
Effect of Two Biostimulants, Based on Ascophyllum nodosum Extracts, on Strawberry Performance under Mild Drought Stress

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Growing Conditions, Biostimulant Application and Water Stress Treatments
2.3. Chlorophyll Measurement and Analyses of Physical Parameters
2.4. Biochemical Analyses of Fruits
2.5. Statistical Analyses
3. Results
3.1. Vegetative Growth
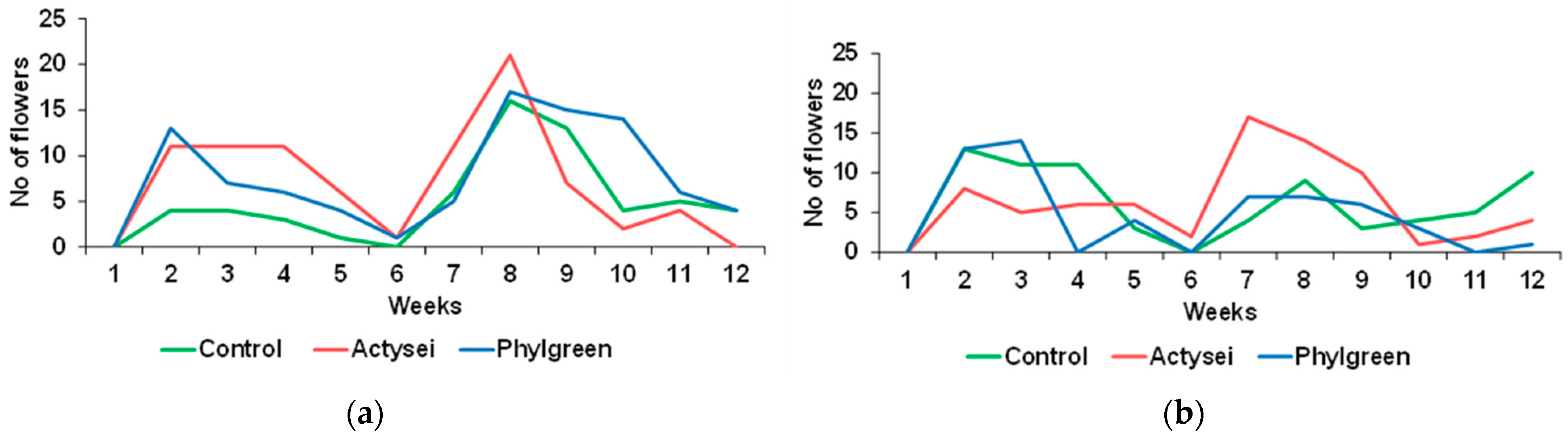
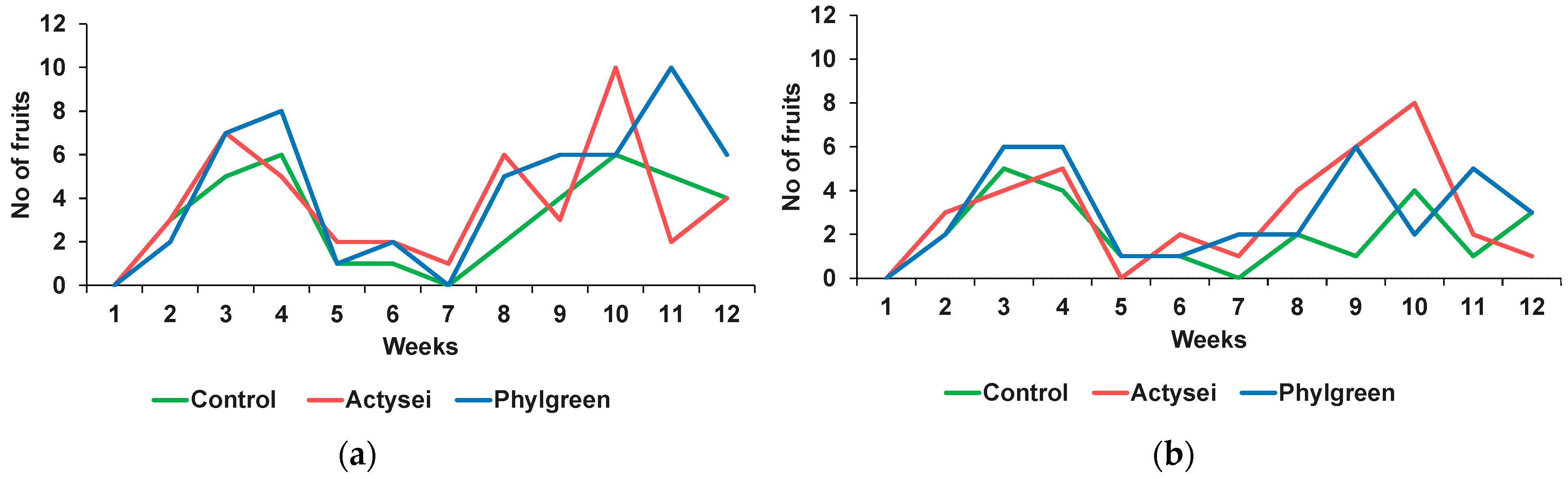
3.2. Reproductive Traits
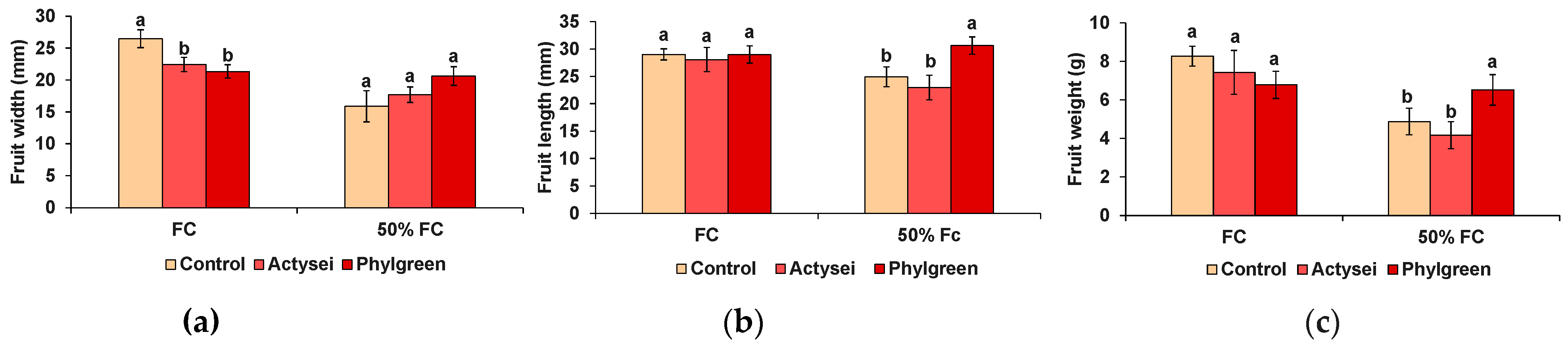
3.3. Biochemical Analyses of the Fruits
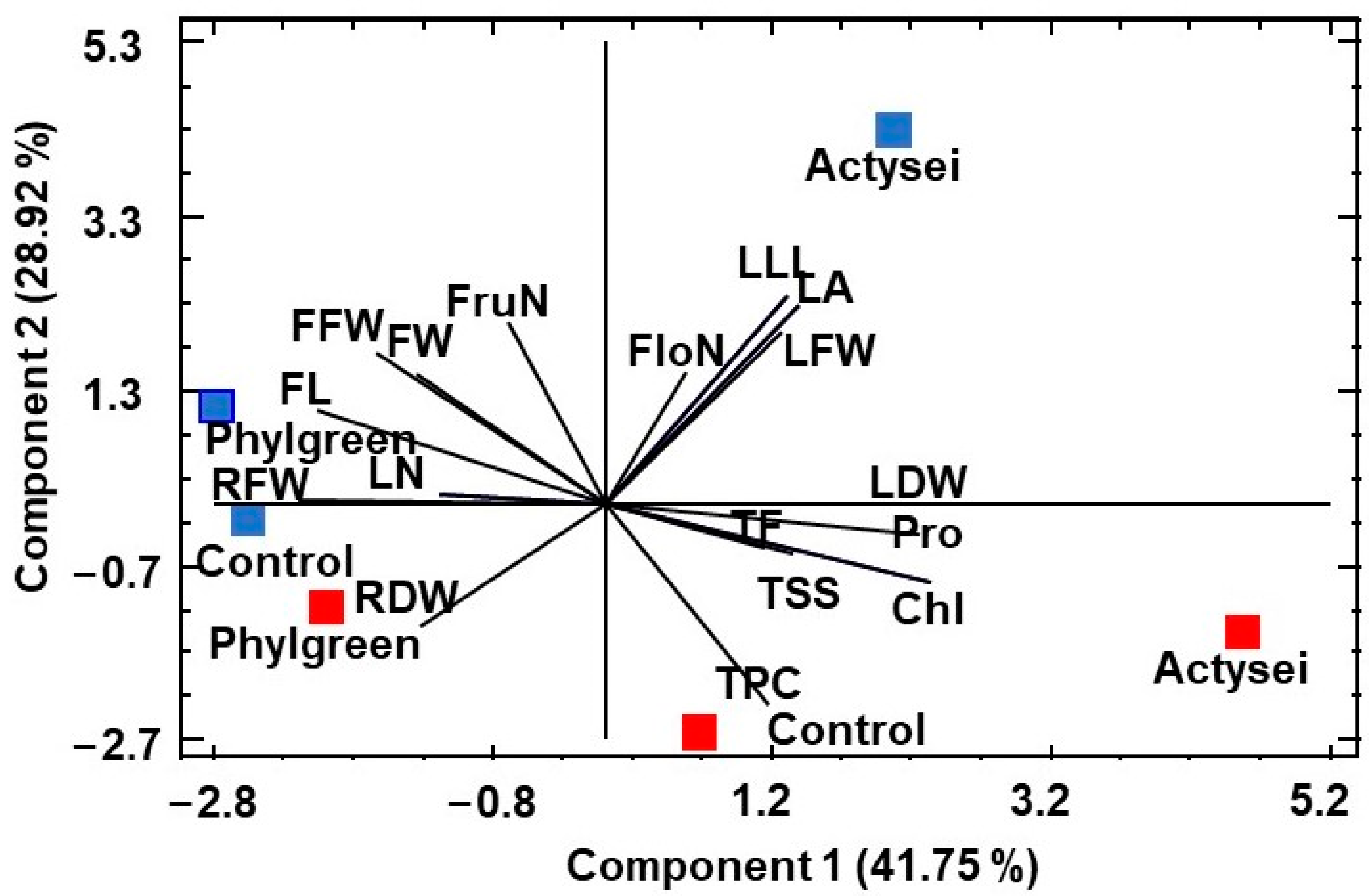
3.4. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. World Population Prospects: The 2019 Revision Population Database. Available online: https://population.un.org/wpp/ (accessed on 15 June 2023).
- Intergovernmental Panel on Climate Change. Special Report on Climate Change and Land: Summary for Policymakers; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Biostimulants in agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.; Menon, U.; Subramanian, S.; Jithesh, M.; Rayorath, P.; Hodges, D.; Crithcley, A.; Craigie, J.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth. Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- El Boukhari, M.E.M.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in seaweed extract based biostimulants: Manufacturing process and beneficial effect on soil-plant systems. Plants 2020, 9, 359. [Google Scholar] [CrossRef]
- Espinosa-Antón, A.; Hernández-Herrera, R.; González, M. Extractos bioactivos de algas marinas como bioestimulantes del crecimiento y la protección de las plantas. Biot. Veg. 2020, 20, 257–282. [Google Scholar]
- Stirk, W.A.; Rengasamy, K.R.R.; Kulkarni, M.G.; van Staden, J. Plant biostimulants from seaweed. In The Chemical Biology of Plant Biostimulants; Geelen, D., Xu, L., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 31–55. [Google Scholar]
- Vera, J.; Castro, J.; González, A.; Moenne, A. Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef]
- Crouch, I.J.; van Staden, J. Evidence for the presence of plant growth regulators in commercial seaweed products. Plant Growth Reg. 1993, 13, 21–29. [Google Scholar] [CrossRef]
- Tarakhovskaya, E.R.; Maslov, Y.I.; Shishova, M.F. Phytohormones in algae. Russ. J. Plant Physiol. 2007, 54, 163–170. [Google Scholar] [CrossRef]
- Yokoya, N.S.; Stirk, W.A.; van Standen, J.; Novák, O.; Turečková, V.; Pĕnčík, A.; Strnad, M. Endogenous cytokinins, auxins and abscisic acid in red algae from Brazil. J. Phycol. 2010, 46, 1198–1205. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Yalçin, S.; Okudan, E.S.; Karakaș, O.; Önem, A.N.; Bașkan, K.S. Identification and quantification of some phytohormones in seaweeds using UPLC-MS/MS. J. Liq. Chromatogr. Relat. Technol. 2019, 42, 475–484. [Google Scholar] [CrossRef]
- Nabti, E.; Jha, B.; Hartmann, A. Impact of seaweeds on agricultural crop production as biofertilizer. Int. J. Environ. Sci. Technol. 2017, 14, 1119–1134. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Zou, P.; Lu, X.; Zhao, H.; Yuan, Y.; Meng, L.; Zhang, C.; Li, Y. Polysaccharides derived from the brown algae Lessonia nigrescens enhance salt stress tolerance to wheat seedlings by enhancing the antioxidant system and modulating intracellular ion concentration. Front. Plant Sci. 2019, 10, 48. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.C.G.A.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N. Current trends on seaweeds: Looking at chemical composition, phytopharmacology, and cosmetic applications. Molecules 2019, 24, 4182. [Google Scholar] [CrossRef]
- Sharma, H.S.; Fleming, C.; Selby, C.; Rao, J.R.; Martin, T. Plant biostimulants: A review on the processing of macroalgae and use of extracts for crop management to reduce abiotic and biotic stresses. J. Appl. Phycol. 2014, 26, 465–2014. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babbohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Di Stasio, E.; Van Oosten, M.J.; Silletti, S.; Raimondi, G.; Carrillo, P.; Maggio, A. Ascophyllum nodosum-based algal extracts act as enhancers of growth, fruit quality, and adaptation to stress in salinized tomato plants. J. Appl. Phycol. 2018, 30, 2675–2686. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2014, 26, 619–2014. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Sangiorgio, D.; Cellini, A.; Spinelli, F.; Donati, I. Promoting strawberry (Fragaria × ananassa) stress resistance, growth, and yield using native bacterial biostimulants. Agronomy 2023, 13, 529. [Google Scholar] [CrossRef]
- Ganhão, R.; Pinheiro, J.; Tino, C.; Faria, H.; Gil, M.M. Characterization of nutritional, physicochemical, and phytochemical composition and antioxidant capacity of three strawberry “Fragaria × ananassa Duch.” cultivars (“Primoris”, “Endurance”, and “Portola”) from western region of Portugal. Foods 2019, 8, 682. [Google Scholar] [CrossRef]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A novel type of seaweed extract as a natural alternative to the use of iron chelates in strawberry production. Sci. Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Soltaniband, V.; Brégard, A.; Gaudreau, L.; Dorais, M. Biostimulants promote plant development, crop productivity, and fruit quality of protected strawberries. Agronomy 2022, 12, 1684. [Google Scholar] [CrossRef]
- Bogunovic, I.; Duralija, B.; Gadze, J.; Ivica, K. Biostimulant usage for preserving strawberries to climate damages. Hort. Sci. 2015, 42, 132–140. [Google Scholar] [CrossRef]
- Tomić, J.; Pešaković, M.; Milivojević, J.; Karaklajić-Stajić, Ž. How to improve strawberry productivity, nutrients composition, and beneficial rhizosphere microflora by biofertilization and mineral fertilization? J. Plant Nutr. 2018, 41, 2009–2021. [Google Scholar] [CrossRef]
- Saidimoradi, D.; Ghaderi, N.; Javadi, T. Salinity stress mitigation by humic acid application in strawberry (Fragaria × ananassa Duch.). Sci. Hortic. 2019, 256, 108594. [Google Scholar] [CrossRef]
- Eurosemillas.com. Lo que pide la tierra. Available online: http://www.eurosemillas.com/es/variedades/fresa/item/99-camarosa.html (accessed on 29 June 2023).
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.; de Mello, J. Application and analysis of the Folin Ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Matteazzi, A.; Andreotti, C. Foliar applications of biostimulants promote growth, yield and fruit quality of strawberry plants grown under nutrient limitation. Agronomy 2019, 9, 483. [Google Scholar] [CrossRef]
- Marfa, O.; Caceres, R.; Polo, J.; Rodenas, J. Animal protein hydrolysate as a biostimulant for transplanted strawberry plants subjected to cold stress. Acta Hortic. 2009, 842, 315–318. [Google Scholar] [CrossRef]
- Rahman, M.; Sabir, A.A.; Mukta, J.A.; Khan, M.M.A.; Mohi-Ud-Din, M.; Miah, M.G.; Rahman, M.; Islam, M.T. Plant probiotic bacteria Bacillus and Paraburkholderi improve growth, yield and content of antioxidants in strawberry fruit. Sci. Rep. 2018, 8, 2504. [Google Scholar] [CrossRef]
- Sinclair, G.; Charest, C.; Dalpe, Y.; Khanizadeh, S. Influence of colonization by arbuscular mycorrhizal fungi and a root endophyte on selected strawberry cultivars under salt conditions. Can. J. Plant Sci. 2014, 93, 997–999. [Google Scholar] [CrossRef]
- Alam, M.; Braun, G.; Norrie, J.; Hodges, D. Effect of Ascophyllum extract application on plant growth, fruit yield and soil microbial communities of strawberry. Can. J. Plant Sci. 2013, 93, 23–36. [Google Scholar] [CrossRef]
- Mattner, S.W.; Milinkovic, M.; Arioli, T. Increased growth response of strawberry roots to a commercial extract from Durvillaea potatorum and Ascophyllum nodosum. J. Appl. Phycol. 2018, 30, 2943–2951. [Google Scholar] [CrossRef]
- Ashour, M.; Al-Souti, A.S.; Hassan, S.M.; Ammar, G.A.G.; Goda, A.M.A.-S.; El-Shenody, R.; Abomohra, A.E.-F.; El-Haroun, E.; Elshobary, M.E. Commercial seaweed liquid extract as strawberry biostimulants and bioethanol production. Life 2023, 13, 85. [Google Scholar] [CrossRef]
- El-Miniawy; Ragab, M.E.; Youssef, S.; Metwally, A. Influence of foliar spraying of seaweed extract on growth, yield and quality of strawberry plants. J. Appl. Sci. Res. 2014, 10, 88–94. [Google Scholar]
- Dong, C.; Wang, G.; Du, M.; Niu, C.; Zhang, P.; Zhang, X.; Ma, D.; Ma, F.; Bao, Z. Biostimulants promote plant vigor of tomato and strawberry after transplanting. Sci. Hortic. 2020, 267, 109355. [Google Scholar] [CrossRef]
- Phylgreen. Tradecorp. Bionutrición-Ecológicos. Available online: https://tradecorp.es/producto/phylgreen/ (accessed on 24 October 2023).
- Actisey. Vademecum de Productos Fitosanitarios y Nutricionales España Carlos de Liñán. Portal TecnoAgrícola. Available online: https://www.buscador.portaltecnoagricola.com/vademecum/esp/producto/13411/ACTYSEI (accessed on 24 October 2023).
- Grzebisz, W.; Gransee, A.; Szczepaniak, W.; Diatta, J. The effects of potassium fertilization on water-use efficiency in crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 355–374. [Google Scholar] [CrossRef]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in root growth and development. Plants 2019, 8, 435. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.F.; Wu, W.H. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2021, 63, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Hossen, M.S.; Masud, A.A.C.; Moumita; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef]
- Mosa, W.; Salem, M.; Al-huqail, A.; Ali, H. Application of glycine, folic acid, and moringa extract as biostimulants for enhancing the production of ‘Flame Seedless’ grape cultivar. BioResources 2021, 16, 3391–3410. [Google Scholar] [CrossRef]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef]
- Ali, J.; Jan, I.; Ullah, H.; Ahmed, N.; Alam, M.; Ullah, R.; El-Sharnouby, M.; Kesba, H.; Shukry, M.; Sayed, S.; et al. Influence of Ascophyllum nodosum extract foliar spray on the physiological and biochemical attributes of okra under drought stress. Plants 2022, 11, 790. [Google Scholar] [CrossRef]
- Kumari, S.; Sehrawat, K.D.; Phogat, D.; Sehrawat, A.R.; Chaudhary, R.; Sushkova, S.N.; Voloshina, M.S.; Rajput, V.D.; Shmaraeva, A.N.; Marc, R.A.; et al. Ascophyllum nodosum (L.) Le Jolis, a pivotal biostimulant toward sustainable agriculture: A comprehensive review. Agriculture 2023, 13, 1179. [Google Scholar] [CrossRef]
- Blunden, G.; Jenkins, T.; Liu, Y. Enhanced leaf chlorophyll levels in plants treated with seaweed extract. J. Appl. Phycol. 1997, 8, 535543. [Google Scholar] [CrossRef]
- Pohl, A.; Grabowska, A.; Kalisz, A.; Sękara, A. The eggplant yield and fruit composition as affected by genetic factor and biostimulant application. Not. Bot. Horti. Agrobot. 2019, 47, 929–938. [Google Scholar] [CrossRef]
- Carmody, N.; Goñi, O.; Łangowski, Ł.; O’Connell, S. Ascophyllum nodosum extract biostimulant processing and its impact on enhancing heat stress tolerance during tomato fruit set. Front. Plant Sci. 2020, 25, 807. [Google Scholar] [CrossRef]
- De Clercq, P.; Pauwels, E.; Top, S.; Steppe, K.; Van Labeke, M.-C. Effect of seaweed-based biostimulants on growth and development of Hydrangea paniculata under continuous or periodic drought stress. Horticulturae 2023, 9, 509. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Marra, R.; Vitale, S.; Pironti, A.; Fiorentino, N.; Mori, M. Yield and quality of processing tomato as improved by biostimulants based on Trichoderma sp. and Ascophyllum nodosum and biodegradable mulching films. Agronomy 2023, 13, 901. [Google Scholar] [CrossRef]
- Weber, N.; Schmitzer, V.; Jakopic, J.; Stampar, F. First fruit in season: Seaweed extract and silicon advance organic strawberry (Fragaria × ananassa Duch.) fruit formation and yield. Sci. Hortic. 2018, 242, 103–109. [Google Scholar] [CrossRef]
- Roussos, P.; Denaxa, N.-K.; Damvakaris, T. Strawberry fruit quality attributes after application of plant growth stimulating compounds. Sci. Hortic. 2009, 119, 138–146. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant biostimulants: Importance of the quality and yield of horticultural crops and the improvement of plant tolerance to abiotic stress—A review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants research in some horticultural plant species-A review. Food Energy Secur. 2018, 8, e00162. [Google Scholar] [CrossRef]
- Garza-Alonso, C.A.; Olivares-Sáenz, E.; González-Morales, S.; Cabrera-De la Fuente, M.; Juárez-Maldonado, A.; González-Fuentes, J.A.; Tortella, G.; Valdés-Caballero, M.V.; Benavides-Mendoza, A. Strawberry biostimulation: From mechanisms of action to plant growth and fruit quality. Plants 2022, 11, 3463. [Google Scholar] [CrossRef] [PubMed]
- Righini, H.; Roberti, R.; Baraldi, E. Use of algae in strawberry management. J. Appl. Phycol. 2018, 30, 3551–3564. [Google Scholar] [CrossRef]
- Nanda, S.; Kumar, G.; Hussain, S. Utilization of seaweed-based biostimulants in improving plant and soil health: Current updates and future prospective. Int. J. Environ. Sci. Technol. 2022, 19, 12839–12852. [Google Scholar] [CrossRef]
- Kapur, B.; Sarıdaș, M.A.; Çeliktopuz, E.; Kafkas, E.; Paydaș Kargı, S. Health and taste related compounds in strawberries under various irrigation regimes and bio-stimulant application. Food Chem. 2018, 263, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Actisey. Bioestimulantes Agrícolas. Seipasa. Available online: https://www.seipasa.com/es_ES/extracto-de-algas-actysei/ (accessed on 24 October 2023).
- Martínez-Ferri, E.; Soria, C.; Ariza, M.T.; Medina, J.J.; Miranda, L.; Domíguez, P.; Muriel, J.L. Water relations, growth and physiological response of seven strawberry cultivars (Fragaria × ananassa Duch.) to different water availability. Agric. Water Manag. 2016, 164, 73–82. [Google Scholar] [CrossRef]
- De Saeger, J.; Van Praet, S.; Vereecke, D.; Park, J.; Jacques, S.; Han, T.; Depuydt, S. Toward the molecular understanding of the action mechanism of Ascophyllum nodosum extracts on plants. J. Appl. Phycol. 2020, 32, 573–597. [Google Scholar] [CrossRef]
- Zhang, X.; Ervin, E.H. Cytokinin-containing seaweed and humic acid extracts associated with creeping bentgrass leaf cytokinins and drought resistance. Crop Sci. 2004, 44, 1737–1745. [Google Scholar] [CrossRef]
- Aziz, N.G.A.; Mahgoub, M.H.; Siam, H.S. Growth, flowering and chemical constituents performance of Amaranthus tricolor plants as influenced by seaweed (Ascophyllum nodosum) extract application under salt stress conditions. J. Appl. Sci. Res. 2011, 7, 1472–1484. [Google Scholar]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Enhancement of phenolic and flavonoid compounds in cabbage (Brassica oleracea) following application of commercial seaweed extracts of the brown seaweed (Ascophyllum nodosum). Agric. Food Sci. 2013, 22, 288–295. [Google Scholar] [CrossRef]
- Iwamoto, K.; Shiraiwa, Y. Salt-regulated mannitol metabolism in algae. Mar. Biotechnol. 2005, 7, 407–415. [Google Scholar] [CrossRef]
- Badran, E.G.; Abogadallah, G.M.; Nada, R.M.; Nemat Alla, M.M. Role of glycine in improving the ionic and ROS homeostasis during NaCl stress in wheat. Protoplasma 2015, 252, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Hannum, S.M. Potential impact of strawberries on human health: A review of the science. Crit. Rev. Food Sci. Nutr. 2004, 44, 1–17. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Temperature (°C) | Relative Humidity (%) | ||||
|---|---|---|---|---|---|---|
| Mean | Max | Min | Mean | Max | Min | |
| February 2021 | 19.46 | 33.03 | 14.84 | 55.53 | 75.03 | 19.46 |
| March 2021 | 20.49 | 37.07 | 11.83 | 53.41 | 87.66 | 14.92 |
| April 2021 | 20.84 | 36.04 | 12.92 | 59.46 | 59.46 | 59.46 |
| May 2021 | 24.16 | 39.46 | 16.82 | 56.39 | 86.17 | 19.52 |
| June 2021 | 26.07 | 39.91 | 17.56 | 57.72 | 84.31 | 15.38 |
| Parameter | Substrate Moisture (A) | Treatment (B) | A × B |
|---|---|---|---|
| LN | 0.12 ns | 1.24 ns | 1.35 ns |
| LLL | 3.08 ns | 6.59 ** | 0.38 ns |
| LA | 2.04 ns | 4.99 * | 1.03 ns |
| Chl | 19.61 *** | 14.66 *** | 0.98 ns |
| RFW | 20.76 *** | 25.52 *** | 2.10 ns |
| LFW | 3.45 ns | 6.10 ** | 4.70 * |
| RDW | 1.37 ns | 6.38 ** | 1.77 ns |
| LDW | 2.24 ns | 2.86 ** | 0.76 ** |
| Parameter | Substrate Moisture (A) | Treatment (B) | A × B |
|---|---|---|---|
| FW | 12.04 *** | 0.68 ns | 2.53 ns |
| FL | 2.69 ns | 2.93 ns | 2.03 ns |
| FFW | 23.32 *** | 0.39 ns | 6.66 ** |
| Pro | 0.05 * | 6.80 ** | 1.74 ns |
| TSS | 0.68 ns | 0.70 ns | 0.50 * |
| TPC | 2.30 ns | 0.34 ns | 0.86 ns |
| TF | 0.12 ns | 0.30 ns | 0.64 ns |
| Trait | Component 1 | Component 2 |
|---|---|---|
| LN | −0.149 | −0.021 |
| LLL | 0.125 | 0.441 |
| LA | 0.134 | 0.426 |
| Chl | 0.369 | −0.048 |
| RFW | −0.328 | −0.073 |
| RDW | −0.172 | −0.295 |
| LFW | 0.112 | 0.369 |
| LDW | 0.333 | 0.094 |
| FloN | 0.041 | 0.290 |
| FruN | −0.145 | 0.330 |
| FW | −0.234 | 0.187 |
| FL | −0.329 | 0.083 |
| FFW | −0.291 | 0.208 |
| Pro | 0.351 | 0.044 |
| TSS | 0.227 | −0.048 |
| TPC | 0.244 | −0.319 |
| TF | 0.207 | −0.019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shakya, R.; Capilla, E.; Torres-Pagán, N.; Muñoz, M.; Boscaiu, M.; Lupuţ, I.; Vicente, O.; Verdeguer, M. Effect of Two Biostimulants, Based on Ascophyllum nodosum Extracts, on Strawberry Performance under Mild Drought Stress. Agriculture 2023, 13, 2108. https://doi.org/10.3390/agriculture13112108
Shakya R, Capilla E, Torres-Pagán N, Muñoz M, Boscaiu M, Lupuţ I, Vicente O, Verdeguer M. Effect of Two Biostimulants, Based on Ascophyllum nodosum Extracts, on Strawberry Performance under Mild Drought Stress. Agriculture. 2023; 13(11):2108. https://doi.org/10.3390/agriculture13112108
Chicago/Turabian StyleShakya, Rashmi, Elvira Capilla, Natalia Torres-Pagán, Marta Muñoz, Monica Boscaiu, Ioan Lupuţ, Oscar Vicente, and Mercedes Verdeguer. 2023. "Effect of Two Biostimulants, Based on Ascophyllum nodosum Extracts, on Strawberry Performance under Mild Drought Stress" Agriculture 13, no. 11: 2108. https://doi.org/10.3390/agriculture13112108
APA StyleShakya, R., Capilla, E., Torres-Pagán, N., Muñoz, M., Boscaiu, M., Lupuţ, I., Vicente, O., & Verdeguer, M. (2023). Effect of Two Biostimulants, Based on Ascophyllum nodosum Extracts, on Strawberry Performance under Mild Drought Stress. Agriculture, 13(11), 2108. https://doi.org/10.3390/agriculture13112108