
Novel 3-D Spacer Textiles to Protect Crops from Insect Infestation and That Enhance Plant Growth

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
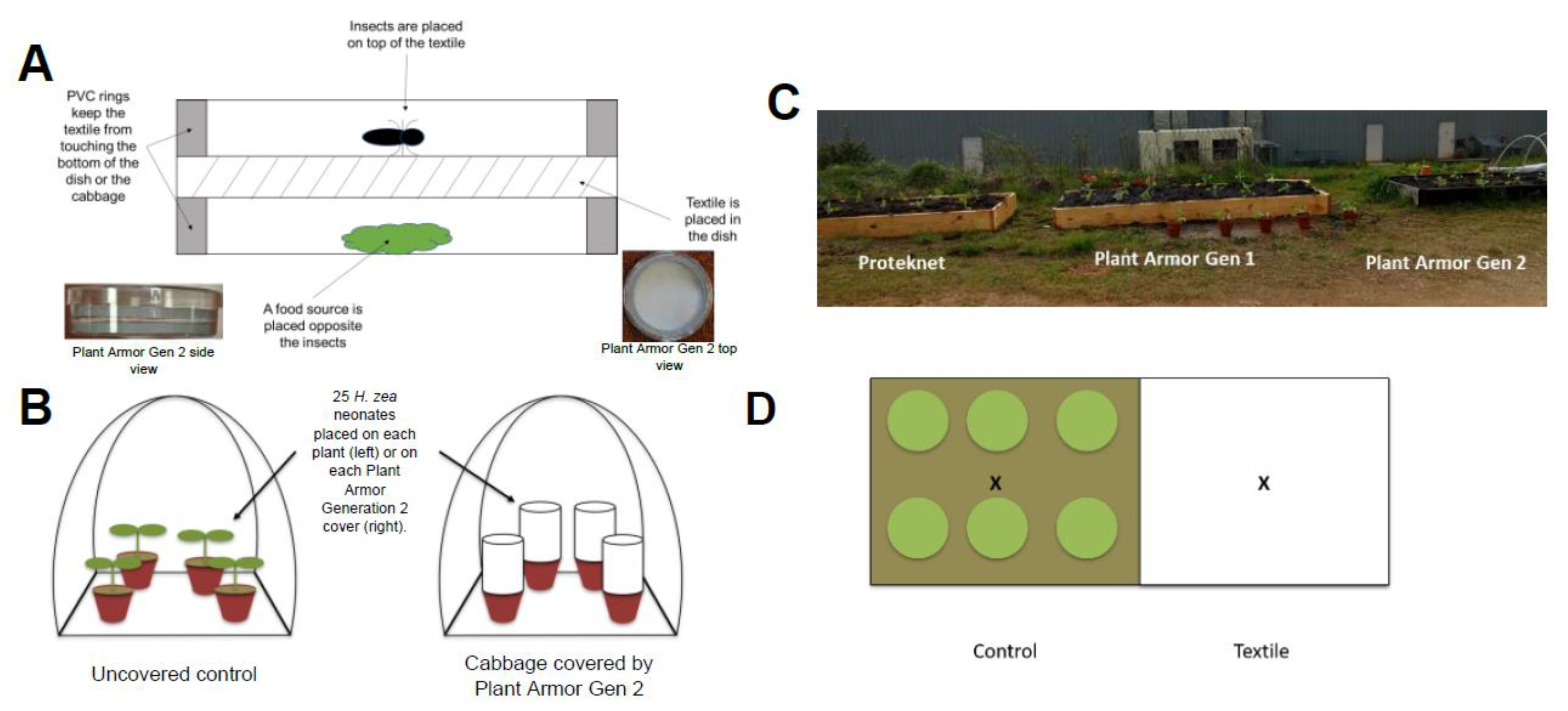
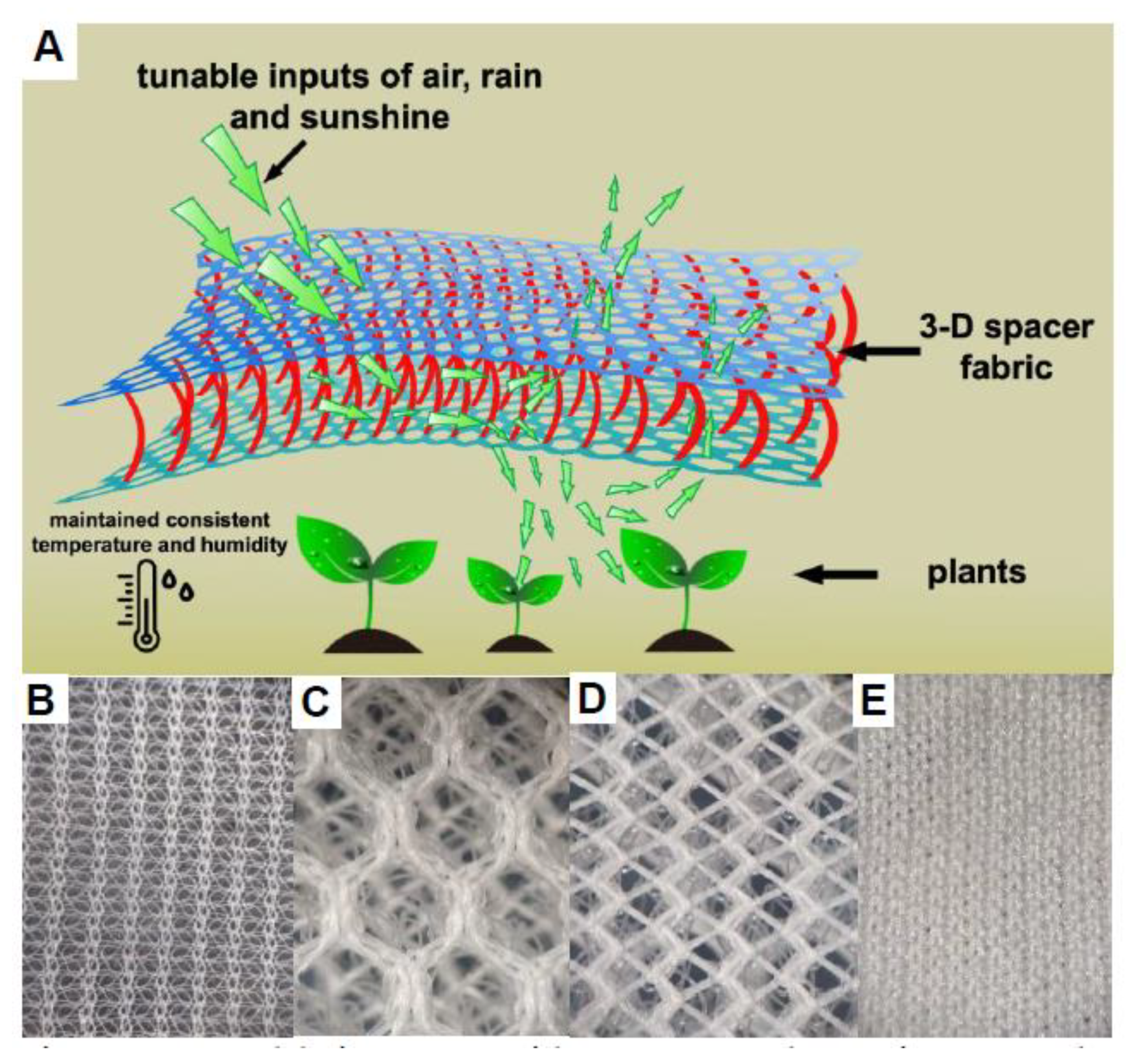
2.1. Textiles
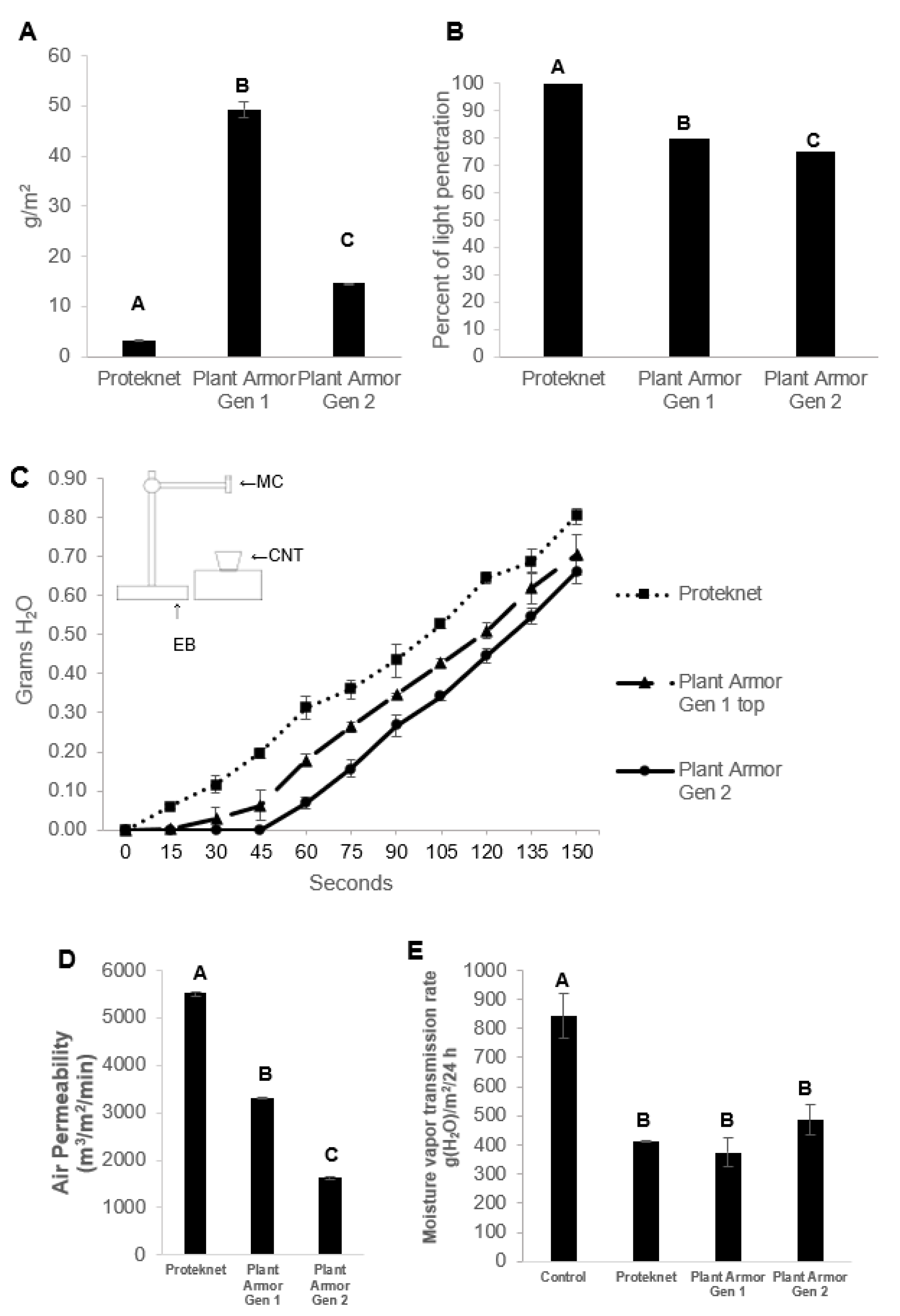
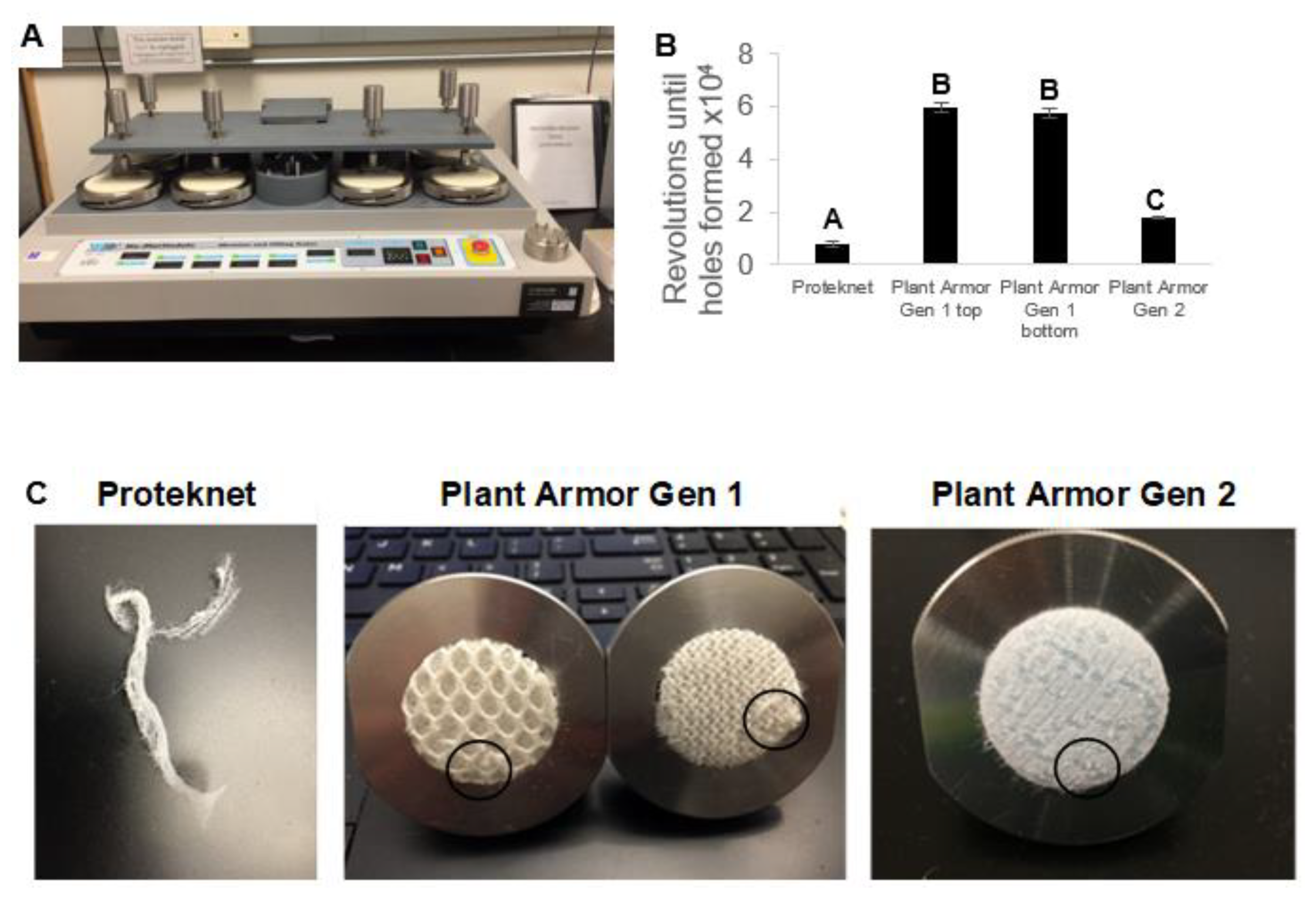
2.2. Textile Metrics
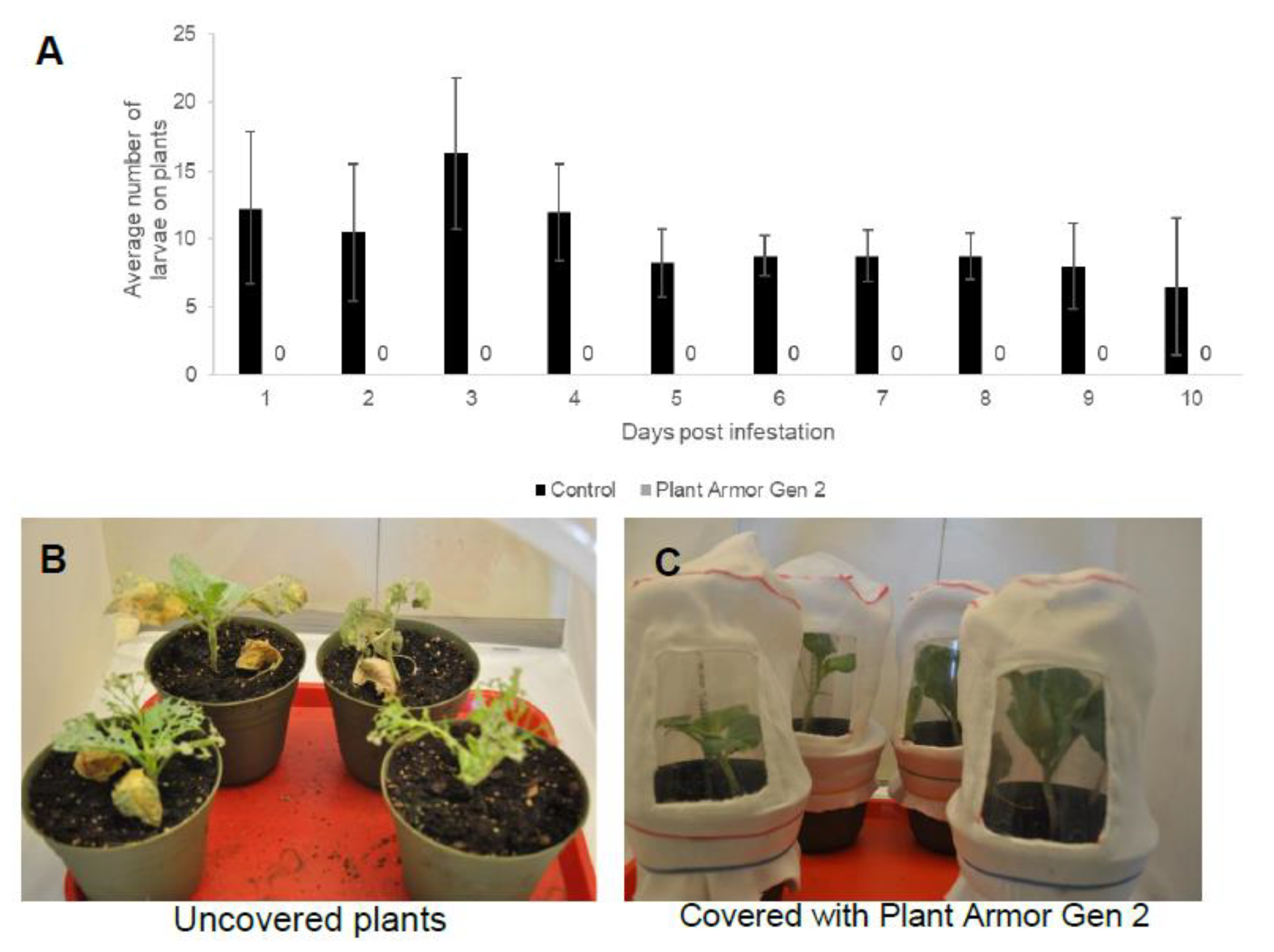
2.3. Insects
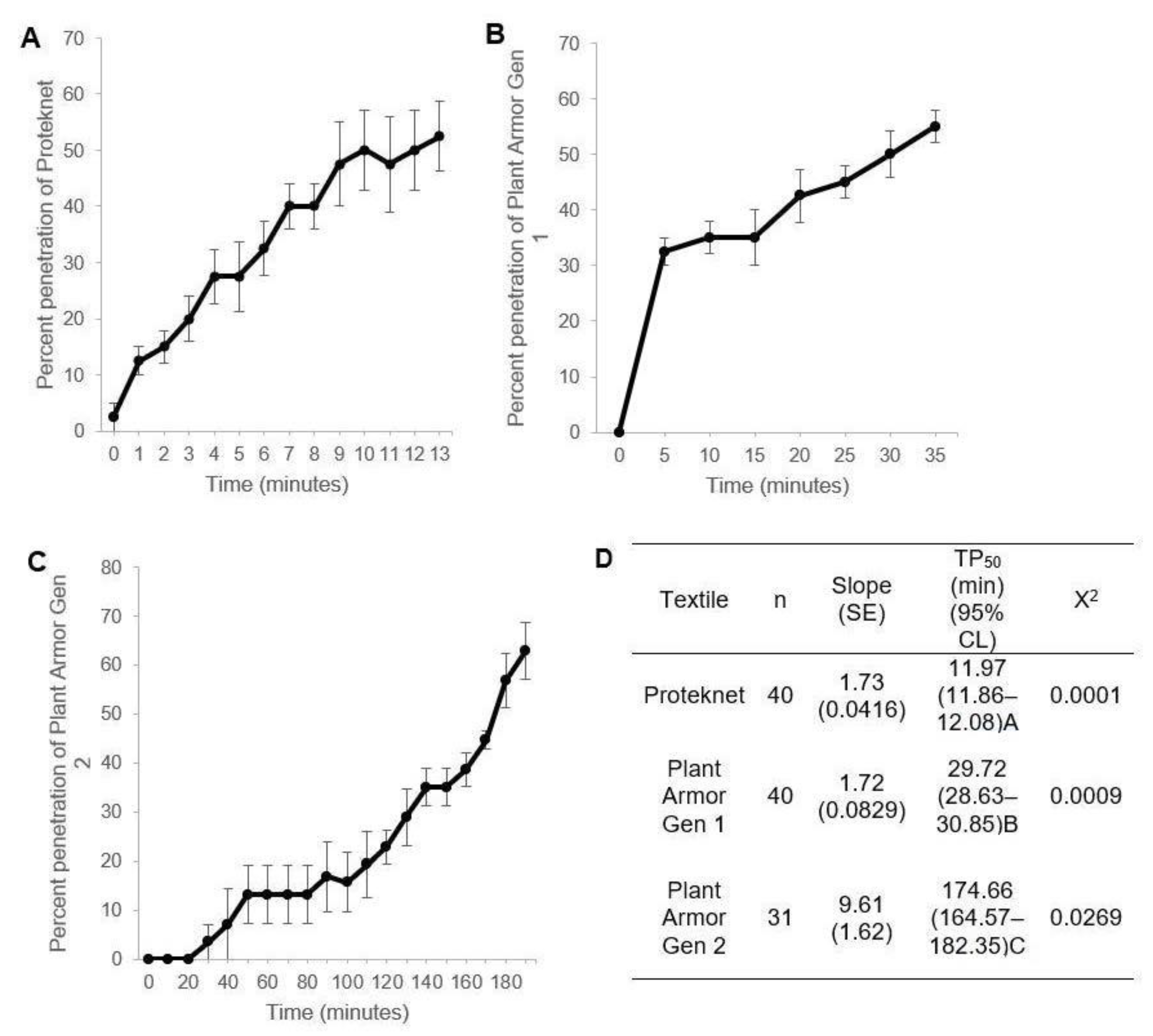
2.4. Petri Dish Penetration Assay
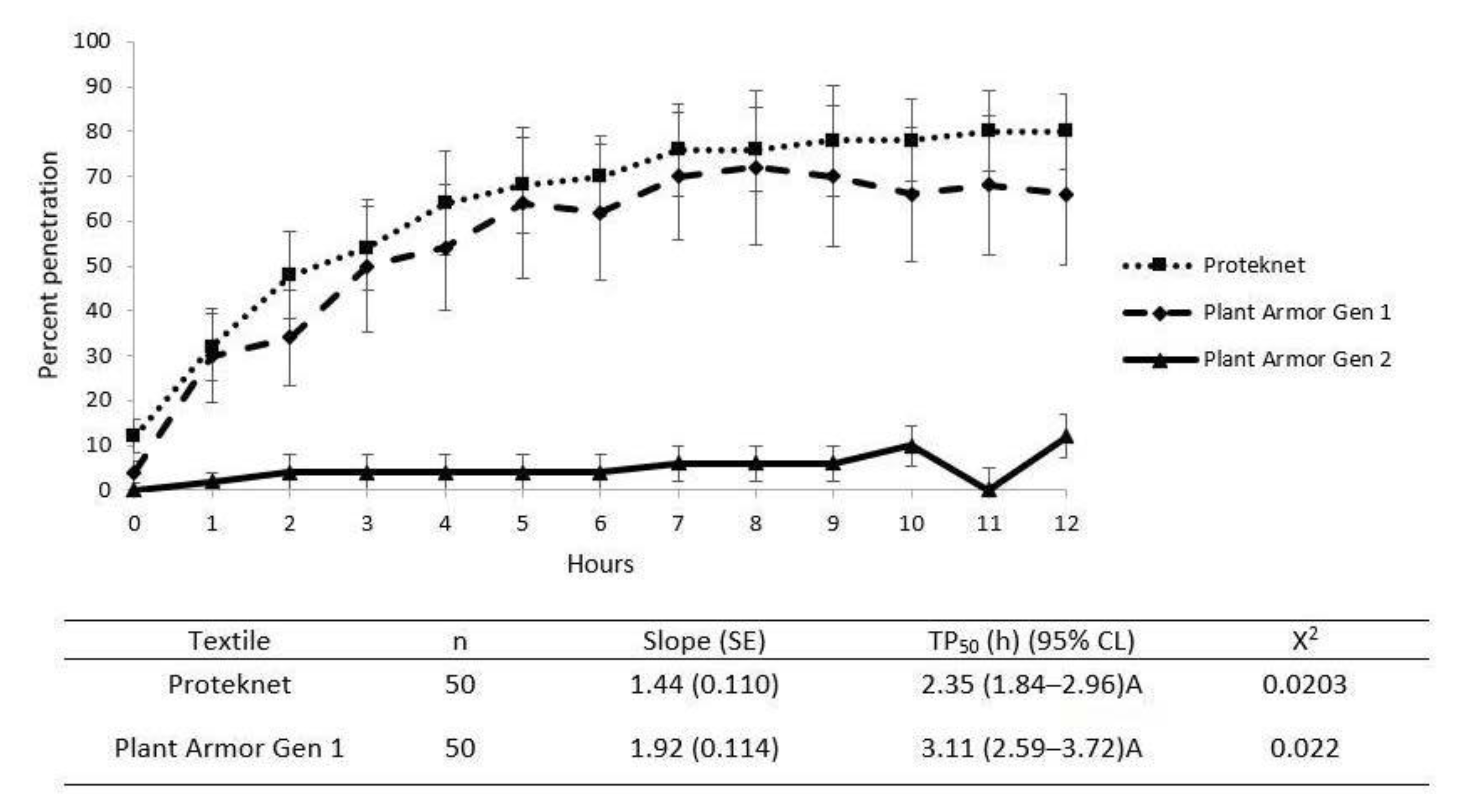
2.5. Cage Penetration Assay
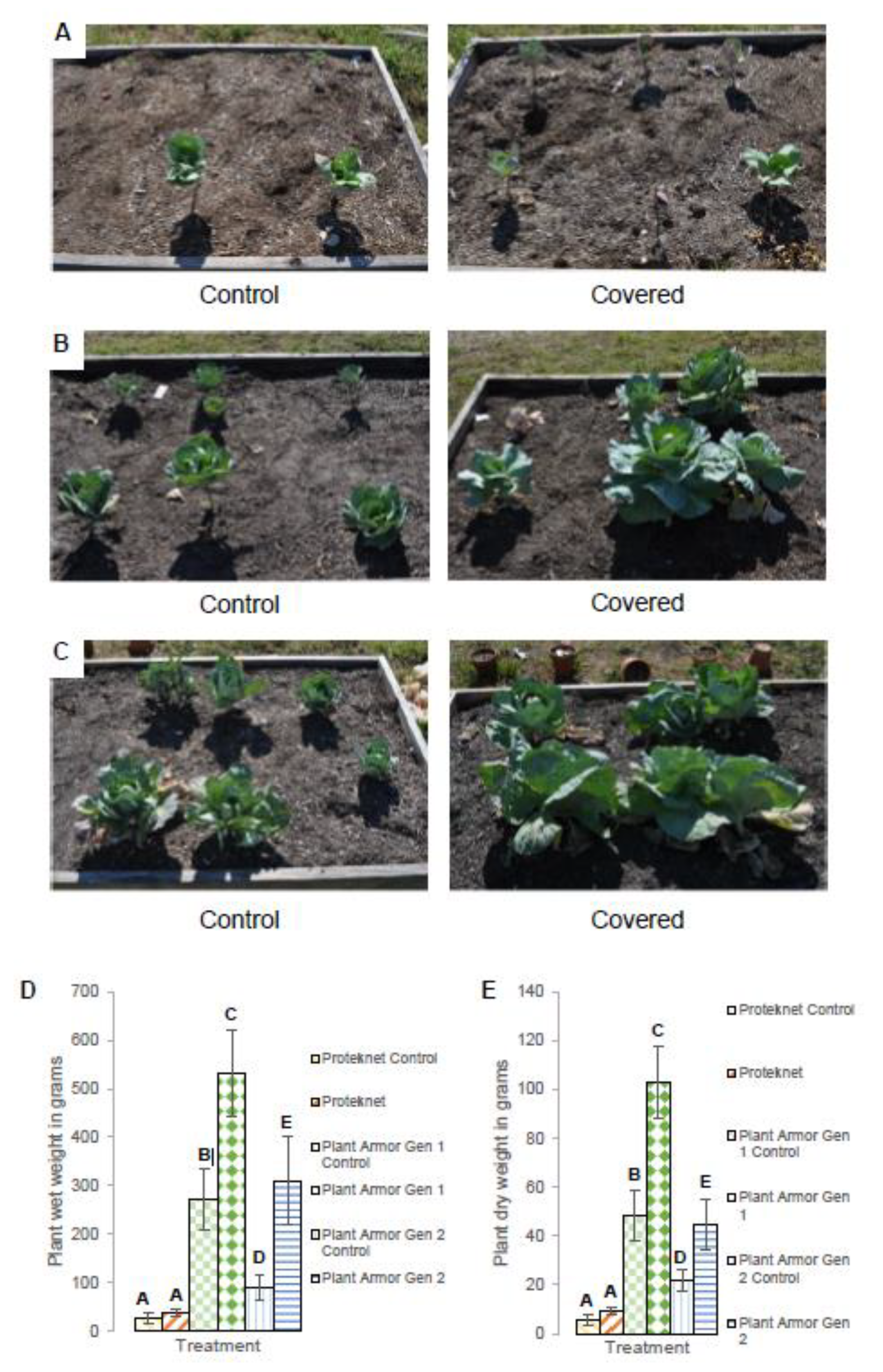
2.6. Small Field Plot Trials
2.7. Statistical Analyses
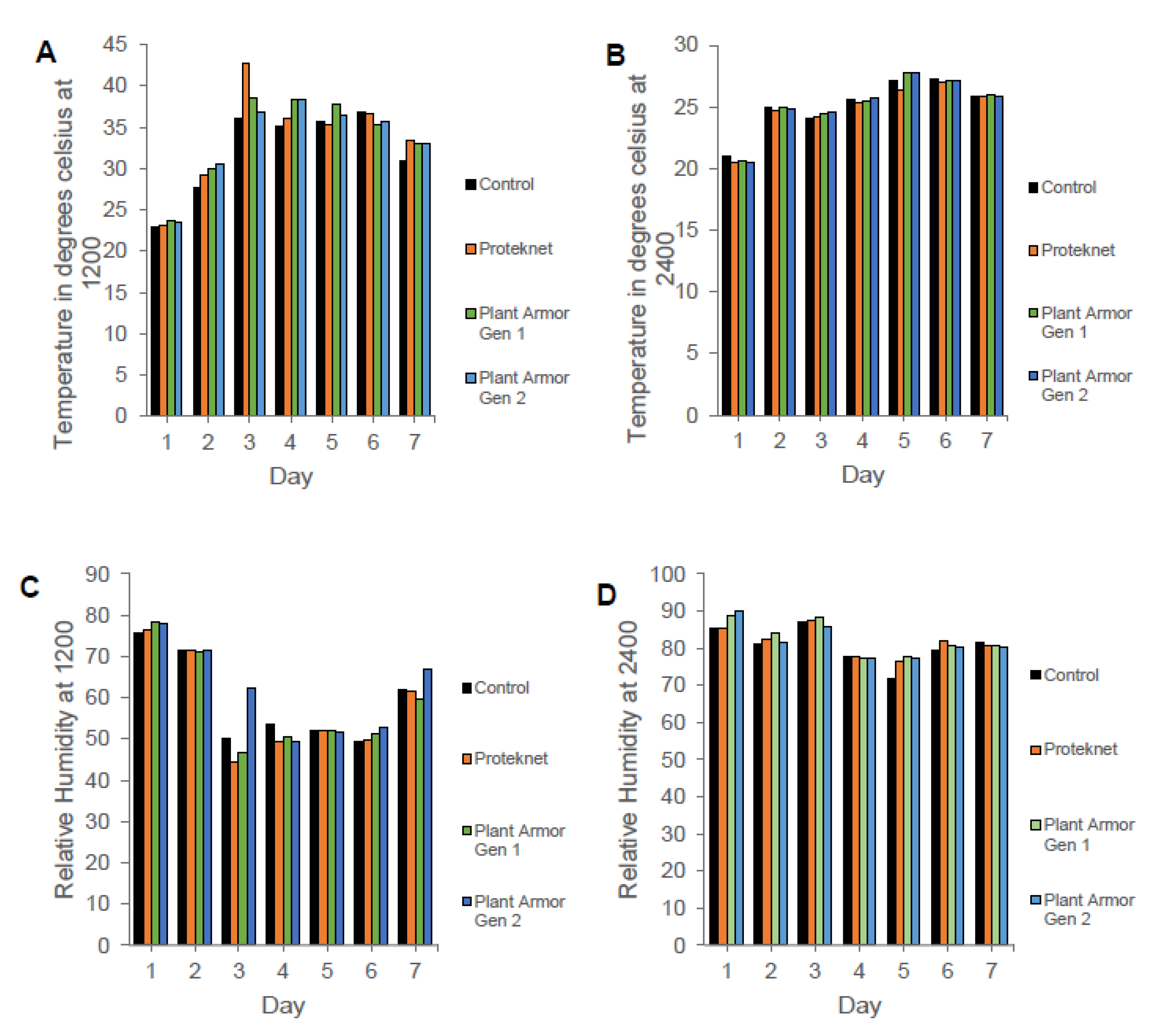
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Radcliffe, E.; Hutchison, W.; Cancelado, R. Integrated Pest Management: Concepts, Tactics, Trategies and Case Studies; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Abrol, D.P. Integrated Pest Management: Current Concepts and Ecological Perspective; Elsevier: New York, NY, USA, 2014. [Google Scholar]
- Roe, R.M.; North Carolina State University, Raleigh, NC, USA. Personal Communication. 2022. [Google Scholar]
- Moar, W.; Roush, R.; Shelton, A.; Ferré, J.; MacIntosh, S.; Leonard, B.R.; Abel, C. Field-evolved resistance to Bt toxins. Nat. Biotechnol. 2008, 26, 1072–1074. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Qureshi, J.A.; Meagher, R.L.; Reisig, D.D.; Head, G.P.; Andow, D.A.; Ni, X.; Kerns, D.; Buntin, G.D.; Niu, Y.; et al. Cry1F resistance in fall armyworm Spodoptera frugiperda: Single gene versus pyramided Bt maize. PLoS ONE 2014, 9, e112958. [Google Scholar] [CrossRef] [Green Version]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef]
- Reisig, D.D.; Huseth, A.S.; Bacheler, J.S.; Aghaee, M.A.; Braswell, L.; Burrack, H.J.; Flanders, K.; Greene, J.K.; Herbert, D.A.; Jacobson, A.; et al. Long-term empirical and observational evidence of practical helicoverpa zea resistance to cotton with pyramided bt toxins. J. Econ. Entomol. 2018, 111, 1824–1833. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.A.; Tsagkarakou, A.; Vontas, J.; Nauen, R. Insecticide resistance in the tomato pinworm Tuta absoluta: Patterns, spread, mechanisms, management and outlook. J. Pest Sci. 2019, 92, 1329–1342. [Google Scholar] [CrossRef]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt Maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Carrire, Y. Field-evolved resistance to Bt cotton: Bollworm in the U.S. and pink bollworm in India. Southwest. Entomol. 2010, 35, 417–424. [Google Scholar] [CrossRef]
- Tefera, T.; Mugo, S.; Mwimali, M.; Anani, B.; Tende, R.; Beyene, Y.; Gichuki, S.; Oikeh, S.O.; Nang’ayo, F.; Okeno, J.; et al. Resistance of Bt-maize (MON810) against the stem borers Busseola fusca (Fuller) and Chilo partellus (Swinhoe) and its yield performance in Kenya. Crop Prot. 2016, 89, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Shew, A.M.; Danforth, D.M.; Nalley, L.L.; Nayga, R.M.; Tsiboe, F.; Dixon, B.L. New innovations in agricultural biotech: Consumer acceptance of topical RNAi in rice production. Food Control 2017, 81, 189–195. [Google Scholar] [CrossRef]
- Mouhamadou, C.S.; Luan, K.; Fodjo, B.K.; West, A.J.; McCord, M.G.; Apperson, C.S.; Roe, R.M. Development of an insecticide-free trapping bednet to control mosquitoes and manage resistance in malaria vector control: A new way of thinking. Insects 2020, 11, 732. [Google Scholar] [CrossRef] [PubMed]
- Luan, K.; West, A.J.; McCord, M.G.; Denhartog, E.A.; Shi, Q.; Bettermann, I.; Li, J.; Travanty, N.V.; Mitchell, R.D.; Cave, G.L.; et al. Mosquito-textile physics: A mathematical roadmap to insecticide-free, bite-proof clothing for everyday life. Insects 2021, 12, 636. [Google Scholar] [CrossRef] [PubMed]
- Basile, B.; Giaccone, M.; Cirillo, C.; Ritieni, A.; Graziani, G.; Shahak, Y.; Forlani, M. Photo-selective hail nets affect fruit size and quality in Hayward kiwifruit. Sci. Hortic. 2012, 141, 91–97. [Google Scholar] [CrossRef]
- Chouinard, G.; Veilleux, J.; Pelletier, F.; Larose, M.; Philion, V.; Cormier, D. Impact of exclusion netting row covers on arthropod presence and crop damage to ‘Honeycrisp’ apple trees in North America: A five-year study. Crop Prot. 2017, 98, 248–254. [Google Scholar] [CrossRef]
- Bosančić, B.; Mićić, N.; Blanke, M.; Pecina, M. A main effects meta principal components analysis of netting effects on fruit: Using apple as a model crop. Plant Growth Regul. 2018, 86, 455–464. [Google Scholar] [CrossRef]
- Kong, Y.; Avraham, L.; Perzelan, Y.; Alkalai-Tuvia, S.; Ratner, K.; Shahak, Y.; Fallik, E. Pearl netting affects postharvest fruit quality in “Vergasa” sweet pepper via light environment manipulation. Sci. Hortic. 2013, 150, 290–298. [Google Scholar] [CrossRef]
- Nordey, T.; Deletre, E.; Mlowe, N.; Martin, T. Small mesh nets protect tomato plants from insect pests and increase yields in eastern Africa. J. Hortic. Sci. Biotechnol. 2020, 95, 222–228. [Google Scholar] [CrossRef]
- Nordey, T.; Deletre, E.; Mlowe, N.; Martin, T. Nethouses protect cucumber plants from insect pests and increase yields in eastern Africa. J. Hortic. Sci. Biotechnol. 2020, 95, 673–678. [Google Scholar] [CrossRef]
- Simon, S.; Komlan, F.A.; Adjaïto, L.; Mensah, A.; Coffi, H.K.; Ngouajio, M.; Martin, T. Efficacy of insect nets for cabbage production and pest management depending on the net removal frequency and microclimate. Int. J. Pest Manag. 2014, 60, 208–216. [Google Scholar] [CrossRef]
- Kiptoo, J.; Kasina, M.; Wanjala, F.; Kipyab, P.; Wasilwa, L.A.; Ngouajio, M.; Martin, T. Use of low cost pest exclusion nets can boost cabbage yield. East Afr. Agric. For. J. 2015, 81, 112–119. [Google Scholar] [CrossRef]
- Bergquist, S.Å.M.; Gertsson, U.E.; Nordmark, L.Y.G.; Olsson, M.E. Ascorbic acid, carotenoids, and visual quality of baby spinach as affected by shade netting and postharvest storage. J. Agric. Food Chem. 2007, 55, 8444–8451. [Google Scholar] [CrossRef] [PubMed]
- Vatsanidou, A.; Bartzanas, T.; Papaioannou, C.; Kittas, C. Efficiency of physical means of IPM on insect population control in greenhouse crops. Acta Hortic. 2011, 893, 1247–1254. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: New York, NY, USA, 1971. [Google Scholar]
- Costa, L.C.B.; Pinto, J.E.B.P.; Castro, E.M.; Alves, E.; Rosal, L.F.; Bertolucci, S.K.V.; Alves, P.B.; Evangelino, T.S. Yield andcomposition of the essential oil of ocimum selloi benth. Cultivated under colored netting. J. Essent. Oil Res. 2010, 22, 34–39. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Milenković, L.; Šunić, L.; Fallik, E. Effect of coloured shade-nets on plant leaf parameters and tomato fruit quality. J. Sci. Food Agric. 2015, 95, 2660–2667. [Google Scholar] [CrossRef] [PubMed]
- Mira-García, A.B.; Conejero, W.; Vera, J.; Ruiz-Sánchez, M.C. Leaf water relations in lime trees grown under shade netting and open-air. Plants 2020, 9, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCaskill, M.R.; McClymont, L.; Goodwin, I.; Green, S.; Partington, D.L. How hail netting reduces apple fruit surface temperature: A microclimate and modelling study. Agric. For. Meteorol. 2016, 226–227, 148–160. [Google Scholar] [CrossRef]
- Gogo, E.O.; Saidi, M.; Ochieng, J.M.; Martin, T.; Baird, V.; Ngouajio, M. Microclimate modification and insect pest exclusion using agronet improve pod yield and quality of French bean. HortScience 2014, 49, 1298–1304. [Google Scholar] [CrossRef]
- Martin, T.; Simon, S.; Parrot, L.; Komlan, F.A.; Vidogbena, F.; Adegbidi, A.; Baird, V.; Saidi, M.; Kasina, M.; Wasilwa, L.A.; et al. Eco-friendly nets to improve vegetable production and quality in sub-Saharan Africa. Acta Hortic. 2015, 1105, 221–227. [Google Scholar] [CrossRef]
- Mashabela, M.N.; Selahle, K.M.; Soundy, P.; Crosby, K.M.; Sivakumar, D. Bioactive Compounds and Fruit Quality of Green Sweet Pepper Grown under Different Colored Shade Netting during Postharvest Storage. J. Food Sci. 2015, 80, H2612–H2618. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cave, G.L.; West, A.J.; McCord, M.G.; Koene, B.; Beck, J.B.; Deguenon, J.M.; Luan, K.; Roe, R.M. Novel 3-D Spacer Textiles to Protect Crops from Insect Infestation and That Enhance Plant Growth. Agriculture 2022, 12, 498. https://doi.org/10.3390/agriculture12040498
Cave GL, West AJ, McCord MG, Koene B, Beck JB, Deguenon JM, Luan K, Roe RM. Novel 3-D Spacer Textiles to Protect Crops from Insect Infestation and That Enhance Plant Growth. Agriculture. 2022; 12(4):498. https://doi.org/10.3390/agriculture12040498
Chicago/Turabian StyleCave, Grayson L., Andre J. West, Marian G. McCord, Bryan Koene, J. Benjamin Beck, Jean M. Deguenon, Kun Luan, and R. Michael Roe. 2022. "Novel 3-D Spacer Textiles to Protect Crops from Insect Infestation and That Enhance Plant Growth" Agriculture 12, no. 4: 498. https://doi.org/10.3390/agriculture12040498
APA StyleCave, G. L., West, A. J., McCord, M. G., Koene, B., Beck, J. B., Deguenon, J. M., Luan, K., & Roe, R. M. (2022). Novel 3-D Spacer Textiles to Protect Crops from Insect Infestation and That Enhance Plant Growth. Agriculture, 12(4), 498. https://doi.org/10.3390/agriculture12040498