Abstract
Corn is an important crop for animal production systems. Owing to climate change, it is important to understand the behavior of commercial corn hybrids under low soil water availability. The objective of this study was to evaluate the morphogenic characteristics and yields of silage corn hybrids subjected to different water conditions using univariate and multivariate analyses. The experimental design was randomized blocks in a 5 × 2 factorial scheme with four replications. Corn hybrids (AGRI104, AGRI320, AGRI330, AGRI340, and AGRI360) and water replacement levels (50% and 100% replacement of crop evapotranspiration—ETc) were considered as factors. A water replacement level of 50% ETc was defined as a severe water deficit that persisted for 46 days from V7 until harvesting (R3). Plant height, stem diameter, leaf area, chlorophyll index, water productivity, fresh and dry mass, and dry matter of the shoot and root systems were evaluated. An analysis of variance and a mean test of isolated variables were carried out, and a multivariate perspective, through principal components was performed. In general, the replacement of 50% ETc changed the morphophysiological characteristics of the plants; however, it was only possible to classify the hybrids through a principal components analysis. The best performing hybrids, regardless of the water replacement levels, were AGRI340 and AGRI360.
1. Introduction
Water deficits are a frequent cause of reduced food production. It is estimated that 40% of the global population will suffer from water scarcity in the next few years [1], and the predicted global climate change will imply a significant increase in the water footprint of corn [2].
Recently, Conab [3] estimated that the average yield of corn grains has been reduced owing to the high fluctuations in the rainfall regime that occurred throughout the Brazilian territory, especially in the second crop. In addition to these climate changes, agricultural crops in general cultivation during the second season coincide with periods of severe water deficit and lower soil water storage for a long period, especially in regions located in the Cerrado biome [4].
Corn is an important crop for livestock but more sensitive to water deficit than other grasses [5]; however, some hybrids use mechanisms to reduce the productivity losses which occur as a consequence of the biochemical and physiological disorganization induced by this stress [6,7]. Water deficit can be thus tackled through the identification and use of corn hybrids that make better use of water during development [8], which decreases the water footprint and contributes to the sustainability of agricultural systems [5].
Various methodologies and data analyses have been carried out to select corn genotypes resistant to water stress, indicating promising lines for genetic improvement programs [9]. However, this goal is challenging due to variations in the results depending on when morphological assessments were performed and given the existence of plants with low productivity and others that, by employing mechanisms to avoid dehydration, present the detriment of the yield grains and biomass [10,11].
Commercial hybrids have the advantage of having already been selected for their productivity by companies holding their patents. In the present work, we use AGRI cultivars, which according to MAPA [12], have a 120-day cultivation cycle. The phases—I, germination/emergence; II, growth/development; III, flowering/grain filling; and IV, physiological maturation/harvest—last 15, 45, 40, and 20 days, respectively. Before using these materials, information about their performance in places with water restriction is necessary. Thus, it is possible to carry out efficient agroclimatic zoning.
Based on the hypothesis that water deficit has strong negative influences on productivity and on important morphological, physiological, and biochemical characteristics of maize plants [6,7,8,9,10,11,13], different plant cultivars can have different responses to water deficit, which can lead to variations in productivity and quality of harvested products. Furthermore, different ways of analyzing the results should be tested in order to better detect differences in and groupings of materials that are more tolerant to water deficits. Therefore, the objective of the present study was to evaluate the performance of silage Brazilian corn hybrids under water scarcity conditions using univariate and multivariate analyses.
2. Materials and Methods
The present study was carried out to evaluate different corn hybrids for silage in two water conditions (with and without water deficit). During corn cultivation in pots and in a protected environment, growth parameters were obtained throughout the cycle. In corn’s R5 stage, parameters related to shoot and root system production were evaluated. Data were analyzed using traditional statistics, and principal component analysis was also used to support the conclusions obtained in the study.
The experiment was carried out in a greenhouse located in the municipality of Campo Grande, state of Mato Grosso do Sul, Brazil. The climate of the region is Aw, characterized as tropical, with a well-defined dry season in the cold months and heavy rainfall during the summer, according to the Köppen classification [14].
The temperature and relative humidity of the air were recorded daily (Figure 1) by using a thermo-hygrometer placed inside the greenhouse. The minimum and maximum air temperatures were 19.0 and 37.6 °C, respectively, and the relative humidity ranged from 23.5 to 86.6%.
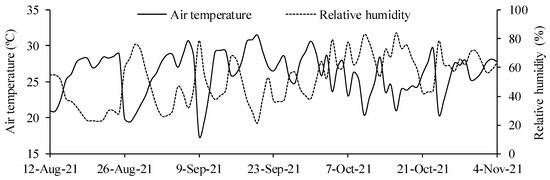
Figure 1.
Daily behavior of medium temperature (T °C) and relative humidity (RH %) during the experimental period.
The experimental design was a randomized block in a 5 × 2 factorial arrangement, with four replications. Five corn hybrids grown for silage (AGRI104, AGRI320, AGRI330, AGRI340, and AGRI360), and two water replacement levels (50% and 100% crop evapotranspiration—ETc) were selected as factors. All hybrids studied are recommended for silage and have an early cycle, and the grain type is orange semi-dented [15]. The chosen hybrids were recently launched in the Brazilian market, although their behavior under water scarcity conditions is unknown.
The study was carried out in a pot so that it was possible to provide the amount of water precisely, guaranteeing the correct levels of water in the imposition of treatments. The pots had a base width, top width, and height of 19, 24, and 29 cm, respectively. The experimentation in pot allowed less physical space requirements, which made it possible to carry out the same experiment in a greenhouse. In this way, there was precise control of the cultivation environment, mainly in relation to the exclusion of the contribution of water from rainfall.
The soil was identified as dystrophic Red Latosol (Oxisol), collected from an area previously cultivated with forage, and the chemical and physical attributes were organic matter (g dm−3): 44.9; pH (CaCl2): 5.6; P (Mehlich): 86; K (mmolc dm−3): 18.6; Ca (mmolc dm−3): 67; Mg (mmolc dm−3): 26; H + Al (mmolc dm−3): 36; Clay (g kg−1): 637; Silt (g kg−1): 157; Sand (g kg−1): 206; field capacity (g g−1): 0.3001; wilting point (g g−1): 0.2278.
Samples of soil were crushed in 5 mm sieves, homogenized, corrected, and fertilized according to physical and chemical analyses. After corrective fertilization (30 mg dm−3 of K2O, 15.05 mg dm−3 of N, and 52.5 mg dm−3 of P2O5), the volume of 8-dm3 of soil was deposited in the pots, which were carefully labeled and placed in a greenhouse.
Sowing was performed manually, with seven seeds per pot inserted at a depth of approximately 1 cm. Uniform thinning in all pots was performed on the tenth day after sowing to maintain one plant per pot. Thus, each experimental unit consisted of a plant grown in a single pot. In total, the experiment had 40 plants and 40 pots.
All plants were maintained at field capacity with daily irrigation to replace 100% ETc until the end of the V6 vegetative stage (six expanded leaves). When they reached the seventh expanded leaf (V7), at 37 days after emergence (DAE), the two treatments were imposed with replacement levels of 100% ETc and 50% ETc. The replacement of 50% ETc was defined as a severe and persistent water deficit, as it took place for 46 days, that is, until the plants had pasty grains (R3).
The replacement of evapotranspirated water was performed using the mass required to raise the soil to field capacity [16,17]. For this, at the time of installation of the experiment, all pots received the same volume of soil. Then, the soils contained in the pots were saturated, and the upper area was covered to avoid loss of water by evaporation. After the soil had drained and the pot had constant mass, it was assumed that the soil was in field capacity condition, that is, the soil had the maximum amount of water retained. From this value, the pots were weighed daily and the difference between the current mass and the mass equivalent to the field capacity was the water that evapotranspired in the last 24 h. In the 100% ETc treatment, all the lost water was replaced, and in the 50% ETc treatment, half of the calculated volume was used in irrigation.
The plants received two foliar sprays with micronutrients in the vegetative stage (V4 and V5) using 1 g L−1 of a foliar fertilizer (Mg: 1.2%, B: 0.85%, Fe: 3,4%, Zn: 4.2%, Mn: 3.2%, Cu: 0.5%, Mo: 0.06%). When they were in V5, the plants were also fertilized with 90 mg dm−3 of N in topdressing in the form of urea (45% N). Defoliating caterpillars were controlled by spraying insecticides registered for the crop.
Corn growth evaluations were carried out at 47, 60 and 73 DAE. On those dates, the corn was in phases V14 (vegetative phase with 14 leaves), Vt (feathering), and R2 (milky grains), respectively. Plant height, stem diameter, and leaf area index (LAi): the plant height was measured as the distance from the last exposed ligule to the ground, stem diameter was measured using a digital caliper, and the LAi was estimated from the average of the last three expanded leaves.
The quantification of the chlorophyll index (CLi) was performed indirectly using a portable chlorophyll meter at leaf CHL BLUE [18]. According to the authors, the chlorophyll meter emits blue light on the leaf and measures the amount of reflected light. Chlorophyll absorbs blue light and reflects light close to red. The device compares the intensity of the reflected light with a reference value and provides a reading indicating the amount of chlorophyll present in the leaves. An average of 20 readings were performed on the last newly expanded sheet to obtain the CLi on dates 47, 60, 67, 70, and 73 DAE.
The experiment was conducted during the vegetative phase and up to the R5 phase, when the corn begins to show dent and the milk line, a period in which water deficit significantly affects corn productivity [19]. Thus, at 83 DAE, the plants were cut approximately 5 cm above the ground level and chopped to estimate the fresh mass, and dry mass of the aerial part (g pot−1), as well as the dry matter content [20].
After removing the aerial part, the soil together with the roots was removed from the pots. To separate both, the root systems of the plants were deposited on a 1–2 mm sieve and washed in running water so that the fresh mass, dry matter, and dry matter content could be further estimated [21].
The sum of the volumes of the water used daily during the experiment was recorded to obtain the average consumption of each treatment. Water productivity was obtained from the ratio of dry matter produced by the plant to total water consumption.
All data were submitted to analysis of variance; when necessary, the Scott–Knott test of means was performed, with a significant effect at 5%. In the analysis of the principal components (PC), the results obtained at 73 DAE were used, and the dataset was standardized so that each descriptor had a null mean and variance equal to unity. Statistical analyses were performed using the R software program (version 4.0).
3. Results and Discussion
The total water consumption was found to be 57.0 and 34.7 L for the cultivated plants, with replacements of 100 and 50% ETc, respectively (Figure 2). Up to 36 DAE, all hybrids received water replacement equivalent to 100% of ETc. From that date, the plants in the 50% ETc treatment began to receive less water, reducing the matric potential of the soil and triggering the effects that will be presented later [22,23]. In the treatment of 100% of ETc, it is verified that the slope of the curve is increased from 36 DAE. This result is a function of the increase in the leaf area index and consequent increase in the transpiration component [24].
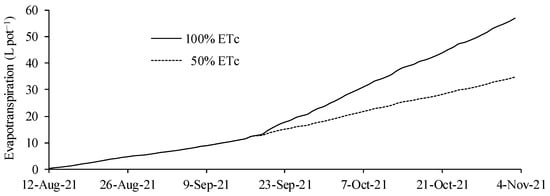
Figure 2.
Water consumption of corn plants grown in pots and irrigated with water replacement levels of 50 and 100% of crop evapotranspiration—ETc.
Some growth parameters were analyzed at different times to monitor corn throughout the cycle. When evaluating plant height (Table 1), no interaction was observed between the hybrids and water replacement levels. A reduction in plant height was identified when irrigation was applied to meet 50% ETc. With the reduction in water supply in this treatment, the water potential of the soil was reduced. This contributes to the reduction in water flow from the soil to the plant [13]. In moments of greater water vapor pressure deficit, so that transpiration does not exceed the flow of water from the soil to the plant and lead to high leaf temperatures, the plant regulates its stomatal opening, reducing transpiration [25]. In this regulation, gas exchange is also reduced, triggering lower rates of photosynthesis and production of photoassimilates [26]. These photoassimilates are responsible for the growth and development of the aerial part of the corn, partly explaining the smaller plant heights observed in the hybrids submitted to the water replacement treatment of 50% of the ETc.

Table 1.
Height and stem diameter of corn hybrids (HBD, cm) as a function of water replacement levels (WRL, %ETc) evaluated on different days after emergence (DAE).
The replacement of 100% ETc promoted the development of taller plants by meeting the water demand for tissue production. The plants reached an average height of 105.49 and 142.39 cm when irrigated with 50 and 100% ETc, respectively. Despite this, a lower stature of the hybrids evaluated in this study was observed when they were cultivated under field conditions [20]. These differences may be attributable to the physical limitations to the development of the root system and soil exploration caused by the smaller volume of the pot.
The reduction of approximately 25.9% in plant height at 73 days between treatments with different water conditions corroborates the results observed by Gomaa et al. [27]. They studied the effect of water scarcity on corn cultivation, imposed as a function of three irrigation intervals, and found a significant effect of water volume on plant height. In previous studies, reduced plant height was also found to be associated with lower water supply, which was explained by the physiological effects of stomatal guard cell closure [28,29].
It was observed that the plants had a smaller stem diameter, approximately 15%, only after 10 days of water deficit imposition (Table 1). No significant interactions were found between the hybrids and water replacement levels, with fewer differences between the hybrids at 60 and 73 DAE. These results indicated that the date of evaluation of the stem diameter can influence the results and interpretation of their response to water deficit.
There was no significant interaction found between the factors for the LAi (Table 2). Within the hybrid factor, differences were found at 47 DAE, and the highest index was observed in the hybrids AGRI320 and AGRI360.
Table 2.
Leaf area index of corn hybrids (HBD, cm3) as a function of water replacement levels (WRL, %ETc) evaluated on different days after emergence (DAE).
Under the effect of water replacement levels and in all evaluations performed, the highest LAi was observed in the treatment with 100% ETc. In agreement with these results, Avramova et al. [8] and Alves et al. [30] observed reduced leaf development in corn plants under water scarcity, which was associated with a reduced cell division rate.
Table 3 shows the chlorophyll index (CLi) values of corn at different dates and treatments. The values measured using the chlorophyll meter varied between 42.3 and 55.8. These values are similar to those obtained by Soaud [31], who, applying different doses of nitrogen, obtained CLi values ranging between 44.1 and 56.0. There was an interaction between the factors at 67 and 70 DAE for the CLi. There was also a significant difference (p < 0.05) in the water replacement levels at 47 DAE.
Table 3.
Chlorophyll index in corn hybrid leaves (HBD) as a function of water replacement levels (WRL, %ETc) evaluated on different days after emergence (DAE).
At 67 DAE, the interaction under the replacement of 100% ETc provided the hybrid AGRI360 with the highest CLi. At 70 DAE, the hybrid AGRI360 exhibited the highest chlorophyll transmittance values within the group of hybrids and with irrigation of 100% ETc. At 47 DAE, the replacement of 100% ETc provided an increase in the leaf index by approximately 10.5%, in relation to the treatment with irrigation of 50% ETc. At 73 DAE, the same hybrids showed increases of 8.2 (AGRI320) and 11.7% (AGRI360) in relation to the lowest average, found in the AGRI104 hybrid.
Although these results depended on the evaluation time after plant emergence, maybe the results could be more accurately obtained with modern equipment for investigating the photosynthetic efficiency of corn under water deficit. Recently, Chiango et al. [32] used a portable fluorometer to analyze the performance of photosystem II in the vegetative stage, which allowed the grouping of corn hybrids according to their tolerance to water suppression.
It was observed that the 50% reduction in the water supply to the plants reduced the production of fresh mass and dry mass in the aerial parts of the plants per pot (Table 4). No significant effect of the water regime on shoot dry mass content was identified, nor was there a difference (p > 0.05) between hybrids.
Table 4.
Fresh mass, dry mass, and shoot dry matter content and water productivity of corn hybrids (HBD) as a function of water replacement levels (WRL, %ETc).
The highest yields of fresh mass (61.9%) and dry mass (70.8%) occurred in the treatment with 100% ETc. This shows that the corn plants suffered the effects of the lower water availability in the soil imposed by the 50% ETc. It is worth noting that the effects in experiments in pots are intensified by the lower volume of soil explored [33,34]. Thus, small changes in soil moisture make plants experience lower water potential compared with the conventional cultivation situation.
Regarding water productivity, there was no significant interaction between the evaluated factors or isolated effects of hybrids and water replacement levels (Table 4). Corn water productivity increased only 5.4% when the soil was irrigated with 100% ETc, not being enough to cause statistical difference. This reveals that the ratio between the dry mass of corn produced and the amount of water applied was not affected by the reduction in water supply. The average value of water productivity, considering all treatments, was 1.91 g L−1. This result indicates that a volume of irrigation water of 524 L was needed to produce 1 kg of dry mass of corn silage.
The water productivity results for corn in this study must be analyzed with great care to avoid erroneous generalizations, since the variable dry mass of the aerial part was used to obtain these data and not corn grain yield. Another factor worth mentioning is that the water productivity was obtained through an experiment carried out in pots; therefore, its comparison with other work and extrapolation to situations of cultivation in crops must be carried out with prudence.
The productivity values for fresh mass, dry mass, and dry matter content of the corn root system are shown in Table 5. There was a significant interaction (p < 0.05) between the hybrids and water replacement levels for fresh mass, and root dry mass. An increase of 79.4% was observed in the dry mass of the hybrid AGRI320 compared with that in the AGRI104.
Table 5.
Fresh mass, dry mass, and dry mass content of the root system of corn hybrids (HBD) as a function of water replacement levels (WRL, %ETc).
The highest values for fresh mass of the root system were observed for the hybrids AGRI320 and AGRI330 when treated without water deficit (100% ETc). Under the effect of water replacement of 50% of ETc, despite the significant interaction, the high coefficient of variation interfered with the separation of means, and it was not possible to identify a significant difference between the hybrids. It should also be considered that the root system was limited in growth and development because of the smaller volume of soil imposed by the pots. The AGRI320 hybrid was the only hybrid with the highest productivity of root system dry mass in the irrigation treatment with 100% ETc. Regarding the dry matter content, the average treatment with water deficit was 44.5% higher than that the treatment 100% ETc.
According to the results shown in Table 5, and upon comparing the hybrids with each other, it was not possible to indicate the presence of any genes that would indicate a greater tolerance to severe water deficit, since the plants developed the same amount of dry mass as the root system when subjected to a water deficit of V7 until the moment of cutting (R3). It is hypothesized that severe and persistent water deficit was the reason for the results observed because, based on the methodology used in this work, the plants remained under low water supply during several critical periods in the development of the root system [35].
The effects of water deficit on corn morphology found in this study were in partial agreement with other studies of the same nature. Some authors have noted the effect of water deficit on the loss of plant phenotypic expression, especially on the reduction in LAi and plant height [36,37,38]. Sah et al. [19] argued that root system development is not always suppressed by water deficits, as it varies according to the genetic background of the hybrid. In the present study, variation was observed in the fresh and dry mass of the root system of the hybrids evaluated against water deficits (Table 5), with emphasis on AGRI320, which was the only one with a significant reduction in root system dry mass when cultivated with replacement of 50% ETc.
In this experiment, we attempted to select corn hybrids for silage in greenhouse; however, the variables analyzed alone were not sufficient to provide an assertive conclusion. As seen in Table 1, Table 2, Table 3, Table 4 and Table 5, there were rare situations in which some variables allowed the indication of a promising hybrid for cultivation under water restriction, and there were different responses depending on the moment of the evaluation. The implication of this result lies in the time required to conduct experiments for the selection of water-deficit-tolerant corn genotypes [38,39]. If the experiments are finished early, even in the vegetative stages, they may underestimate the effects of water deficit on plant performance.
In the present study, CLi and LAi varied according to the moment of evaluation of the hybrids used, potentially generating discordant conclusions if the evaluations were made at different phenological stages (Table 2 and Table 3). It is suggested that indirect analyses of chlorophyll content or photosynthetic efficiency in corn should be performed at different times after emergence and according to the period of exposure of plants to water stress.
An alternative that would allow the isolated analysis of variables would be the use of genotypes with previously known behavior in the face of water deficit as instruments for comparison [9,32]. However, seeds of these genotypes are not always available, and if water productivity is to be used as a selection criterion, the grain or biomass yield for silage may be lower than achieved by modern hybrids, even under full supply of water for the plants [19].
However, from a multidimensional perspective, with the use of the PC method, it was possible to explain about 75 and 84% of the total variance in the data obtained in two PCs, when the plants were subjected to the replacement of 100 and 50% ETc, respectively. The significance of each variable can be observed via the correlation between the correlation matrix of the data and PCs.
When the hybrids were submitted to irrigation for 100% ETc replacement, the most relevant variables for the first principal component (PC1), which explained 46% of the data variance, were LAi, shoot dry mass, and water productivity. In the second principal component (PC2), which explained 30% of the variance, the most representative variables were stem diameter, plant height, and root dry mass.
Under these conditions, without a water deficit, it was possible to separate the corn hybrids from a multivariate perspective and as a function of the two PCs observed in Figure 3A. Regarding PC1, it is possible to observe a high positive correlation of the AGRI340 hybrid with the shoot dry mass and water productivity variables, and a negative correlation with the LAi. The hybrid AGRI360 was also highlighted in one of the quadrants of the biplot because it was positively related to shoot dry mass, water productivity, and CLi, with a lower correlation with LAi. According to PC2, the AGRI320 hybrid had a high correlation with root dry mass and plant height; however, it was in the opposite position to stem diameter. The hybrids AGRI104 and AGRI330 were in the opposite quadrant to the shoot dry mass and water productivity variables, indicating a lower performance than the other hybrids that were irrigated with 100% ETc.
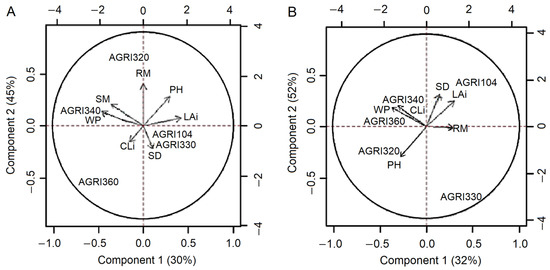
Figure 3.
Biplot of the first (x-axis) and second (y-axis) principal component referring to the five corn hybrids cultivated with the replacements of 100% (A) and 50% (B) of the crop evapotranspiration. SD: stem diameter; PH: plant height; LAi: leaf area index; CLi: chlorophyll index; SM: shoot dry mass; RM: root dry mass; WP: water productivity.
When the plants were cultivated with the replacement of only 50% ETc, there was a change in the positioning of the variables in relation to the first two PCs (Figure 3B). In PC1 (32%), the most significant variables were related to productivity and efficient use of water (CLi, shoot dry mass, root dry mass, and water productivity) and, in this scenario, with the aggregation of the hybrids AGRI340 and AGRI360 in the same quadrant of the biplot. For PC2, which explains 52% of the observed variation, the stem diameter and LAi variables were positively related to the AGRI104 hybrid, whereas the plant height was associated with the AGRI320.
A principal component analysis was more effective in understanding the variables and allowed us to conclude that the hybrids with the best performance under conditions of lower water availability were AGRI340 and AGRI360. Similar results were obtained by Badr and Brüggemann [40], who used a PC analysis to classify ten corn genotypes under the water deficit, chlorophyll fluorescence, and physiological attributes related to photosynthesis.
Regardless of the water replacement levels, the LAi variable was associated with hybrids with inefficient use of water for the production of shoot dry matter (AGRI330 and AGRI104). The results obtained from the present investigation suggest a criterion that can be used for discarding genotypes in breeding programs to launch corn hybrids capable of facing the water deficit and of being environmentally sustainable when cultivated.
It is expected that our results can be used to improve corn crop management practices in areas subject to water deficit. For example, water management strategies can be developed that take into account the characteristics of the more tolerant cultivars identified by the study. Furthermore, our study provided valuable information about the morphological mechanisms that regulate water-deficit tolerance in different maize cultivars. This information can be used by breeders to develop new maize cultivars with greater tolerance to water scarcity.
Despite the hybrids AGRI340 and AGRI360 presenting the best performances in relation to growth and production (quantity), we recommend that future studies be carried out to evaluate the quality of the silage in function of the water availability. This information will provide a solid basis for safely recommending which hybrid to cultivate considering specific needs and objectives in corn silage production.
4. Conclusions
Water deficit is one of the main causes of the reduction in corn productivity, and this was also observed in our study. Water replacement of 50% of ETc reduced the fresh and dry mass productivity in shoots and root systems. The effects were observed on the morphological characteristics of all the corn hybrids studied.
It was possible to classify the hybrids according to their performance based on the variables evaluated through an analysis of the PCs.
The best-performing hybrids were AGRI340 and AGRI360, regardless of the evapotranspired water replacement level.
The results of this study may have a significant impact on maize production in regions with low water availability, in addition to contributing to the development of new cultivars that are better adapted to these conditions. The information obtained can be used both by farmers and by researchers and breeders, aiming to improve the productivity and sustainability of agricultural production.
Author Contributions
Conceptualization: A.R.D.C. and G.d.F.T.; methodology: F.F.d.C. and G.d.F.T.; investigation: A.R.D.C., L.G.M., A.S.M. and G.d.F.T.; data curation: A.R.D.C., L.G.M. and A.S.M.; formal analysis: A.R.D.C., F.F.d.C. and G.d.F.T.; writing—original draft: A.R.D.C., F.F.d.C. and G.d.F.T.; writing—review and editing: F.F.d.C. and G.d.F.T.; visualization: A.R.D.C., L.G.M., A.S.M., F.F.d.C. and G.d.F.T.; supervision: G.d.F.T.; funding acquisition: F.F.d.C. and G.d.F.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Coordination for the Improvement of Higher Education Personnel—Brazil (CAPES)—Finance Code 001 and the National Council for Scientific and Technological Development—Brazil (CNPq)—Process 308769/2022-8.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the Graduate Program in Animal Science (PPGCA) at the Federal University of Mato Grosso do Sul (UFMS) and the Graduate Program in Agricultural Engineering (PPGEA) at the Federal University of Viçosa (UFV) for supporting the researchers.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Unesco WWAP. United Nations World Report on Water Resources Development 2020: Water and Climate Change, Executive Summary. 2020. Available online: https://unesdoc.unesco.org/ark:/48223/pf0000372882_eng (accessed on 8 March 2023).
- Elbeltagi, A.; Aslam, M.R.; Malik, A.; Mehdinejadiani, B.; Srivastava, A.; Bhatia, A.S.; Denga, J. The impact of climate changes on the water footprint of wheat and corn production in the Nile Delta, Egypt. Sci. Total Environ. 2020, 743, 140770. [Google Scholar] [CrossRef] [PubMed]
- Conab. Companhia Nacional de Abastecimento. Acompanhamento da Safra Brasileira—Grãos, Safra 2020/21—8º Levantamento. 2021. Available online: https://www.conab.gov.br/info-agro/safras/graos (accessed on 10 March 2023).
- Aparecido, L.; Torsoni, G.B.; Mesquita, D.Z.; Meneses, K.C.; Moraes, J.R.S.C. Modeling of safrinha corn yield in function of the climate conditions of Mato Grosso do Sul. Rev. Bras. Clim. 2020, 26, 113–132. [Google Scholar] [CrossRef]
- Abbade, E.B. Land and water footprints associated with rice and corn losses in Brazil. Land Use Pol. 2020, 99, 105–106. [Google Scholar] [CrossRef]
- Costa, J.R.; Pinho, J.L.N.; Parry, M.M. Dry matter production of corn cultivars under different water stress levels. Rev. Bras. Eng. Agr. Amb. 2008, 12, 443–450. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.X.; Jacinthe, P.A. Global synthesis of drought effects on corn and wheat production. PLoS ONE 2016, 11, 0156362. [Google Scholar] [CrossRef]
- Avramova, V.; Nagel, K.A.; AbdElgawad, H.; Bustos, D.; DuPlessis, M.; Fiorani, F.; Beemster, G.T.S. Screening for drought tolerance of corn hybrids by multi-scale analysis of root and shoot traits at the seedling stage. J. Exp. Bot. 2016, 67, 2453–2466. [Google Scholar] [CrossRef]
- Rocha, D.S.; Rodrigues, C.S.; Gallo, P.B.; Ticelli, M.; Paterniani, M. Drought tolerance in intervarietal maize hybrids. Caatings 2021, 34, 80–89. [Google Scholar] [CrossRef]
- Derera, J.; Tongoona, P.; Vivek, B.S. Gene action controlling grain yield and secondary traits in southern African corn hybrids under drought and non-drought environments. Euphytica 2008, 162, 411–422. [Google Scholar] [CrossRef]
- Pires, M.V.; Castro, E.M.; Freitas, B.S.M.; Lira, J.M.S.; Magalhães, P.C.; Pereira, M.P. Yield-related phenotypic traits of drought resistant corn genotypes. Environ. Exp. Bot. 2019, 171, 103962. [Google Scholar] [CrossRef]
- Brasil. Ministério da Agricultura, Pecuária e Abastecimento. In Portaria nº 189, de 20 de Agosto de 2020; SPA: Brasília, Brazil, 2020; p. 5. [Google Scholar]
- Kaya, C.; Şenbayram, M.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Sulfur-enriched leonardite and humic acid soil amendments enhance tolerance to drought and phosphorus deficiency stress in maize (Zea mays L.). Sci. Rep. 2020, 10, 6432. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- LatinaSeeds. Sementes de Milho Para Silagem. 2023. Available online: https://www.latinaseeds.com.br/sementes-milho-silagem (accessed on 28 May 2023).
- Cunha, F.F.; Soares, A.A.; Sediyama, G.C.; Mantovani, E.C.; Pereira, O.G.; Abreu, F.V.S.; Souza, D.O. The root system of Tanzania grass as affected by different intervals and levels of irrigation. Eng. Agr. 2007, 15, 200–211. [Google Scholar]
- Chapuis, R.; Delluc, C.; Debeuf, R.; Tardieu, F.; Welcker, C. Resiliences to water deficit in a phenotyping platform and in the field: How related are they in corn? Eur. J. Agron. 2012, 42, 59–67. [Google Scholar] [CrossRef]
- Zhu, J.; Tremblay, N.; Liang, Y. Comparing SPAD and atLEAF values for chlorophyll assessment in crop species. Can. J. Soil Sci. 2012, 92, 645–648. [Google Scholar] [CrossRef]
- Sah, R.P.; Chakraborty, M.; Prasad, K.; Pandit, M.; Tudu, V.K.; Chakravarty, M.K.; Narayan, S.C.; Rana, M.; Moharana, D. Impact of water deficit stress in maize: Phenology and yield components. Sci. Rep. 2020, 10, 2944. [Google Scholar] [CrossRef] [PubMed]
- Rezende, R.P.; Golin, H.O.; Abreu, V.L.S.; Theodoro, G.F.; Franco, G.L.; Brumatti, R.C.; Fernandes, P.B.; Bento, A.L.L.; Rocha, R.F.A.T. Does intercropping corn with forage sorghum effect biomass yield, bromatological silage quality and economic viability? Res. Soc. Devel. 2020, 9, 1–25. [Google Scholar] [CrossRef]
- Fernandes, P.B.; Bitencourt, L.P.; Theodoro, G.F.; Curcio, U.A.; Theodoro, W.A.; Arruda, C.O.C.B. Influence of calcium silicate on soil fertility and corn morphology. J. Agric. Stud. 2020, 8, 51–63. [Google Scholar] [CrossRef]
- Rosskopf, U.; Uteau, D.; Peth, S. Development of mechanical soil stability in an initial homogeneous loam and sand planted with two maize (Zea mays L.) genotypes with contrasting root hair attributes under in-situ field conditions. Plant Soil 2022, 478, 143–162. [Google Scholar] [CrossRef]
- Vetterlein, D.; Phalempin, M.; Lippold, E.; Schlüter, S.; Schreiter, S.; Ahmed, M.A.; Carminati, A.; Duddek, P.; Jorda, H.; Bienert, G.P.; et al. Root hairs matter at field scale for maize shoot growth and nutrient uptake, but root trait plasticity is primarily triggered by texture and drought. Plant Soil 2022, 478, 119–141. [Google Scholar] [CrossRef]
- Ding, J.; Jiao, X.; Bai, P.; Hu, Y.; Zhang, J.; Li, J. Effect of vapor pressure deficit on the photosynthesis, growth, and nutrient absorption of tomato seedlings. Sci. Hortic. 2022, 293, 110736. [Google Scholar] [CrossRef]
- Santos, C.C.; Silva, L.F.P.; Piesanti, M.; Scalon, S.P.Q.; Goelzer, A.; Silverio, J.M.; Reis, L.C. Salicylic acid on gas exchange and growth of Hymenaea courbaril L. seedlings under flooding. Rev. Bras. Eng. Agric. Ambient. 2023, 27, 327–334. [Google Scholar] [CrossRef]
- Zheng, J.; Fan, J.; Zhang, F.; Zhuang, Q. Evapotranspiration partitioning and water productivity of rainfed maize under contrasting mulching conditions in Northwest China. Agric. Water Manag. 2021, 243, 106473. [Google Scholar] [CrossRef]
- Gomaa, M.A.; Kandil, E.E.; Eldein, A.A.M.Z.; Aboudonia, M.E.M.; Ali, H.M.; Abdelsalam, N.R. Increase maize productivity and water use efficiency through application of potassium silicate under water stress. Sci. Rep. 2021, 11, 224. [Google Scholar] [CrossRef]
- Almeida, A.C.S.; Bonifácio, J.; Pusch, M.; Oliveira, F.C.; Geseinhoff, L.O.; Biscaro, G.A. Produtividade e eficiência de uso da água em milho cultivado com diferentes estratégias de manejo hídrico. Rev. Bras. Agr. Irrig. 2017, 11, 1448–1457. [Google Scholar] [CrossRef]
- Ali, M.A.; Ibrahim, S.; Abou-Amer, I. Water deficit stress mitigation by foliar application of potassium silicate for sugar beet grown in a saline calcareous soil. Egyp. J. Soil Sci. 2019, 59, 15–23. [Google Scholar] [CrossRef]
- Alves, B.E.; Cardoso, M.J.; Andrade, J.A.S.; Born, F.N.; Ramos, H.M. Desempenho da produtividade de espigas de milho verde sob diferentes regimes hídricos. Rev. Bras. Milho Sorgo. 2017, 16, 94–108. [Google Scholar] [CrossRef]
- Soaud, A.A.; Abou-Zeid, S.T.; Ali, A.M.; Hassan, D.A. Chlorophyll meter-based precision nitrogen management in maize grown in alluvial soil in Egypt. In Proceedings of the African Conference on Precision Agriculture, Benguérir, Morocco, 7–9 December 2022; pp. 137–142. [Google Scholar]
- Chiango, H.; Figueiredo, A.; Sousa, L.; Sinclair, T.; Silva, J.M. Assessing drought tolerance of traditional corn genotypes of Mozambique using chlorophyll fluorescence parameters. S. Afr. J. Bot. 2021, 138, 311–317. [Google Scholar] [CrossRef]
- Dariva, F.D.; Person, H.P.; Copati, M.G.F.; Almeida, G.Q.; Castro, F.M.N.; Picoli, E.A.T.; Cunha, F.F.; Nick, C. Yield and fruit quality attributes of selected tomato introgression lines subjected to long-term deficit irrigation. Sci. Hortic. 2021, 289, 110426. [Google Scholar] [CrossRef]
- Nascimento, H.R.; Moura, L.O.; Duarte, A.B.; Dantas, S.A.G.; Ferreira, D.O.; Rosmaninho, L.B.C.; Cavallin, I.C.; Cunha, F.F.; Silva, F.L. A new methodological approach for simulating water deficit in soybean genotypes. J. Agron. Crop Sci. 2021, 207, 946–955. [Google Scholar] [CrossRef]
- Souza, T.C.; Magalhães, P.C.; Castro, E.M.; Duarte, V.P.; Lavinsky, A.O. Corn root morphoanatomy at different development stages and yield under water stress. Pesq. Agropec. Bras. 2016, 51, 330–339. [Google Scholar] [CrossRef]
- Çakir, R. Effect of water stress at different development stages on vegetative and reproductive growth of corn. Field Crop Res. 2004, 89, 1–16. [Google Scholar] [CrossRef]
- Ge, T.; Sui, F.; Bai, L.; Tong, C.; Sun, N. Effects of water stress on growth, biomass partitioning, and water-use efficiency in summer corn (Zea mays L.) throughout the growth cycle. Acta Physiol. Plant 2012, 34, 1043–1053. [Google Scholar] [CrossRef]
- Gheysari, M.; Pirnajmedin, F.; Movahedrad, H.; Majidi, M.M.; Zareian, M.J. Crop yield and irrigation water productivity of silage maize under two water stress strategies in semi-arid environment: Two different pot and field experiments. Agric. Water Manag. 2021, 255, 106999. [Google Scholar] [CrossRef]
- Maitah, M.; Malec, K.; Maitah, K. Influence of precipitation and temperature on maize production in the Czech Republic from 2002 to 2019. Sci. Rep. 2021, 11, 10467. [Google Scholar] [CrossRef] [PubMed]
- Badr, A.; Brüggemann, W. Comparative analysis of drought stress response of corn genotypes using chlorophyll fluorescence measurements and leaf relative water content. Photosynthetica 2020, 58, 638–645. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).