
The Combination of Plant Extracts and Probiotics Improved Jejunal Barrier and Absorption Capacity of Weaned Piglets

Abstract
:1. Introduction
2. Materials and Methods
2.1. IVE and Probiotics Supplemented Diet
2.2. Preparation and Determination of IVE
2.3. Animals, Experimental Design, and Management
2.4. Sample Collections for Analysis of Nutrient Availability
2.5. Sampling for Determination of Jejunal Barrier and Absorption Capacity
2.6. Quantitative Real-Time PCR
2.7. Histological and Immunohistochemical Analysis
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. Nutrients Digestibility
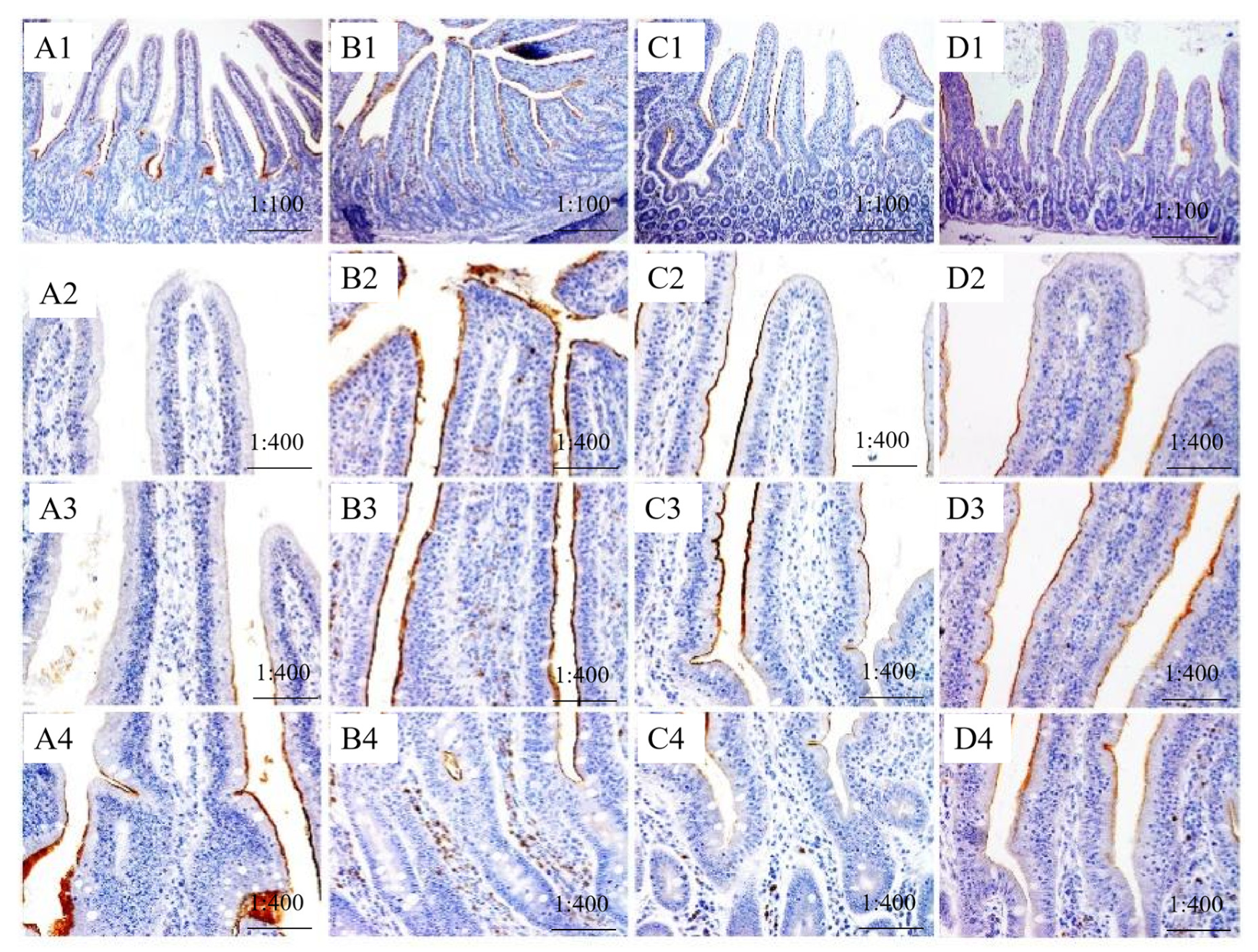
3.2. Localization of SGLT1 and ZO-1
3.3. The Expression of mRNA and Protein
4. Discussion
4.1. Nutrients Digestibility
4.2. Jejunum Histomorphology
4.3. Localization and Expression
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.; Zhou, H.; Guan, M.; Zhou, X.; Zhang, L. Lactobacillus casei YRL577 combined with plant extracts reduce markers of non-alcoholic fatty liver disease in mice. Br. J. Nutr. 2020, 125, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.C.; Fu, J.; Zhu, L.Y.; Li, Z.; Jin, M.L.; Wang, Y.Z. Overall assessment of antibiotic substitutes for pigs: A set of meta-analyses. J. Anim. Sci. Biotechnol. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.L. Effect of probiotics on intestinal flora and its application in livestock and poultry production. Hunan Feed. 2019, 1, 23–26. [Google Scholar]
- Liao, S.F.; Nyachoti, M. Using probiotics to improve swine gut health and nutrient utilization. Anim. Nutr. 2017, 3, 331–343. [Google Scholar] [CrossRef]
- Huang, W.; Chang, J.; Wang, P.; Liu, C.Q.; Yin, Q.Q.; Song, A.D.; Gao, T.Z.; Dang, X.W.; Lu, F.S. Effect of Compound Probiotics and Mycotoxin Degradation Enzymes on Alleviating Cytotoxicity of Swine Jejunal Epithelial Cells Induced by Aflatoxin B1 and Zearalenone. Toxins 2019, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Berlec, A. Novel techniques and findings in the study of plant microbiota: Search for plant probiotics. Plant Sci. Int. J. Exp. Plant Biol. 2012, 193, 96–102. [Google Scholar] [CrossRef]
- Belorkar, S.A. Biostimulants: Promising Probiotics for Plant Health; Woodhead Publishing: Shaston, UK, 2021. [Google Scholar]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef]
- Lata, P. Probiotics for Human Health; Springer: Singapore, 2021. [Google Scholar]
- O’Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2017, 2, 1–6. [Google Scholar] [CrossRef]
- Sánchez, B.; Delgado, S.; Blanco-míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2016, 61, 1600240. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.; Zhao, D.H.; Jiang, M. probiotics regulate the intestinal epithelial barrier in vivo and in vitro via the p38 and ERK signaling pathways. Int. J. Mol. Med. 2012, 29, 202. [Google Scholar]
- Liu, H.; Zhang, J.; Zhang, S.; Yang, F.; Thacker, P.A.; Zhang, G.; Qiao, S.; Ma, X. Oral administration of Lactobacillus fermentum I5007 favors intestinal development and alters the intestinal microbiota in formula-fed piglets. J. Agric. Food Chem. 2014, 62, 860–866. [Google Scholar] [CrossRef]
- Yang, J.F.; Yang, C.H.; Chang, H.W. Chemical composition and antibacterial activities of Illicium verum against antibiotic-resistant pathogens. J. Med. Food 2010, 13, 1254–1262. [Google Scholar] [CrossRef]
- Wang, G.W.; Hu, W.T.; Huang, B.K.; Qin, L.P. Illicium verum: A review on its botany, traditional use, chemistry and pharmacology. J. Ethnopharmacol. 2011, 136, 10–20. [Google Scholar] [CrossRef]
- Ding, X.; Yang, C.; Yang, Z.; Ren, X.J.; Wang, P. Effects of star anise (Illicium verum Hook.f) oil on the nuclear factor E2–related factor 2 signaling pathway of chickens during subclinical Escherichia coli challenge. Poult. Sci. 2020, 99, 3092–3101. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, G.; Chen, K.; Chen, X.; Kan, J. Assessment of fresh star anise (Illicium verum Hook.f.) drying methods for influencing drying characteristics, color, flavor, volatile oil and shikimic acid. Food Chem. 2020, 342, 128359. [Google Scholar] [CrossRef]
- Luís, Â.; Sousa, S.; Wackerlig, J.; Dobusch, D.; Duarte, A.P.; Pereira, L.; Domingues, F. Star anise (Illicium verum Hook. f.) essential oil: Antioxidant properties and antibacterial activity against Acinetobacter baumannii. Flavour Fragr. J. 2019, 34, 260–270. [Google Scholar] [CrossRef]
- Wei, H.K.; Wang, J.; Cheng, C.S.; Jin, L.Z.; Peng, J. Application of plant essential oils in pig diets. Feed Addit. 2020, 13, 227–237. [Google Scholar]
- Yi, Y.; You, X.; Long, Y.; Huang, Y. Efficacy of different plant extracts in the prevention of radiation dermatitis in radiotherapy patients with nasopharyngeal carcinoma: A protocol for a Bayesian network meta-analysis. Medicine 2020, 99, 23523. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.J.; Yang, Z.B.; Yang, W.R.; Huang, L.B.; Xu, C.; Liu, M.; Ge, J.S.; Wang, Y.X.; Jiang, S.Z. Illicium verum extracts and probiotics with added glucose oxidase promote antioxidant capacity through upregulating hepatic and jejunal Nrf2/Keap1 of weaned piglets. J. Anim. Sci. 2020, 98, skaa077. [Google Scholar] [CrossRef]
- Holkem, A.T.; Silva, M.P.; Favaro-Trindade, C.S. Probiotics and plant extracts: A promising synergy and delivery systems. Crit. Rev. Food Sci. Nutr. 2022, 4, 1–19. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine, 10th ed.; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- AOAC. Association of Official Analytical Chemists, Gaithersburg, 8th ed.; AOAC: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Yang, L.J.; Min, Z.; Libo, H.; Weiren, Y.; Zaibin, Y.; Shuzhen, J.; Jinshan, G. Zearalenone promotes follicle growth through modulation of Wnt-1/β-catenin signaling pathway and expression of estrogen receptor genes in ovaries of post-weaning piglets. J. Agric. Food Chem. 2018, 66, 7899–7906. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, K.X.; Liang, P.X.; Yang, K.Z.; Zou, L.J.; Yang, Y.F. Effects of compound probiotics on growth performance and apparent nutrient digestibility of piglets. China Feed. 2019, 21, 57–59. [Google Scholar]
- Zhou, S.; Huang, H.S.; Yang, W.R. Effects of enzyme and probiotics on nutrient utilization and growth performance of Weaned Piglets. Feed. Ind. 2011, 32, 40–43. [Google Scholar]
- Chen, P. Study on the Effect of Illicium Verum and Eucommia Leaf Extract on the Antioxidant Abilities and Immunity in Weaned Piglets; Shandong Agricultural University: Tai’an, China, 2017. [Google Scholar]
- Ding, X.; Ren, X.J.; Yang, Z.B.; Li, G.; Zhang, X.Q. Effects of star anise, essential oil and residues on amino acid utilization in Broilers. Feed. Ind. 2018, 39, 48–51. [Google Scholar]
- Yu, C.Y. Study on the Effects of Star Anise oil (Illicium verum) on the Laying Performance, Nutrient Utilization and Antioxidant Status in Laying Hens; Shandong Agricultural University: Tai’an, China, 2019. [Google Scholar]
- Curry, S.M.; Schwartz, K.J.; Yoon, K.J.; Gabler, K.J.; Burrough, E.R. Effects of porcine epidemic diarrhea virus infection on nursery pig intestinal function and barrier integrity. Vet. Microbiol. 2017, 211, 58–66. [Google Scholar] [CrossRef]
- Shen, X.X. Effects of Sugar Cane Extract on Intestinal Histological Moprhology of Growing Pigs and Heat Stressed Rats; Nanjing Agricultural University: Tai’an, China, 2016. [Google Scholar]
- Cairo, P.L.G.; Gois, F.D.; Sbardella, M.; Silveira, H.; Oliveira, R.M.; Allaman, I.B.; Cantarelli, V.S.; Costa, L.B. Effects of dietary supplementation of red pepper (Schinus terebinthifolius Raddi) essential oil on performance, small intestinal morphology and microbial counts of weanling pigs. J. Sci. Food Agric. 2018, 98, 541–548. [Google Scholar] [CrossRef]
- Kim, J.S.; Hosseindoust, A.; Lee, S.H.; Choi, Y.H.; Chae, B.J. Bacteriophage cocktail and multi-strain probiotics in the feed for weanling pigs: Effects on intestine morphology and targeted intestinal coliforms and Clostridium. Animal 2017, 1, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Mishra, D.K.; Verma, A.K.; Agarwal, N.; Singh, P. Effect of Probiotics on Blood Biochemical Profile, Immunity and Small Intestine Morphology in Growing Finishing Pigs. Agric. Res. 2016, 5, 1–6. [Google Scholar] [CrossRef]
- Dowarah, R.; Verma, A.K.; Agarwal, N.; Patel, B.H.M.; Singh, P. Effect of swine based probiotic on performance, diarrhoea scores, intestinal microbiota and gut health of grower-finisher crossbred pigs. Livest. Sci. 2017, 195, 74–79. [Google Scholar] [CrossRef]
- Yoshida, A.; Takata, K.; Kasahara, T.; Aoyagi, T.; Saito, S.; Hirano, H. Immunohistochemical localization of Na(+)-dependent glucose transporter in the rat digestive tract. Histochem. J. 1995, 27, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.W.; Al-Rammahi, M.A.; Arora, D.K.; Batchelor, D.J.; Coulter, E.A.; Ionescu, C.; Bravo, D.; Shirazi-Beechey, S.P. Expression of Na+/glucose co-transporter 1 (SGLT1) is enhanced by supplementation of the diet of weaning piglets with artificial sweeteners. Br. J. Nutr. 2010, 104, 637–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, H.; Melo, A.D.B.; Bortoluzzi, C.; Costa, L.B.; Rostagno, M.H.; Schinckel, A.P.; Garbossa, C.A.P.; Cantarelli, V.D.S. Feed additives can differentially modulate NF-κB (Rel A/p65), IGF-1, GLUT2, and SGLT1 gene expression in porcine jejunal explants. Rev. Bras. Zootec. 2018, 47, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic Lactobacillus plantarum promotes intestinal barrier function by strengthening the epithelium and modulating gut microbiota. Front. Microbiol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Jahromi, M.F.; Altaher, Y.W.; Shokryazdan, P.; Ebrahimi, R.; Ebrahimi, M.; Idrus, Z.; Tufarelli, V.; Liang, J.B. Dietary supplementation of a mixture of Lactobacillusstrains enhances performance of broiler chickens raised under heat stress conditions. Int. J. Biometeorol. 2016, 60, 1099–1110. [Google Scholar] [CrossRef]
- Inoue, K.; Oyamada, M.; Mitsufuji, S.; Okanoue, T.; Takamatsu, T. Different changes in the expression of multiple kinds of tight-junction proteins during ischemia-reperfusion injury of the rat ileum. Acta Histochem. Cytochem. 2006, 39, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Wei, H.K.; Xiang, Q.H.; Wang, J.; Zhou, Y.F.; Peng, J. Protective effect of quercetin on pig intestinal integrity after transport stress is associated with regulation oxidative status and inflammation. J. Vet. Med. Sci. 2016, 78, 1487–1494. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.F.; Zhou, L.L.; Hu, H.C.; Zhou, H.L. Effects of extract of Pseudostellaria on histological morphology and permeability of ileal mucosa in Weaned Piglets. Chin. J. Anim. Sci. 2018, 54, 110–114. [Google Scholar]
- Roselli, M.; Finamore, A.; Britti, M.S.; Konstantinov, S.R.; Smidt, H.; de Vos, W.M.; Mengheri, E. The novel porcine Lactobacillus sobrius strain protects intestinal cells from enterotoxigenic Escherichia coli K88 infection and prevents membrane barrier damage. J. Nutr. 2007, 137, 2709–2716. [Google Scholar] [CrossRef] [Green Version]
- Bhat, M.I.; Sowmya, K.; Kapila, S.; Kapila, R. Potential Probiotic Lactobacillus rhamnosus (MTCC-5897) Inhibits Escherichia coli Impaired Intestinal Barrier Function by modulating the Host Tight Junction Gene Response. Probiotics Antimicrob. Proteins 2019, 12, 1149–1160. [Google Scholar] [CrossRef]
- Du, W.; Xu, H.; Mei, X.; Cao, X.; Gong, L.; Wu, Y.; Li, Y.; Yu, D.; Liu, S.; Wang, Y.; et al. Probiotic Bacillus enhance the intestinal epithelial cell barrier and immune function of piglets. Benef. Microbes 2018, 9, 743–754. [Google Scholar] [CrossRef]
- Xu, B.L.; Wang, S.; Hao, Z.M.; Wei, Z.Y. Research progress of basic amino acid transporters in animal cells. Feed. Ind. 2010, 31, 12–16. [Google Scholar]
- Zhi, A.M.; Zuo, J.J.; Zhou, X.Y.; Zou, S.K.; Huang, Z.Y.; Wang, X.L.; Ye, H.; Dai, F.W.; Feng, D.Y. The Influence of Different Lysine Concentration on the Cationic Amino Acid Transporter mRNA Expression of Porcine IEC. Chin. Agric. Sci. Bull. 2010, 26, 6–11. [Google Scholar]
- Xu, X.; Chen, X.; Chen, D.; Yu, B.; Yin, J.; Huang, Z. Effects of dietary apple polyphenol supplementation on carcass traits, meat quality, muscle amino acid and fatty acid composition in finishing pigs. Food Funct. 2019, 10, 7426–7434. [Google Scholar] [CrossRef]
- Chen, L.X.; Hu, T.; Xu, X. Effects of probiotics on the intestinal flora of piglets. China Anim. Husb. Vet. Med. 2013, 40, 172–174. [Google Scholar]
- Wang, S.; Ahmadi, S.; Nagpal, R.; Jian, S.; Mishra, S.P.; Kavanagh, K.; Zhu, X.W.; Wang, Z.; Mcclain, D.; Kritchevsky, S.B.; et al. Lipoteichoic acid from the cell wall of a heat killed Lactobacillus paracasei D3-5 ameliorates aging-related leaky gut, inflammation and improves physical and cognitive functions: From C. elegans to mice. Geroscience 2020, 42, 333–352. [Google Scholar] [CrossRef]
- Wegh, C.A.M.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and their potential applications in early life nutrition and beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [Green Version]
- Klemashevich, C.; Wu, C.; Howsmon, D.; Alaniz, R.C.; Lee, K.; Jayaraman, A. Rational identification of diet-derived postbiotics for improving intestinal microbiota function. Curr. Opin. Biotechnol. 2014, 26, 85–90. [Google Scholar] [CrossRef]
- Wilkins, T.; Sequoia, J. Probiotics for gastrointestinal conditions: A summary of the evidence. Am. Fam. Physician 2017, 96, 170–178. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content (%) | Nutrients 2 | |
|---|---|---|---|
| Corn | 64.50 | Digestible Energy, MJ/kg | 13.81 |
| Whey powder | 5.00 | Crude Protein (%) | 19.82 |
| Soybean meal | 23.00 | Calcium (%) | 0.70 |
| Fish meal | 5.00 | Total Phosphorus (%) | 0.64 |
| L-Lysine HCl | 0.20 | Lysine (%) | 1.22 |
| CaHPO4 | 0.70 | Sulfur Amino Acid (%) | 0.65 |
| Pulverized Limestone | 0.30 | Threonine (%) | 0.75 |
| NaCl | 0.30 | Tryptophan (%) | 0.22 |
| Premix 1 | 1.00 | ||
| Total | 100.0 | ||
| Genes | Accession No. | Primer Sequence (5′ to 3′) | Product Size bp |
|---|---|---|---|
| GADPH | NM_001206359.1 | F: ATGGTGAAGGTCGGAGTGAA | 154 |
| R: CGTGGGTGGAATCATACTGG | |||
| SGLT1 | M34044 | F: ATGTACCTGTCTGTCCTGTCG | 381 |
| R: GTGTTGGAGATGGTCTTGGAG | |||
| CAT1 | NM_001012613 | F: CATCAAAAACTGGCAGCTCA | 185 |
| R: TGGTAGCGATGCAGTCAAAG | |||
| Occludin | U79554 | F: TATGAGACAGACTACACAACTGGCGGCGAGTCC | 363 |
| R: ATCATAGTCTCCAACCATCTTCTTGATGTG | |||
| ZO-1 | MX003353439.2 | F: CAGCCCGAGGCGTGTTTA | 150 |
| R: AAGGTGGGAGGATGCTGTTG |
| Items | Apparent Digestibility | BV | NPU | ||||
|---|---|---|---|---|---|---|---|
| DM | OM | EE | CP | ||||
| IVE− | PGO− | 80.46 | 83.75 | 87.97 c | 76.30 c | 88.02 | 67.16 b |
| PGO+ | 80.80 | 84.31 | 88.90 b | 77.92 a | 88.39 | 68.87 a | |
| IVE+ | PGO− | 80.55 | 83.84 | 89.22 ab | 77.49 b | 88.87 | 68.87 a |
| PGO+ | 80.87 | 84.33 | 89.69 a | 77.48 b | 89.00 | 68.96 a | |
| SEM | 0.201 | 0.309 | 0.180 | 0.397 | 0.527 | 0.546 | |
| Pooled | IVE− | 80.63 | 84.03 | 88.44 b | 77.11 | 88.21 | 68.02 b |
| IVE+ | 80.71 | 84.08 | 89.45 a | 77.49 | 88.94 | 68.91 a | |
| PGO− | 80.51 | 83.79 | 88.59 b | 76.90 b | 88.45 | 68.01 b | |
| PGO+ | 80.83 | 84.32 | 89.30 a | 77.70 a | 88.70 | 68.92 a | |
| p-Value | IVE+ | 0.949 | 0.901 | 0.002 | 0.127 | 0.643 | 0.048 |
| PGO+ | 0.519 | 0.227 | 0.044 | 0.043 | 0.327 | 0.026 | |
| PGO × IVE | 0.876 | 0.929 | 0.452 | 0.046 | 0.316 | 0.141 | |
| Items | Lysine | Methionine | Tryptophan | Arginine | Histidine | Leucine | Isoleucine | Phenylalanine | Threonine | Valine | TEAA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IVE− | PGO− | 83.94 c | 84.78 | 78.41 c | 88.24 | 86.53 | 79.55 | 77.34 | 78.52 | 78.25 | 77.97 | 81.65 |
| PGO+ | 85.15 b | 83.84 | 81.95 a | 88.45 | 86.15 | 79.84 | 75.92 | 78.48 | 77.52 | 76.55 | 81.21 | |
| IVE+ | PGO− | 85.21 b | 84.24 | 80.92 b | 88.31 | 86.11 | 79.45 | 77.75 | 79.15 | 77.43 | 76.58 | 81.63 |
| PGO+ | 85.96 a | 85.72 | 81.07 b | 88.78 | 86.26 | 81.21 | 78.01 | 78.77 | 78.74 | 76.93 | 81.99 | |
| SEM | 0.23 | 0.41 | 0.36 | 0.25 | 0.33 | 0.46 | 0.47 | 0.40 | 0.43 | 0.44 | 0.24 | |
| Pooled | IVE− | 84.55 b | 84.31 | 80.18 | 88.35 | 86.34 | 79.70 | 76.63 | 78.50 | 77.89 | 77.26 | 81.43 |
| IVE+ | 85.59 a | 84.98 | 81.00 | 88.55 | 86.19 | 80.33 | 77.88 | 78.96 | 78.09 | 76.76 | 81.81 | |
| PGO− | 84.58 b | 84.51 | 79.67 b | 88.28 | 86.32 | 79.50 | 77.55 | 78.84 | 77.84 | 77.28 | 81.64 | |
| PGO+ | 85.56 a | 84.78 | 81.51 a | 88.62 | 86.21 | 80.53 | 76.97 | 78.63 | 78.13 | 76.74 | 81.60 | |
| p-Value | IVE+ | 0.013 | 0.410 | 0.191 | 0.660 | 0.814 | 0.488 | 0.079 | 0.533 | 0.808 | 0.600 | 0.523 |
| PGO+ | 0.017 | 0.829 | 0.009 | 0.614 | 0.866 | 0.269 | 0.418 | 0.756 | 0.681 | 0.594 | 0.942 | |
| PGO × IVE | 0.046 | 0.111 | 0.021 | 0.857 | 0.693 | 0.425 | 0.256 | 0.832 | 0.260 | 0.319 | 0.549 | |
| Items | Villus Length, μm | Crypt Depth, μm | Villus Length/Crypt Depth | |
|---|---|---|---|---|
| IVE− | PGO− | 461.46 b | 390.17 a | 1.18 b |
| PGO+ | 596.56 a | 333.80 a | 1.78 a | |
| IVE+ | PGO− | 552.86 ab | 300.32 b | 1.84 a |
| PGO+ | 551.61 ab | 324.91 a | 1.69 ab | |
| SEM | 14.461 | 12.312 | 0.093 | |
| Pooled | IVE− | 529.02 | 361.99 | 1.48 |
| IVE+ | 552.24 | 312.62 | 1.77 | |
| PGO− | 507.17 | 345.25 | 1.51 | |
| PGO+ | 574.09 | 329.36 | 1.74 | |
| p-Value | IVE+ | 0.360 | 0.018 | 0.049 |
| PGO+ | 0.034 | 0.896 | 0.033 | |
| PGO × IVE | 0.663 | 0.058 | 0.010 | |
| Items | Occludin | ZO-1 | SGLT1 | CAT1 | |
|---|---|---|---|---|---|
| IVE− | PGO− | 1.00 b | 1.00 c | 1.00 b | 1.00 b |
| PGO+ | 1.48 a | 2.30 a | 1.36 a | 1.83 a | |
| IVE+ | PGO− | 1.28 ab | 1.57 b | 1.41 a | 1.24 ab |
| PGO+ | 1.48 a | 1.92 a | 1.35 a | 1.63 ab | |
| SEM | 0.201 | 0.288 | 0.177 | 0.314 | |
| Pooled | IVE− | 1.24 | 1.65 | 1.18 | 1.42 |
| IVE+ | 1.28 | 1.75 | 1.38 | 1.44 | |
| PGO− | 1.04 | 1.29 | 1.21 | 1.12 | |
| PGO+ | 1.48 | 2.11 | 1.36 | 1.73 | |
| p-value | IVE+ | 0.704 | 0.071 | 0.032 | 0.165 |
| PGO+ | 0.010 | <0.001 | 0.002 | <0.001 | |
| PGO × IVE | 0.266 | 0.04 | 0.013 | 0.093 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Ma, X.; Yang, C.; Jiang, S.; Yang, W.; Jiang, S. The Combination of Plant Extracts and Probiotics Improved Jejunal Barrier and Absorption Capacity of Weaned Piglets. Agriculture 2022, 12, 912. https://doi.org/10.3390/agriculture12070912
Yang L, Ma X, Yang C, Jiang S, Yang W, Jiang S. The Combination of Plant Extracts and Probiotics Improved Jejunal Barrier and Absorption Capacity of Weaned Piglets. Agriculture. 2022; 12(7):912. https://doi.org/10.3390/agriculture12070912
Chicago/Turabian StyleYang, Lijie, Xiangming Ma, Chongwu Yang, Shan Jiang, Weiren Yang, and Shuzhen Jiang. 2022. "The Combination of Plant Extracts and Probiotics Improved Jejunal Barrier and Absorption Capacity of Weaned Piglets" Agriculture 12, no. 7: 912. https://doi.org/10.3390/agriculture12070912
APA StyleYang, L., Ma, X., Yang, C., Jiang, S., Yang, W., & Jiang, S. (2022). The Combination of Plant Extracts and Probiotics Improved Jejunal Barrier and Absorption Capacity of Weaned Piglets. Agriculture, 12(7), 912. https://doi.org/10.3390/agriculture12070912