
Month of Harvest and Leaf Age Impact the Bromatological Composition and Polyphenol Content of Gymnopodium floribundum Rolfe Leaves

 ,
,  , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Selection of Trees for Sampling
2.3. Leaf Sampling
- (i)
- 18–21 December (winter solstice) 2017 and 2018,
- (ii)
- 18–21 March (spring equinox) 2018,
- (iii)
- 18–21 June (summer solstice) 2018, and
- (iv)
- 18–21 September (fall equinox) 2018.
- (a)
- Mixed-age leaves were sampled from the 12 non-defoliated trees randomly selected for that month. These leaves were used to prepare three composite samples (each containing all of the leaves from four trees).
- (b)
- To obtain samples of 90-day-old leaves, the 12 trees defoliated 90 days earlier (in the previous sampling month) were defoliated again. These 90-day-old leaves were used to prepare a new set of three composite samples (each containing all of the leaves from four trees).
2.4. Environmental Data
2.5. Fresh Biomass and Dry Matter of Gymnopodium floribundum Leaves
2.6. Chemical Analysis and In Vitro Digestibility
2.7. Statistical Analysis
3. Results
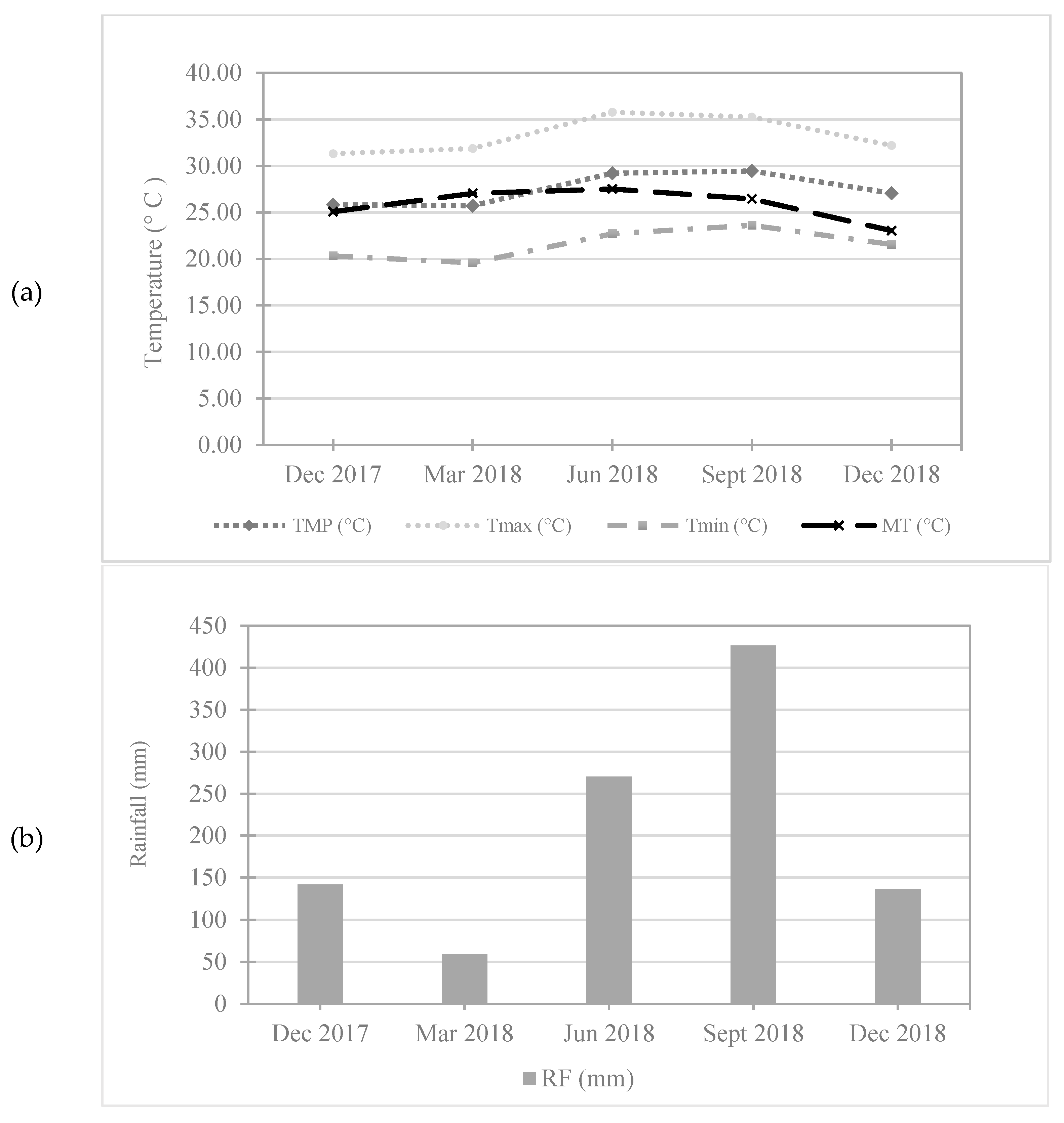
3.1. Climatic Factors during the Experimental Period
3.2. Effects of Harvest Month, Leaf Age, and their Interaction on the Chemical Composition
3.3. Relationship between Climatic Factors and the Chemical Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ortiz-Díaz, J.J. Polygonaceae. Etnoflora Yucatanense No.10, 1st ed.; Universidad Autónoma de Yucatán (UADY): Mérida, Mexico, 1994; pp. 38–41. [Google Scholar]
- White, D.A.; Hood, C.S. Vegetation patterns and environmental gradients in tropical dry forests of the northern Yucatan Peninsula. J. Veg. Sci. 2004, 15, 151–161. [Google Scholar] [CrossRef]
- Balam-Narváez, R. Una Planta Común con Varias Funciones. ¡Aunque Usted no lo Crea! 2009. Available online: https://www.cicy.mx/Documentos/CICY/Desde_Herbario/2009/2009-09-10-Balam-Gymnopodium.pdf/ (accessed on 6 June 2022).
- Ortíz-Ocampo, G.I.; Tun-Garrido, J.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.J. El Dzidzilché (Gymnopodium floribundum): Una planta con varios servicios ecosistémicos en la península de Yucatán. Bioagrociencias 2019, 12, 28–33. [Google Scholar]
- González-Pech, P.G.; Torres-Acosta, J.F.; Sandoval-Castro, C.A.; Tun-Garrido, J. Feeding behavior of sheep and goats in a deciduous tropical forest during the dry season: The same menu consumed differently. Small. Ruminant. Res. 2015, 133, 128–134. [Google Scholar] [CrossRef]
- Ventura-Cordero, J.; González-Pech, P.G.; Jaimez-Rodríguez, P.R.; Ortíz-Ocampo, G.I.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.J. Gastrointestinal nematode infection does not affect selection of tropical foliage by goats in a cafeteria trial. Trop. Anim. Health Prod. 2017, 49, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Ventura-Cordero, J.; González-Pech, P.G.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.J.; Tun-Garrido, J. Feed resource selection by Criollo goats browsing a tropical deciduous forest. Anim. Prod. Sci. 2019, 58, 2314–2320. [Google Scholar] [CrossRef]
- Ventura-Cordero, J.; González-Pech, P.G.; Jaimez-Rodríguez, P.R.; Ortiz-Ocampo, G.I.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.J. Feed resource selection of Criollo goats artificially infected with Haemonchus contortus: Nutritional wisdom and prophylactic self-medication. Animal 2018, 12, 1269–1276. [Google Scholar] [CrossRef]
- Torres-Fajardo, R.A.; Navarro-Alberto, J.A.; Ventura-Cordero, J.; González-Pech, P.G.; Sandoval-Castro, C.A.; Chan-Pérez, J.I.; Torres-Acosta, J.F.J. Intake and selection of goats grazing heterogeneous vegetation: Effect of gastrointestinal nematodes and condensed tannins. Rangel. Ecol. Manag. 2019, 72, 946–953. [Google Scholar] [CrossRef]
- Méndez-Ortiz, F.A.; Sandoval-Castro, C.A.; Ventura-Cordero, J.; Sarmiento-Franco, L.A.; Santos-Ricalde, R.H.; Torres-Acosta, J.F.J. Gymnopodium floribundum fodder as a model for the in vivo evaluation of nutraceutical value against Haemonchus contortus. Trop. Anim. Health. Prod. 2019, 51, 1501–1599. [Google Scholar] [CrossRef]
- Bullock, S. La fenología de plantas en Chamela. In Historia natural de Chamela, 1st ed.; Noguera, F.A., Vega Rivera, J.H., García Aldrete, A.N., Quesada Avendaño, M., Eds.; Universidad Nacional Autónoma de México: México City, Mexico, 2002; pp. 491–494. [Google Scholar]
- Camarena Gutiérrez, G. Señales en la interacción planta insecto. Rev. Chapingo. Ser. Cienc. For. Ambiente 2009, 15, 81–85. [Google Scholar]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental facts. Molecules 2018, 23, 762. [Google Scholar] [CrossRef]
- Nosko, P.; Embury, K. Induction and persistence of allelochemicals in the foliage of balsam fir seedlings following simulated browsing. Plant. Ecol. 2018, 219, 611–619. [Google Scholar] [CrossRef]
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen, 5th ed.; Instituto de Geografía-Universidad Nacional Autónoma de México: México City, Mexico, 2004; pp. 21–65. [Google Scholar]
- García, J.; Mizrahi, A.; Bautista, F. Manejo campesino de la selva baja y selección de especies arbóreas para barbechos mejorados en Hocabá, Yucatán. In Caracterización y Manejo de los suelos de la Península de Yucatán: Implicaciones Agropecuarias, Forestales y Ambientales, 1st ed.; Bautista, F., Palacio, G., Eds.; Universidad Autónoma de Campeche: Campeche, Mexico, 2005; pp. 195–208. [Google Scholar]
- Borges-Gómez, L.; Moo-Kauil, C.; Ruíz-Novelo, J.; Osalde-Balam, M.; González-Valencia, C.; Yam-Chimal, C.; Can-Puc, F. Suelos destinados a la producción de chile habanero en Yucatán: Características físicas y químicas predominantes. Agrociencia 2014, 48, 347–359. [Google Scholar]
- Valdez-Hernández, J.I.; Vaquera-Huerta, H.; García-Moya, E.; Romero-Manzanares, A.; Borja-De-La-Rosa, M.A.; Interián-Ku, V.M. Arquitectura y morfometría de dos especies arbóreas en una selva baja caducifolia del sur de Yucatán, México. Bol. Soc. Bot. México 2009, 85, 17–29. [Google Scholar]
- Comisión Nacional del Agua (CONAGUA). Normales climatológicas por estado: Yucatán, México. Available online: https://smn.conagua.gob.mx/es/informacion-climatologica-por-estado?estado=yuc.C (accessed on 20 June 2019).
- Makkar, H.P.S. Quantification of Tannins in Tree and Shrub Foliage. A Laboratory Manual, 1st ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 49–53. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 13th ed.; Association of Official Analytical Chemists International: Washington, DC, USA, 1980. [Google Scholar]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G. A critical evaluation of the vanillin reaction assay for tannin in sorghum grain. J. Agric. Food. Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Barros-Rodríguez, M.; Solorio-Sánchez, J.; Ku-Vera, A.; Ayala-Burgos, A.; Sandoval-Castro, C.; Solís-Pérez, G. Productive performance and urinary excretion of mimosine metabolites by hair sheep grazing in a silvopastoral system with high densities of Leucaena leucocephala. Trop. Anim. Health Prod. 2012, 44, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Theodorou, M.; Williams, B.; Dhanoa, M.; Mcallan, A.; France, J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed. Sci. Technol. 1994, 48, 185–197. [Google Scholar] [CrossRef]
- Agriculture and Food Research Council (AFRC). Energy and Protein Requirements of Ruminants: An Advisory Manual Prepared by the AFRC Technical Committee on Responses to Nutrients; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Minitab 16 Statistical software; Computer software Minitab ver.16.2.4. Minitab Inc.: State College, PA, USA, 2013. Available online: www.minitab.com (accessed on 3 November 2020).
- Balvanera, P.; Islas, A.; Aguirre, E.; Quijas, S. Las selvas secas. Ciencias 2000, 57, 18–24. [Google Scholar]
- Valladares, F.; Vilagrosa, A.; Peñuelas, J.; Ogaya, R.; Julio Camarero, J.; Corcuera, L.; Sisó, S.; Gil Pelegrín, E. Estrés hídrico: Ecofisiología y escalas de la sequía. In Ecofisiología del Bosque Mediterráneo en un Mundo Cambiante, 1st ed.; Valladares, F., Ed.; Ministerio del Medio ambiente, EGRAF, S.A.: Madrid, Spain, 2004; pp. 163–190. [Google Scholar]
- Yepes, A.; Silveira-Buckeridge, M. Respuesta de las plantas ante los factores ambientales del cambio climático global—Revisión. Colomb. For. 2011, 14, 213–232. [Google Scholar] [CrossRef][Green Version]
- Enríquez-Quiroz, J.F.; Meléndez-Nava, F.; Bolaños-Aguilar, E.D. Tecnología Para la Producción y Manejo de Forrajes Tropicales en México, 1st ed.; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP); Campo Experimenta Papaloapan: Veracruz, Mexico, 1999. [Google Scholar]
- Vanholme, R.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin engineering. Curr. Opin. Plant. Biol. 2008, 11, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, A.E.; Butler, L.G. Tannins and lignins. In Herbivores: Their Interactions with Secondary Plant Metabolites, 2nd ed.; Rosenthal, G.A., Berehaum, M.R., Eds.; Academic Press: San Diego, CA, USA, 1991; Volume 1, pp. 355–388. [Google Scholar]
- Minson, J.D. Composición Química y Valor Nutritivo de las Gramíneas Tropicales. In Gramíneas Tropicales; Skerman, P.J., Riveros, F., Eds.; FAO: Roma, Italia, 1992; pp. 181–189. [Google Scholar]
- Castañeda-Ramírez, G.S.; Rodríguez-Labastida, M.; Ortiz-Ocampo, G.I.; González-Pech, P.G.; Ventura-Cordero, J.; Borges-Argáez, R.; Torres-Acosta, J.F.J.; Sandoval-Castro, C.A.; Mathieu, C. An in vitro approach to evaluate the nutraceutical value of plant foliage against Haemonchus contortus. Parasitol. Res. 2018, 117, 33979–33991. [Google Scholar] [CrossRef]
- Coley, P.D. Herbivory and defensive characteristics of tree species in a lowland tropical forest. Ecol. Monogr. 1983, 53, 209–234. [Google Scholar] [CrossRef]
- del Val, E. Herbivoría. In Ecología y Evolución de las Interacciones Bióticas, 1st ed.; del Val, E., Boege, K., Eds.; Fondo de Cultura Económica: México City, México, 2012; pp. 43–75. [Google Scholar]
- Chakraborty, U.; Chakraborty, N. Impact of environmental factors on infestation of tea leaves by Helopeltis theivora, and associated changes in flavonoid flavor components and enzyme activities. Phytoparasitica 2005, 33, 88–96. [Google Scholar] [CrossRef]
- Gobbo-Neto, L.; Lopes, P. Plants Medicinals: Factores de Influência no conteúdo de metabólites secundarios. Quim. Nova 2007, 30, 374–381. [Google Scholar] [CrossRef]
- García-Rodríguez, Y.; Bravo-Monzón, A.; Martínez-Díaz, Y.; Torres-Gurrola, G.; Espinosa-García, F.J. Variación Fitoquímica defensiva en ecosistemas terrestres. In Temas Selectos en Ecología Química de Insectos, 1st ed.; Rojas, J.C., Malo, E., Eds.; El Colegio de la Frontera Sur: San Cristobal de Las Casas, Mexico, 2012; pp. 217–252. [Google Scholar]
- Sepúlveda-Jiménez, G.; Porta-Doucoing, H.; Rocha-Sosa, M. Participación de los metabolitos secundarios en la defensa de las plantas. Rev. Mex. Fitopatol. 2003, 21, 355–363. [Google Scholar]
- Ramakrishna, A.; Aswathanarrayana, G. Influence of abiotic stress signals on secondary metabolites in plants. Plant. Signal. Behav. 2011, 11, 1720–1731. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, B.; Ruusunen, V.; Laaksonen, O.; Tahvonen, R.; Hellsten, J.; Kallio, H. Compositional differences of phenolic compounds between black currant (Ribes nigrum L.) cultivars and their response to latitude weather conditions. Agric. Food. Chem. 2012, 60, 6581–6593. [Google Scholar] [CrossRef] [PubMed]
- Uleberg, E.; Rohloff, J.; Jaakola, L.; Trôst, K.; Junttila, O.; Häggman, H.; Martinussen, I. Effects of temperature and photoperiod on yield and chemical composition of northern and southern clones of bilberry (Vaccinium myrtillus L.). J. Agric. Food. Chem. 2012, 60, 10406–10414. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Harvest Time | FB | DM | CP | ADF | NDF | Lignin | TP | TT | CT | IVDMD | IVOMD | EM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mixed-age leaves | ||||||||||||
| March | 84.7 d | 65.6 a | 15.4 | 18.3 c | 35.3 d | 8.0 cd | 6.7 abc | 2.44 | 23.1 c | 36.8 b | 38.6 b | 5.7 b |
| June | 1071.0 a | 32.3 e | 14.6 | 23.1 b | 46.5 b | 10.2 c | 5.7 c | 1.58 | 22.7 c | 26.2 de | 27.0 c | 3.9 c |
| September | 903.0 ab | 41.46 d | 14.3 | 32.3 a | 51.8 a | 18.2 a | 6.3 bc | 1.26 | 41.3 a | 25.6 de | 28.0 c | 4.1 c |
| December | 545.7 bc | 52.96 c | 13.9 | 25.4 b | 40.2 c | 12.6 b | 4.6 d | 1.4 | 24.0 c | 22.9 e | 26.8 c | 3.9 c |
| 90-day-old leaves | ||||||||||||
| March | 151.0 cd | 59.23 b | 14.9 | 13.6 c | 33.5 d | 5.6 d | 7.9 a | 1.1 | 31.3 b | 43.1 a | 42.7 a | 6.3 a |
| June | 833.6 ab | 33.3 e | 14.4 | 24.7 b | 42.7 b | 11.5 bc | 4.6 d | 1.2 | 19.3 c | 29.1 cd | 29.4 c | 4.3 c |
| September | 318.0 cd | 36.46 d | 14.3 | 32.2 a | 53.1 a | 17.1 a | 6.7 abc | 1.4 | 44.3 a | 25.1 de | 25.9 c | 3.8 c |
| December | 127.7 cd | 58.9 b | 13.5 | 24.8 b | 47.9 b | 11.8 b | 7.4 ab | 2.8 | 43.7 a | 34.2 bc | 34.3 b | 5.1 b |
| SE | 90.17 | 1.13 | 0.68 | 1.76 | 1.77 | 1.23 | 0.30 | 0.39 | 1.15 | 1.19 | 1.08 | 0.15 |
| FB | DM | CP | ADF | NDF | Lignin | TP | TT | CT | |
|---|---|---|---|---|---|---|---|---|---|
| Mixed-age leaves | |||||||||
| SH | 0.806 ** | −0.879 ** | − | − | 0.829 ** | − | − | − | − |
| RH | − | − | − | 0.712 ** | 0.625 * | 0.787 ** | − | − | 0.965 ** |
| TMP | 0.822 ** | −0.874 ** | − | − | 0.885 ** | − | − | − | − |
| RF | 0.761 ** | −0.762 ** | − | 0.789 ** | 0.946 ** | 0.779 ** | − | −0.587 * | 0.805 ** |
| MT | − | − | − | − | − | − | 0.655 * | − | − |
| MH | 0.738 ** | −0.702 * | − | 0.864 ** | 0.863 ** | 0.838 ** | − | 0.753 ** | 0.707 * |
| 90-day-old leaves | |||||||||
| SH | 0.768 ** | −0.996 ** | − | 0.643 * | − | 0.613 * | −0.780 ** | − | − |
| RH | − | − | − | 0.688 * | 0.816 ** | 0.690 * | − | − | 0.872 ** |
| TMP | 0.650 * | −0.992 ** | − | 0.810 ** | 0.663 * | 0.771 ** | −0.717 ** | − | − |
| RF | − | −0.857 ** | − | 0.841 ** | 0.727 ** | 0.836 ** | − | − | − |
| MT | − | − | − | − | − | − | − | −0.705 * | −0.668 * |
| MH | − | −0.704 * | − | 0.931 ** | 0.881 ** | 0.908 ** | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortíz-Ocampo, G.I.; Sandoval-Castro, C.A.; González-Pech, P.G.; Mancilla-Montelongo, G.; Ventura-Cordero, J.; Castañeda-Ramírez, G.S.; Tun-Garrido, J.; Torres-Acosta, J.F.d.J. Month of Harvest and Leaf Age Impact the Bromatological Composition and Polyphenol Content of Gymnopodium floribundum Rolfe Leaves. Agriculture 2022, 12, 1110. https://doi.org/10.3390/agriculture12081110
Ortíz-Ocampo GI, Sandoval-Castro CA, González-Pech PG, Mancilla-Montelongo G, Ventura-Cordero J, Castañeda-Ramírez GS, Tun-Garrido J, Torres-Acosta JFdJ. Month of Harvest and Leaf Age Impact the Bromatological Composition and Polyphenol Content of Gymnopodium floribundum Rolfe Leaves. Agriculture. 2022; 12(8):1110. https://doi.org/10.3390/agriculture12081110
Chicago/Turabian StyleOrtíz-Ocampo, Guadalupe Isabel, Carlos Alfredo Sandoval-Castro, Pedro Geraldo González-Pech, Gabriela Mancilla-Montelongo, Javier Ventura-Cordero, Gloria Sarahi Castañeda-Ramírez, Juan Tun-Garrido, and Juan Felipe de Jesús Torres-Acosta. 2022. "Month of Harvest and Leaf Age Impact the Bromatological Composition and Polyphenol Content of Gymnopodium floribundum Rolfe Leaves" Agriculture 12, no. 8: 1110. https://doi.org/10.3390/agriculture12081110
APA StyleOrtíz-Ocampo, G. I., Sandoval-Castro, C. A., González-Pech, P. G., Mancilla-Montelongo, G., Ventura-Cordero, J., Castañeda-Ramírez, G. S., Tun-Garrido, J., & Torres-Acosta, J. F. d. J. (2022). Month of Harvest and Leaf Age Impact the Bromatological Composition and Polyphenol Content of Gymnopodium floribundum Rolfe Leaves. Agriculture, 12(8), 1110. https://doi.org/10.3390/agriculture12081110