
Exposure to Cattle Slurry of Different Concentrations Influence Germination and Initial Growth of Selected Grass and Legume Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment I: Germination of Forage Grasses and Legumes
2.2. Experiment II: Initial Growth of of Forage Grasses and Legumes
2.3. Data Analyses
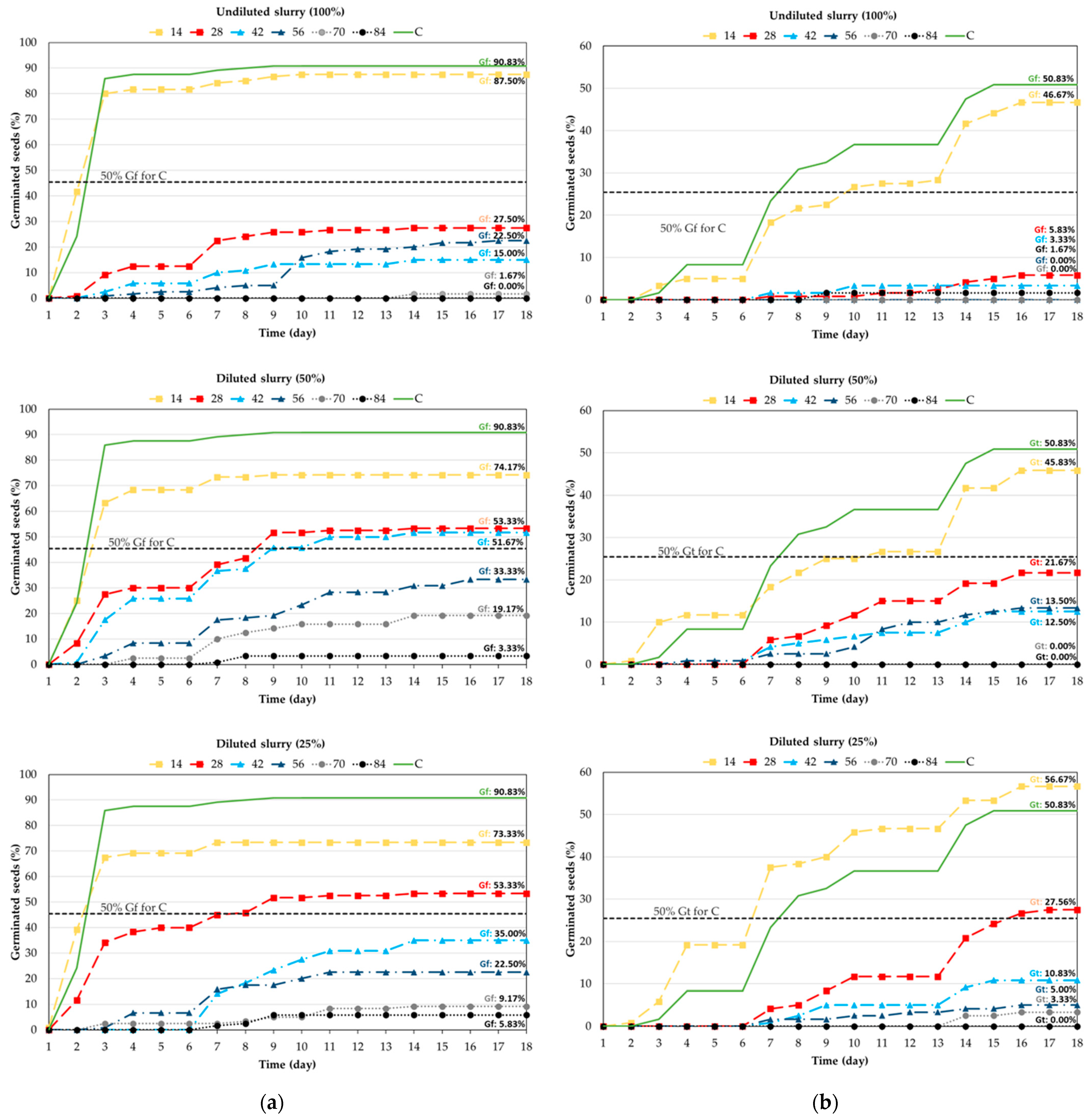
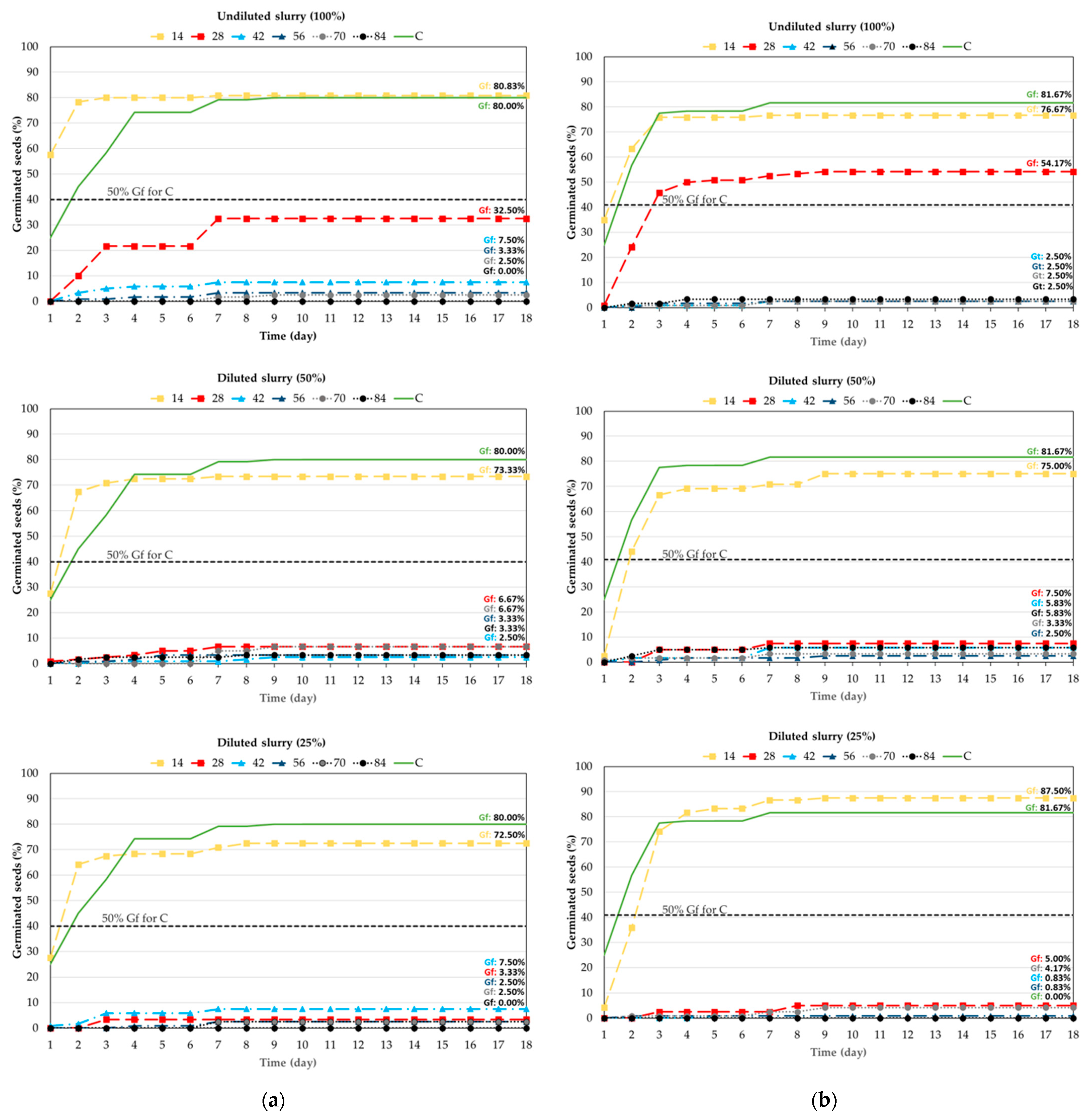
3. Results
3.1. Experiment I: Germination of Forage Grasses and Legumes
3.2. Experiment II: Initial Growth of Forage Grasses and Legumes
4. Discussion
5. Conclusions
- -
- Experiment I: A short 14-day soaking in cattle slurry has no negative effect on germination of L. multiflorum, D. glomerata, T. repens and T. pratense. However, longer soaking (more than 14 days) resulted in reduced and delayed germination of these tested plants, especially in undiluted slurries for grasses and diluted slurries for clovers.
- -
- Experiment II: Soaking seeds in cattle slurry of different concentrations for 14 days only affected the initial growth of L. multiflorum. Seedlings of seeds soaked in undiluted slurry have higher RRL than the corresponding control.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huyghe, C.; De Vliegher, A.; van Gils, B.; Huyghe, C. Grasslands and Herbivore Production in Europe and Effects of Common Policies; Éditions Quæ: Versailles, France, 2014. [Google Scholar]
- Veen, P.; Molnár, Z.; Pärtel, M.; Nagy, S. Grassland Ecosystems in Central and Eastern Europe-Prepared in the Framework of the High Level Conference on EU Enlargement’The Relation between Agriculture and Nature Management’-22/24 January 2001; Veen Ecology: Nunspeet, The Netherlands, 2001. [Google Scholar]
- Dauber, J.; Bengtsson, J.A.N.; Lenoir, L. Evaluating Effects of Habitat Loss and Land-Use Continuity on Ant Species Richness in Seminatural Grassland Remnants. Conserv. Biol. 2006, 20, 1150–1160. [Google Scholar] [CrossRef]
- Plantureux, S.; Peeters, A.; McCracken, D. Biodiversity in intensive grasslands: Effect of management, improvement and challenges. Agron. Res. 2005, 3, 153–164. [Google Scholar]
- Walker, K.J.; Stevens, P.A.; Stevens, D.P.; Mountford, J.O.; Manchester, S.J.; Pywell, R.F. The restoration and re-creation of species-rich lowland grassland on land formerly managed for intensive agriculture in the UK. Biol. Conserv. 2004, 119, 1–18. [Google Scholar] [CrossRef]
- Hodgson, J.G.; Montserrat-Martí, G.; Tallowin, J.; Thompson, K.; Díaz, S.; Cabido, M.; Grime, J.P.; Wilson, P.J.; Band, S.R.; Bogard, A.; et al. How much will it cost to save grassland diversity? Biol. Conserv. 2005, 122, 263–273. [Google Scholar] [CrossRef]
- Hopkins, A.; Holz, B. Grassland for agriculture and nature conservation: Production, quality and multi-functionality. Agron. Res. 2006, 4, 3–20. [Google Scholar]
- Davies, A.; Waite, S. The persistence of calcareous grassland species in the soil seed bank under developing and established scrub. Plant Ecol. 1998, 136, 27–39. [Google Scholar] [CrossRef]
- Bossuyt, B.; Butaye, J.; Honnay, O. Seed bank composition of open and overgrown calcareous grassland soils—A case study from Southern Belgium. J. Environ. Manag. 2006, 79, 364–371. [Google Scholar] [CrossRef]
- Fagan, K.C.; Pywell, R.F.; Bullock, J.M.; Marrs, R.H. The seed banks of English lowland calcareous grasslands along a restoration chronosequence. Plant Ecol. 2010, 208, 199–211. [Google Scholar] [CrossRef]
- Hermoso, V.; Carvalho, S.B.; Giakoumi, S.; Goldsborough, D.; Katsanevakis, S.; Leontiou, S.; Markantonatou, V.; Rumes, B.; Vogiatzakis, I.N.; Yates, K.L. The EU Biodiversity Strategy for 2030: Opportunities and challenges on the path towards biodiversity recovery. Environ. Sci. Policy 2022, 127, 263–271. [Google Scholar] [CrossRef]
- European Commission Environment, D.-G. EU Biodiversity Strategy for 2030: Bringing Nature Back into Our Lives; Publications Office of the European Union: Luxembourg, 2021. [Google Scholar] [CrossRef]
- Kull, K.; Zobel, M. High species richness in an Estonian wooded meadow. J. Veg. Sci. 1991, 2, 715–718. [Google Scholar] [CrossRef]
- Hansson, M.; Fogelfors, H. Management of a semi-natural grassland; results from a 15-year-old experiment in southern Sweden. J. Veg. Sci. 2000, 11, 31–38. [Google Scholar] [CrossRef]
- Mitlacher, K.; Poschlod, P.; Rosén, E.; Bakker, J.P. Restoration of wooded meadows—A comparative analysis along a chronosequence on Öland (Sweden). Appl. Veg. Sci. 2002, 5, 63–73. [Google Scholar] [CrossRef]
- Dahlstrom, A.; Lennartsson, T.; Wissman, J. Biodiversity and traditional land use in South-Central Sweden: The significance of management timing. Environ. Hist. 2008, 14, 385–403. [Google Scholar] [CrossRef]
- Barbaro, L.; Dutoit, T.; Cozic, P. A six-year experimental restoration of biodiversity by shrub-clearing and grazing in calcareous grasslands of the French Prealps. Biodivers. Conserv. 2001, 10, 119–135. [Google Scholar] [CrossRef]
- Fagan, K.C.; Pywell, R.F.; Bullock, J.M.; Marrs, R.H. Do restored calcareous grasslands on former arable fields resemble ancient targets? The effect of time, methods and environment on outcomes. J. Appl. Ecol. 2008, 45, 1293–1303. [Google Scholar] [CrossRef]
- Stampfli, A.; Zeiter, M. Plant Species Decline Due to Abandonment of Meadows Cannot Easily Be Reversed by Mowing. A Case Study from the Southern Alps. J. Veg. Sci. 1999, 10, 151–164. [Google Scholar] [CrossRef]
- Pywell, R.F.; Bullock, J.M.; Hopkins, A.; Walker, K.J.; Sparks, T.H.; Burke, M.J.W.; Peel, S. Restoration of species-rich grassland on arable land: Assessing the limiting processes using a multi-site experiment. J. Appl. Ecol. 2002, 39, 294–309. [Google Scholar] [CrossRef]
- Pywell, R.F.; Bullock, J.M.; Tallowin, J.B.; Walker, K.J.; Warman, E.A.; Masters, G. Enhancing diversity of species-poor grasslands: An experimental assessment of multiple constraints. J. Appl. Ecol. 2007, 44, 81–94. [Google Scholar] [CrossRef]
- Edwards, A.R.; Mortimer, S.R.; Lawson, C.S.; Westbury, D.B.; Harris, S.J.; Woodcock, B.A.; Brown, V.K. Hay strewing, brush harvesting of seed and soil disturbance as tools for the enhancement of botanical diversity in grasslands. Biol. Conserv. 2007, 134, 372–382. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties, 2nd ed.; Wiley: Chichester, West sussex, UK, 2001. [Google Scholar]
- Čop, J.; Vidrih, M.; Hacin, J. Influence of cutting regime and fertilizer application on the botanical composition, yield and nutritive value of herbage of wet grasslands in Central Europe. Grass Forage Sci. 2009, 64, 454–465. [Google Scholar] [CrossRef]
- Chytrý, M.; Hejcman, M.; Hennekens, S.M.; Schellberg, J. Changes in vegetation types and Ellenberg indicator values after 65 years of fertilizer application in the Rengen Grassland Experiment, Germany. Appl. Veg. Sci. 2009, 12, 167–176. [Google Scholar] [CrossRef]
- Poschlod, P.; Bonn, S. Changing dispersal processes in the central European landscape since the last ice age: An explanation for the actual decrease of plant species richness in different habitats? Acta Bot. Neerl. 1998, 47, 27–44. [Google Scholar]
- Bruun, H.H.; Fritzbøger, B. The Past Impact of Livestock Husbandry on Dispersal of Plant Seeds in the Landscape of Denmark. AMBIO A J. Hum. Environ. 2002, 31, 425–431, 427. [Google Scholar] [CrossRef] [PubMed]
- Hyde, B.; Carton, O.T. Manure management facilities on farms and their relevance to efficient nutrient use. Proc. Fertil. Assoc. Irel. 2005, 42, 27–43. [Google Scholar]
- Kirk, J. The Survival and Transport of Seeds of Wild Oat (Avena fatua L.) in Agricultural Situations. Ph.D. Thesis, Queen’s University Belfast Belfast, Belfast, Northern Ireland, 1975. [Google Scholar]
- Humphreys, J.; Culleton, N.; Jansen, T.; O’Riordan, E.; Storey, T. Aspects of the role of cattle slurry in dispersal and seedling establishment of Rumex obtusifolius seed in grassland. Ir. J. Agric. Food Res. 1997, 36, 39–49. [Google Scholar]
- Nesheim, L. Slurry seeding in grassland in Norway. In Grassland in a Changing World, Proceedings of the 23th General Meeting of the European Grassland Federation, Kiel, Germany, 29 August–2 September 2010; Mecke Druck und Verlag: Duderstadt, Germany, 2010; pp. 181–183. [Google Scholar]
- Volden, B. Renewal of grassland without ploughing by wet sowing techniques, with special attention to establishement of clover in the plant stand. In Abstracts of the 5th Circumpolar Agricultural Conference, Umeå, Sweden, 27–29 September 2004; Swedish University of Agricultural Science: Uppsala, Sweden, 2004; pp. 37–38. [Google Scholar]
- Jones, E.L.; Roberts, J.E. Sward maintenance of Lolium multiflorum by slurry seeding. Grass Forage Sci. 1989, 44, 27–30. [Google Scholar] [CrossRef]
- Kader, M. A comparison of seed germination calculation formulae and the associated interpretation of resulting data. J. Proceeding R. Soc. New South Wales 2005, 138, 65–75. [Google Scholar] [CrossRef]
- Šarapatka, B.; Holub, M.; Lhotská, M. The Effect of Farmyard Manure Anaerobic Treatment on Weed Seed Viability. Biol. Agric. Hortic. 1993, 10, 1–8. [Google Scholar] [CrossRef]
- Weed Seed Survival in Anaerobic Digesters. Available online: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=1c10f5599ebba60ca9e7560ff8b389a1266c9f97 (accessed on 31 August 2023).
- Zaller, J.G. Seed germination of the weed Rumex obtusifolius after on-farm conventional, biodynamic and vermicomposting of cattle manure. Ann. Appl. Biol. 2007, 151, 245–249. [Google Scholar] [CrossRef]
- Wollan, E.; Davis, R.D.; Jenner, S. Effects of sewage sludge on seed germination. Environ. Pollut. (1970) 1978, 17, 195–205. [Google Scholar] [CrossRef]
- Wong, M.H.; Cheung, Y.H.; Cheung, C.L. The effects of ammonia and ethylene oxide in animal manure and sewage sludge on the seed germination and root elongation of Brassica parachinensis. Environ. Pollut. Ser. A Ecol. Biol. 1983, 30, 109–123. [Google Scholar] [CrossRef]
- Harrigan, T.M.; Mutch, D.R.; Snapp, S.S. Manure Slurry-Enriched Micro-Site Seeding of Biosuppressive Covers. Appl. Eng. Agric. 2006, 22, 827–834. [Google Scholar] [CrossRef]
- Allred, S.E.; Ohlrogge, A.J. Principles of Nutrient Uptake From Fertilizer Bands. VI. Germination and Emergence of Corn as Affected by Ammonia and Ammonium Phosphate1. Agron. J. 1964, 56, 309–313. [Google Scholar] [CrossRef]
- Suarez, E.A. Slurry Seeding Cover Crops. Master’s Thesis, Purdue University, West Lafayette, IN, USA, 2013. [Google Scholar]
- Hoekstra, N.J.; Bosker, T.; Lantinga, E.A. Effects of cattle dung from farms with different feeding strategies on germination and initial root growth of cress (Lepidium sativum L.). Agric. Ecosyst. Environ. 2002, 93, 189–196. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soaking Time (Days) | L. multiflorum | D. glomerata | ||||||
|---|---|---|---|---|---|---|---|---|
| Cattle Slurry Concentration (%) | Cattle Slurry Concentration (%) | |||||||
| 100 | 50 | 25 | Mean | 100 | 50 | 25 | Mean | |
| 14 | 423 ± 40.4 a* | 355 ± 56.8 a* | 359 ± 57.1 a* | 379 A | 125 ± 26.4 b | 131 ± 8.5 b* | 193 ± 33.2 a | 150 A |
| 28 | 109 ± 21.1 cd | 220 ± 78.3 b | 228 ± 36.1 b | 186 B | 11 ± 4.1 d | 55 ± 15.8 c | 57 ± 6.6 c | 41 B |
| 42 | 55 ± 5.5 d | 201 ± 80.9 bc | 105 ± 31.0 cd | 120 C | 11 ± 7.2 d | 32 ± 12.5 cd | 24 ± 8.8 cd | 22 C |
| 56 | 57 ± 2.9 d | 106 ± 22.2 cd | 82 ± 28.5 d | 82 C | 0 ± 0.0 d | 32 ± 17.5 cd | 12 ± 8.5 d | 15 C |
| 70 | 3 ± 1.9 d | 61 ± 16.0 d | 29 ± 6.9 d | 31 D | 0 ± 0.0 d | 0 ± 0.0 d | 5 ± 1.0 d | 2 D |
| 84 | 0 ± 0.0 d | 11 ± 6.0 d | 19 ± 8.5 d | 10 E | 5 ± 10.0 d | 0 ± 0.0 d | 0 ± 0.0 d | 2 D |
| Mean | 108 B | 159 A | 137 A | 25 B | 42 A | 48 A | ||
| C | 438 ± 13.1 | 153 ± 7.1 | ||||||
| Soaking Time (Days) | T. repens | T. pratense | ||||||
|---|---|---|---|---|---|---|---|---|
| Cattle Slurry Concentration (%) | Cattle Slurry Concentration (%) | |||||||
| 100 | 50 | 25 | Mean | 100 | 50 | 25 | Mean | |
| 14 | 428 ± 54.1 a* | 379 ± 44.2 a* | 370 ± 22.1 a* | 392 A | 397 ± 30.4 a* | 364 ± 28.7 a* | 416 ± 17.3 a* | 392 A |
| 28 | 146 ± 37.4 b | 23 ± 7.6 c | 16 ± 11.4 c | 62 B | 258 ± 33.0 b | 33 ± 5.3 c | 20 ± 11.7 c | 104 B |
| 42 | 35 ± 3.7 c | 9 ± 5.4 c | 35 ± 11.6 c | 26 C | 10 ± 6.9 c | 24 ± 8.3 c | 4 ± 0.3 c | 12 C |
| 56 | 14 ± 5.0 c | 8 ± 4.6 c | 10 ± 5.9 c | 11 C | 11 ± 6.8 c | 10 ± 3.7 c | 4 ± 0.4 c | 9 C |
| 70 | 9 ± 5.7 c | 23 ± 7.5 c | 9 ± 0.9 c | 14 C | 10 ± 6.1 c | 15 ± 9.9 c | 15 ± 2.0 c | 13 C |
| 84 | 0 ± 0.0 c | 15 ± 5.8 c | 0 ± 0.0 c | 5 C | 16 ± 9.3 c | 28 ± 9.7 c | 0 ± 0.0 c | 15 C |
| Mean | 105 A | 76 B | 73 B | 117 A | 79 B | 77 B | ||
| C | 393 ± 21.5 | 412 ± 16.7 | ||||||
| Slurry Conc. RRL and RSL (%) | L. multiflorum | D. glomerata | T. repens | T. pratense | ||||
|---|---|---|---|---|---|---|---|---|
| RRL * | RSL ns | RRL ns | RSL ns | RRL ns | RSL ns | RRL ns | RSL ns | |
| 100 | 117 ± 20.3 a | 87 ± 20.0 | 112 ± 16.5 | 105 ± 19.7 | 105 ± 24.1 | 84 ± 6.1 | 71 ± 9.8 | 76 ± 7.6 |
| 50 | 91 ± 4.8 b | 96 ± 16.1 | 100 ± 22.0 | 96 ± 7.6 | 87 ± 28.2 | 77 ± 17.4 | 95 ± 20.2 | 66 ± 2.1 |
| 25 | 87 ± 9.3 b | 85 ± 9.4 | 128 ± 20.8 | 115 ± 20.3 | 111 ± 12.2 | 91 ± 20.5 | 83 ± 13.2 | 80 ± 10.3 |
| C lenghts (mm) | 161.6 | 94.2 | 58.0 | 112.1 | 48.1 | 16.0 | 62.9 | 23.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gselman, A.; Sem, V.; Grobelnik Mlakar, S. Exposure to Cattle Slurry of Different Concentrations Influence Germination and Initial Growth of Selected Grass and Legume Species. Agriculture 2023, 13, 1999. https://doi.org/10.3390/agriculture13101999
Gselman A, Sem V, Grobelnik Mlakar S. Exposure to Cattle Slurry of Different Concentrations Influence Germination and Initial Growth of Selected Grass and Legume Species. Agriculture. 2023; 13(10):1999. https://doi.org/10.3390/agriculture13101999
Chicago/Turabian StyleGselman, Anastazija, Vilma Sem, and Silva Grobelnik Mlakar. 2023. "Exposure to Cattle Slurry of Different Concentrations Influence Germination and Initial Growth of Selected Grass and Legume Species" Agriculture 13, no. 10: 1999. https://doi.org/10.3390/agriculture13101999
APA StyleGselman, A., Sem, V., & Grobelnik Mlakar, S. (2023). Exposure to Cattle Slurry of Different Concentrations Influence Germination and Initial Growth of Selected Grass and Legume Species. Agriculture, 13(10), 1999. https://doi.org/10.3390/agriculture13101999