google Scholar] [CrossRef]
- Pinto, J.; Zempleni, J. Riboflavin. Adv. Nutr. 2016, 7, 973–975. [Google Scholar] [CrossRef]
- Whatley, F.R.; Allen, M.B.; Arnon, D.L. Photosynthesis by isolated chloroplasts. VII. Vitamin K and riboflavin phosphate as cofactors of cyclic photophosphorylation. BBA 1959, 32, 32–46. [Google Scholar]
- Deng, B.L.; Jin, X.H.; Yang, Y.; Lin, Z.W.; Zhang, Y.L. The regulatory role of riboflavin in the drought tolerance of tobacco plants depends on ROS production. Plant Growth Regul. 2014, 72, 269–277. [Google Scholar] [CrossRef]
- Hu, H.T.; Ren, D.Y.; Hu, J.; Jiang, H.Z.; Chen, P.; Zeng, D.L.; Qian, Q.; Guo, L.B. WHITE AND LESION-MIMIC LEAF1, encoding a lumazine synthase, affects reactive oxygen species balance and chloroplast development in rice. Plant J. 2021, 108, 1690–1703. [Google Scholar] [CrossRef]
- Tian, Q.Z.; Wang, G.; Ma, X.X.; Shen, Q.W.; Ding, M.L.; Yang, X.Y.; Luo, X.L.; Li, R.R.; Wang, Z.H.; Wang, X.H.; et al. Riboflavin integrates cellular energetics and cell cycle to regulate maize seed development. Plant Biotechnol. J. 2022, 20, 1487–1501. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.L.; Zhao, J.; Pan, X.M.L.; Zhao, J.F.; Dong, H.S.; Wang, J.S.; Liu, B.X.; Liu, G.Y.; Cheng, Y.J. Riboflavin activates growth signal transduction pathway in plants. J. Nanjing Agric. Univ. 2002, 4, 33–36. (In Chinese) [Google Scholar]
- Zhu, Q.; Cao, X.; Luo, Y. Growth analysis of rice grain filling. Acta Agron. Sin. 1988, 14, 182–193. (In Chinese) [Google Scholar]
- Tang, X. Crop Cultivation and Physiological Experiment Guidance; Guangdong Higher Education Press: Guangzhou, China, 2021. [Google Scholar]
- Nakamura, Y.; Yuki, K.; Park, S.Y.; Ohya, T. Carbohydrate metabolism in the developing endosperm of rice grains. Plant Cell Physiol. 1989, 30, 833–839. [Google Scholar] [CrossRef]
- Dai, L.; Ren, Y.; Ashraf, U.; Gui, R.F.; Wang, Z.M.; Tang, X.R.; Duan, M.Y.; Mo, Z.W. Optimized liquid fertilizer management increases 2-acetyl-1-pyrroline content and grain quality in aromatic rice. J. Food Compos. Anal. 2024, 133, 106433. [Google Scholar] [CrossRef]
- Jun, Z.; Peng, D.; Zhang, H.Z.; Meng, C.R.; Zhang, X.J.; Hou, J.W.; Wei, C.Z. Low soil temperature reducing the yield of drip irrigated rice in arid area by influencing anther development and pollination. J. Arid Land 2019, 11, 419–430. [Google Scholar]
- Powers, H.J. Riboflavin (vitaminB-2) and health. Am. J. Clin. Nutr. 2003, 77, 1352–1360. [Google Scholar] [CrossRef]
- Massey, V. The chemical and biological versatility of riboflavin. Biochem. Soc. Trans. 2000, 28, 283–296. [Google Scholar] [CrossRef]
- Peng, J.; Zhao, J.; Pan, X.; Zhao, J.; Dong, H. Preliminary study on the activation of plant growth signaling pathways by riboflavin. J. Nanjing Agric. Univ. 2002, 4, 33–36. (In Chinese) [Google Scholar]
- Namba, J.; Harada, M.; Shibata, R.; Toda, Y.; Maruta, T.; Ishikawa, T.; Shigeoka, S.; Yoshimura, K.; Ogawa, T. AtDREB2G is involved in the regulation of riboflavin biosynthesis in Arabidopsis thaliana. Plant Sci. 2024, 347, 112196. [Google Scholar] [CrossRef]
- Wilkie, K.; Wootton, M.; Paton, J.E. Sensory testing of Australian fragrant, imported fragrant, and non-fragrant rice aroma. Int. J. Food Prop. 2004, 7, 27–36. [Google Scholar] [CrossRef]
- Ben-Asher, J.; Tsuyuki, I.; Bravdo, B.A.; Sagih, M. Irrigation of grapevines with saline water: I. Leaf area index, stomatal conductance, transpiration and photosynthesis. Agric. Water Manag. 2006, 83, 13–21. [Google Scholar] [CrossRef]
- Hoshida, H.; Tanaka, Y.; Hibino, T.; Hayashi, Y.; Tanaka, A.; Takabe, T.; Takabe, T. Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast glutamine synthetase. Plant Mol. Biol. 2000, 43, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Huang, Y.; Huang, J.; Xia, G.; Gong, N. Comparative study on the growth and physiological characteristics of plants with different salt tolerance under salt stress. J. Zhejiang Univ. (Agric. Life Sci.) 2006, 4, 420–427. (In Chinese) [Google Scholar]
- Li, X.; Liu, B.; Guo, Z.; Chang, Y.; He, L.; Chen, F.; Lu, B. Changes in photosynthetic characteristics and rapid chlorophyll fluorescence induction kinetics of Populus euphratica leaves under NaCl stress. Chin. J. Appl. Ecol. 2013, 24, 2479–2484. (In Chinese) [Google Scholar]
- Engelaar, W.M.H.G.; Matsumaru, T.; Yoneyama, T. Combined effects of soil water logging and compaction on rice (Oryza sativa L.) growth, soil aeration, soil N transformations and 15 N discrimination. Biol. Fertil. Soils 2000, 32, 484–493. [Google Scholar] [CrossRef]
- Luo, H.; He, L.; Du, B.; Wang, Z.; Zheng, A.; Lai, R.; Tang, X. Foliar application of selenium (Se) at heading stage induces regulation of photosynthesis, yield formation, and quality characteristics in fragrant rice. Photosynthetica 2019, 57, 1007–1014. [Google Scholar] [CrossRef]
- Wu, Q.X.; Du, B.; Jiang, S.C.; Zhang, H.W.; Zhu, J.Q. Side deep fertilizing of machine-transplanted rice to guarantee rice yield in conservation tillage. Agriculture 2022, 12, 528. [Google Scholar] [CrossRef]
- Hu, Q.; Jiang, W.Q.; Qiu, S.; Xing, Z.P.; Hu, Y.J.; Guo, B.W.; Liu, G.D.; Gao, H.; Zhang, H.C.; Wei, H.Y. Effect of wide-narrow row arrangement in mechanical pot-seedling transplanting and plant density on yield formation and grain quality of japonica rice. J. Integr. Agric. 2020, 19, 1197–1214. [Google Scholar] [CrossRef]
- An, N.; Zhang, L.; Liu, Y.X.; Shen, S.; Li, N.; Wu, Z.C.; Yang, J.F.; Han, W.; Han, X.R. Biochar application with reduced chemical fertilizers improves soil pore structure and rice productivity. Chemosphere 2022, 298, 134304. [Google Scholar] [CrossRef]
- Wie, H.Y.; Hu, L.; Zhu, Y.; Xu, D.; Zheng, L.M.; Chen, Z.F.; Hu, Y.J.; Cui, P.Y.; Guo, B.W.; Dai, Q.G.; et al. Different characteristics of nutrient absorption and utilization between inbred Japonica super rice and inter-sub-specific hybrid super rice. Field Crop. Res. 2018, 218, 88–96. [Google Scholar]
- Wu, H.; Xiang, J.; Zhang, Y.P.; Zhang, Y.K.; Peng, S.B.; Chen, H.Z.; Zhu, D.F. Effects of post-anthesis nitrogen uptake and translocation on photosynthetic production and rice yield. Sci. Rep. 2018, 8, 2045–2322. [Google Scholar] [CrossRef] [PubMed]
- You, C.C.; Chen, L.; He, H.B.; Wu, L.Q.; Wang, S.H.; Ding, Y.F.; Ma, C.X. iTRAQ-based proteome profile analysis of superior and inferior spikelets at early grain filling stage in japonica rice. BMC Plant Biol. 2017, 17, 100. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Shon, J.; Lee, C.K.; Yang, W.; Yoon, Y.; Yang, W.H.; Kim, Y.G.; Lee, B.W. Relationship between grain filling duration and leaf senescence of temperate rice under high temperature. Field Crop. Res. 2011, 122, 207–213. [Google Scholar] [CrossRef]
- Ahmed, N.; Tetlow, I.; Nawaz, S.; Iqbal, A.; Mubin, M.; Shah, N.; Butt, A.; Lightfoot, D.; Maekawa, M. Effect of high temperature on grain filling period, yield, amylose content and activity of starch biosynthesis enzymes in endosperm of basmati rice. J. Sci. Food Agric. 2015, 95, 2237–2243. [Google Scholar] [CrossRef]
- Pasuquin, E.M.; Eberbach, P.L.; Hasegawa, T.; Lafarge, T.; Harnpichitvitaya, D.; Wade, L.J. Responses to elevated daytime air and canopy temperature during panicle development in four rice genotypes under paddy conditions in large field chambers. Crop Environ. 2023, 2, 147–156. [Google Scholar] [CrossRef]
- Taub, D.R.; Seemann, J.R.; Coleman, J.S. Growth in elevated CO2 protects photosynthesis against high-temperature damage. Plant Cell Environ. 2000, 23, 649–656. [Google Scholar] [CrossRef]
- Lu, G.H.; Wu, Y.F.; Bai, W.B.; Ma, B.; Wang, C.Y.; Song, J.Q. Influence of high temperature stress on net photosynthesis, dry matter partitioning and rice grain yield at flowering and grain filling stages. J. Integr. Agric. 2013, 12, 603–609. [Google Scholar] [CrossRef]
- Xu, H.F.; Wang, Z.X.; Xiao, F.; Yang, L.; Li, G.H.; Ding, Y.F.; Paul, M.J.; Li, W.W.; Liu, Z.H. Dynamics of dry matter accumulation in internodes indicates source and sink relations during grain-filling stage of japonica rice. Field Crop. Res. 2021, 263, 108009. [Google Scholar] [CrossRef]
- Wu, Q.; Wu, F.; Hu, Y.; Kang, Z.; Zhang, Y.; Yang, J.; Yue, K.; Ni, X.; Yang, Y. Comparison of nonstructural carbohydrate contents in young and old leaves of 11 tree species in a subtropical homogeneous garden. Acta Ecol. Sin. 2021, 45, 771–779. (In Chinese) [Google Scholar]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Pérez-Garcia, M.D.; Ogé, L.; Hamama, L.; Atanassova, R. The sugar-signaling hub: Overview of regulators and interaction with the hormonal and metabolic network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yin, F.; Xu, K.; Jia, X.; Zhou, S.; Ma, C. The role of sucrose metabolism and signaling in plant development and stress responses. Agric. Sci. Technol. 2021, 35, 2044–2055. (In Chinese) [Google Scholar]
- Ye, X.Q.; Meng, J.L.; Zeng, B.; Ming, W. Improved flooding tolerance and carbohydrate status of flood-tolerant plant Arundinella anomala at lower water temperature. PLoS ONE 2018, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, H.P.; Batish, D.P.; Kaur, S.; Kohli, R.K. EMF radiations (1800 MHz)-inhibited early seedling growth of maize (Zea mays) involves alterations in starch and sucrose metabolism. Protoplasma 2016, 253, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Lee, J.; Kichey, T.; Gerentes, D.; Zivy, M.; Tatout, C.; Dubois, F.; Balliau, T.; Valot, B.; Davanture, M.; et al. Two cytosolic glutamine synthetase isoforms of maize are specifically involved in the control of grain production. Plant Cell 2006, 18, 3252–3274. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Treatments | Panicle Number per Hill | Grains Number per Panicle | Grain Filling (%) | 1000-Grain Weight (g) | Yield (g·Pot−1) |
|---|---|---|---|---|---|---|
| Meixiangzhan 2 | T0 | 13.29 a | 118.50 a | 78.74 b | 17.19 b | 72.35 c |
| T1 | 13.27 a | 117.75 a | 82.15 a | 17.20 b | 77.60 b | |
| T2 | 13.18 a | 122.75 a | 82.65 a | 17.67 a | 82.34 a | |
| Mean | 13.24 | 119.50 | 81.18 | 17.35 | 78.10 | |
| Nanjingxiangzhan | T0 | 12.59 a | 135.25 a | 79.61 b | 19.77 b | 87.54 b |
| T1 | 12.57 a | 133.25 a | 83.35 a | 19.82 b | 93.28 ab | |
| T2 | 12.40 a | 132.50 a | 83.48 a | 20.53 a | 96.75 a | |
| Mean | 12.52 | 133.67 | 82.45 | 20.04 | 92.52 |
| Varieties (V) | Treatment (T) | V × T | |
|---|---|---|---|
| Yield | * | ** | ns |
| Productive panicle | ** | ns | ns |
| Spikelet per panicle | ns | ns | ns |
| Grain filling | ns | ** | ns |
| 1000-grain weight | ** | ** | ns |
| TAB—EG | ** | ns | ns |
| TAB—MG | ns | * | ns |
| TAB—MS | ns | ** | * |
| Total starch—EG | ns | ** | ns |
| Total starch—MG | ns | * | ns |
| Total starch—MS | ns | ** | ns |
| Amylose content—EG | * | ** | ** |
| Amylose content—MG | * | ** | ** |
| Amylose content—MS | * | ** | * |
| Sucrose—EG—L | ns | ** | ** |
| Sucrose—MG—L | * | * | ns |
| Sucrose—MS—L | * | ** | ** |
| Sucrose—EG—G | ** | ** | ** |
| Sucrose—MG—G | ** | ** | ** |
| Sucrose—MS—G | * | ** | ** |
| Soluble Sugar—EG—L | * | ** | * |
| Soluble Sugar—MG—L | ** | ** | ** |
| Soluble Sugar—MS—L | ** | ** | ** |
| Soluble Sugar—EG—G | ns | ** | ** |
| Soluble Sugar—MG—G | ** | ** | ns |
| Soluble Sugar—MS—G | ** | ** | ** |
| SPS—EG—L | * | ** | ns |
| SPS—MG—L | ** | ** | ** |
| SPS—MS—L | * | ** | ns |
| SPS—EG—G | ** | ns | ns |
| SPS—MG—G | ** | ** | ns |
| SPS—MS—G | ns | * | ns |
| SS—EG—L | ns | ** | ** |
| SS—MG—L | ns | ** | ns |
| SS—MS—L | * | ** | ns |
| SS—EG—G | * | ** | ns |
| SS—MG—G | * | ** | ns |
| SS—MS—G | * | ** | ns |
| Chlorophyll a—EG | ns | ** | ns |
| Chlorophyll a—MG | * | ** | ns |
| Chlorophyll a—MS | ** | ** | ** |
| Chlorophyll b—EG | * | ** | ns |
| Chlorophyll b—MG | ns | ** | ** |
| Chlorophyll b—MS | ns | ** | ns |
| Carotenoids—EG | ns | ** | ns |
| Carotenoids—MG | * | ** | ns |
| Carotenoids—MS | ** | ** | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Cheng, S.; Ding, X.; Lin, X.; Deng, S.; Peng, L.; Tian, H. Exogenous Riboflavin Application at Different Growth Stages Regulates Photosynthetic Accumulation and Grain Yield in Fragrant Rice. Agriculture 2024, 14, 1979. https://doi.org/10.3390/agriculture14111979
Wu Y, Cheng S, Ding X, Lin X, Deng S, Peng L, Tian H. Exogenous Riboflavin Application at Different Growth Stages Regulates Photosynthetic Accumulation and Grain Yield in Fragrant Rice. Agriculture. 2024; 14(11):1979. https://doi.org/10.3390/agriculture14111979
Chicago/Turabian StyleWu, Yizhu, Siren Cheng, Xinfang Ding, Xueer Lin, Sicheng Deng, Ligong Peng, and Hua Tian. 2024. "Exogenous Riboflavin Application at Different Growth Stages Regulates Photosynthetic Accumulation and Grain Yield in Fragrant Rice" Agriculture 14, no. 11: 1979. https://doi.org/10.3390/agriculture14111979
APA StyleWu, Y., Cheng, S., Ding, X., Lin, X., Deng, S., Peng, L., & Tian, H. (2024). Exogenous Riboflavin Application at Different Growth Stages Regulates Photosynthetic Accumulation and Grain Yield in Fragrant Rice. Agriculture, 14(11), 1979. https://doi.org/10.3390/agriculture14111979