
Pyramiding Submergence Tolerance and Three Bacterial Blight Resistance Genes in Popular Rice Variety Hasanta through Marker-Assisted Backcross Breeding
 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Breeding Strategy
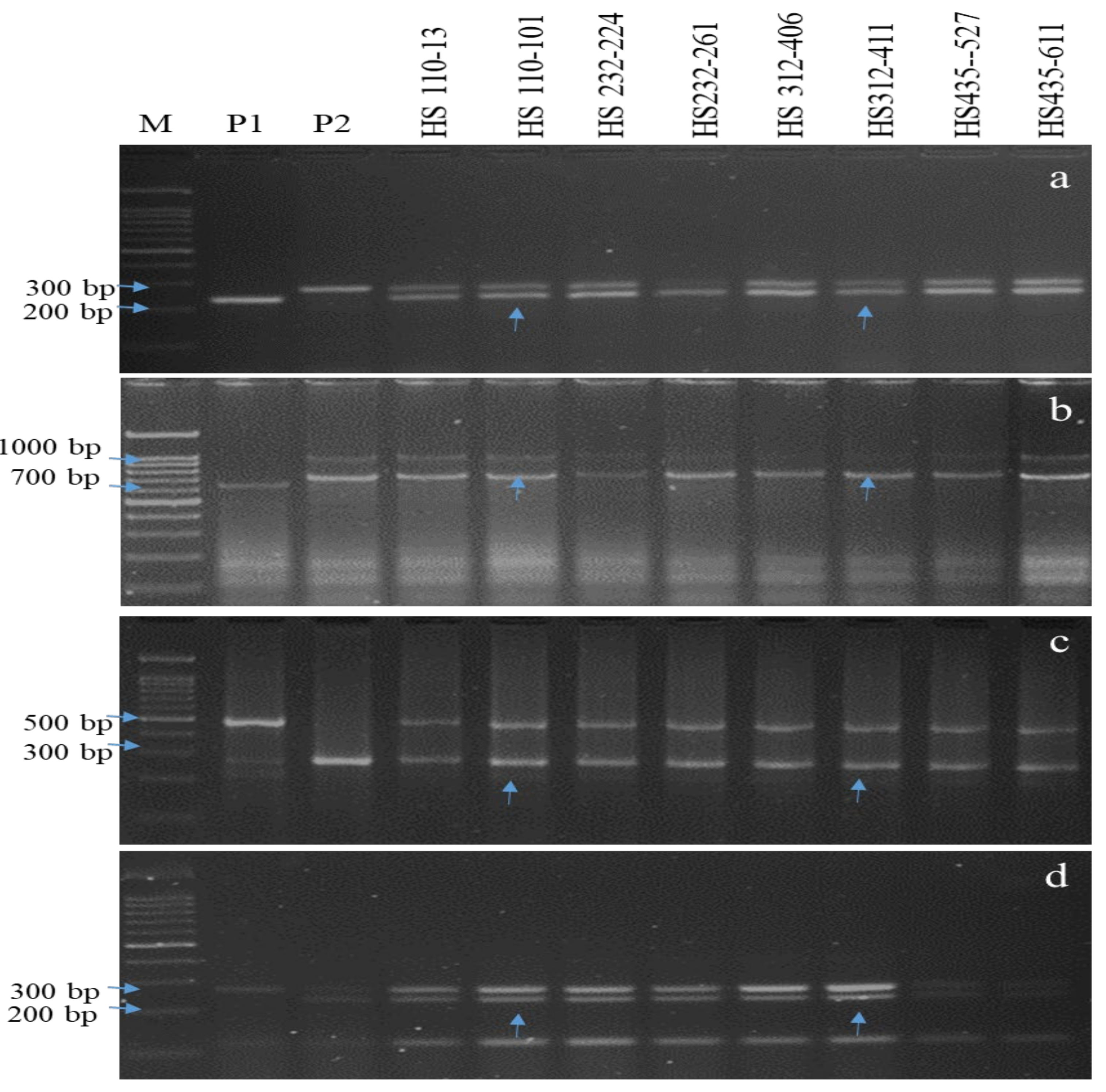
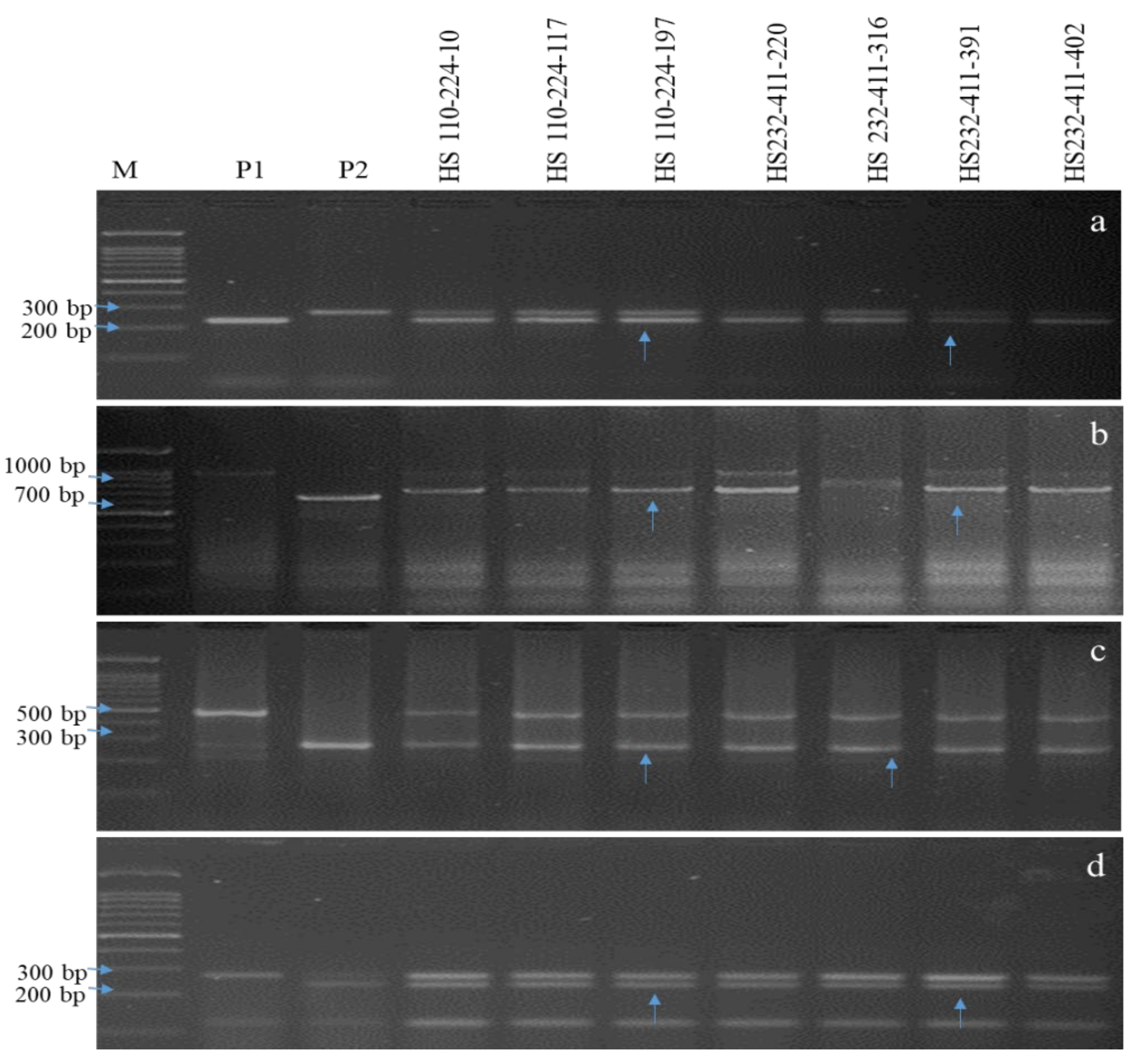
2.2. PCR and Marker Analysis
2.3. Screening for Submergence Tolerance
2.4. Disease Bioassay Analysis
2.5. Speed Breeding and Single Seed Descent (SSD) Strategies for Generation Advancement
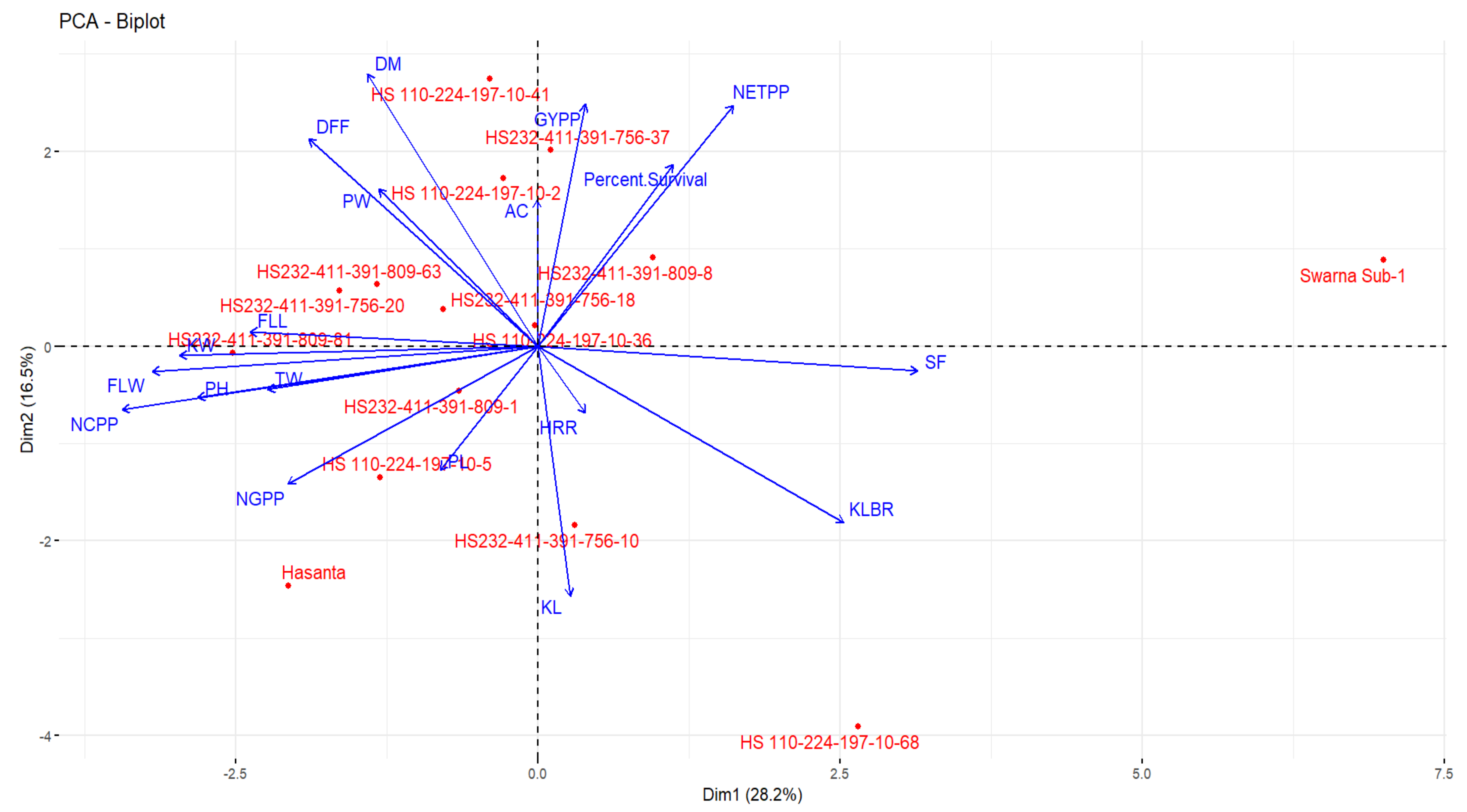
2.6. Agro-Morpho Evaluation of NILs
3. Results
3.1. Molecular Characterization of Parents and Selection of Donor
3.2. MAB Based Introgression of Sub1 and Three BB Resistance Genes in Hasanta
3.3. Genome Introgression on the Carrier and Non-Carrier Chromosomes
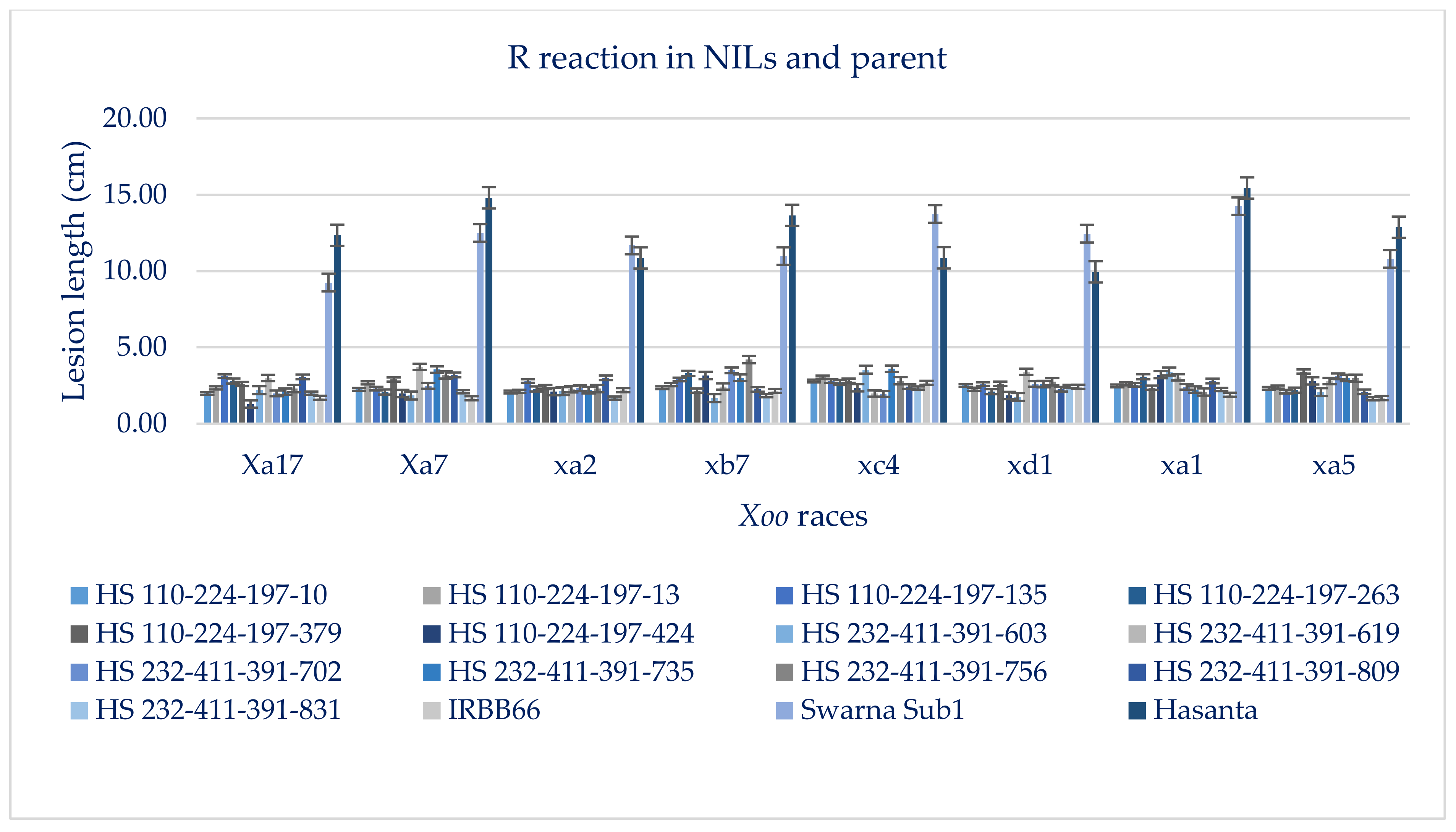
3.4. Screening of NILs for Submergence Tolerance and BB Resistance
3.5. Physiological Responses of Sub1 Gene in NILs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ismail, A.M.; Singh, U.S.; Singh, S.; Dar, M.H.; Mackill, D.J. The contribution of submergence tolerant (Sub1) rice varieties to food security in flood prone rainfed lowland areas in Asia. Field Crops Res. 2013, 152, 83–93. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Baksh, S.Y.; Mukherjee, A.K.; Mohanty, S.P. Development of flash-flood tolerant and durable bacterial blight resistant versions of mega rice variety ‘Swarna’ through marker-assisted backcross breeding. Sci. Rep. 2019, 9, 12810. [Google Scholar] [CrossRef] [PubMed]
- Iftekharuddaula, K.M.; Ahmed, H.U.; Ghosal, S.; Moni, Z.R.; Amin, A.; Ali, M.S. Development of a new submergence tolerant rice variety for Bangladesh using marker-assisted backcrossing. Rice Sci. 2015, 22, 15–26. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Nayak, D.K.; Mohanty, S.; Behera, L.; Barik, S.R.; Pandit, E.; Lenka, S.; Anandan, A. Pyramiding of three bacterial blight resistance genes for broad-spectrum resistance in deepwater rice variety, Jalmagna. Rice 2015, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Khush, G.S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef]
- Kumar, A.; Nayak, A.K.; Hanjagi, P.S.; Kumari, K.; Vijaykumar, S.; Mohanthy, S.; Tripathi, R.; Pannerselvam, P. Submergence stress in rice: Adaptive mechanism, coping strategies and future research needs. Environ. Exp. Bot. 2021, 186, 104448. [Google Scholar] [CrossRef]
- Mackill, D.J.; Ismail, A.M.; Singh, U.S.; Labios, R.V.; Paris, T.R. Development and rapid adoption of submergence-tolerant (Sub1) rice varieties. Ad. Agron. 2012, 115, 252–299. [Google Scholar]
- Sarkar, R.K.; Das, K.K.; Panda, D.; Reddy, J.N.; Patnaik, S.S.C.; Patra, B.C.; Singh, D.P. Submergence Tolerance in Rice: Biophysical Constraints, Physiological Basis and Identification of Donors; Central Rice Research Institute: Cuttack, India, 2014; Volume 36. [Google Scholar]
- Pradhan, K.C.; Barik, S.R.; Mohapatra, S.; Nayak, D.K.; Pandit, E.; Jena, B.K.; Sangeeta, S.; Pradhan, A.; Samal, A.; Meher, J.; et al. Incorporation of two bacterial blight resistance genes into the popular rice variety, ranidhan through marker-assisted breeding. Agriculture 2022, 12, 1287. [Google Scholar] [CrossRef]
- Sanchez, A.C.; Brar, D.S.; Huang, N.; Li, Z.; Khush, G.S. Sequence tagged site marker-assisted selection for three bacterial blight resistance genes in rice. Crop Sci. 2000, 40, 792–797. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Hasan, M.M.; Rafii, M.Y.; Ismail, M.R.; Mahmood, M.; Rahim, H.A.; Alam, M.A. Marker assisted backcrossing: A useful method for rice improvement. Biotech. Biotechnol. Equip. 2015, 29, 237–254. [Google Scholar] [CrossRef]
- Dash, A.K.; Rao, R.N.; Rao, G.J.N.; Verma, R.L.; Katara, J.L.; Mukherjee, A.K.; Singh, O.N.; Bagchi, T.B. Phenotypic and Marker-Assisted Genetic Enhancement of Parental Lines of Rajalaxmi, an Elite Rice Hybrid. Front. Plant Sci. 2016, 7, 1005. [Google Scholar] [CrossRef]
- Kumar, M.; Singh, R.P.; Singh, O.N.; Singh, P.; Arsode, P.; Jena, D.; Samantaray, S.; Verma, R.L. Generation mean analysis for bacterial blight resistance and yield traits in rice. J. Pharm. Phytochem. 2019, 8, 2120–2124. [Google Scholar]
- Ronald, P.C.; Albano, B.; Tabien, R.; Abenes, M.L.P.; Wu, K.S.; McCouch, S.R. Genetic and physical analysis of the rice bacterial blight disease resistance locus Xa21. Mol. Gen. Genet. 1992, 236, 113–120. [Google Scholar] [CrossRef]
- Adkins, S.W.; Shiraishi, T.; McComb, J.A. Submergence tolerance of rice; a new glasshouse method for the experimental submergence of plants. Plant Physiol. 1990, 80, 642–646. [Google Scholar] [CrossRef]
- Park, C.; Ronald, P.C. Cleavage and nuclear localization of the rice Xa21 immune receptor. Nat. Commun. 2012, 3, 920. [Google Scholar] [CrossRef]
- Catling, D. Rice in Deep Water; International Rice Research Institute: Manila, Philippines, 1992. [Google Scholar]
- Cen, M.M.; Li, W.; Wang, R.; Luo, S. The molecular regulatory pathways and metabolic adaptation in the seed germination and early seedling growth of rice in response to low O2 stress. Plants 2020, 9, 1363. [Google Scholar]
- Samanta, P.; Ganie, S.A.; Chakraborty, A.; Dey, N. Study on regulation of carbohydrate usage in a heterogeneous rice population under submergence. J. Plant Biochem. Biotechnol. 2021, 30, 138–146. [Google Scholar] [CrossRef]
- Das, K.K.; Sarkar, R.K.; Ismail, A.M. Elongation ability and non-structural carbohydrate levels in relation to submergence tolerance in rice. Plant Sci. 2005, 168, 131–136. [Google Scholar] [CrossRef]
- Kende, H.; Knaap, E.; Cho, H. Deep water rice: A model plant to study stem elongation. Plant Physiol. 1998, 118, 1105–1110. [Google Scholar] [CrossRef]
- Rajpurohit, D.; Kumar, R.; Kumar, M.; Paul, P.; Awasthi, A.; Osman, B.P.; Puri, A.; Jhang, T.; Singh, K.; Dhaliwal, H.S. Pyramiding of two bacterial blight resistance and a semi dwarfing gene in type 3 Basmati using marker-assisted selection. Euphytica 2011, 178, 111–126. [Google Scholar] [CrossRef]
- Chetia, S.K.; Kalita, M.; Verma, R.K.; Barua, B.; Ahmed, T.M.; Modi, K.; Singh, N.K. Flood proofing of Ranjit, a popular variety of North-Eastern India through transfer of Sub1 rice QTL by modified marker-assisted backcross breeding. Indian J. Genet. 2018, 78, 166–173. [Google Scholar] [CrossRef]
- Kabir, M.E.; Iftekharuddaula, K.M.; Khan, M.A.I.; Mian, M.A.K.; Ivy, N.A. Marker assisted introgression of bacterial blight resistant gene into submergence tolerance rice variety BRRI Dhan52. Bangladesh J. Agril. Res. 2017, 42, 403–411. [Google Scholar] [CrossRef]
- Manivong, P.; Korinsak, K.; Korinsak, S.; Siangliw, J.L.; Vanavichit, A.; Toojinda, T. Marker-assisted selection to improve submergence tolerance, blast resistance and strong fragrance in glutinous rice. Thai J. Genet. 2014, 7, 110–122. [Google Scholar]
- Nair, M.M.; Shylaraj, K.S. Introgression of dual abiotic stress tolerance QTLs (Saltol QTL and Sub1 gene) into rice (Oryza sativa L.) variety Aiswarya through marker assisted backcross breeding. Physiol. Mol. Biol. Plants 2021, 27, 497–514. [Google Scholar] [CrossRef] [PubMed]
- Neeraja, C.N.; Rodriguez, M.R.; Pamplona, A.; Heuer, S.; Collard, B.C.; Septiningsih, E.M.; Vergara, G.; Sanchez, D.; Xu, K.; Ismail, A.M.; et al. A marker-assisted backcross approach for developing submergence-tolerant rice cultivars. Theor. Appl. Genet. 2007, 115, 767–776. [Google Scholar] [CrossRef] [PubMed]
- ICAR-National Rice Research Institute (NRRI). Annual-Report; NRRI: Cuttack, India, 2018; pp. 5–8. [Google Scholar]
- Rahman, H.; Dakshinamurthi, V.; Ramasamy, S.; Manickam, S.; Kaliyaperumal, A.K.; Raha, S.; Paneerselvam, N.; Ramanathan, V.; Nallathambi, J.; Sabariyappan, R.; et al. Introgression of submergence tolerance into CO43, a popular rice variety of india, through marker-assisted backcross breeding. Czech J. Genet. Plant Breed. 2018, 54, 101–108. [Google Scholar]
- Singh, P.; Sinha, A.K. A positive feedback loop governed by SUB1A1 interaction with MITOGEN-ACTIVATED PROTEIN KINASE3 imparts submergence tolerance in rice. Plant Cell 2016, 28, 1127–1143. [Google Scholar] [CrossRef]
- Singh, R.; Singh, Y.; Xalaxo, S.; Verulkar, S.; Yadav, N.; Singh, S.; Singh, N.; Prasad, K.S.N.; Kondayya, K.; Rao, P.V.R.; et al. From QTL to variety- harnessing the benefits of QTLs for drought, flood and salt tolerance in mega rice varieties of India through a multi-institutional network. Plant Sci. 2016, 242, 278–287. [Google Scholar] [CrossRef]
- Chen, M.; Presting, G.; Barbazuk, W.B.; Goicoechea, J.L.; Blackmon, B.; Fang, G.; Kim, H.; Frisch, D.; Yu, Y.; Sun, S.; et al. An integrated physical and genetic map of the rice genome. Plant Cell 2002, 14, 537–545. [Google Scholar] [CrossRef]
- Toojinda, T.; Siangliw, M.; Tragoonrung, S.; Vanavichit, A. Molecular genetics of submergence tolerance in rice: QTL analysis of key traits. Ann. Bot. 2003, 91, 243–253. [Google Scholar] [CrossRef]
- Verma, R.L.; Singh, S.; Singh, P.; Kumar, V.; Singh, S.P.; Singh, S.; Samantaray, S.; Singh, O.N. Genetic purity assessment of indica rice hybrids through DNS fingerprinting and grow-out test. J. Environ. Biol. 2017, 38, 1321–1331. [Google Scholar] [CrossRef]
- Xu, K.; Mackill, D.J. A major locus for submergence tolerance mapped on rice chromosome 9. Mol. Breed. 1996, 2, 219–224. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey- Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef]
- Ou, S.H. Rice Diseases, 2nd ed.; Commonwealth Mycological Institute: Kew, UK, 1985; p. 380. [Google Scholar]
- Rout, D.; Jena, D.; Singh, V.; Kumar, M.; Arsode, P.; Singh, P.; Katara, J.L.; Samantaray, S.; Verma, R.L. Hybrid rice research: Current status and prospects. In Recent Advances in Rice Research, 1st ed.; Ansari, M.R., Ed.; IntechOpen: London, UK, 2020; p. 93668. [Google Scholar]
- Khush, G.S.; Mackill, D.J.; Sidhu, G.S. Breeding Rice for Resistance to Bacterial Leaf Blight; IRRI, Ed.; IRRI: Manila, Philippines, 1989; pp. 207–217. [Google Scholar]
- Das, G.; Rao, G.J.N.; Varier, M.; Prakash, A.; Prasad, D. Improved Tapaswini having four BB resistance genes pyramided with six genes/QTLs, resistance/ tolerance to biotic and abiotic stresses in rice. Sci. Rep. 2018, 8, 2413. [Google Scholar] [CrossRef]
- Ellur, R.K.; Khanna, A.; Yadav, A.; Pathania, S.; Rajashekar, H.; Singh, V.K.; Gopala Krishnan, S.; Bhowmicka, P.K.; Nagaraj, M.; Vinod, K.K.; et al. Improvement of basmati rice varieties for resistance to blast and bacterial blight diseases using marker assisted back-cross breeding. Plant Sci. 2016, 242, 330–341. [Google Scholar] [CrossRef]
- Dokku, P.; Das, K.M.; Rao, G.J.N. Pyramiding of four resistance genes of bacterial blight in Tapaswini, an elite rice cultivar, through marker-assisted selection. Euphytica 2013, 192, 87–96. [Google Scholar] [CrossRef]
- Suh, J.P.; Jeung, J.U.; Noh, T.H.; Cho, Y.C.; Park, S.H.; Park, H.S.; Shin, M.S.; Kim, C.K.; Jena, K.K. Development of breeding lines with three pyramided resistance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice 2013, 6, 5. [Google Scholar] [CrossRef]
- Yamada, S.; Katsuhara, M.W.; Michalowski, C.B.; Bohnert, H.J. A family or transcripts encoding MIP: Homologues tissue specificity of expression under salt-stress in Masembryanthemum crystallinum. Plant Cell 1995, 7, 1129–1142. [Google Scholar]
- Yoshimura, S.; Yoshimura, A.; Iwata, N.; McCouch, S.R.; Abenes, M.L.; Baraoidan, M.R.; Mew, T.W.; Nelson, R.J. Tagging and combining of bacterial blight resistance genes in rice using RAPD and RFLP markers. Mol. Breed. 1995, 1, 375–387. [Google Scholar] [CrossRef]
- Stafford, H.A.; Vennesland, B. Alcohol dehydrogenase of wheat germ. Arch. Biochem. Biop. 1953, 44, 404–414. [Google Scholar] [CrossRef]
- Toojinda, T.; Tragoonrung, S.; Vanavichit, A.; Siangliw, J.L.; Pa-In, N.; Jantaboon, J.; Siangliw, M.; Fukai, S. Molecular breeding for rainfed lowland rice in the Mekong region. Plant Prod. Sci. 2005, 8, 330–333. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Chakraborti, M.; Chakraborty, K.; Behera, L.; Meher, J.; Subudhi, H.N.; Mishra, S.K.; Pandit, E.; Reddy, J.N. Genetic improvement of rainfed shallowlowland rice for higher yield and climate resilience. In Rice Research for Productivity, Profitability and Climate Resilience, 1st ed.; NRRI: Cuttack, India, 2018; pp. 107–121. [Google Scholar]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.K.; Laha, G.S.; Reddy, G.A.; Rani, S.N.; Sharma, N.P.; Sonti, R.V. Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 2008, 160, 411–422. [Google Scholar] [CrossRef]
- Nandakumar, N.; Singh, A.K.; Sharma, R.K.; Mohapatra, T.; Prabhu, K.V.; Zaman, F.U. Molecular fingerprinting of hybrids and assessment of genetic purity of hybrid seeds in rice using microsatellite markers. Euphytica 2004, 136, 257–264. [Google Scholar] [CrossRef]
- Dutta, S.S.; Divya, D.; Durga, R.C.V.; Reddy, D.T.; Visalakshmi, V.; Cheralu, C. Characterization of gall midge resistant rice genotypes using resistance gene specific markers. J. Exp. Biol. Agric. Sci. 2014, 2, 339–446. [Google Scholar]
- Kauffman, H.E.; Reddy, A.; Hsieh, S.P.Y.; Merca, S.D. An improved technique for evaluating resistance of varieties to Xanthomonas oryzae pv. oryzae. Plant Dis. Rep. 1973, 57, 537–541. [Google Scholar]
- Raskin, I.; Kende, H. Role of gibberellin in the growth response of submerged deep water rice. Plant Physiol. 1985, 76, 947–950. [Google Scholar] [CrossRef]
- Septiningsih, E.M.; Pamplona, A.M.; Sanchez, D.L.; Neeraja, C.N.; Vergara, G.V.; Heuer, S.; Ismail, A.M.; Mackill, D.J. Development of submergence tolerant rice cultivars: The Sub1 locus and beyond. Ann. Bot. 2009, 103, 151–160. [Google Scholar] [CrossRef]
- Perrier, X.; Flori, A.; Bonnot, F. Data analysis methods. In Genetic Diversity of Cultivated Tropical Plants; Hamon, P., Seguin, M., Perrier, X., Glaszmann, J.C., Eds.; Enfield, Science Publishers: Montpellier, France, 2003; pp. 43–76. [Google Scholar]
- IRRI. Injuries caused by diseases. In Standard Evaluation System for Rice (SES), 5th ed.; International Rice Research Institute: Los Baños, Philippines, 2013; pp. 18–27. [Google Scholar]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- SSR Markers. Available online: https://www.gramene.org/SSR (accessed on 27 July 2019).
- Krishnaveni, S.; Balasubramanian, T.; Sadasivam, S. Sugar distribution in sweet stalk sorghum. Food Chem. 1984, 15, 229–232. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.; Hamilton, J.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Witham, F.H.; Blaydes, D.F.; Devlin, R.M. Experiments in Plant Physiology; Van Nostrand Reinhold: New York, NY, USA, 1971. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Gnanamanickam, S.S.; Brindha, P.V.; Narayanan, N.N.; Vasudevan, P.; Kavitha, S. An overview of bacterial blight disease of rice and strategies for its management. Curr. Sci. 1999, 77, 1435–1443. [Google Scholar]
- Chen, X.; Temnykh, S.; Xu, Y.; Cho, Y.G.; McCouch, S.R. Development of a microsatellite framework map providing genome wide coverage in rice (Oryza sativa L.). Theor. Appl. Genet. 1997, 95, 553–567. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Khush, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.; Singh, S.P.; Ellur, R.K.; Singh, D.; Krishnan, S.G.; Bhowmick, P.K.; Nagarajan, M.; Vinod, K.K.; Singh, U.D.; et al. Marker-assisted simultaneous but stepwise backcross breeding for pyramiding blast resistance genes Pi2 and Pi54 into an elite basmati rice restorer line PRR78. Plant Breed. 2013, 132, 486–495. [Google Scholar]
- Yazid, S.N.; Ahmad, K.; Razak, M.S.F.A.; Rahman, Z.A.; Ramachandran, K.; Mohamad, S.N.A.; Ghaffar, M.B.A. Introgression of bacterial leaf blight (BLB) resistant gene, Xa7 into MARDI elite variety, MR219 by marker assisted backcrossing (MABC) approach. Braz. J. Biol. 2021, 84, e248359. [Google Scholar] [CrossRef]
- Berloo, R.V. GGT 2.0: Versatile software for visualization and analysis of genetic data. J. Hered. 2008, 99, 232–236. [Google Scholar] [CrossRef]
- Fahy, P.C.; Persley, G.J. Plant Bacterial Diseases: A Diagnostic Guide; Academic Press: New York, NY, USA, 1983; p. 393. [Google Scholar]
- IRRI. International Rice Research Institute, Rice Fact Sheet, Bacterial Blight. March 2010. Available online: https://download.ceris.purdue.edu/file/1503 (accessed on 22 July 2022).
- Mondal, K.K.; Meena, B.R.; Junaid, A.; Verma, G.; Mani, C.; Majumder, D.; Khicher, M.; Kumar, S.; Banik, S. Pathotyping and genetic screening of type III effectors in Indian strains of Xanthomonas oryzae pv. oryzae causing bacterial leaf blight of rice. Physiol. Mol. Plant Pathol. 2014, 86, 98–106. [Google Scholar] [CrossRef]
- Bhaduri, D.; Chakraborty, K.; Nayak, A.K.; Shahid, M.; Tripathi, R.; Behera, R.; Singh, S.; Srivastava, A.K. Alteration in plant spacing improves submergence tolerance in Sub1 and non-Sub1 rice (cv. IR64) by better light interception and effective carbohydrate utilisation under stress. Funct. Plant Biol. 2020, 47, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. 2020. R Package Version 1.0.7. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 10 December 2021).
- Hsu, Y.C.; Chiu, C.H.; Yap, R.; Tseng, U.C.; Wu, Y.P. Pyramiding bacterial blight resistance genes in Tainung82 for broad-spectrum resistance using marker-assisted selection. Inter. J. Mol. Sci. 2020, 21, 1281. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Fukao, F.; Ronald, P.; Ismail, A.; Heuer, S.; Mackill, D. Submergence tolerant rice: Sub1’s journey from landrace to modern cultivar. Rice 2010, 3, 138–147. [Google Scholar] [CrossRef]
- Singh, A.K.; Gopalakrishnan, S.; Singh, V.P.; Prabhu, K.V.; Mohapatra, T.; Singh, N.K.; Sharma, T.R.; Nagarajan, M.; Vinod, K.K.; Singh, D.; et al. Marker assisted selection: A paradigm shift in Basmati breeding. Indian J. Genet. 2011, 71, 120–128. [Google Scholar]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Flooding stress: Acclimations and genetic diversity. Ann. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [PubMed]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Hasan, M.M.; Oladosu, Y.A.; Magaji, U.G.; Akos, I.; Olalekan, K.K. Bacterial leaf blight resistance in rice: A review of conventional breeding to molecular approach. Mol. Biol. Rep. 2019, 46, 1519–1532. [Google Scholar] [CrossRef]
- Khanna, A.; Sharma, V.; Ellur, R.K.; Shikari, A.B.; Gopalakrishnan, S.; Singh, U.D.; Prakash, G.; Sharma, T.R.; Rathour, R.; Variar, M.; et al. Development and evaluation of near-isogenic lines for major blast resistance gene(s) in Basmati rice. Theor. Appl. Genet. 2015, 128, 1243–1259. [Google Scholar] [CrossRef]
- Balachiranjeevi, C.; Bhaskar, N.S.; Abhilash, V.; Akanksha, S.; Viraktamath, B.C.; Madhav, M.S.; Hariprasad, A.S.; Laha, G.S.; Prasad, M.S.; Balachandran, S.; et al. Marker-assisted introgression of bacterial blight and blast resistance into DRR17B, an elite, fine-grain type maintainer line of rice. Mol. Breed. 2015, 35, 151. [Google Scholar] [CrossRef]
- Singh, G.P.; Singh, S.R.; Singh, R.V.; Singh, R.M. Variation and qualitative losses caused by bacterial blight in different rice varieties. Indian Phytopathol. 1997, 30, 180–185. [Google Scholar]
- Singh, S.; Mackill, D.J.; Ismail, A.M. Responses of Sub1 rice introgression lines to submergence in the field: Yield and grain quality. Field Crops Res. 2009, 113, 12–23. [Google Scholar] [CrossRef]
- Cobb, J.N.; Biswas, P.S.; Platten, J.D. Back to the future: Revisiting MAS as a tool for modern plant breeding. Theor. Appl. Genet. 2019, 132, 647–667. [Google Scholar] [CrossRef]
- Cheema, K.; Grewal, N.; Vikal, Y.; Sharma, R.; Lore, J.S.; Das, A.; Bhatia, D.; Mahajan, R.; Gupta, V.; Bharaj, T.S.; et al. A novel bacterial blight resistant gene from Oryza nivara mapped to 38 kb region on chromosome 4L and transferred to Oryza sativa L. Genet. Res. Camb. 2008, 90, 397–407. [Google Scholar] [CrossRef]
- Ikmal, A.M.; Noraziyah, A.A.S.; Wickneswari, R.; Amira, I.; Ellina, Z.P.D. Interaction of submergence tolerance and drought yield QTLs (Sub1 and qDTYs) enhances morpho-physiological traits and survival of rice (Oryza sativa L.) under submergence. Ann. Appl Biol. 2021, 178, 355–366. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.; Singh, P.; Ellur, R.K.; Choudhary, V.; Sarkel, S.; Singh, D.; Gopalakrishnan, S.; Nagarajan, M.; Vinod, K.K.; et al. Incorporation of blast resistance into “PRR78”, an elite basmati rice restorer line, through marker assisted backcross breeding. Field Crops Res. 2012, 128, 8–16. [Google Scholar] [CrossRef]
- Sarkar, R.K.; Reddy, J.N.; Sharma, S.G.; Ismail, A.M. Physiological basis of submergence tolerance in rice and implications for crop improvement. Cur. Sci. 2006, 91, 899–906. [Google Scholar]
- Adak, M.K.; Gupta, D.K.D. Prolonged waterlogging on photosynthesis and related characters in rice. Indian J. Plant Physiol. 2000, 5, 380–382. [Google Scholar]
- Fukao, T.; Yeung, E.; Bailey-Serres, J. The submergence tolerance gene SUB1A delays leaf senescence under prolonged darkness through hormonal regulation in rice. Plant Physiol. 2012, 4, 1795–1807. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Nanda, B.B.; Swami, S.G.; Nayak, B.B. Influence of nitrogen on the physico-chemical characteristics of rice grain. Oryza 1971, 891, 87–97. [Google Scholar]
- Ashraf, M. Relationships between leaf gas exchange characteristics and growth of differently adapted populations of blue panicgrass panicum antidotale Retz. under salinity or water logging. Plant Sci. 2003, 165, 69–75. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Deka, M.; Baruah, K.K. Comparable studies of rainfed upland winter rice (Oryza sativa) cultivars for drought tolerance. Indian J. Agril. Sci. 2000, 70, 135–139. [Google Scholar]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Verma, R.L.; Singh, R.S.; Singh, R.P.; Singh, H.B.; Arsode, P.; Kumar, M.; Singh, P.K. Biotic stress management in rice (Oryza sativa L.) through conventional and molecular approaches. In New Frontiers in Stress Management for Durable Agriculture; Springer: Singapore, 2020; pp. 609–644. [Google Scholar]
- Luo, F.L.; Chen, Y.; Huang, L.; Wang, A.; Zhang, M.X.; Yu, F.H. Shifting effects of physiological integration on performance of a clonal plant during submergence and de-submergence. Ann. Bot. 2014, 113, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Loreti, E.; Striker, G.G. Plant responses to hypoxia: Signaling and adaptation. Plants 2020, 9, 1704. [Google Scholar] [CrossRef] [PubMed]
- Joshi, E.; Kumar, D.; Lal, B.V.; Gautam, P.; Vyas, A.K. Management of direct seeded rice for enhanced resource—Use efficiency. Plant Knowl. J. 2013, 2, 119–134. [Google Scholar]
- Xu, K.; Xu, X.; Ronald, P.C.; Mackill, D.J. A high-resolution linkage map in the vicinity of the rice submergence tolerance locus Sub1. Mol. Gen. Genet. 2000, 263, 681–689. [Google Scholar] [CrossRef]
- Ismail, A.M.; Ella, E.S.; Vergara, G.V.; Mackill, D.J. Mechanisms associated with tolerance to flooding during germination and early seedling growth in rice (Oryza sativa). Ann. Bot. 2009, 103, 197–209. [Google Scholar] [CrossRef]
- Chen, C.T.; Kao, C.H. Osmotic stress and water stress have opposite effects on putrescine and proline production in excised rice leaves. Plant Growth Regul. 1993, 13, 197–202. [Google Scholar] [CrossRef]
- Caballero, J.L.; Verderzco, C.V.; Galam, J.; Jimenez, E.S.D. Proline accumulation as a symptom of water stress in maize: A tissue differentiation requirement. J. Exp. Bot. 2005, 39, 889–897. [Google Scholar] [CrossRef]
- Bradbury, L.M.T.; Fitzgerald, T.L.; Henry, R.J.; Jin, Q.S.; Waters, D.L.E. The gene for fragrance in rice. Plant Biotechnol. J. 2005, 3, 363–370. [Google Scholar] [CrossRef]
- Juliano, B.O. A simplified assay for milled rice amylose. Cereal Sci. Today 1971, 16, 340–360. [Google Scholar]
- Kumar, M.; Singh, R.P.; Singh, O.N.; Singh, P.; Arsode, P.; Namrata; Chaudhary, M.; Jena, D.; Singh, V.; Rout, D.; et al. Genetic analysis for bacterial blight resistance in indica rice (Oryza sativa L.) cultivars. Oryza 2019, 56, 247–255. [Google Scholar] [CrossRef]
- Rahman, M.; Grover, A.; Peacock, W.J.; Dennis, E.S.; Ellis, M.H. Effects of manipulation of pyruvate decarboxylase and alcohol dehydrogenase levels on the submergence tolerance of rice. Aust. J. Plant Physiol. 2001, 28, 1231–1241. [Google Scholar] [CrossRef]
- Peng, H.; Chen, Z.; Fang, Z.; Zhou, J.; Xia, Z.; Gao, L.; Chen, L.; Li, L.; Li, T.; Zhai, W.; et al. Rice Xa21 primed genes and pathways that are critical for combating bacterial blight infection. Sci. Rep. 2015, 5, 12165. [Google Scholar] [CrossRef]
- Katara, J.L.; Verma, R.L.; Parida, M.; Ngangkham, U.; Molla, K.A.; Barbadikar, K.M.; Mukherjee, M.; Samantaray, S.; Ravi, N.R.; Singh, O.N.; et al. Differential expression of genes at panicle initiation and grain filling stages implied in heterosis of rice hybrids. Int. J. Mol. Sci. 2020, 21, 1080. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Pedigree | Special Features |
|---|---|---|
| Hasanta | OR 1206-26-2 x OR 1534-129 | Medium late duration, semi-dwarf, high yielding with small bold grains, MR to leaf folder, leaf blast, sheath blight and bacterial leaf blast for Odisha |
| Swarna Sub1 | Swarna 3/IR 49830-7-1-2-3 | High yielding (45–55 q/ha), semi-dwarf, medium-late (140–145 days) duration, medium slender grains, >65% HRR, tolerant to complete submergence between 15–17 days |
| IRBB66 | IR 24 NIL | Medium duration (135 days), semi-dwarf (95–100 cm), medium slender grain, medium amylose, bacterial blight resistant |
| Gene/QTL | Chr. No. | Marker/Primer | Physical Position (Mb) | Type | Marker/Primer/Sequence | Reference |
|---|---|---|---|---|---|---|
| Sub1 | 9 | Sub1bc2 | 0.0–4.80 | FS | Forward-5′ aaaacaatggttccatacgagac3′ Reverse-5′gcctatcaatgcgtgctctt3′ | [2] |
| RM8300 | 1.0 | RS | Forward-5′gctagtgcagggttgacaca3′ Reverse-5′ctctggccgtttcatggtat3′ | |||
| A203 | 2.0 | RS | Forward-5′cttcttgctcaacgacaacg3′ Reverse-5′aggctccagatgtccatgtc3′ | |||
| C173 | 4.0 | RS | Forward-5′aacgccaagaccaacttcc3′ Reverse-5′aggaggctgtccatcaggt3′ | |||
| xa5 | 5 | RM122 | 1.65 | FS | Forward-5′gcactgcaaccatcaatgaatc3′ Reverse-5′cctaggagaaactagccgtcca3′ | [65] |
| RM17941 | 3.40 | RS | Forward-5′gcctcgaagaaccagtagaacagc3′ Reverse-5′cttgtcttctcctcctcctgtgc3′ | |||
| xa13 | 8 | xa13prom | 26.81 | FS | Forward-5′ggccatggctcagtgtttat3′ Reverse-5′gagctccagctctccaaatg3′ | [13,44,55] |
| RM23356 | 24.20 | RS | Forward-5′gcctccaacagatctcctatctgg3′ Reverse-5′tttggcgctaatgagagattgg3′ | |||
| RM22914 | 30.90 | RS | Forward-5′ccaatcattaacccctgagc3′ Reverse-5′gccttcatgcttcagaagac3′ | |||
| Xa21 | 11 | pTA248 | 22.60 | FS | Forward-5′agacgcggaagggtggttcccgga3′ Reverse-5′agacgcggtaatcgaaagatgaaa3′ | [23] |
| RM26969 | 21.50 | RS | Forward-5′ctcacacttgcaacatcctagc3′ Reverse-5′aaggctctagttggtgaagacc3′ |
| Generation | Total Plants Raised | Gene Positive Plants | Plants Advanced | % RPG Recovery | Selection Criteria |
|---|---|---|---|---|---|
| F1s | 87 (P1) & 47 (P2) | 78 & 38 | 5 of each P1&P2 | * | Hybridity testing with gene linked markers |
| IC1F1 | 532 | 11 | 10 | * | Hybridity testing with gene linked markers |
| BC1F1 | 678 | 8 | 6 | 68.43–80.08 | FS, BS, phenomics and bioassay |
| BC2F1 | 465 | 7 | 7 | 79.65–89.40 | FS, BS, phenomics and bioassay |
| BC2F2 | 847 | 13 | 3 | 91.35–95.19 | FS, BS, phenomics and bioassay |
| BC2F3 | 13 families (each 150 plants) | 13 families (28 plants) | 13 NILs | >93.75 | Phenomics and screening/bioassay analysis |
| BC2F4 | 13 NILs | 13 NILs | 13 NILs | >93.75 | Phenomics and screening/bioassay analysis |
| Pyramided Lines (NILs) | Gene Combination | Disease Lesion Length (LL) in cm (Mean ± Standard Error) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Xa17 | Xa7 | xa2 | xb7 | xc4 | xd1 | xa1 | xa5 | MLL | Disease Reaction | ||
| HS 110-224-197-10-2-2 | Sub1 + Xa21 + xa13 + xa5 | 1.98 ± 0.67 | 2.25 ± 0.23 | 2.08 ± 0.19 | 2.36 ± 0.39 | 2.8 ± 0.87 | 2.5 ± 0.13 | 2.48 ± 0.29 | 2.32 ± 0.58 | 2.35 | R |
| HS 110-224-197-10-2-5 | Sub1 + Xa21 + xa13 + xa5 | 2.35 ± 0.33 | 2.68 ± 0.94 | 2.12 ± 0.28 | 2.56 ± 0.13 | 3.05 ± 0.74 | 2.26 ± 0.42 | 2.62 ± 1.09 | 2.4 ± 0.31 | 2.51 | R |
| HS 110-224-197-10-2-36 | Sub1 + Xa21 + xa13 + xa5 | 3.12 ± 0.92 | 2.3 ± 0.54 | 2.8 ± 0.91 | 2.9 ± 0.48 | 2.8 ± 0.76 | 2.6 ± 0.82 | 2.55 ± 0.86 | 2.10 ± 0.74 | 2.65 | R |
| HS 110-224-197-10-2-41 | Sub1 + Xa21 + xa13 + xa5 | 2.8 ± 0.32 | 2.08 ± 0.67 | 2.28 ± 1.18 | 3.3 ± 0.35 | 2.7 ± 0.69 | 2.10 ± 0.39 | 3.08 ± 1.11 | 2.20 ± 0.62 | 2.57 | R |
| HS 110-224-197-10-2-68 | Sub1 + Xa21 + xa13 + xa5 | 2.6 ± 0.46 | 2.9 ± 0.33 | 2.4 ± 0.48 | 2.17 ± 0.43 | 2.82 ± 0.12 | 2.62 ± 0.22 | 2.36 ± 1.00 | 3.42 ± 0.53 | 2.66 | R |
| HS 232-411-391-809-63-8-10 | Sub1 + Xa21 + xa13 + xa5 | 1.29 ± 0.12 | 1.98 ± 0.65 | 2.11 ± 0.34 | 3.16 ± 0.54 | 2.36 ± 0.21 | 1.85 ± 0.39 | 3.22 ± 0.64 | 2.81 ± 0.19 | 2.35 | R |
| HS 232-411-391-809-63-8-18 | Sub1 + Xa21 + xa13 + xa5 | 2.2 ± 0.92 | 1.85 ± 0.42 | 2.14 ± 0.94 | 1.68 ± 0.85 | 3.54 ± 0.67 | 1.75 ± 0.38 | 3.42 ± 1.20 | 2.07 ± 0.66 | 2.33 | R |
| HS 232-411-391-809-63-8-20 | Sub1 + Xa21 + xa13 + xa5 | 3.0 ± 0.59 | 3.72 ± 0.27 | 2.26 ± 0.87 | 2.44 ± 0.72 | 1.98 ± 0.22 | 3.40 ± 0.65 | 3.04 ± 1.22 | 2.8 ± 0.77 | 2.83 | R |
| HS 232-411-391-809-63-8-37 | Sub1 + Xa21 + xa13 + xa5 | 1.99 ± 0.32 | 2.48 ± 0.47 | 2.33 ± 1.15 | 3.5 ± 0.28 | 1.95 ± 1.02 | 2.61 ± 0.82 | 2.42 ± 0.36 | 3.11 ± 0.62 | 2.55 | R |
| HS 232-411-391-809-63-1 | Sub1 + Xa21 + xa13 + xa5 | 2.1 ± 1.24 | 3.55 ± 0.75 | 2.2 ± 0.63 | 3.02 ± 0.22 | 3.6 ± 1.24 | 2.6 ± 0.46 | 2.24 ± 0.47 | 3.00 ± 0.56 | 2.79 | R |
| HS 232-411-391-809-63-8 | Sub1 + Xa21 + xa13 + xa5 | 2.3 ± 0.69 | 3.2 ± 0.46 | 2.3 ± 0.53 | 4.2 ± 0.64 | 2.82 ± 0.33 | 2.75 ± 1.12 | 2.08 ± 0.18 | 2.98 ± 0.39 | 2.83 | R |
| HS 232-411-391-809-63-63 | Sub1 + Xa21 + xa13 + xa5 | 3.07 ± 0.62 | 3.2 ±0.63 | 3.01 ± 0.45 | 2.25 ± 0.24 | 2.44 ± 0.52 | 2.26 ± 0.37 | 2.8 ± 0.19 | 2.09 ± 0.12 | 2.64 | R |
| HS 232-411-391-809-63-81 | Sub1 + Xa21 + xa13 + xa5 | 2.0 ± 0.33 | 2.1 ± 1.14 | 1.68 ± 0.15 | 1.84 ± 0.42 | 2.34 ± 0.51 | 2.44 ± 0.86 | 2.24 ± 0.47 | 1.64 ± 0.39 | 2.04 | R |
| IRBB66 | Xa21 + xa13 + xa5 + Xa4 | 1.7 ± 0.13 | 1.67 ± 0.26 | 2.2 ± 1.08 | 2.15 ± 0.26 | 2.68 ± 0.23 | 2.42 ± 0.41 | 1.9 ± 0.34 | 1.68 ± 0.21 | 2.05 | R |
| Swarna Sub1 (donor) | Sub1 + xa21 + Xa13 + Xa5 | 9.25 ± 1.35 | 12.5 ± 1.28 | 11.68 ± 1.54 | 10.98 ± 0.98 | 13.74 ± 1.46 | 12.45 ± 1.28 | 14.25 ± 1.35 | 10.80 ± 1.16 | 11.96 | S |
| Hasanta (recipient) | sub1 + xa21 + Xa13 + Xa5 | 12.34 ± 1.08 | 14.82 ± 1.38 | 10.86 ± 1.25 | 13.65 ± 1.32 | 10.87 ± 1.27 | 9.95 ± 1.74 | 15.44 ± 1.95 | 12.87 ± 1.32 | 12.60 | S |
| Trait | Environment | RP | DP | NILs | RUs | CV | LSD | |
|---|---|---|---|---|---|---|---|---|
| Mean | Range | |||||||
| DFF | NS | 112.6 | 114.5 | 112.7 | 110.2–115.2 | −4.34 | 10.25 | 2.41 |
| S | 118.5 | 115.2 | 117.6 | 112.5–122.5 | 8.15 | 1.83 | ||
| DM | NS | 148.3 | 146.5 | 144.7 | 140.2–148.4 | −2.14 | 8.65 | 2.33 |
| S | 150.2 | 146.5 | 147.8 | 138.5–155.2 | 18.32 | 0.72 | ||
| PH (cm) | NS | 111.45 | 96.35 | 112.9 | 109.5–116.5 | 2.12 | 9.08 | 2.84 |
| S | 108.22 | 93.12 | 110.5 | 106.5–114.5 | 7.08 | 4.42 | ||
| FLL (cm) | NS | 24.5 | 19.3 | 24.8 | 22.9–26.6 | 2.41 | 8.65 | 3.02 |
| S | 24.05 | 18.8 | 24.2 | 19.4–28.15 | 11.24 | 8.42 | ||
| FLW (cm) | NS | 1.21 | 1.05 | 1.2 | 1.12–1.34 | 0.83 | 10.25 | 0.08 |
| S | 1.19 | 0.99 | 1.19 | 1.13–1.28 | 8.05 | 0.25 | ||
| NETPP | NS | 12.55 | 11.75 | 11.0 | 10.05–12.5 | 13.63 | 9.40 | 1.95 |
| S | 5.75 | 10.95 | 9.5 | 8.2–11.7 | 12.87 | 1.3 | ||
| PL (cm) | NS | 20.97 | 21.17 | 21.5 | 19.3–24.8 | −0.93 | 8.65 | 1.7 |
| S | 20.42 | 16.62 | 21.7 | 19.2–24.5 | 11.62 | 2.4 | ||
| PW (g) | NS | 10.4 | 9.55 | 9.0 | 7.3–10.3 | 2.22 | 10.25 | 1.84 |
| S | 9.1 | 8.2 | 8.8 | 6.7–9.8 | 13.16 | 2.64 | ||
| SF% | NS | 81.2 | 82.6 | 81.1 | 75.8–86.2 | 1.72 | 3.90 | 11.90 |
| S | 77.3 | 84.9 | 79.7 | 76.2–82.2 | 6.82 | 6.18 | ||
| TW | NS | 20.5 | 18.8 | 20.4 | 19.3–22.4 | −1.47 | 7.71 | 2.04 |
| S | 20.4 | 18.7 | 20.7 | 19.1–23.5 | 11.08 | 1.24 | ||
| GY (q/ha) | NS | 78.1 | 81.6 | 80.9 | 62.4–89.4 | 33.12 | 6.03 | 1.65 |
| S | 5.5 | 52.8 | 54.1 | 40.13–63.1 | 12.7 | 1.40 | ||
| KL (mm) | NS | 4.6 | 4.5 | 4.5 | 4.0–5.1 | −2.22 | 10.2 | 0.03 |
| S | 4.6 | 4.5 | 4.6 | 4.2–5.1 | 10.08 | 0.03 | ||
| KW (mm) | NS | 1.87 | 1.50 | 1.77 | 1.54–1.88 | −1.12 | 7.59 | 0.2 |
| S | 1.87 | 1.50 | 1.79 | 1.65–1.88 | 8.02 | 0.2 | ||
| KLBR | NS | 2.46 | 3.00 | 2.56 | 2.21–3.30 | 0.0 | 8.25 | 0.1 |
| S | 2.47 | 3.01 | 2.56 | 2.34–3.09 | 5.18 | 0.09 | ||
| HRR% | NS | 64.3 | 64.5 | 56.2 | 43.9–62.04 | −3.38 | 4.65 | 0.8 |
| S | 59.8 | 60.02 | 58.1 | 48.2–64.6 | 4.72 | 0.6 | ||
| AC% | NS | 23.06 | 22.2 | 22.4 | 20.08–25.6 | −3.12 | 3.65 | 0.14 |
| S | 22.6 | 23.08 | 23.1 | 21.6–25.1 | 3.06 | 0.06 | ||
| %Survival | NS | 86.0 | 96.0 | 94.0 | 88.0–99.0 | 10.53 | 6.46 | 3.93 |
| S | 02.0 | 89.0 | 84.1 | 76.0–91.0 | 18.45 | 8.22 | ||
| Source | DF | Mean Sum of Squares | |||
|---|---|---|---|---|---|
| ADH | TSS | PA | TCC | ||
| Replications | 2 | 39.017 ** | 0.00019 * | 0.0023 * | 2.424 * |
| Genotype (A) | 14 | 269.125 ** | 0.249 ** | 0.165 ** | 4.234 ** |
| Treatment (B) | 3 | 26.491 ** | 0.0622 ** | 0.0226 ** | 2.571 ** |
| AxB | 42 | 22.230 ** | 0.172 ** | 0.0422 ** | 3.121 ** |
| Error | 42 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das, G.; Pradhan, B.; Bastia, D.; Samantaray, S.; Jena, D.; Rout, D.; Arsode, P.B.; Singh, V.; Mukherjee, A.K.; Mohan, C.; et al. Pyramiding Submergence Tolerance and Three Bacterial Blight Resistance Genes in Popular Rice Variety Hasanta through Marker-Assisted Backcross Breeding. Agriculture 2022, 12, 1815. https://doi.org/10.3390/agriculture12111815
Das G, Pradhan B, Bastia D, Samantaray S, Jena D, Rout D, Arsode PB, Singh V, Mukherjee AK, Mohan C, et al. Pyramiding Submergence Tolerance and Three Bacterial Blight Resistance Genes in Popular Rice Variety Hasanta through Marker-Assisted Backcross Breeding. Agriculture. 2022; 12(11):1815. https://doi.org/10.3390/agriculture12111815
Chicago/Turabian StyleDas, Goutam, Banshidhar Pradhan, Debendranath Bastia, Sanghamitra Samantaray, Debarchana Jena, Diptibala Rout, Paduranga Bhagwan Arsode, Vineeta Singh, Arup Kumar Mukherjee, Chander Mohan, and et al. 2022. "Pyramiding Submergence Tolerance and Three Bacterial Blight Resistance Genes in Popular Rice Variety Hasanta through Marker-Assisted Backcross Breeding" Agriculture 12, no. 11: 1815. https://doi.org/10.3390/agriculture12111815
APA StyleDas, G., Pradhan, B., Bastia, D., Samantaray, S., Jena, D., Rout, D., Arsode, P. B., Singh, V., Mukherjee, A. K., Mohan, C., & Verma, R. (2022). Pyramiding Submergence Tolerance and Three Bacterial Blight Resistance Genes in Popular Rice Variety Hasanta through Marker-Assisted Backcross Breeding. Agriculture, 12(11), 1815. https://doi.org/10.3390/agriculture12111815