

The Combinations of White, Blue, and UV-A Light Provided by Supplementary Light-Emitting Diodes Promoted the Quality of Greenhouse-Grown Cucumber Seedlings

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Treatment Design
2.3. Growth Measurements
2.3.1. Plant Morphology and Growth Traits
2.3.2. Determinations of Photosynthetic Performance
2.3.3. Measurement of Root Activity, Stem Firmness, and Cellulose Content of Cucumber Seedling
2.3.4. Measurement of Hormone Content of Cucumber Seedlings
2.3.5. Supplementary Light Use Efficiency
2.4. Statistical Analysis
3. Results
3.1. Impacts of Supplementary Light on Morphological Performances of Greenhouse-Grown Cucumber Seedlings
3.2. Impacts of Supplementary Light on Photosynthetic Performances of Cucumber Seedlings Cultivated in the Greenhouse
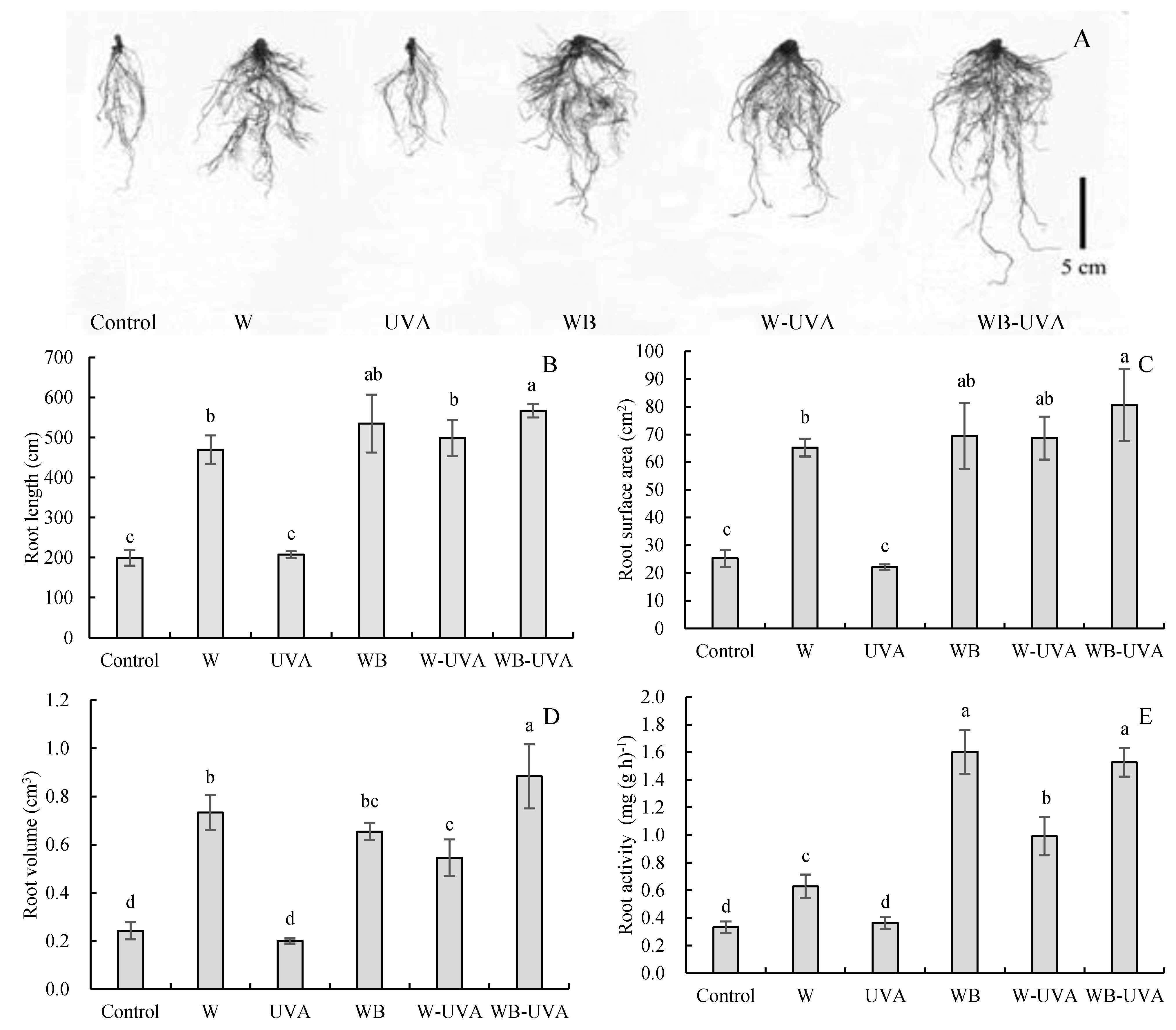
3.3. Impacts of Supplementary Light on Growth Characteristics and Root Architecture on Greenhouse-Grown Cucumber Seedlings
3.4. Influences of Supplementary Light on Stem Firmness and Cellulose Content of Greenhouse-Grown Cucumber Seedlings
3.5. Hormone Content of Cucumber Seedlings Cultivated in the Greenhouse as Affected by Supplementary Light
3.6. Supplementary Light Use Efficiency of Greenhouse-Grown Cucumber Seedlings
3.7. Heat Map Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. 2022. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 7 July 2022).
- Chang, C.L.; Chang, K.P. The growth response of leaf lettuce at different stages to multiple wavelength-band light-emitting diode lighting. Sci. Hortic. 2014, 179, 78–84. [Google Scholar] [CrossRef]
- Nakano, A.; Ohtake, N.; Yoneda, T.; Shinoda, A. Effects of LED alternating irradiation on tomato seedling quality and root development after transplanting. Root Res. 2017, 26, 3–9. [Google Scholar] [CrossRef]
- Ji, F.; Wei, S.Q.; Liu, N.; Xu, L.J.; Yang, P. Growth of cucumber seedlings in different varieties as affected by light environment. Int. J. Agric. Biol. Eng. 2020, 13, 73–78. [Google Scholar] [CrossRef]
- Özdemir, Y.; Demirel, H.; Yildirim, Y.E. Developing of DLI (Daily Light Integral) and spectrum control systems for scientific cultivation in agriculture. Int. J. Agric. For. Sci. 2016, 1, 14–17. [Google Scholar]
- Pennisi, G.; Pistillo, A.; Orsini, F.; Cellini, A.; Spinelli, F.; Nicola, S.; Juan, A.; Crepaldi, F.A.; Gianquinto, G.; Marcelis, L.F.M. Optimal light intensity for sustainable water and energy use in indoor cultivation of lettuce and basil under red and blue LEDs. Sci. Hortic. 2020, 272, 109508. [Google Scholar] [CrossRef]
- Dou, H.J.; Niu, G.; Gu, M.M.; Masabni, J.G. Responses of sweet basil to different daily light integrals in photosynthesis, morphology, yield, and nutritional quality. HortScience 2018, 53, 496–503. [Google Scholar] [CrossRef]
- Gao, M.; He, R.; Shi, R.; Zhang, Y.; Song, S.; Su, W.; Liu, H. Differential Effects of Low Light Intensity on Broccoli Microgreens Growth and Phytochemicals. Agronomy 2021, 11, 537. [Google Scholar] [CrossRef]
- Ke, X.; Yoshida, H.; Hikosaka, S.; Goto, E. Optimization of photosynthetic photon flux density and light quality for increasing radiation-use efficiency in dwarf tomato under LED light at the vegetative growth stage. Plants 2022, 11, 121. [Google Scholar] [CrossRef]
- Kelly, N.; Choe, D.; Meng, Q.W.; Runkle, E.S. Promotion of lettuce growth under an increasing daily light integral depends on the combination of the photosynthetic photon flux density and photoperiod. Sci. Hortic. 2020, 272, 109565. [Google Scholar] [CrossRef]
- Cui, J.; Song, S.; Yu, J.; Liu, H. Effect of daily light integral on cucumber plug seedlings in artificial light plant factory. Horticulturae 2021, 7, 139. [Google Scholar] [CrossRef]
- Wang, Y.; Chu, Y.; Wan, Z.; Zhang, G.; Liu, L.; Yan, Z. Root architecture, growth and photon yield of cucumber seedlings as influenced by daily light integral at different stages in the closed transplant production system. Horticulturae 2021, 7, 328. [Google Scholar] [CrossRef]
- Yan, Z.N.; Wang, L.; Wang, Y.; Chu, Y.; Lin, D.; Yang, Y. Morphological and physiological properties of greenhouse-grown cucumber seedlings as influenced by supplementary light-emitting diodes with same daily light integral. Horticulturae 2021, 7, 361. [Google Scholar] [CrossRef]
- Dyśko, J.; Kaniszewski, S. Effects of LED and HPS lighting on the growth, seedling morphology and yield of greenhouse tomatoes and cucumbers. Hortic. Sci. 2021, 48, 22–29. [Google Scholar] [CrossRef]
- Sobczak, A.; Sujkowska-Rybkowska, M.; Gajc-Wolska, J.; Kowalczyk, W.; Borucki, W.; Kalaji, H.M.; Kowalczyk, K. Photosynthetic efficiency and anatomical structure of pepper leaf (Capsicum annuum L.) transplants grown under high-pressure sodium (HPS) and light-emitting diode (LED) supplementary lighting systems. Plants 2021, 10, 1975. [Google Scholar] [CrossRef] [PubMed]
- Hogewoning, S.W.; Wientjes, E.; Douwstra, P.; Trouwborst, G.; Van Ieperen, W.; Croce, R.; Harbinson, J. Photosynthetic quantum yield dynamics: From photosystems to leaves. Plant Cell 2012, 24, 1921–1935. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Hernandez, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LED. Environ. Exp. Bot. 2016, 121, 66–74. [Google Scholar] [CrossRef]
- Clavijo-Herrera, J.; van Santen, E.; Gómez, C. Growth, Water-use efficiency, stomatal conductance, and nitrogen uptake of two lettuce cultivars grown under different percentages of blue and red light. Horticulturae 2018, 16, 3–13. [Google Scholar] [CrossRef]
- Wang, J.; Wei, L.; Tong, Y.; Yang, Q.C. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef]
- Azad, M.O.K.; Kjaer, K.H.; Adnan, M.; Naznin, M.T.; Lim, J.D.; Sung, I.J.; Park, C.H.; Lim, Y.S. The evaluation of growth performance, photosynthetic capacity, and primary and secondary metabolite content of leaf lettuce grown under limited irradiation of blue and red LED light in an urban plant factory. Agriculture 2020, 10, 28. [Google Scholar] [CrossRef]
- Yang, Z.C.; He, W.; Mou, S.T.; Wang, X.X.; Chen, D.Y.; Hu, X.T.; Chen, L.H.; Bai, J.Y. Plant growth and development of pepper seedlings under different photoperiods and photon flux ratios of red and blue LEDs. Trans. CSAE 2017, 33, 173–180. [Google Scholar]
- Liang, Y.; Kang, C.Q.; Kaiser, E.; Kuang, Y.; Yang, Q.C.; Li, T. Red/blue light ratios induce morphology and physiology alterations differently in cucumber and tomato. Sci. Hortic. 2021, 281, 109995. [Google Scholar] [CrossRef]
- Pennisi, G.; Blasioli, S.; Cellini, A.; Maia, L.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Unraveling the role of red:blue LED lights on resource use efficiency and nutritional properties of indoor grown sweet basil. Front. Plant Sci. 2019, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Hammock, H.A.; Kopsell, D.A.; Sams, C.E. Supplementary blue and red LED narrowband wavelengths improve biomass yield and nutrient uptake in hydroponically grown basil. HortScience 2020, 55, 1888–1897. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Spectral effects of light-emitting diodes on plant growth, visual color quality, and photosynthetic photon efficacy: White versus blue plus red radiation. PLoS ONE 2018, 13, e0202386. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.N.; He, D.X.; Niu, G.H.; Zhou, Q.; Qu, Y.H. Growth, nutritional quality, and energy use efficiency in two lettuce cultivars as influenced by white plus red versus red plus blue LEDs. Int. J. Agric. Biol. Eng. 2020, 13, 33–40. [Google Scholar] [CrossRef]
- Yan, Z.N.; He, D.X.; Niu, G.H.; Zhou, Q.; Qu, Y.H. Growth, nutritional quality and energy use efficiency of hydroponic lettuce as influenced by daily light integrals exposed to white versus white plus red light-emitting diodes. HortScience 2019, 54, 1737–1744. [Google Scholar] [CrossRef]
- Gao, W.; He, D.X.; Ji, F.; Zhang, S.; Zheng, J.F. Effects of daily light integral and LED spectrum on growth and nutritional quality of hydroponic spinach. Agronomy 2020, 10, 1082. [Google Scholar] [CrossRef]
- Zheng, J.F.; Peidian, G.; Ji, F.; He, D.X.; Yang, P. Growth and energy use efficiency of grafted tomato transplants as affected by LED light quality and photon flux density. Agriculture 2021, 11, 816. [Google Scholar] [CrossRef]
- Kalaitzoglou, P.; Taylor, C.; Calders, K.; Hogervorst, M.; Ieperen, W.V.; Harbinson, J.; Visser, P.D.; Nicole, C.C.S.; Marcelis, L.F.M. Unraveling the effects of blue light in an artificial solar background light on growth of tomato plants. Environ. Exp. Bot. 2021, 184, 104377. [Google Scholar] [CrossRef]
- Kaiser, E.; Ouzounis, T.; Giday, H.; Schipper, R.; Heuvelink, E.; Marcelis, L.F.M. Adding blue to red supplemental light increases biomass and yield of greenhousegrown tomatoes, but only to an optimum. Front. Plant Sci. 2019, 9, 2002. [Google Scholar] [CrossRef] [PubMed]
- Verdaguer, D.; Jansen, M.A.K.; Llorens, L.; Morales, L.O.; Neugart, S. UV-A radiation effects on higher plants: Exploring the known unknown. Plant Sci. 2017, 255, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Park, S.Y.; Oh, M.M. Supplemental radiation of ultraviolet-A light-emitting diode improves growth, antioxidant phenolics, and sugar alcohols of ice plant. Hortic. Environ. Biotechnol. 2021, 62, 559–570. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Elias, K.; Zhang, Y.Q.; Zou, J.; Bian, Z.H.; Yang, Q.C.; Li, T. UVA radiation promotes tomato growth through morphological adaptation leading to increased light interception. Environ. Exp. Bot. 2020, 176, 104073. [Google Scholar] [CrossRef]
- Kang, S.Y.; Zhang, Y.T.; Zhang, Y.Q.; Zou, J.; Yang, Q.C.; Li, T. Ultraviolet-A radiation stimulates growth of indoor cultivated tomato (Solanum lycopersicum) seedlings. HortScience 2018, 53, 1429–1433. [Google Scholar] [CrossRef]
- Yan, Z.N.; Wang, L.; Cheng, J.; Lin, D.; Yang, Y.J. Morphology, Growth and physiological traits of greenhouse-grown cucumber seedlings as affected by supplementary white and blue light-emitting diodes. In Proceedings of the 2021 International Symposium on Efficient Production of Value-Added Plants, Online, 27–28 December 2021; pp. 57–70. [Google Scholar]
- Han, S.Q.; Wang, X.F.; Wei, M.; Li, Y. Study of plug seedling index if sweet pepper and relationship between seedling index and characters. J. Shandong Agric. Univ. 2004, 35, 187–190. [Google Scholar]
- Li, H.S. Principle and Technology of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2003; pp. 119–120. [Google Scholar]
- Updegraff, D.M. Semi-micro determination of cellulose in biological materials. Anal. Biochem. 1969, 32, 420–424. [Google Scholar] [CrossRef]
- Aguilar, M.L.; Espadas, F.; Maust, B.; Sáenz, L. Endogenous cytokinin content in coconut palms affected by lethal yellowing. J. Plant Pathol. 2009, 91, 141–146. [Google Scholar]
- Wei, H.; Wang, M.W.; Jeong, B.R. Effect of supplementary lighting duration on growth and activity of antioxidant enzymes in grafted watermelon seedlings. Agronomy 2020, 10, 337. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Kobori, M.M.R.G.; Mello, S.D.C.; Freitas, I.S.D.; Silveira, F.F.; Alves, M.C.; Azevedo, R.A. Supplemental light with different blue and red ratios in the physiology, yield and quality of Impatiens. Sci. Hortic. 2022, 306, 111424. [Google Scholar] [CrossRef]
- Robson, T.M.; Klem, K.; Urban, O.; Jansen, M.A.K. Re-interpreting plant morphological responses to UV-B radiation. Plant Cell Environ. 2015, 38, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.J.; Rosenqvist, E.; Flygare, A.M.; Kalbina, I.; Teng, Y.W.; Marcel, A.K.; Jansen, A. UV-A light induces a robust and dwarfed phenotype in cucumber plants (Cucumis sativus L.) without affecting fruit yield. Sci. Hortic. 2020, 263, 109110. [Google Scholar] [CrossRef]
- Liu, B.; Yang, Z.; Gomez, A.; Liu, B.; Lin, C.; Oka, Y. Signaling mechanisms of plant cryptochromes in Arabidopsis thaliana. J. Plant Res. 2016, 129, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radaglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Caldwell, C.R.; Britz, S.J. Effect of supplemental ultraviolet radiation on the carotenoid and chlorophyll composition of greenhouse-grown leaf lettuce (Lactuca sativa L.) cultivars. J. Food Compos. Anal. 2006, 19, 637–644. [Google Scholar] [CrossRef]
- Štroch, M.; Meterová, Z.; Vrábl, D.; Karlický, V.; Šigut, L.; Nezval, J.; Špunda, V. Protective effect of UV-A radiation during acclimation of the photosynthetic apparatus to UV-B treatment. Plant Physiol. Biochem. 2015, 96, 90–96. [Google Scholar] [CrossRef]
- Moreira-Rodriguez, M.; Nair, V.; Benavides, J.; Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. UVA, UVB light doses and harvesting time differentially tailor glucosinolate and phenolic profiles in broccoli sprouts. Molecules 2017, 22, 1065. [Google Scholar] [CrossRef]
- Yang, X.; Xu, H.; Shao, L.; Li, T.; Wang, Y.; Wang, R. Response of photosynthetic capacity of tomato leaves to different LED light wavelength. Environ. Exp. Bot. 2018, 150, 161–171. [Google Scholar] [CrossRef]
- Moosavi-Nezhad, M.; Salehi, R.; Aliniaeifard, S.; Tsaniklidis, G.; Woltering, E.J.; Fanourakis, D.; Zuk-Gołaszewska, K.; Kalaji, H.M. Blue light improves photosynthetic performance during healing and acclimatization of grafted watermelon seedlings. Int. J. Mol. Sci. 2021, 22, 8043. [Google Scholar] [CrossRef]
- Gao, M.; Li, Y.; Jiang, H.; He, R.; Shi, R.; Song, S.; Liu, H. UVA-Radiation Exposure of Different Durations Promoted the Growth, Phytochemicals and Glucosinolate Biosynthesis of Chinese Kale. Int. J. Mol. Sci. 2022, 23, 7619. [Google Scholar] [CrossRef] [PubMed]
- Barbara, F.; Szczak, B.F.; Monika, K.M. The preferences of different cultivars of lettuce seedlings (Lactuca sativa L.) for the spectral composition of light. Agronomy 2021, 11, 1211. [Google Scholar]
- Paucek, I.; Pennisi, G.; Pistillo, A.; Appolloni, E.; Crepaldi, A.; Calegari, B.; Spinelli, F.; Cellini, A.; Gabarrell, X.; Orsini, F.; et al. Supplementary LED interlighting improves yield and precocity of greenhouse tomatoes in the Mediterranean. Agronomy 2020, 10, 1002. [Google Scholar] [CrossRef]
- He, J.; Jawahir, N.K.B.; Lin, Q. Quantity of supplementary LED lightings regulates photosynthetic apparatus, improves photosynthetic capacity and enhances productivity of Cos lettuce grown in a tropical greenhouse. Photosynth. Res. 2021, 149, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.G.; Deng, Y.C.; Hussain, S.; Zou, J.L.; Yuan, J.; Luo, L.; Yang, C.Y.; Yuan, X.Q.; Yang, W.Y. Relationship between cellulose accumulation and lodging resistance in the stem of relay intercropped soybean [Glycine max (L.) men.]. Field Crop. Res. 2016, 196, 261–267. [Google Scholar] [CrossRef]
- Rowe, J.H.; Topping, J.F.; Liu, J.; Lindsey, K. Abscisic acid regulates root growth under osmotic stress conditions via an interacting hormonal network with cytokinin, ethylene and auxin. New Phytol. 2016, 211, 225–239. [Google Scholar] [CrossRef]
- Zürcher, E.; Müller, B. Cytokinin synthesis, signaling, and function—Advances and new insights. Int. Rev. Cell Mol. Biol. 2016, 324, 1–38. [Google Scholar]
- Chory, J.; Reinecke, D.; Sim, S.; Washburn, T.; Brenner, M. A role for cytokinins in de-etiolation in Arabidopsis (det mutants have an altered response to cytokinins). Plant Physiol. 1994, 104, 339–347. [Google Scholar] [CrossRef]
- Marchetti, C.F.; Skrabisova, M.; Galuszka, P.; Novak, O.; Causin, H.F. Blue light suppression alters cytokinin homeostasis in wheat leaves senescing under shading stress. Plant Physiol. Biochem. 2018, 130, 647–657. [Google Scholar] [CrossRef]
- Elkins, C.; van Iersel, M.W. Longer photoperiods with the same daily light integral improve growth of Rudbeckia seedlings in a greenhouse. Hort. Sci. 2020, 55, 1676–1682. [Google Scholar] [CrossRef]
- Lee, M.; Rivard, C.; Pliakoni, E.; Wang, W.; Rajashekar, C.B. Supplemental UV-A and UV-B affect the nutritional quality of lettuce and tomato: Health-promoting phytochemicals and essential nutrients. Am. J. Plant Sci. 2021, 12, 104–126. [Google Scholar] [CrossRef]
- Chen, Y.; Li, T.; Yang, Q.; Zhang, Y.; Zou, J.; Bian, Z.; Wen, X. UVA radiation is beneficial for yield and quality of indoor cultivated lettuce. Front. Plant Sci. 2019, 10, 1563. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Hypocotyl Length (cm) | Stem Diameter (mm) | Leaf Length (cm) | Leaf Width (cm) | Leaf Area (cm2) | Specific Leaf Area (cm2 mg−1) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 18.6 ± 2.2 | a | 13.8 ± 1.4 | a | 3.6 ± 0.4 | b | 6.6 ± 0.6 | c | 6.7 ± 0.6 | c | 30.7 ± 3.1 | c | 0.501 ± 0.035 | a |
| W | 14.0 ± 1.0 | bc | 8.5 ± 0.4 | c | 4.7 ± 0.3 | a | 7.4 ± 0.3 | b | 7.9 ± 0.4 | b | 38.8 ± 2.1 | b | 0.305 ± 0.013 | c |
| UVA | 15.8 ± 0.3 | b | 12.1 ± 0.6 | b | 3.7 ± 0.2 | b | 7.3 ± 0.1 | b | 6.7 ± 0.3 | c | 33.5 ± 2.2 | c | 0.415 ± 0.031 | b |
| WB | 12.7 ± 1.0 | cd | 8.6 ± 0.6 | c | 4.8 ± 0.4 | a | 7.7 ± 0.2 | ab | 8.4 ± 0.3 | a | 42.9 ± 3.6 | ab | 0.271 ± 0.021 | cd |
| W-UVA | 13.5 ± 1.3 | c | 8.5 ± 0.7 | c | 4.9 ± 0.2 | a | 7.3 ± 0.4 | b | 8.0 ± 0.4 | ab | 39.4 ± 2.8 | b | 0.278 ± 0.012 | cd |
| WB-UVA | 11.2 ± 0.4 | d | 7.1 ± 0.5 | d | 4.8 ± 0.4 | a | 8.0 ± 0.1 | a | 8.4 ± 0.2 | ab | 44.4 ± 2.4 | a | 0.262 ± 0.013 | d |
| Treatments | SPAD Value | Net Photosynthetic Rate (μmol m−2 s−1) | Stomatal Conductance (mol m−2 s−1) | Substomatal CO2 Concentration (μmol mol−1) | Transpiration Rate (mmol m−2 s−1) | Apparent Mesophyll Conductance (mol m−2 s−1) | Stomatal Limitation Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 36.3 ± 2.1 | d | 5.2 ± 0.2 | d | 0.06 ± 0.01 | d | 243 ± 24 | b | 0.78 ± 0.09 | d | 0.025 ± 0.004 | d | 0.40 ± 0.06 | a |
| W | 46.3 ± 1.8 | b | 9.6 ± 0.6 | b | 0.19 ± 0.02 | b | 295 ± 8 | a | 2.16 ± 0.04 | b | 0.031 ± 0.002 | c | 0.24 ± 0.02 | c |
| UVA | 39.1 ± 3.5 | c | 6.8 ± 0.3 | c | 0.10 ± 0.01 | c | 281 ± 25 | a | 1.17 ± 0.15 | c | 0.026 ± 0.002 | d | 0.31 ± 0.03 | b |
| WB | 50.5 ± 1.7 | a | 10.8 ± 0.4 | a | 0.24 ± 0.03 | a | 300 ± 8 | a | 2.65 ± 0.22 | a | 0.036 ± 0.001 | b | 0.24 ± 0.02 | c |
| W-UVA | 47.9 ± 1.4 | ab | 10.1 ± 0.6 | b | 0.19 ± 0.03 | b | 293 ± 14 | a | 2.13 ± 0.32 | b | 0.033 ± 0.002 | bc | 0.24 ± 0.03 | c |
| WB-UVA | 50.5 ± 1.5 | a | 11.2 ± 0.8 | a | 0.22 ± 0.02 | ab | 295 ± 12 | a | 2.39 ± 0.21 | ab | 0.039 ± 0.004 | a | 0.23 ± 0.02 | c |
| Treatments | Shoot Fresh Weight (g Per Plant) | Root Fresh Weight (g Per Plant) | Shoot Dry Weight (g Per Plant) | Root Dry Weight (g Per Plant) | Seedling Quality Index | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 2.92 ± 0.13 | d | 0.45 ± 0.03 | d | 0.198 ± 0.030 | c | 0.016 ± 0.002 | d | 0.025 ± 0.003 | c |
| W | 3.49 ± 0.44 | bc | 1.17 ± 0.09 | c | 0.283 ± 0.020 | b | 0.040 ± 0.005 | bc | 0.055 ± 0.006 | b |
| UVA | 3.05 ± 0.27 | cd | 0.44 ± 0.01 | d | 0.216 ± 0.026 | c | 0.015 ± 0.001 | d | 0.022 ± 0.002 | c |
| WB | 4.09 ± 0.19 | a | 1.42 ± 0.18 | b | 0.316 ± 0.014 | ab | 0.045 ± 0.006 | b | 0.065 ± 0.009 | ab |
| W-UVA | 4.13 ± 0.10 | a | 1.09 ± 0.09 | c | 0.339 ± 0.038 | a | 0.037 ± 0.004 | c | 0.057 ± 0.005 | b |
| WB-UVA | 3.82 ± 0.45 | ab | 1.90 ± 0.18 | a | 0.348 ± 0.046 | a | 0.059 ± 0.009 | a | 0.069 ± 0.008 | a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Z.; Wang, C.; Wang, L.; Li, X.; Wang, G.; Yang, Y. The Combinations of White, Blue, and UV-A Light Provided by Supplementary Light-Emitting Diodes Promoted the Quality of Greenhouse-Grown Cucumber Seedlings. Agriculture 2022, 12, 1593. https://doi.org/10.3390/agriculture12101593
Yan Z, Wang C, Wang L, Li X, Wang G, Yang Y. The Combinations of White, Blue, and UV-A Light Provided by Supplementary Light-Emitting Diodes Promoted the Quality of Greenhouse-Grown Cucumber Seedlings. Agriculture. 2022; 12(10):1593. https://doi.org/10.3390/agriculture12101593
Chicago/Turabian StyleYan, Zhengnan, Chunling Wang, Long Wang, Xin Li, Guanjie Wang, and Yanjie Yang. 2022. "The Combinations of White, Blue, and UV-A Light Provided by Supplementary Light-Emitting Diodes Promoted the Quality of Greenhouse-Grown Cucumber Seedlings" Agriculture 12, no. 10: 1593. https://doi.org/10.3390/agriculture12101593
APA StyleYan, Z., Wang, C., Wang, L., Li, X., Wang, G., & Yang, Y. (2022). The Combinations of White, Blue, and UV-A Light Provided by Supplementary Light-Emitting Diodes Promoted the Quality of Greenhouse-Grown Cucumber Seedlings. Agriculture, 12(10), 1593. https://doi.org/10.3390/agriculture12101593