
Dioecy in Flowering Plants: From the First Observations of Prospero Alpini in the XVI Century to the Most Recent Advances in the Genomics Era




{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Prospero Alpini and the History of Dioecy
| […] Hæc arbor alternis tantum annis copiofiores fructus edit, neque (quod dictu valde mirabile videtur) fœminae concipiunt, ac fructificat, ni in ramis maris foœminę ramos aliquis permifcuerit, ac se quasi osculari permiferit. Plerique fœminas ut fœcundent, non ramos, sed pulurem intra maris inuolucrum inuentam supra fœmi narum ramos, atque cor spargunt, vel alijflores maris pulueris loco spargere solent. ni enim Aegyptii hoc fecerint, fine dubio fœmi næ vel nullos frudus ferent, vel quos ferent, non retinebunt, neque ni maturescent. [...] | […] This tree produces abundant fruit only in alternate years, and (which seems to be very admirable) females conceive and bear fruit only if someone has grafted the branches of the female into the branches of the male and allowed them to kiss. Some do not use the branches to fertilize the females, but the dust found on them, while others usually scatter the flowers of the male instead of the dust. In fact, if in fact the Egyptians did not do this, the females either do not produce any fruit, or those who produce will not keep them and these will not reach maturity. […] |
2. Dioecy in Angiosperms
3. Models for Sex Determination in Angiosperms
4. Plant Species Supporting Models
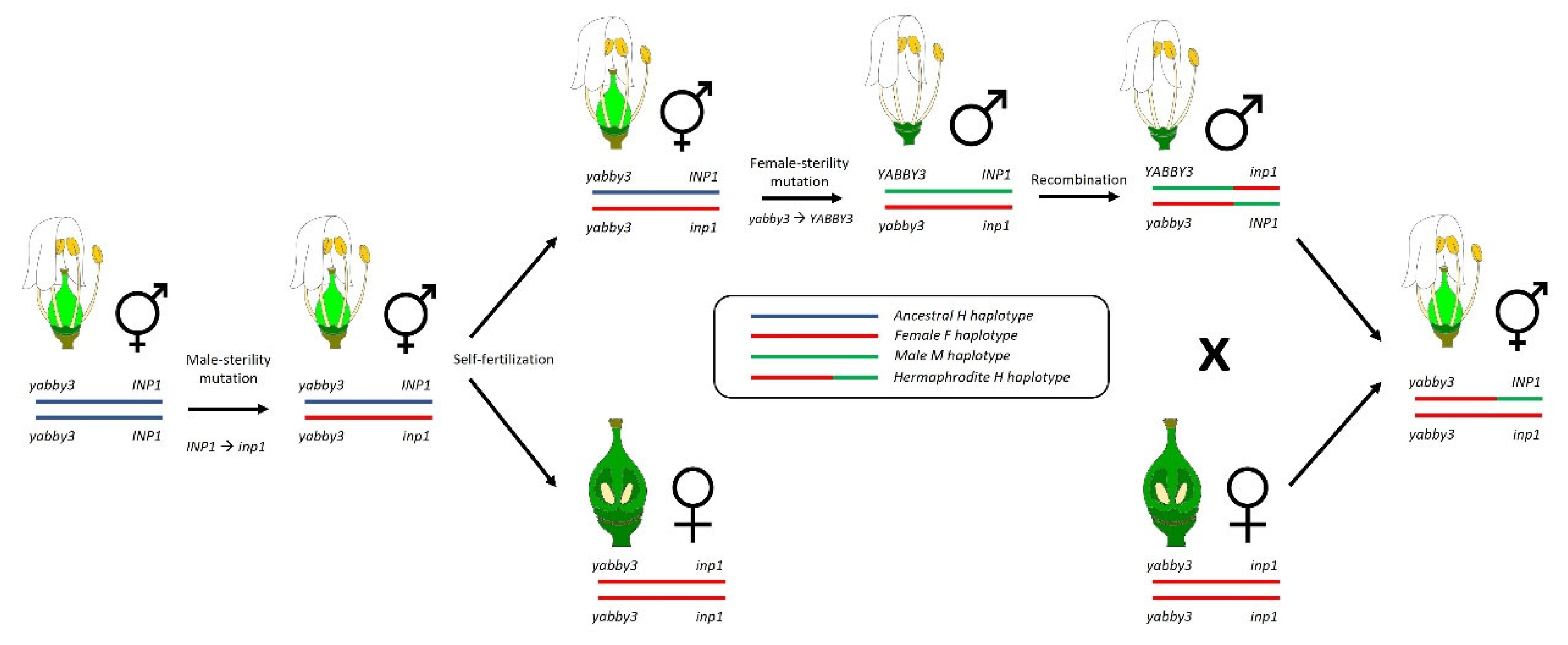
4.1. The Two-Gene Model in the Vitis Genus
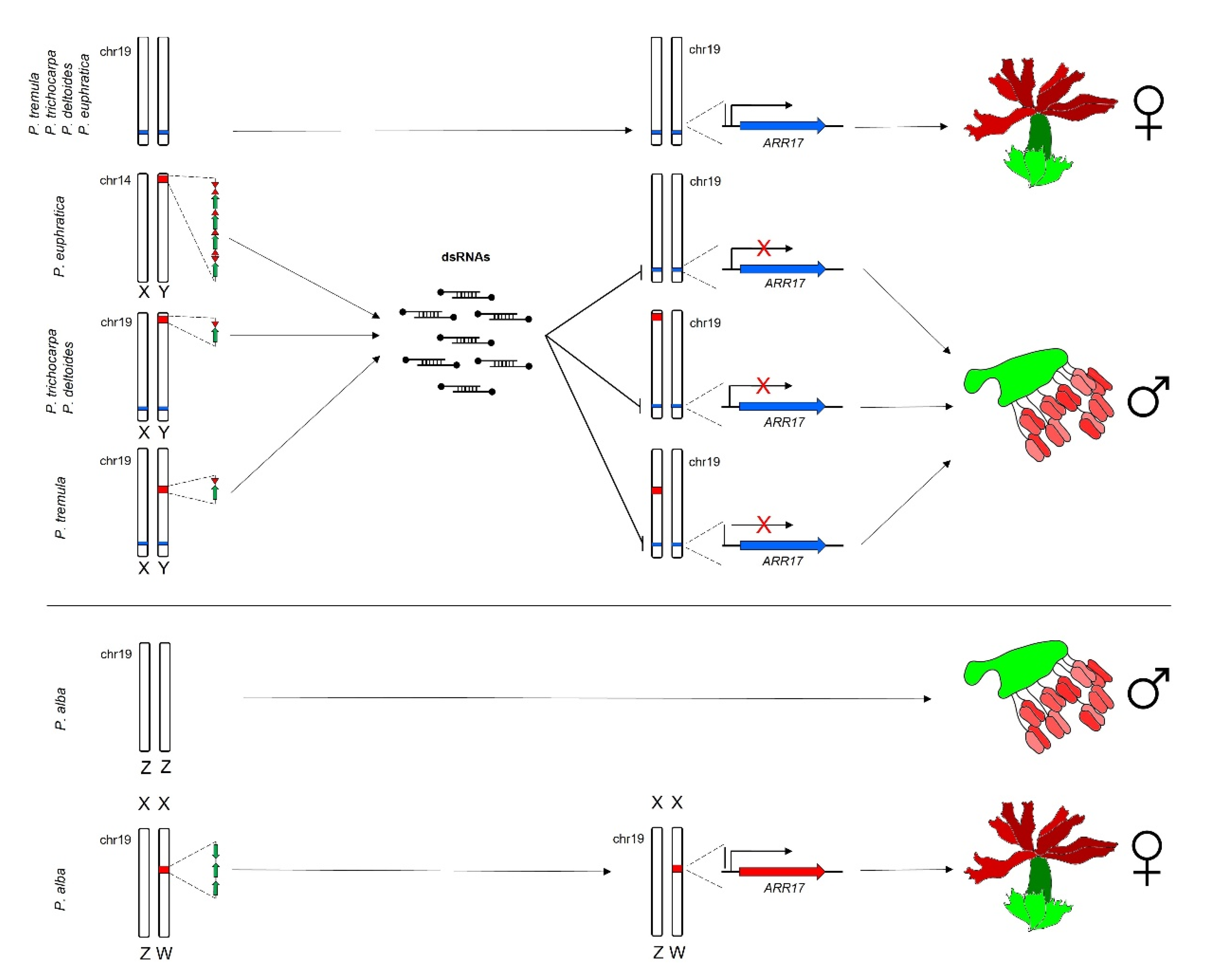
4.2. The One-Gene Model in Poplar Genus
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Alpini, P. De Plantis Aegypti Liber; Venetiis, Apud Franciscum de Franciscis Senensem: Venetia, Italy, 1592. [Google Scholar]
- WFO (2022): World Flora Online. Available online: http://www.worldfloraonline.org/ (accessed on 15 January 2022).
- Renner, S.S.; Ricklefs, R.E. Dioecy and its correlates in the flowering plants. Am. J. Bot. 1995, 82, 596–606. [Google Scholar] [CrossRef] [Green Version]
- Villarreal, J.C.; Renner, S.S. Correlates of monoicy and dioicy in hornworts, the apparent sister group to vascular plants. BMC Evol. Biol. 2013, 13, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walas, Ł.; Mandryk, W.; Thomas, P.A.; Tyrała-Wierucka, Ż.; Iszkuło, G. Sexual systems in gymnosperms: A review. Basic Appl. Ecol. 2018, 31, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Renner, S.S. The relative and absolute frequencies of angiosperm sexual systems: Dioecy, monoecy, gynodioecy, and an updated online database. Am. J. Bot. 2014, 101, 1588–1596. [Google Scholar] [CrossRef] [Green Version]
- Käfer, J.; de Boer, H.; Mousset, S.; Kool, A.; Dufaÿ, M.; Marais, G.A.B. Dioecy is associated with higher diversification rates in flowering plants. J. Evol. Biol. 2014, 27, 1478–1490. [Google Scholar] [CrossRef] [Green Version]
- Heilbuth, J.C. Lower Species Richness in Dioecious Clades. Am. Nat. 2000, 156, 221–241. [Google Scholar] [CrossRef]
- Lewis, B.Y.D. Male Sterility in Natural Populations the Equilibrium Between Females and Hermaphrodites to be Expected with Different. New Phytol. 1941, 40, 56–63. [Google Scholar] [CrossRef]
- Westergaard, M. The Mechanism of Sex Determination in Dioecious Flowering Plants. Adv. Genet. 1958, 9, 217–281. [Google Scholar] [CrossRef]
- Endress, P.K.; Doyle, J.A. Reconstructing the ancestral angiosperm flower and its initial specializations. Am. J. Bot. 2009, 96, 22–66. [Google Scholar] [CrossRef]
- Barcaccia, G.; Palumbo, F.; Sgorbati, S.; Albertini, E.; Pupilli, F. A Reappraisal of the Evolutionary and Developmental Pathway of Apomixis and Its Genetic Control in Angiosperms. Genes 2020, 11, 859. [Google Scholar] [CrossRef]
- King, R.C.; Stansfield, W.D.; Mulligan, P.K. A Dictionary of Genetics, 7th ed.; Oxford: Oxford, UK, 2006. [Google Scholar]
- He, L.; Jia, K.; Zhang, R.; Wang, Y.; Shi, T.; Li, Z.; Zeng, S.; Cai, X.; Wagner, N.D.; Hörandl, E.; et al. Chromosome-scale assembly of the genome of Salix dunnii reveals a male-heterogametic sex determination system on chromosome 7. Mol. Ecol. Resour. 2021, 21, 1966–1982. [Google Scholar] [CrossRef] [PubMed]
- Hallingbäck, H.R.; Pucholt, P.; Ingvarsson, P.K.; Rönnberg-Wästljung, A.C.; Berlin, S. Genome-wide association mapping uncovers sex-associated copy number variation markers and female hemizygous regions on the W chromosome in Salix viminalis. BMC Genom. 2021, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Slancarova, V.; Zdanska, J.; Janousek, B.; Talianova, M.; Zschach, C.; Zluvova, J.; Siroky, J.; Kovacova, V.; Blavet, H.; Danihelka, J.; et al. Evolution of Sex Determination Systems with Heterogametic Males and Females Insilene. Evolution 2013, 67, 3669–3677. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, M.; Natsume, S.; Takagi, H.; White, B.; Yaegashi, H.; Shimizu, M.; Yoshida, K.; Uemura, A.; Oikawa, K.; Abe, A.; et al. Genome sequencing of the staple food crop white Guinea yam enables the development of a molecular marker for sex determination. BMC Biol. 2017, 15, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Pennell, M.W.; Mank, J.E.; Peichel, C.L. Transitions in sex determination and sex chromosomes across vertebrate species. Mol. Ecol. 2018, 27, 3950–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kottler, V.A.; Feron, R.; Nanda, I.; Klopp, C.; Du, K.; Kneitz, S.; Helmprobst, F.; Lamatsch, D.K.; Lopez-Roques, C.; Lluch, J.; et al. Independent Origin of XY and ZW Sex Determination. Genetics 2020, 214, 193–209. [Google Scholar] [CrossRef]
- Charlesworth, B.; Charlesworth, D. A Model for the Evolution of Dioecy and Gynodioecy Published by The University of Chicago Press for The American Society of Naturalists Stable. Am. Nat. 1978, 112, 975–997. [Google Scholar] [CrossRef]
- Pannell, J.R. The Evolution and Maintenance of Androdioecy. Annu. Rev. Ecol. Syst. 2002, 33, 397–425. [Google Scholar] [CrossRef]
- Akagi, T.; Henry, I.M.; Ohtani, H.; Morimoto, T.; Beppu, K.; Kataoka, I.; Tao, R. A Y-Encoded Suppressor of Feminization Arose via Lineage-Specific Duplication of a Cytokinin Response Regulator in Kiwifruit. Plant Cell 2018, 30, 780–795. [Google Scholar] [CrossRef]
- Massonnet, M.; Cochetel, N.; Minio, A.; Vondras, A.M.; Lin, J.; Muyle, A.; Garcia, J.F.; Zhou, Y.; Delledonne, M.; Riaz, S.; et al. The genetic basis of sex determination in grapes. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Torres, M.F.; Mathew, L.S.; Ahmed, I.; Al-Azwani, I.K.; Krueger, R.; Rivera-Nuñez, D.; Mohamoud, Y.A.; Clark, A.G.; Suhre, K.; Malek, J.A. Genus-wide sequencing supports a two-locus model for sex-determination in Phoenix. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, N.A.; Kersten, B.; Leite Montalvão, A.P.; Mähler, N.; Bernhardsson, C.; Bräutigam, K.; Carracedo Lorenzo, Z.; Hoenicka, H.; Kumar, V.; Mader, M.; et al. A single gene underlies the dynamic evolution of poplar sex determination. Nat. Plants 2020, 6, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Ma, X.; Wadlington, W.; Ming, R. Identification of structural variation and polymorphisms of a sex co-segregating scaffold in spinach. Plant Reprod. 2021, 35, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Golenberg, E.M.; West, N.W. Hormonal interactions and gene regulation can link monoecy and environmental plasticity to the evolution of dioecy in plants. Am. J. Bot. 2013, 100, 1022–1037. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Schwaninger, H.R.; Baldo, A.M.; Labate, J.; Zhong, G.-Y.; Simon, C.J. A phylogenetic analysis of the grape genus (Vitis L.) reveals broad reticulation and concurrent diversification during neogene and quaternary climate change. BMC Evol. Biol. 2013, 13, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Coito, J.L.; da Silva, H.S.G.; Ramos, M.J.N.; Montez, M.; Cunha, J.; Amancio, S.; Costa, M.M.R.; Rocheta, M. Vitis Flower Sex Specification Acts Downstream and Independently of the ABCDE Model Genes. Front. Plant Sci. 2018, 9, 1–20. [Google Scholar] [CrossRef]
- Fechter, I.; Hausmann, L.; Daum, M.; Sörensen, T.R.; Viehöver, P.; Weisshaar, B.; Töpfer, R. Candidate genes within a 143 kb region of the flower sex locus in Vitis. Mol. Genet. Genom. 2012, 287, 247–259. [Google Scholar] [CrossRef]
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends Genet. 2006, 22, 511–519. [Google Scholar] [CrossRef]
- Picq, S.; Santoni, S.; Lacombe, T.; Latreille, M.; Weber, A.; Ardisson, M.; Ivorra, S.; Maghradze, D.; Arroyo-Garcia, R.; Chatelet, P.; et al. A small XY chromosomal region explains sex determination in wild dioecious. BMC Plant Biol. 2014, 14, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Henry, I.M.; Akagi, T.; Tao, R.; Comai, L. One Hundred Ways to Invent the Sexes: Theoretical and Observed Paths to Dioecy in Plants. Annu. Rev. Plant Biol. 2018, 69, 553–575. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.; Massonnet, M.; Minio, A.; Patel, S.; Llaca, V.; Karn, A.; Gouker, F.; Cadle-Davidson, L.; Reisch, B.; Fennell, A.; et al. Multiple independent recombinations led to hermaphroditism in grapevine. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Battilana, J.; Lorenzi, S.; Moreira, F.M.; Moreno-Sanz, P.; Failla, O.; Emanuelli, F.; Grando, M.S. Linkage Mapping and Molecular Diversity at the Flower Sex Locus in Wild and Cultivated Grapevine Reveal a Prominent SSR Haplotype in Hermaphrodite Plants. Mol. Biotechnol. 2013, 54, 1031–1037. [Google Scholar] [CrossRef] [Green Version]
- Riaz, S.; Krivanek, A.F.; Xu, K.; Walker, M.A. Refined mapping of the Pierce’s disease resistance locus, PdR1, and Sex on an extended genetic map of Vitis rupestris × V. arizonica. Theor. Appl. Genet. 2006, 113, 1317–1329. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Minio, A.; Massonnet, M.; Solares, E.; Lv, Y.; Beridze, T.; Cantu, D.; Gaut, B.S. The population genetics of structural variants in grapevine domestication. Nat. Plants 2019, 5, 965–979. [Google Scholar] [CrossRef] [PubMed]
- Badouin, H.; Velt, A.; Gindraud, F.; Flutre, T.; Dumas, V.; Vautrin, S.; Marande, W.; Corbi, J.; Sallet, E.; Ganofsky, J.; et al. The wild grape genome sequence provides insights into the transition from dioecy to hermaphroditism during grape domestication. Genome Biol. 2020, 21, 1–24. [Google Scholar] [CrossRef]
- Paolucci, I.; Gaudet, M.; Jorge, V.; Beritognolo, I.; Terzoli, S.; Kuzminsky, E.; Muleo, R.; Mugnozza, G.S.; Sabatti, M. Genetic linkage maps of Populus alba L. and comparative mapping analysis of sex determination across Populus species. Tree Genet. Genomes 2010, 6, 863–875. [Google Scholar] [CrossRef]
- Geraldes, A.; Hefer, C.; Capron, A.; Kolosova, N.; Martinez-Nuñez, F.; Soolanayakanahally, R.Y.; Stanton, B.J.; Guy, R.D.; Mansfield, S.; Douglas, C.J.; et al. Recent Y chromosome divergence despite ancient origin of dioecy in poplars (Populus). Mol. Ecol. 2015, 24, 3243–3256. [Google Scholar] [CrossRef]
- Zhou, R.; Macaya-Sanz, D.; Schmutz, J.; Jenkins, J.W.; Tuskan, G.A.; DiFazio, S.P. Sequencing and Analysis of the Sex Determination Region of Populus trichocarpa. Genes 2020, 11, 843. [Google Scholar] [CrossRef]
- Pakull, B.; Groppe, K.; Meyer, M.; Markussen, T.; Fladung, M. Genetic linkage mapping in aspen (Populus tremula L. and Populus tremuloides Michx.). Tree Genet. Genomes 2009, 5, 505–515. [Google Scholar] [CrossRef]
- Kersten, B.; Pakull, B.; Groppe, K.; Lueneburg, J.; Fladung, M. The sex-linked region in Populus tremuloides Turesson 141 corresponds to a pericentromeric region of about two million base pairs on P. trichocarpa chromosome 19. Plant Biol. 2013, 16, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wang, D.; Li, Y.; Zhang, Z.; Tong, S.; Li, M.; Zhang, X.; Zhang, L.; Ren, L.; Ma, X.; et al. A General Model to Explain Repeated Turnovers of Sex Determination in the Salicaceae. Mol. Biol. Evol. 2020, 38, 968–980. [Google Scholar] [CrossRef] [PubMed]
- McKown, A.D.; Klápště, J.; Guy, R.D.; Soolanayakanahally, R.Y.; La Mantia, J.; Porth, I.; Skyba, O.; Unda, F.; Douglas, C.J.; El-Kassaby, Y.A.; et al. Sexual homomorphism in dioecious trees: Extensive tests fail to detect sexual dimorphism in Populus. Sci. Rep. 2017, 7, 1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, G.; Sanderson, B.; Keefover-Ring, K.; Liu, J.; Ma, T.; Yin, T.; Smart, L.B.; DiFazio, S.P.; Olson, M.S. Pathways to sex determination in plants: How many roads lead to Rome? Curr. Opin. Plant Biol. 2020, 54, 61–68. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vannozzi, A.; Palumbo, F.; Lucchin, M.; Barcaccia, G. Dioecy in Flowering Plants: From the First Observations of Prospero Alpini in the XVI Century to the Most Recent Advances in the Genomics Era. Agriculture 2022, 12, 364. https://doi.org/10.3390/agriculture12030364
Vannozzi A, Palumbo F, Lucchin M, Barcaccia G. Dioecy in Flowering Plants: From the First Observations of Prospero Alpini in the XVI Century to the Most Recent Advances in the Genomics Era. Agriculture. 2022; 12(3):364. https://doi.org/10.3390/agriculture12030364
Chicago/Turabian StyleVannozzi, Alessandro, Fabio Palumbo, Margherita Lucchin, and Gianni Barcaccia. 2022. "Dioecy in Flowering Plants: From the First Observations of Prospero Alpini in the XVI Century to the Most Recent Advances in the Genomics Era" Agriculture 12, no. 3: 364. https://doi.org/10.3390/agriculture12030364
APA StyleVannozzi, A., Palumbo, F., Lucchin, M., & Barcaccia, G. (2022). Dioecy in Flowering Plants: From the First Observations of Prospero Alpini in the XVI Century to the Most Recent Advances in the Genomics Era. Agriculture, 12(3), 364. https://doi.org/10.3390/agriculture12030364