


Portuguese Castanea sativa Genetic Resources: Characterization, Productive Challenges and Breeding Efforts



Abstract
1. Introduction
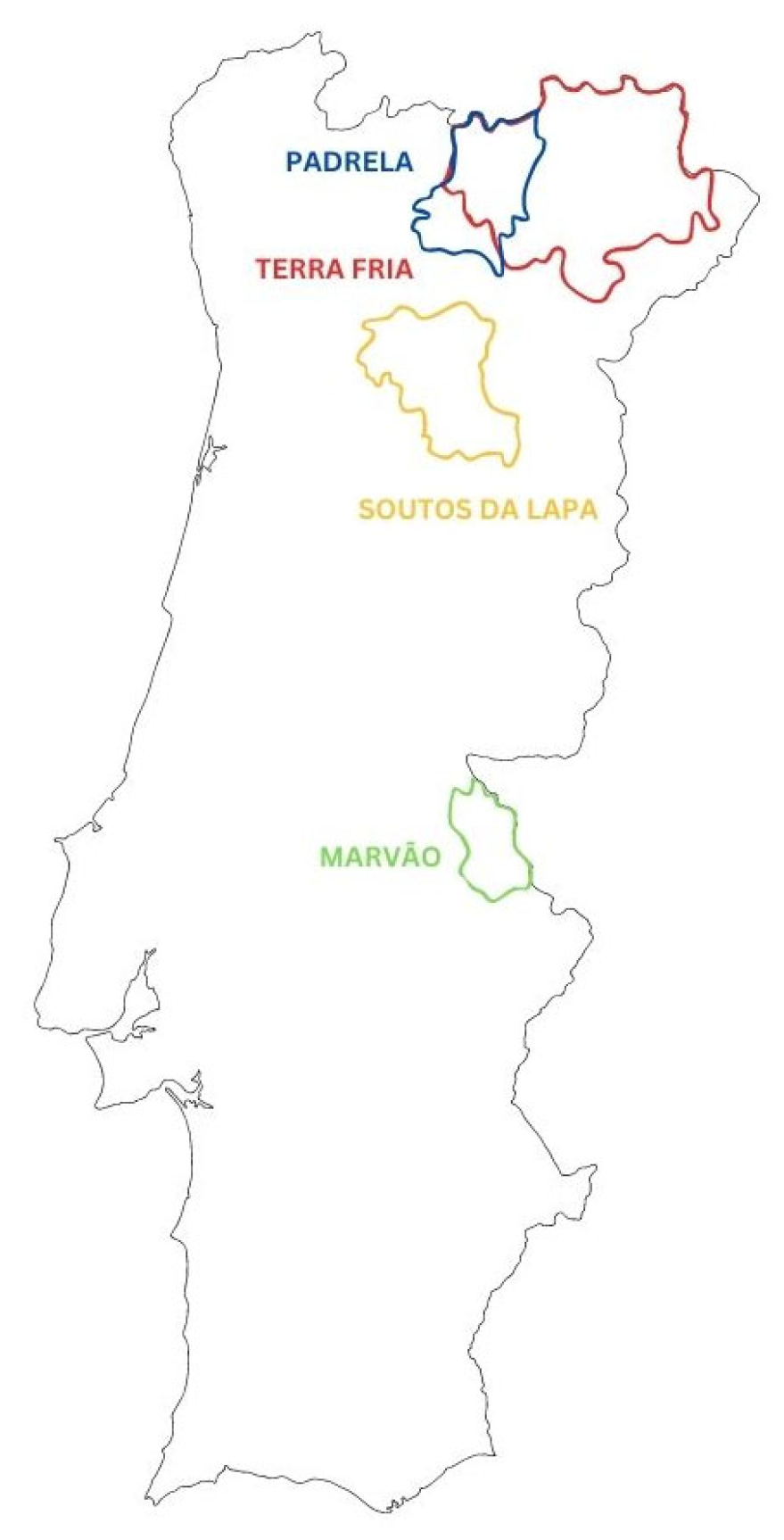
2. Chestnut Varieties in Portugal
3. Ecophysiology and Fruit Characterization
3.1. Plant Growth Requirements
3.2. Morphological Characterization
3.3. Fruit Nutritional Characterization
3.4. Commercialization and Uses
4. The Castanea Genetic Resources, Genetic Diversity and Biotechnology Based Approaches for Improvement
4.1. Intra- and Inter-Varietal Variability
4.2. Portuguese Breeding Programs and Candidate Gene Identification
4.3. The Potential of Genetic Transformation and Genome Editing as a Tool for Pathogen Control
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gomes-Laranjo, J.; Cardoso-Ferreira, J.; Portela, E.; Abreu, G.C. Castanheiros. 2007. Available online: https://www.cm-sabugal.pt/wp-content/uploads/Castanheiros-livro.pdf (accessed on 29 May 2023).
- Fernandes, P.; Colavolpe, M.B.; Serrazina, S.; Costa, R.L. European and American Chestnuts: An Overview of the Main Threats and Control Efforts. Front. Plant Sci. 2022, 13, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Lage, J. Castanea: Uma Dádiva Dos Deuses, 2nd ed.; Oficina São José: Braga, Portugal, 2006; pp. 1–325. [Google Scholar]
- Moura, A.R.; Gomes-Laranjo, J.C.; Cardoso, J.V. Seleção Clonal Na Variedade de Castanha “Longal” Visando a Sua Aptidão Agronómica e Agroalimentar. Master’s Thesis, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal, 2018. [Google Scholar]
- Mellano, M.G.; Beccaro, G.L.; Donno, D.; Marinoni, D.T.; Boccacci, P.; Canterino, S.; Cerutti, A.K.; Bounous, G. Castanea Spp. Biodiversity Conservation: Collection and Characterization of the Genetic Diversity of an Endangered Species. Genet. Resour. Crop. Evol. 2012, 59, 1727–1741. [Google Scholar] [CrossRef]
- Bounous, G. The Chestnut: A Multipurpose Resource for New Millenium. Acta Hortic. 2005, 693, 33–40. [Google Scholar] [CrossRef]
- Gomes-Laranjo, J.; Peixoto, F.; Costa, R.; Ferreira-Cardoso, J. Portugal, Following Chestnut Footprints, Cultivation and Culture, Folklore and History, Traditions and Uses. Scr. Hortic. 2009, 9, 106–111. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#home (accessed on 29 May 2023).
- Serdar, U.; Saito, T.; Cuenca, B.; Akyüz, B.; Laranjo, J.G.; Beccaro, G.; Bounous, G.; Costa, R.L.; Fernandes, P.; Mellano, M.G. Advances in cultivation of Chestnuts. In Achieving Sustainable Cultivation of Tree Nuts, 1st ed.; Serdar, U., Fulbright, D., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; Volume 1, pp. 349–388. [Google Scholar] [CrossRef]
- Instituto Nacional de Investigação Agrária e Veterinária (INIAV). Novos Porta-Enxertos de Castanheiro Com Resistência à Doença Da Tinta. 2018, pp. 1–36. Available online: https://projects.iniav.pt/NewCastRootstocks/images/Manual_INIAV_web.pdf (accessed on 29 May 2023).
- Freitas, G.M.; Igrejas, G.; Santos, H. Caracterização de Cultivares Portuguesas de Castanea Sativa Mill: Uma Análise Proteómica. Master’s Thesis, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal, 2017. [Google Scholar]
- Brito, D.; Gomes-Laranjo, J.; Álvaro, R.C. Produção de Novos Porta-Enxertos Híbridos Para Castanheiro. Caracterização Ecofisiológica Da Geração F1. Master’s Thesis, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal, 2014. [Google Scholar]
- Silva, A.P. Castanha. Um Fruto Saudável. In Projeto Agro 939, Dinamização do Consumo da Castanha Com Denominação de Origem, 1st ed.; Mingo Gráfica: Braga, Portugal, 2007; Volume 1, pp. 1–178. [Google Scholar]
- Costa, R.; Ribeiro, C.; Valdiviesso, T.; Afonso, S.; Borges, O.; Soeiro de Carvalho, J.; Costa, H.A.; Assunção, A.; Fonseca, L.; Augusta, C.; et al. Variedades Portuguesas de Castanha das Regiões Centro e Norte de Portugal, 1st ed.; Instituto Nacional dos Recursos Biológicos, I.P. (INRB, I.P.): Oeiras, Portugal, 2008; pp. 1–78. [Google Scholar]
- Europeia, C. Jornal Oficial Das Comunidades Europeias; European Union: Maastricht, The Netherlands, 1996. [Google Scholar]
- Direção-Geral de Agricultura e Desenvolvimento Rural. Produtos Tradicionais Portugueses. 2018. Available online: https://tradicional.dgadr.gov.pt/pt/ (accessed on 29 May 2023).
- Queijeiro, J.M.; Diaz-Raviña, M.; de la Montaña, J. Edaphic Characterization of Chestnut Tree Orchards in Monterrei (Southeast Galicia, Spain). Ecol. Mediterr. 2000, 26, 163–167. [Google Scholar] [CrossRef]
- Rojo, D.; Santín, J.; Sánchez, J.A. Guía de Buenas Prácticas Para La Conservación y Custodia Del Território Del Castãno; Imprenta Manolete: Asturias, Spain, 2016; pp. 1–37. [Google Scholar]
- Suarez, P.C. Fitopatologia Del Castaño (Castanea Sativa Miller). In Boletin de Sanidad Vegetal; Secretaría General Técnica Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1989; Volume 3, pp. 1–130. [Google Scholar]
- Gonçalves, J.C. Influência de Alguns Factores Na Micropropagação Do Castanheiro (Castanea Miller). Ph.D. Thesis, Higher Institute of Agronomy, Lisbon, Portugal, 1991. [Google Scholar]
- Dinis, L.; Peixoto, F.; Pinto, T.; Costa, R.; Bennett, R.N.; Gomes-Laranjo, J. Study of Morphological and Phenological Diversity in Chestnut Trees (“Judia” Variety) as a Function of Temperature Sum. Environ. Exp. Bot. 2011, 70, 110–120. [Google Scholar] [CrossRef]
- Bradford, M.; Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Valdiviesso, T.; Mederia, C.; Pinto de Abreu, C. Contribution for The Study of the Chestnut Floral Biology. In Proceedings of the International Congress on Chestnut, Spoleto, Italy, 20–23 October 1993; pp. 95–97. [Google Scholar]
- Giordano, E. Biology, Physiology and Ecology of Chestnut, Comunità Montana Monti Martani e Serano of Spoleto and Istituto di Coltivazioni Arboree; University of Perugia: Perugia, Italy, 1993. [Google Scholar]
- De Vasconcelos, M.C.; Bennett, R.N.; Rosa, E.A.; Ferreira-Cardoso, J.V. Composition of European Chestnut (Castanea Sativa Mill.) and Association with Health Effects: Fresh and Processed Products. J. Sci. Food Agric. 2010, 90, 1578–1589. [Google Scholar] [CrossRef]
- Sales, R.L.; Costa, N.M.B.; Monteiro, J.B.R.; Peluzio, M.D.C.G.; Coelho, S.B.; de Oliveira, C.G.; Mattes, R. Efeitos Dos Óleos de Amendoim, Açafrão e Oliva Na Composição Corporal, Metabolismo Energético, Perfil Lipídico e Ingestão Alimentar de Indivíduos Eutróficos Normolipidêmicos. Rev. Nutr. 2005, 18, 499–511. [Google Scholar] [CrossRef]
- Manela-Azulay, M.; Mandarim-De-Lacerda, C.A.; da Perez, M.A.; Filgueira, A.L.; Cuzzi, T. Vitamina C. An. Bras. Dermatol. 2003, 78, 265–272. [Google Scholar] [CrossRef]
- Borges, O.; Gonçalves, B.; de Carvalho, J.L.S.; Correia, P.; Silva, A.P. Nutritional Quality of Chestnut (Castanea Sativa Mill.) Cultivars from Portugal. Food Chem. 2008, 106, 976–984. [Google Scholar] [CrossRef]
- Dinis, L.R.; Gomes-Laranjo, J.; Peixoto, F.; Costa, R. Castanheiros: Seleção Clonal Na Variedade Judia. Ph.D Thesis, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal, 2011. [Google Scholar]
- Montanhas de Investigação. A Castanha Da Terra Fria DOP Inclui 10 Variedades Do Castanheiro (Castanea Sativa Mill)? Available online: https://morecolab.pt/2020/06/15/castanha-da-terra-fria-10-variedades-do-castanheiro/ (accessed on 29 May 2023).
- Gabinete de Planeamento, Políticas e Administração Geral. Available online: https://www.gpp.pt/ (accessed on 29 May 2023).
- Instituto Nacional de Estatística. Available online: https://www.ine.pt/xportal/xmain?xpgid=ine_main&xpid=INE (accessed on 29 May 2023).
- Esquinas-Alcázar, J.T. Plant Breeding. In Plant Breeding: Principles and Prospects; Hayward, M.D., Bosemark, N.O., Romagosa, I., Cerezo, M., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 33–51. [Google Scholar] [CrossRef]
- Carvalho, C. Detecção de Cromatina de Centeio e Análise Da Biodiversidade N landrace de Trigo “Barbela”, Através de Marcadores Genéticos. Ph.D Thesis, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal, 2003. [Google Scholar]
- Kremer, A.; Abbott, A.G.; Carlson, J.E.; Manos, P.S.; Plomion, C.; Sisco, P.; Staton, M.E.; Ueno, S.; Vendramin, G.G. Genomics of Fagaceae. Tree Genet. Genomes 2012, 8, 583–610. [Google Scholar] [CrossRef]
- American Chestnut Foundation. Available online: https://acf.org/ (accessed on 29 May 2023).
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Vidal, N.; Vieitez, A.M.; Fernández, M.R.; Cuenca, B.; Ballester, A. Establishment of Cryopreserved Gene Banks of European Chestnut and Cork Oak. Eur. J. For. Res. 2010, 129, 635–643. [Google Scholar] [CrossRef]
- Guo-Tian, L.; Cheng-Xiang, A.; Li-Si, Z.; Hai-Rong, W.; Qing-Zhong, L. Chestnut Genebank In China National Clonal Plant Germplasm Repository. Acta Hortic. 2009, 199–206. [Google Scholar] [CrossRef]
- Costa, R. Variedades e porta-enxertos. Man. Boas Práticas Frutic. 2020, 6, 4. Available online: https://www.iniav.pt/images/publicacoes/livros-manuais/manual_de_fruticultura_castanheiro.pdf (accessed on 29 May 2023).
- Costa, R.; Valdiviesso, T.; Marum, L.; Fonseca, L.; Borges, O.; Soeiro, J.; Soares, F.M.; Sequeira, J.; Assunção, A.; Correia, P. Characterisation of Traditional Portuguese Chestnut Cultivars by Nuclear SSRS. Acta Hortic. 2005, 437–440. [Google Scholar] [CrossRef]
- Dinis, L.-T.; Peixoto, F.; Costa, R.; Gomes-Laranjo, J. Molecular Characterization Of “Judia” (Castanea Sativa Mill.) From Several Trás-Os-Montes Regions By Nuclear Microsatellite Markers. Acta Hortic. 2010, 225–232. [Google Scholar] [CrossRef]
- Pereira, M.; Castro, L.; Torres-Pereira, J.; Lorenzo, S. Isozyme Polymorphisms in Portuguese Chestnut Cultivars. Acta Hortic. 1999, 283–286. [Google Scholar] [CrossRef]
- Santos, C.; Machado, H.; Serrazina, S.; Gomes, F.; Gomes-Laranjo, J.; Correia, I.; Zhebentyayeva, T.; Duarte, S.; Bragança, H.; Fevereiro, P. Comprehension of Resistance to Diseases in Chestnut. Rev. Ciências Agrárias 2016, 39, 189–193. [Google Scholar] [CrossRef]
- Natividade, J.V. Quatro Anos Na Defesa Da Campanha e Reconstituição Dos Soutos; Boletim Junta Nacional das Frutas: Lisbon, Portugal, 1947; pp. 1–21. [Google Scholar]
- Gomes Guerreiro, M. Alguns Estudos Do Género Castanea. Relatório. Tomos I e II. In Alcobaça: Publicação da Direcção Geral dos Serviços Florestais e Aquícolas; Topografia Alcobacence lda: Alcobaça, Portugal, 1948; Volume XV. [Google Scholar]
- Gomes Guerreiro, M. Castanheiros Alguns Estudos Sobre a Sua Ecologia e o Seu Melhoramento Genético. Master’s Thesis, Higher Institute of Agronomy, Lisbon, Portugal, 1957. [Google Scholar]
- Martins, L.; Anjos, R.; Costa, R.; Gomes-Laranjo, J. Colutad. Um Clone de Castanheiro Resistente à Doença Da Tinta. In Castanheiros. Técnicas e Práticas; Gomes-Laranjo, J., Peixoto, F., Ferreira-Cardoso, J., Eds.; Indústria Criativa and University of Trás-os-Montes and Alto Douro: Vila Real, Portugal, 2009; pp. 135–142. [Google Scholar]
- Costa, R.; Santos, C.; Tavares, F.; Machado, H.; Gomes-Laranjo, J.; Kubisiak, T.; Nelson, C.D. Mapping and Transcriptomic Approches Implemented for Understanding Disease Resistance to Phytophthora Cinammomi in Castanea Sp. BMC Proc. 2011, 5, O18. [Google Scholar] [CrossRef]
- Santos, C.; Machado, H.; Correia, I.; Gomes, F.; Gomes-Laranjo, J.; Costa, R. Phenotyping Castanea Hybrids for Phytophthora Cinnamomi Resistance. Plant Pathol. 2015, 64, 901–910. [Google Scholar] [CrossRef]
- Fernandes, P.; Amaral, A.; Colavolpe, B.; Pereira, A.; Costa, R. Avaliação Agronómica de Genótipos de Castanheiro Selecionados Do Programa de Melhoramento Genético Para a Resistência à Tinta. Vida Rural 2021, 1, 76–82. Available online: https://www.researchgate.net/publication/352878064 (accessed on 29 May 2023).
- Fernandes, P.; Amaral, A.; Colavolpe, B.; Balonas, D.; Serra, M.; Pereira, A.; Costa, R.L. Propagation of New Chestnut Rootstocks with Improved Resistance to Phytophthora cinnamomi—New Cast Rootstocks. Silva Lusit. 2020, 28, 15–29. [Google Scholar] [CrossRef]
- Kubisiak, T.L.; Nelson, C.D.; Staton, M.E.; Zhebentyayeva, T.; Smith, C.; Olukolu, B.A.; Fang, G.-C.; Hebard, F.V.; Anagnostakis, S.; Wheeler, N.; et al. A Transcriptome-Based Genetic Map of Chinese Chestnut (Castanea Mollissima) and Identification of Regions of Segmental Homology with Peach (Prunus Persica). Tree Genet. Genomes 2013, 9, 557–571. [Google Scholar] [CrossRef]
- Serrazina, S.; Santos, C.; Machado, H.; Pesquita, C.; Vicentini, R.; Pais, M.S.; Sebastiana, M.; Costa, R. Castanea Root Transcriptome in Response to Phytophthora Cinnamomi Challenge. Tree Genet. Genomes 2015, 11, 6. [Google Scholar] [CrossRef]
- Zhang, J.; Bernatzky, R.; Anagnostakis, S.L.; Doudrick, R.L.; Kubisiak, F.V.H.T.L.; Ji, F.; Wei, W.; Liu, Y.; Wang, G.; Zhang, Q.; et al. Molecular Mapping of Resistance to Blight in an Interspecific Cross in the Genus Castanea. Phytopathology 1997, 87, 751–759. [Google Scholar] [CrossRef]
- Zhebentyayeva, T.; Chandra, A.; Abbott, A.; Staton, M.; Olukolu, B.; Hebard, F.; Georgi, L.; Jeffers, S.; Sisco, P.; James, J.; et al. Genetic and Genomic Resources For Mapping Resistance To Phytophthora Cinnamomi In Chestnut. Acta Hortic. 2014, 263–270. [Google Scholar] [CrossRef]
- Zhebentyayeva, T.N.; Sisco, P.H.; Georgi, L.L.; Jeffers, S.N.; Perkins, M.T.; James, J.B.; Hebard, F.V.; Saski, C.; Nelson, C.D.; Abbott, A.G. Dissecting Resistance to Phytophthora Cinnamomi in Interspecific Hybrid Chestnut Crosses Using Sequence-Based Genotyping and Qtl Mapping. Phytopathology 2019, 109, 1594–1604. [Google Scholar] [CrossRef]
- Santos, C.; Nelson, C.D.; Zhebentyayeva, T.; Machado, H.; Gomes-Laranjo, J.; Costa, R.L. First Interspecific Genetic Linkage Map for Castanea Sativa x Castanea Crenata Revealed QTLs for Resistance to Phytophthora Cinnamomi. PLoS ONE 2017, 12, e0184381. [Google Scholar] [CrossRef] [PubMed]
- Carraway, D.; Wilde, H.; Merkle, S. Somatic Embryogenesis and Gene Transfer in American Chestnut. J. Am. Chestnut 1994, 6, 29–33. [Google Scholar]
- Seabra, R.C.; Pais, M.S. Genetic transformation of European chestnut. Plant Cell Rep. 1998, 17, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Corredoira, E.; Valladares, S.; Allona, I.; Aragoncillo, C.; Vieitez, A.M. Ballester, Genetic Transformation of European Chestnut Somatic Embryos with a Native Thaumatin-like Protein (CsTL1) Gene Isolated from Castanea Sativa Seeds. Tree Physiol. 2012, 32, 1389–1402. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Corredoira, E.; José, M.C.S.; Vieitez, A.M.; Allona, I.; Aragoncillo, C.; Ballester, A. Agrobacterium-Mediated Transformation of European Chestnut Somatic Embryos with a Castanea Sativa (Mill.) Endochitinase Gene. New For. 2016, 47, 669–684. [Google Scholar] [CrossRef]
- Carlson, E.; Stewart, K.; Baier, K.; McGuigan, L.; Ii, T.C.; Powell, W. Pathogen-induced Expression of a Blight Tolerance Transgene in American Chestnut. Mol. Plant Pathol. 2022, 23, 370–382. [Google Scholar] [CrossRef]
- Polin, L.D.; Liang, H.; Rothrock, R.E.; Nishii, M.; Diehl, D.L.; Newhouse, A.E.; Nairn, C.J.; Powell, W.A.; Maynard, C.A. Agrobacterium-Mediated Transformation of American Chestnut (Castanea Dentata (Marsh.) Borkh.) Somatic Embryos. Plant Cell Tissue Organ Cult. (PCTOC) 2006, 84, 69–79. [Google Scholar] [CrossRef]
- Rothrock, R.E.; Polin-McGuigan, L.D.; Newhouse, A.E.; Powell, W.A.; Maynard, C.A. Plate Flooding as an Alternative Agrobacterium-Mediated Transformation Method for American Chestnut Somatic Embryos. Plant Cell Tissue Organ Cult. (PCTOC) 2007, 88, 93–99. [Google Scholar] [CrossRef]
- McGuigan, L.; Fernandes, P.; Oakes, A.; Stewart, K.; Powell, W. Transformation of American Chestnut (Castanea Dentata (Marsh.) Borkh) Using RITA® Temporary Immersion Bioreactors and We Vitro Containers. Forests 2020, 11, 1196. [Google Scholar] [CrossRef]
- Pavese, V.; Moglia, A.; Corredoira, E.; Martínez, M.T.; Marinoni, D.T.; Botta, R. First Report of CRISPR/Cas9 Gene Editing in Castanea Sativa Mill. Front. Plant Sci. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Pavese, V.; Moglia, A.; Gonthier, P.; Marinoni, D.T.; Cavalet-Giorsa, E.; Botta, R. Identification of Susceptibility Genes in Castanea sativa and Their Transcription Dynamics following Pathogen Infection. Plants 2021, 10, 913. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Name | Port | Cup | Leaves | Catkins | Blooming | Fruit | Maturation |
|---|---|---|---|---|---|---|---|
| “Aveleira” | Open | Spherical-oval | Petiolate, mucronate, obtuse base, oblong-lanceolate, acute on some leaves, slightly asymmetric and slightly concave, light green on the abaxial surface and glabrous on the adaxial surface | Unisex male astaminates inserted very close to each other, in the axilla of the leaves, and androgynics at the base of the catkin | Early, 2nd week of June | Medium fruit, ovoid to broad ovoid, shiny strong brown, red tint with well-spaced dark streaks. Apex with sericeous star-shaped pubescence. Ventral surface predominantly planoconcave and convex dorsal surface, almost semispherical | Early, fall between the 2nd and 3rd decade of September |
| “Martaínha” | Semi-erect to open, branch insertion angles of branches 45° to 60° | Round-oval | Petiolate, base predominantly obtuse, oblong-lanceolate, toothed, straight to slightly concave and slightly asymmetric, glabrous only on the adaxial surface, light green on the abaxial surface, shiny medium green on the adaxial surface | Unisex male brachyaminates, inserted in the leaf axil of the shoots and androgynics | Early, 2nd week of June | Medium to large fruit, ovoid, brown, satiny luster, with dark bordeaux dissipated longitudinal stripes. Triangular pubescence sericeous at the apex on the dorsal side. Flat concave on the ventral surface and predominantly convex on the dorsal surface | Early, fall in the 3rd decade of September |
| “Longal” | Erect to semi-erect, branch insertion angles of branches ≤ 45° | Pyramidal | Petiolate, obtuse base oblong-lanceolate, dentate, slightly asymmetric and straight to slightly concave, glabrous only on the adaxial surface, light green on the abaxial surface and shiny light green on the adaxial surface | Unisex male mesostaminates, inserted in the leaf axil and androgynics | Average, 4th week of June | Medium to large fruit, ovoid, shiny light brown, reddish tint, with well-defined longitudinal stripes, forming slightly prominent, slightly protruding edges on the integument. Small, silky white pubescence at the apex, involving the stipes up to the stigmas. Planoconvex on the ventral surface and convex on the dorsal surface | Late, fall in the 2nd decade of October. |
| “Judia” | Erect to semi-erect, big insertion angles, small branches < 45° to more open angles (about 70°) | Spherical-pyramidal | Petiolate, obtuse base, oblong-lanceolate, toothed, slightly asymmetric and slightly concave, glabrous on the adaxial surface, shiny and more faded green on the abaxial surface and medium green on the adaxial surface | Unisex male astaminated with fillets and anthers not visible on the outside of the flower and androgynics | Average, 3rd week of June | Medium fruit, large ovoid, intense brightness brick-brown, lighter and dark brown tint. Apex with reduced sericeous pubescence. Ventral surface concave-plane to slightly convex in some fruits and convex on the dorsal surface | Average, fall in the 2nd decade of October |
| “Côta” | Semi-erect | Pyramidal | Mucronate, lamina obtuse base, very concave, slightly asymmetrical cross-section leaf, whitish on the abaxial surface and medium green on the adaxial surface | Unisex male brachyaminates | Male flowering (1st week of July) and female flowering (2nd week of July) | Small fruit, large ovoid, dark brown | Average, fall between the 1st and 2nd decades of October |
| “Negral” | Erect | Rounded conic | Petiolate, cordiform limb base, oblong-lanceolate, asymmetrical leave, toothed, slightly asymmetrical cross-section, whitish abaxial surface and dark green adaxial surface | Unisex male brachyaminates | Male flowering (1st week of July) and female flowering (3rd week of July) | Large fruit, globular, dark brown | Average, fall in the 2nd decade of October |
| “Amarelal” | Semi-erect to open, branch insertion angles of branches 45° to 60° | Pyramidal | Petiolate, obtuse base, oblong-lanceolate, mucronate, slightly asymmetric, slightly concave in cross section. Glabrous light green on the abaxial surface and bright green on the adaxial surface | Unisex male mesostaminates, inserted in the axillae of the leaves and androgynics | Average, occurring in the 2nd/3rd weeks of June | Large to medium fruit, globular, light brown with clearly visible dark and shiny longitudinal well, occasionally with a slight concavity. Silky white pubescence on the apex. Almost flat ventral surface and convex on the dorsal surface | Early, occurring in the last week of September/1st week of October |
| “Lamela” | Open, the insertion angles of most of the lower branches > 45° | Rounded | Petiolate, limb with acute base, oblong-lanceolate, toothed, symmetrical, lighter green color on the abaxial surface and glabrous medium green on the adaxial surface | Unisex male brachyaminates to mesostaminates inserted in the leaf axils and androgynics | Average | Large, globular, light brown color, dull, near the hilum shows a dark brown color. Apex with reduced sericeous pubescence. Large hilum. Flat concave on ventral side | Average, fall in the 2nd decade of October |
| Plant Material | Approach for Improvement | Resources | Description | References |
|---|---|---|---|---|
| C. molissima × C. dentata F2 hybrids | Disease resistance (chestnut blight) | Study genetic architecture of CP resistance; future MAS | 3 QTLs: Cbr1 (LG B), Cbr2 (LG F), Cbr3 (LG G) | [56] |
| BC1, BC4 C. dentata × C. dentata molíssima “ Nanking” and “Mahogany” | Disease resistance (chestnut blight and ink disease) | Study genetic architecture of CP and PC resistance; future MAS | 1 QTL: LG E | [57] |
| BC1F1 BC3F1 C. dentata × C. dentata molíssima “ Nanking” and “Mahogany” | Disease resistance (chestnut blight and ink disease) | Study genetic architecture of CP and PC resistance; future MAS | 22 QTLs: hb52208, hb39959 (LG A), nk12394 (LG C), h25723, h54539, hb54410, jb79599, jb32342, jb13258, nk29352, nk35044, nk19473 (LG E), h31744, hb7824, hb27106 (LG K) | [58] |
| C. sativa × C. crenata F1 hybrids | Disease resistance (ink disease) | Study genetic architecture of PC resistance; future MAS | 17 QTLs: CC_3129_774, CmSNP00773E, CC_48142_849_CmSNP00522E, AC_32934_470, AC_36335_960 (LG E), CC_46475_1222, CC_6279_2669, AC_14650_453 (LG K) | [59] |
| Explant | Gene of Interest | Gene Function | Target Pathogen | References |
|---|---|---|---|---|
| C. sativa somatic embryos | CsTL1 | Thaumatin-like protein; promotes osmotic rupture in the pathogen | Phytophthora cinnamomi | [62] |
| C. sativa somatic embryos | CsCH3 | Chitinase-like protein; hydrolyses chitin from pathogen’s cell wall | Cryphonectria parasitica | [63] |
| C. dentata somatic embryos | OxO | Detoxifying enzyme; degrades oxalic acid | Cryphonectria parasitica | [64,65,66,67] |
| C. dentata somatic embryos | Cast_Gnk2-like | Antifungal | Phytophthora cinnamomi | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braga, M.; Araújo, S.d.S.; Sales, H.; Pontes, R.; Nunes, J. Portuguese Castanea sativa Genetic Resources: Characterization, Productive Challenges and Breeding Efforts. Agriculture 2023, 13, 1629. https://doi.org/10.3390/agriculture13081629
Braga M, Araújo SdS, Sales H, Pontes R, Nunes J. Portuguese Castanea sativa Genetic Resources: Characterization, Productive Challenges and Breeding Efforts. Agriculture. 2023; 13(8):1629. https://doi.org/10.3390/agriculture13081629
Chicago/Turabian StyleBraga, Megan, Susana de Sousa Araújo, Hélia Sales, Rita Pontes, and João Nunes. 2023. "Portuguese Castanea sativa Genetic Resources: Characterization, Productive Challenges and Breeding Efforts" Agriculture 13, no. 8: 1629. https://doi.org/10.3390/agriculture13081629
APA StyleBraga, M., Araújo, S. d. S., Sales, H., Pontes, R., & Nunes, J. (2023). Portuguese Castanea sativa Genetic Resources: Characterization, Productive Challenges and Breeding Efforts. Agriculture, 13(8), 1629. https://doi.org/10.3390/agriculture13081629