
Factors Affecting Tolerance to Low Night Temperature Differ by Fruit Types in Tomato


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. The Effect of Low Night Temperature on the Growth and Development of Tomato Accessions
2.3. Electrolyte Conductivity, Total Chlorophyll and Proline Content in Leaves
2.4. Statistical Analyses
3. Results
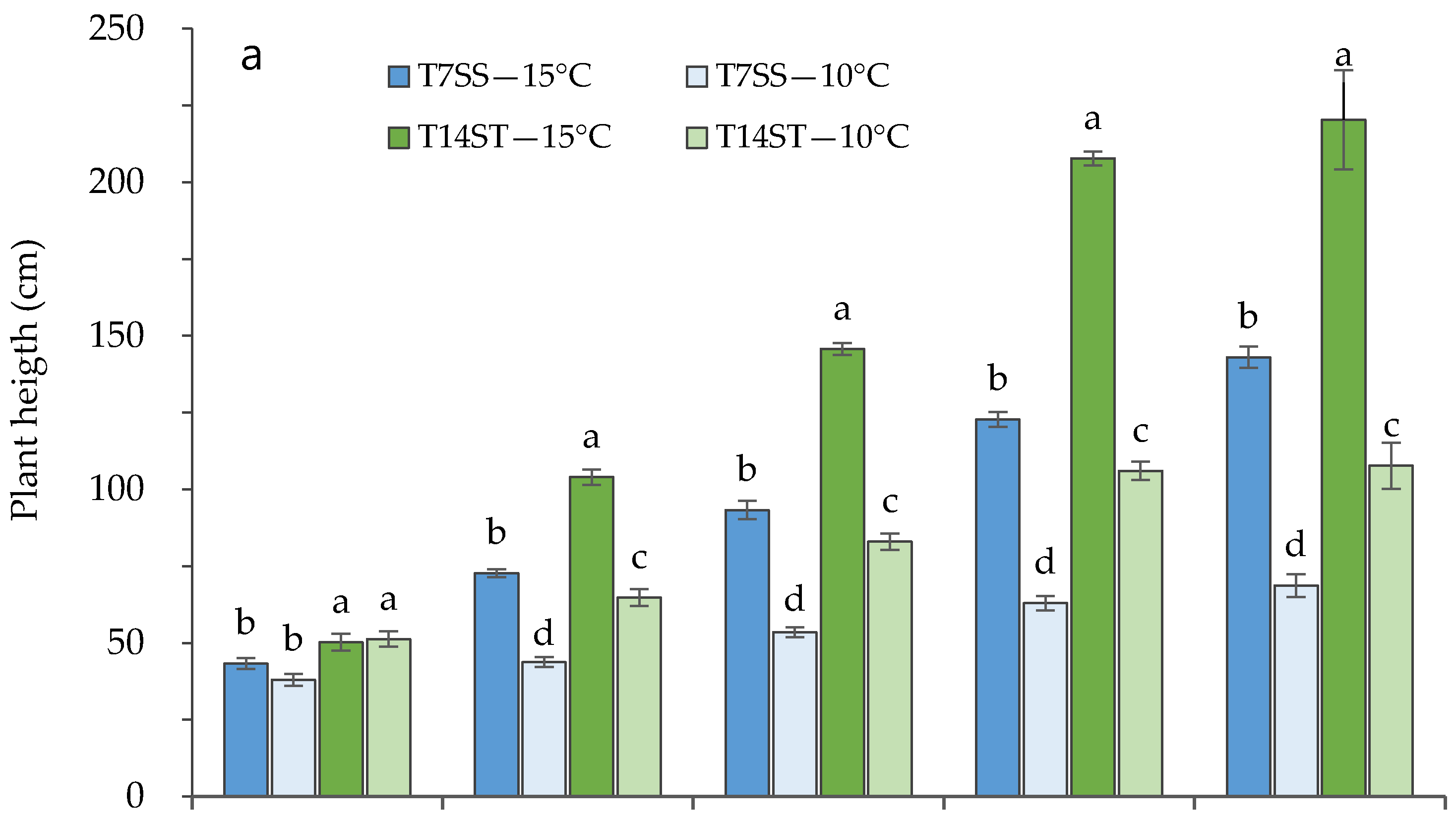
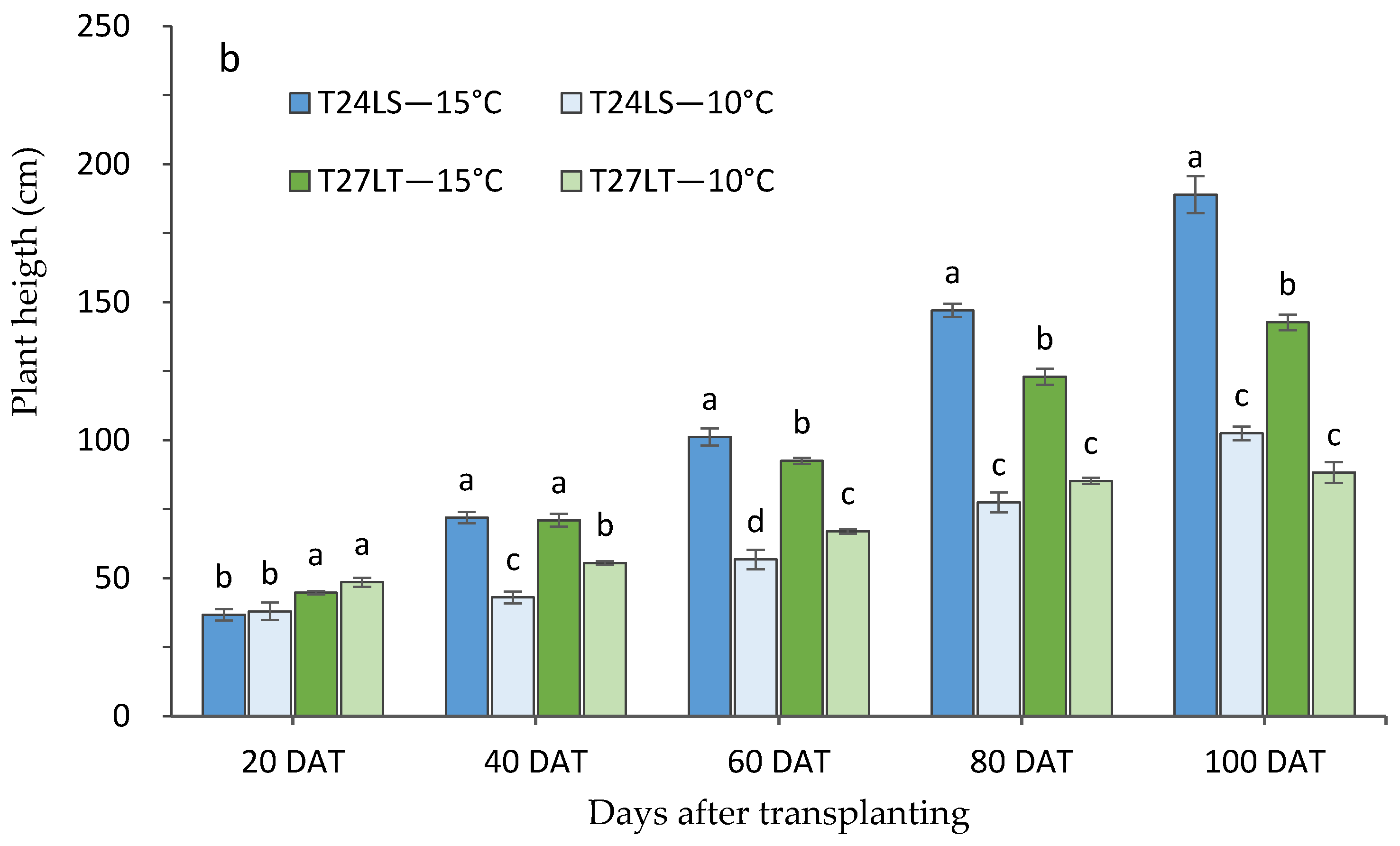
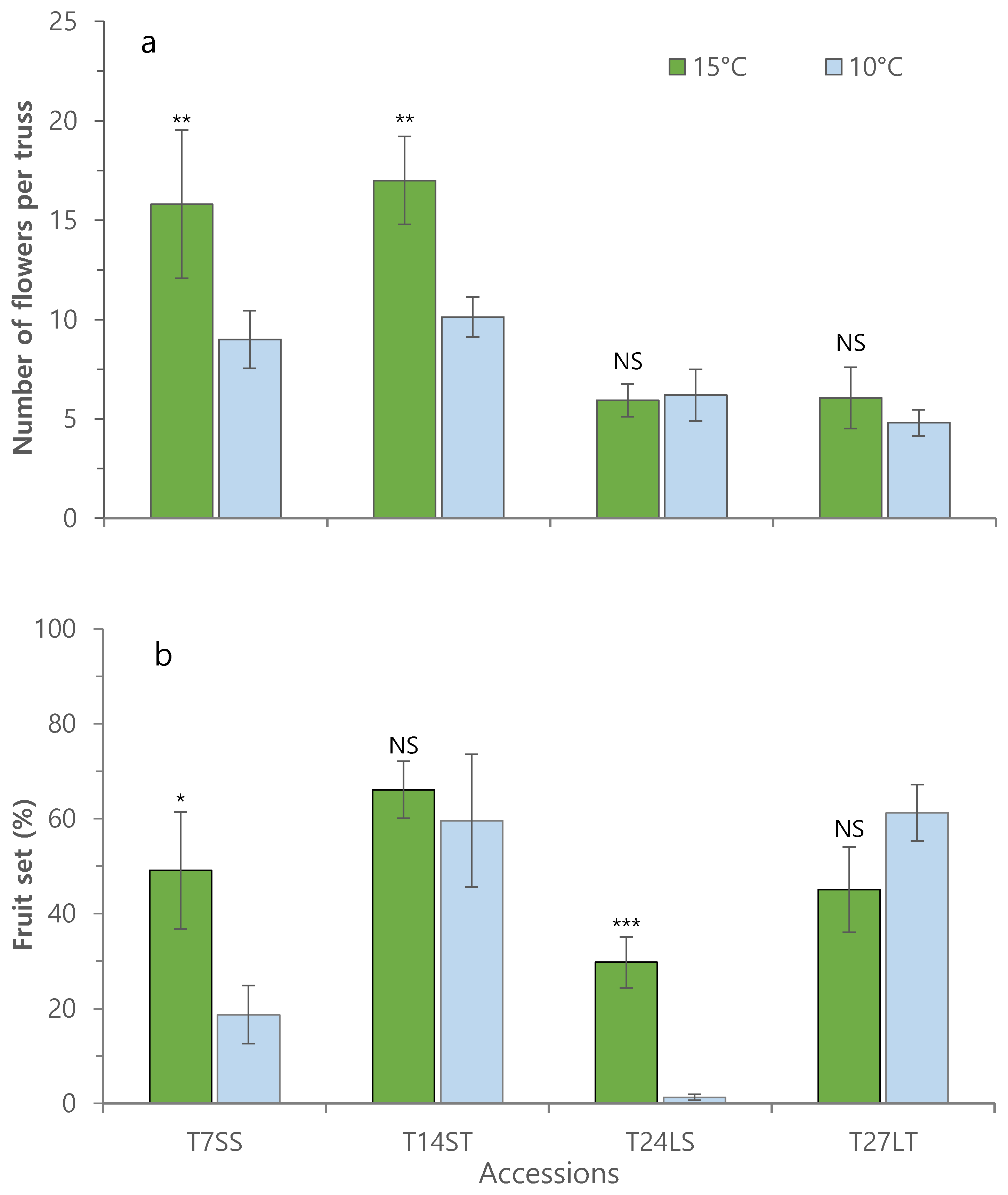
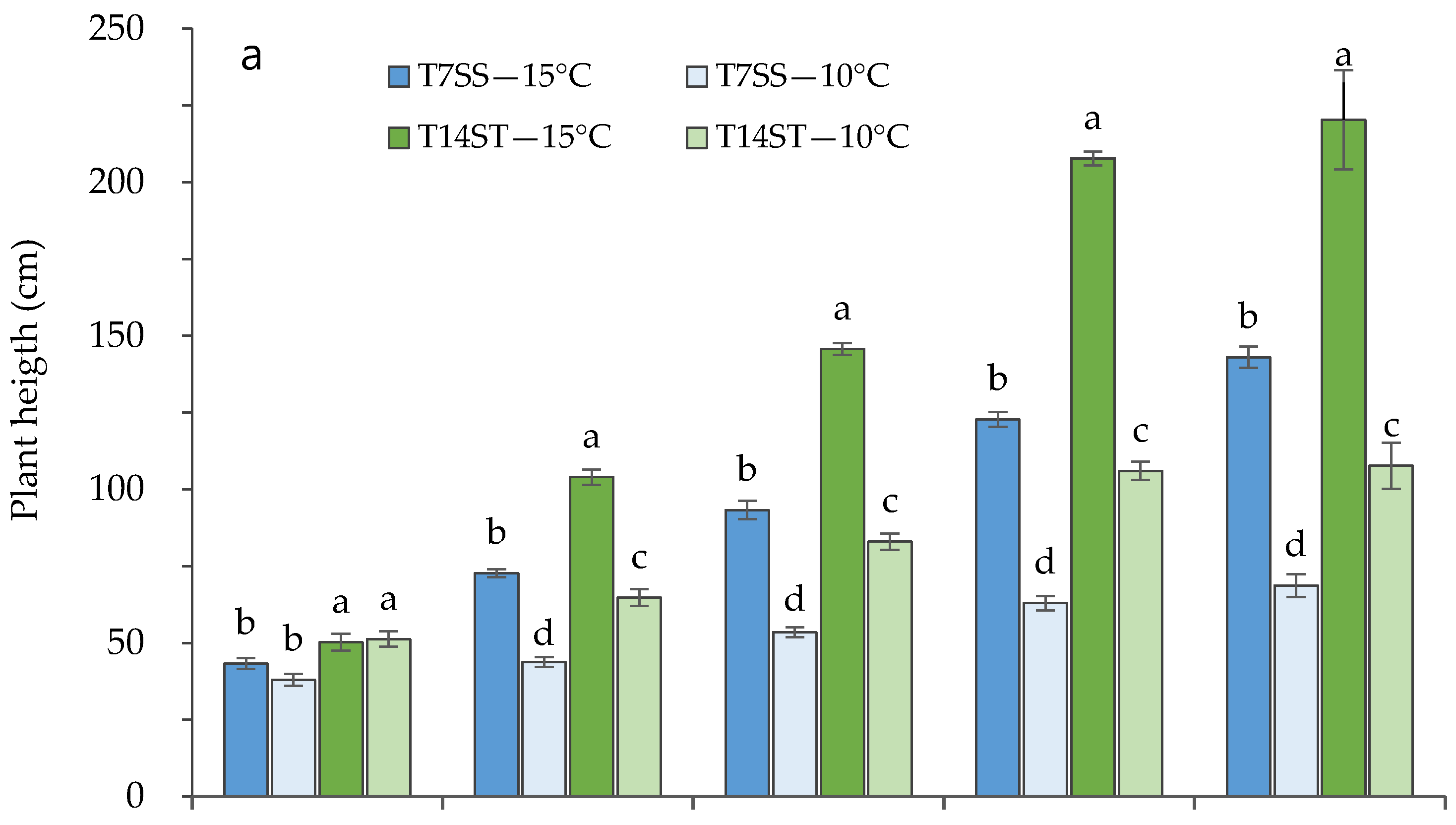
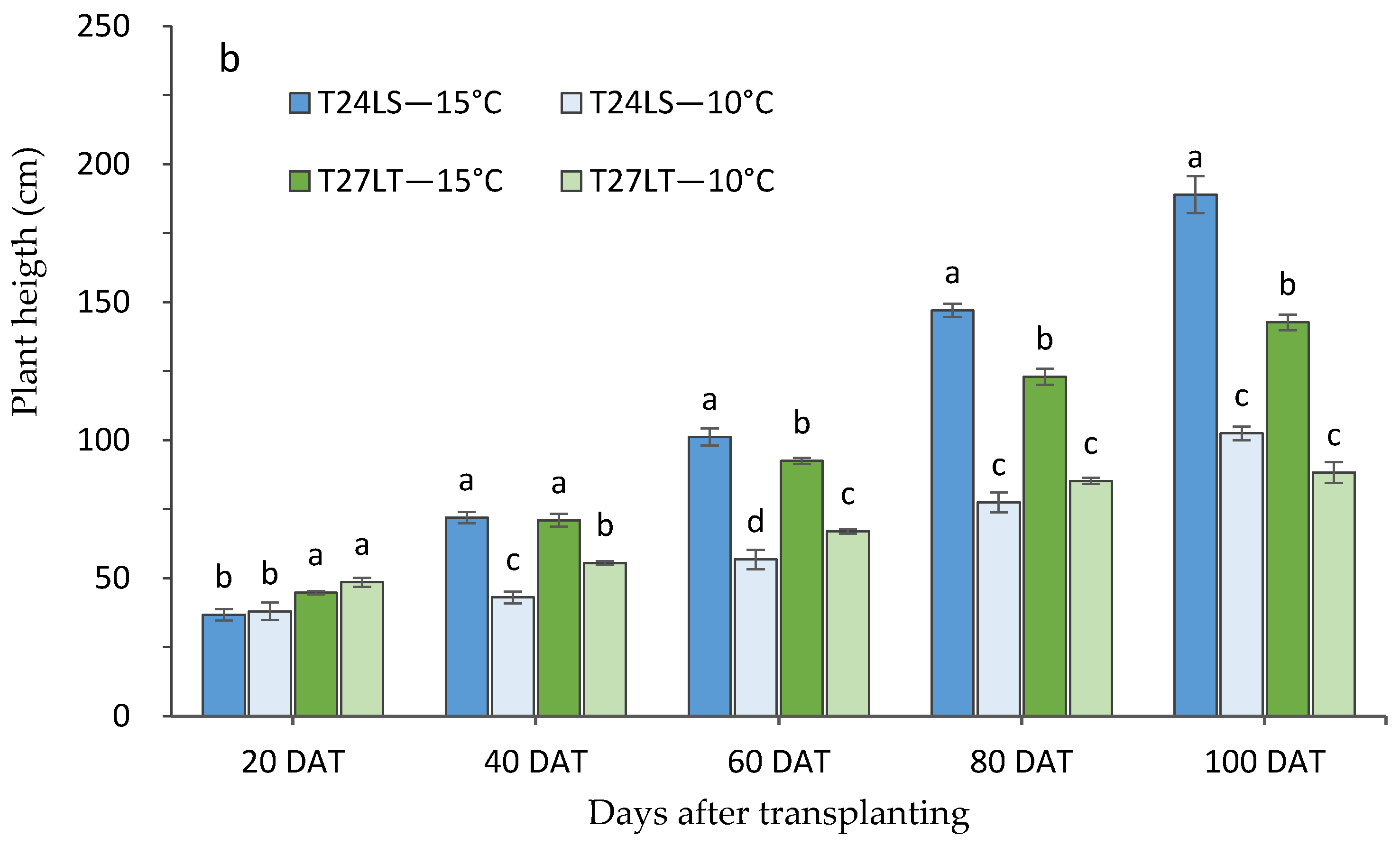
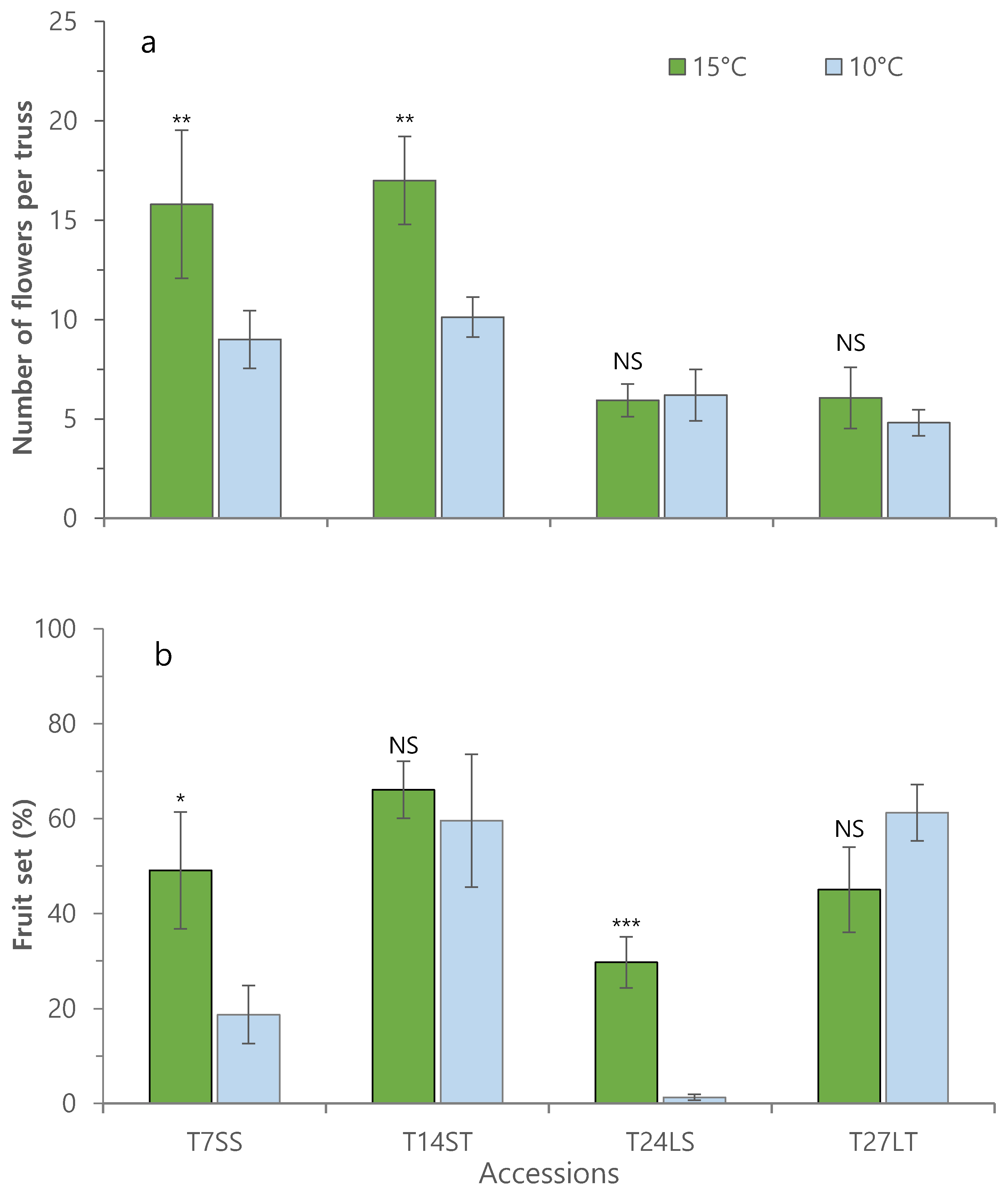
3.1. Effect of Low Night Temperature on the Growth and Development of Tomato Accessions
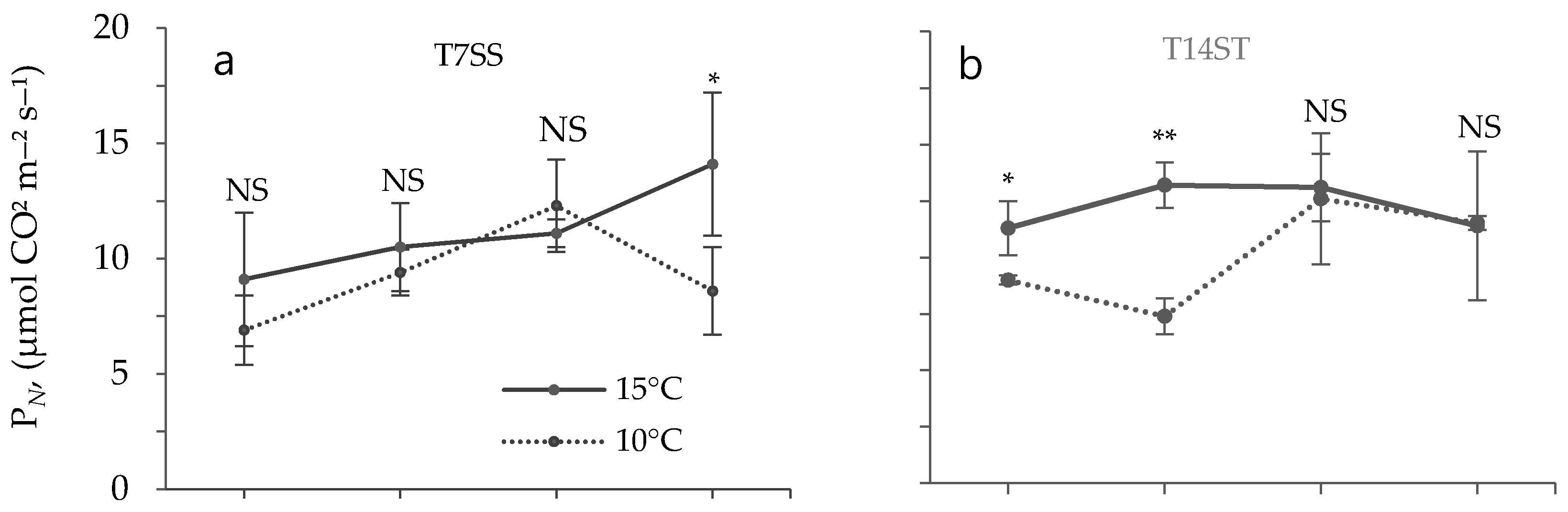
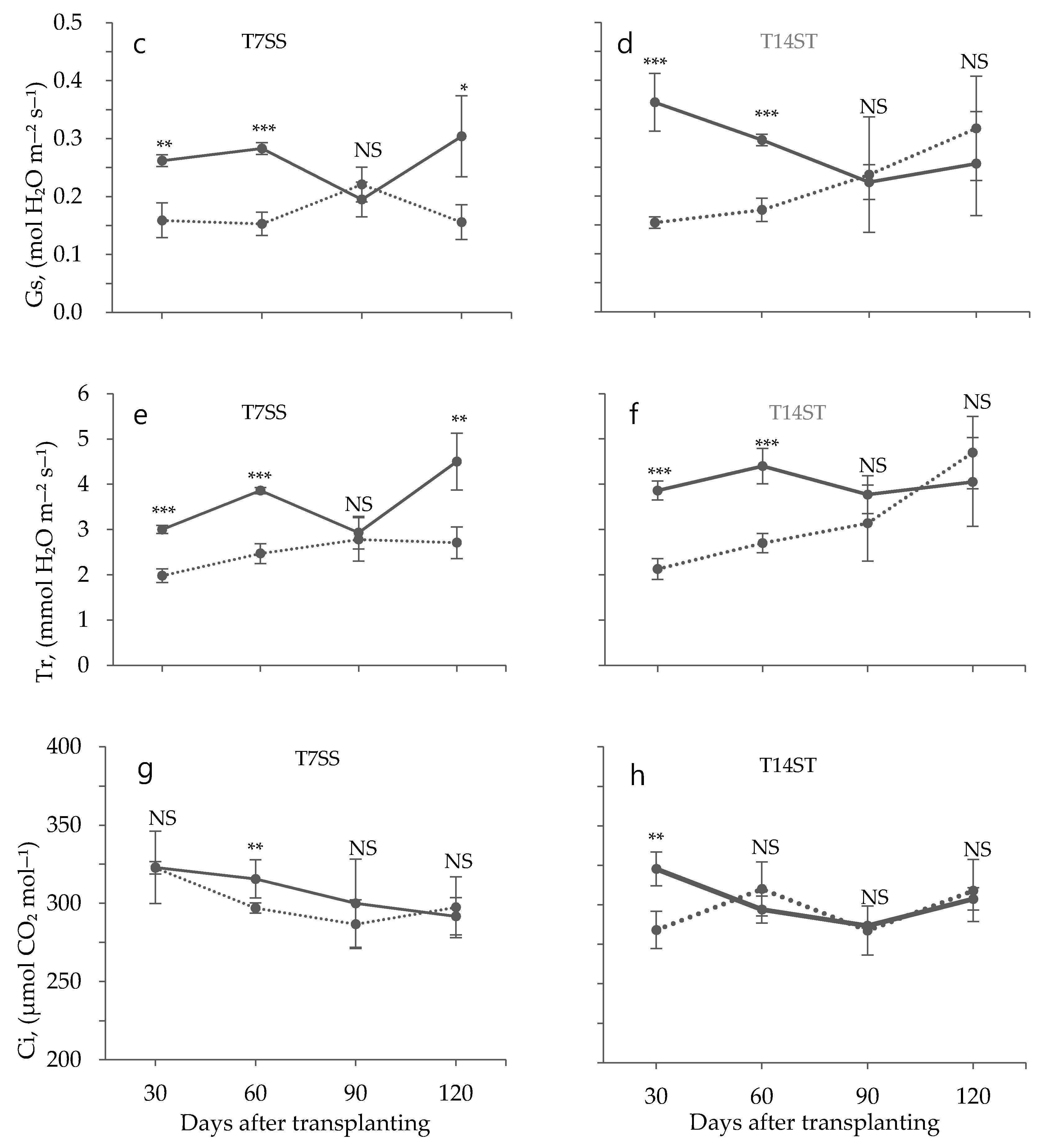
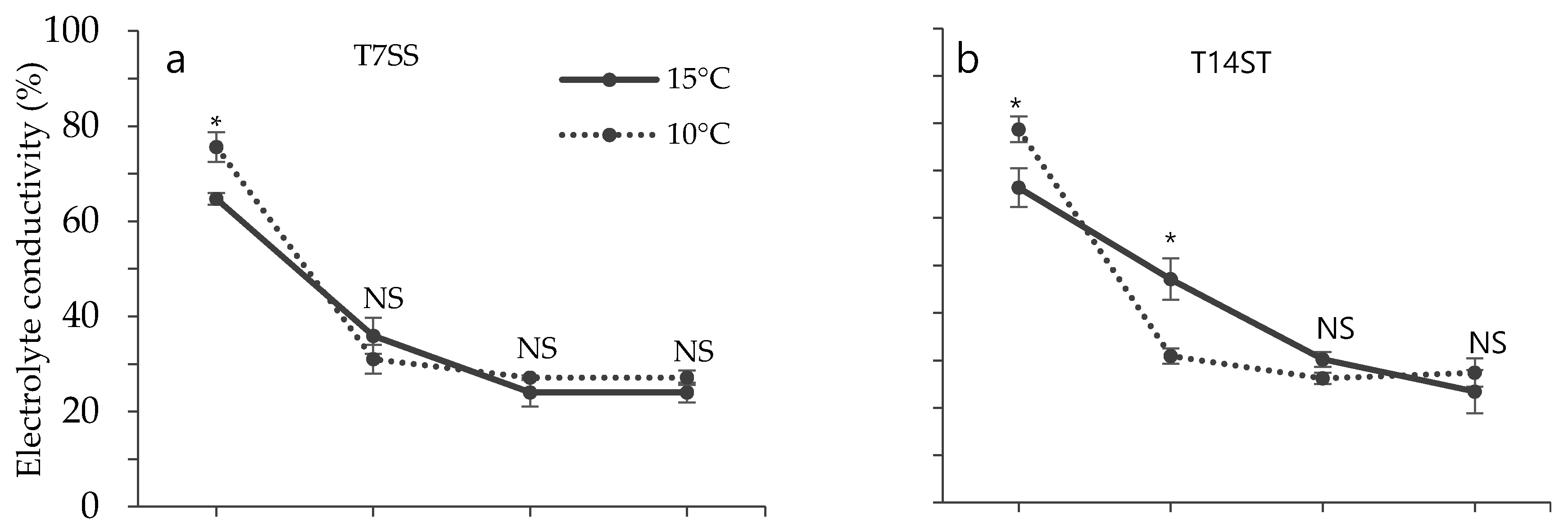
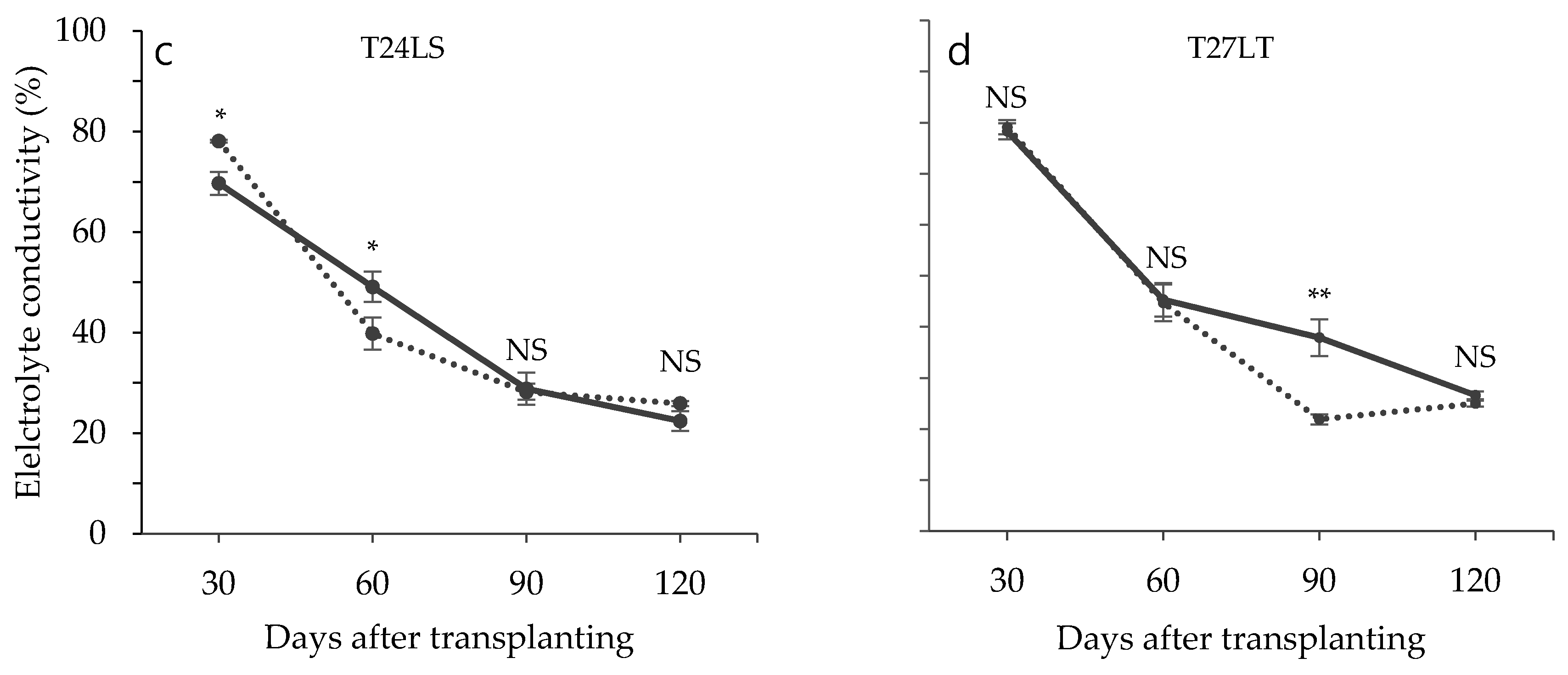
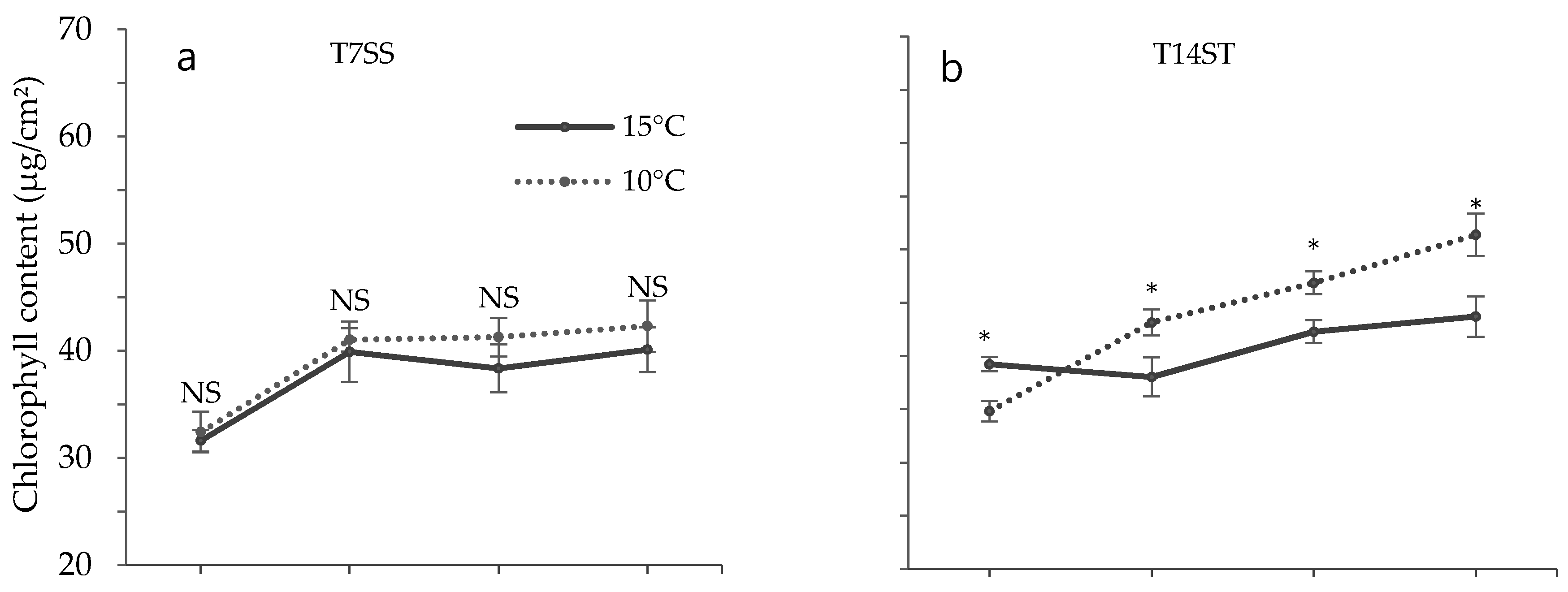
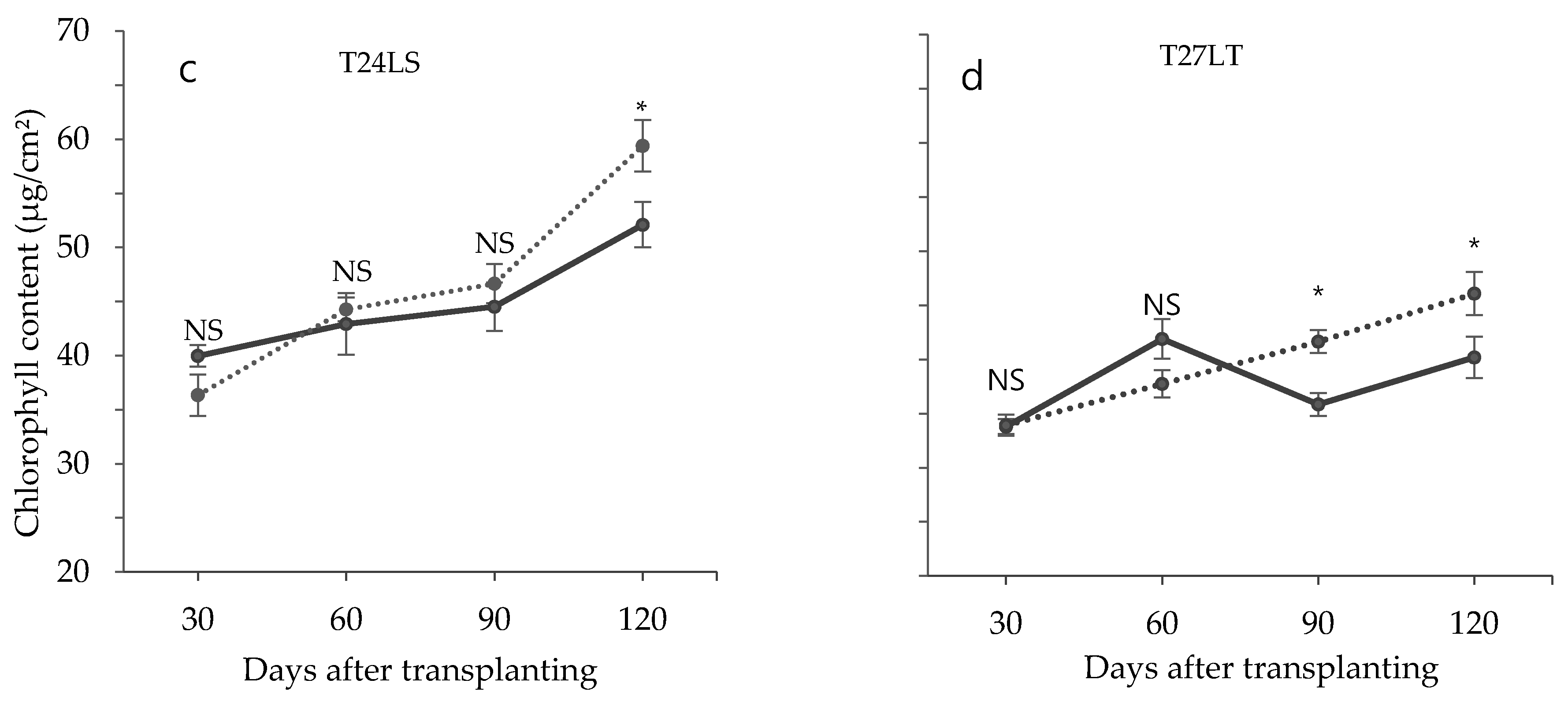
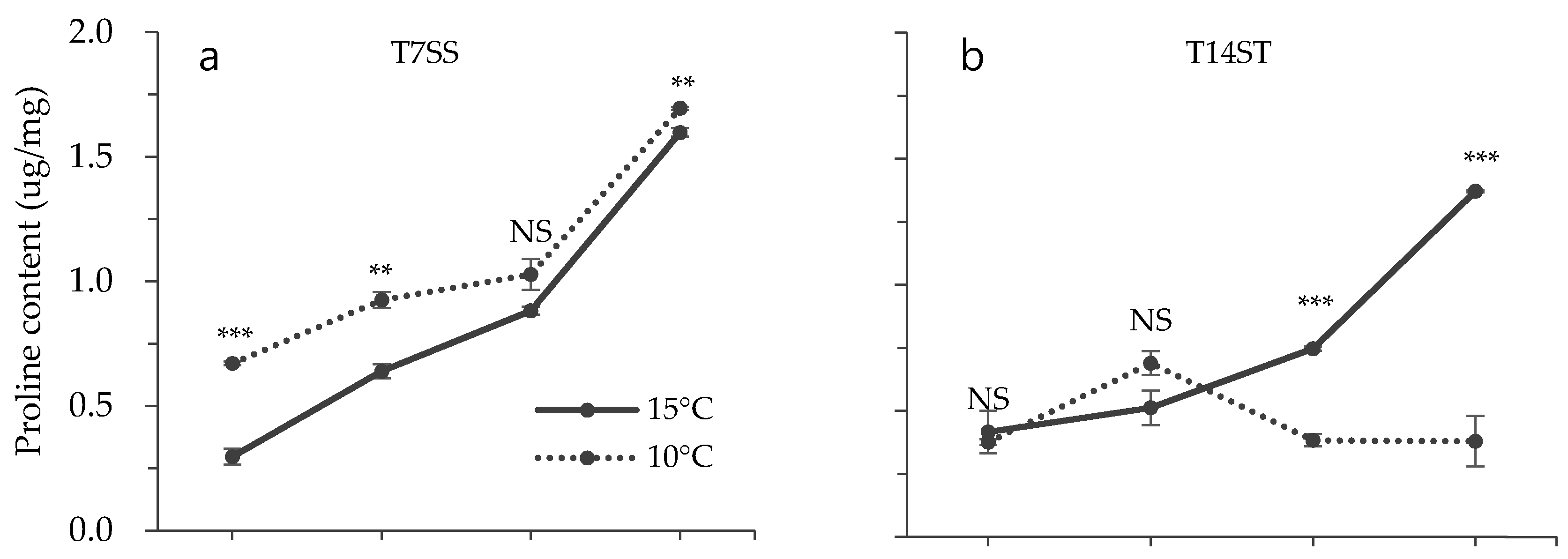
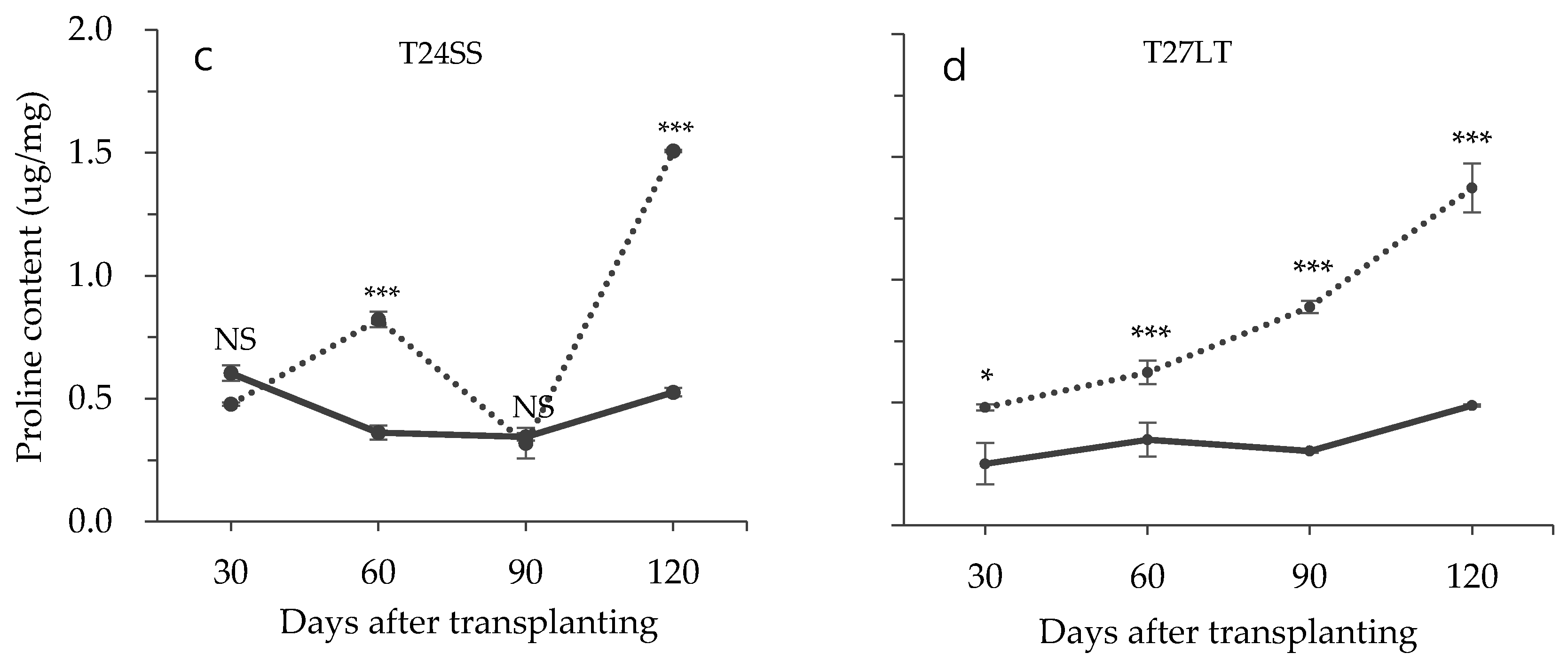
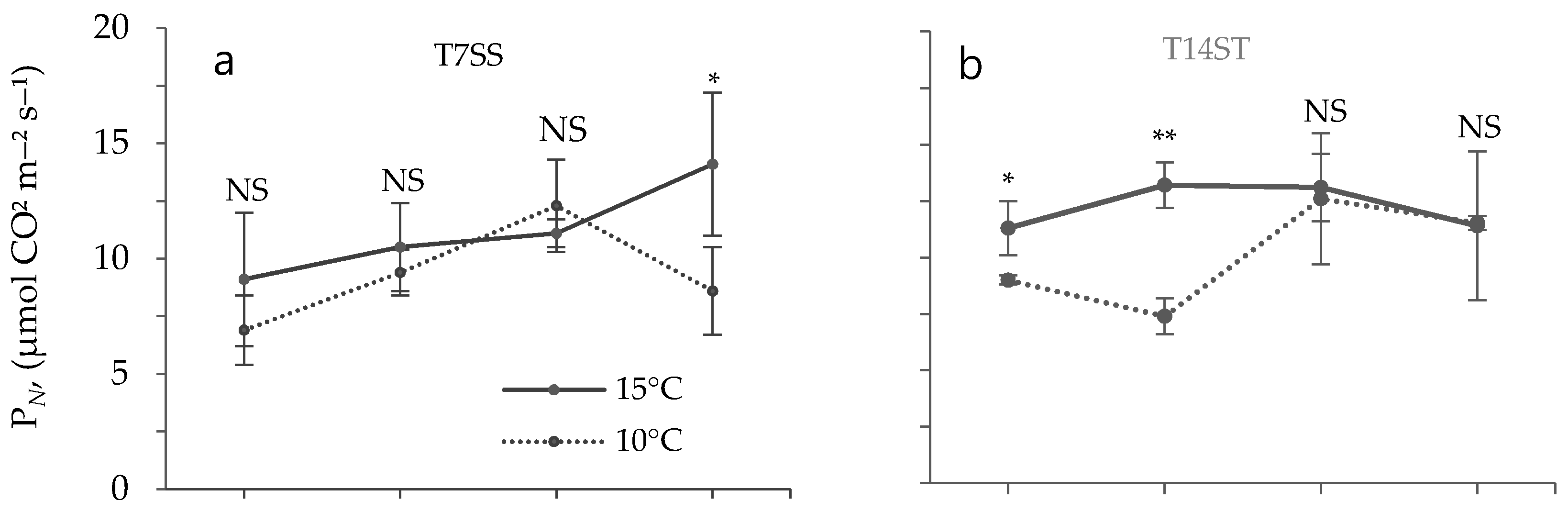
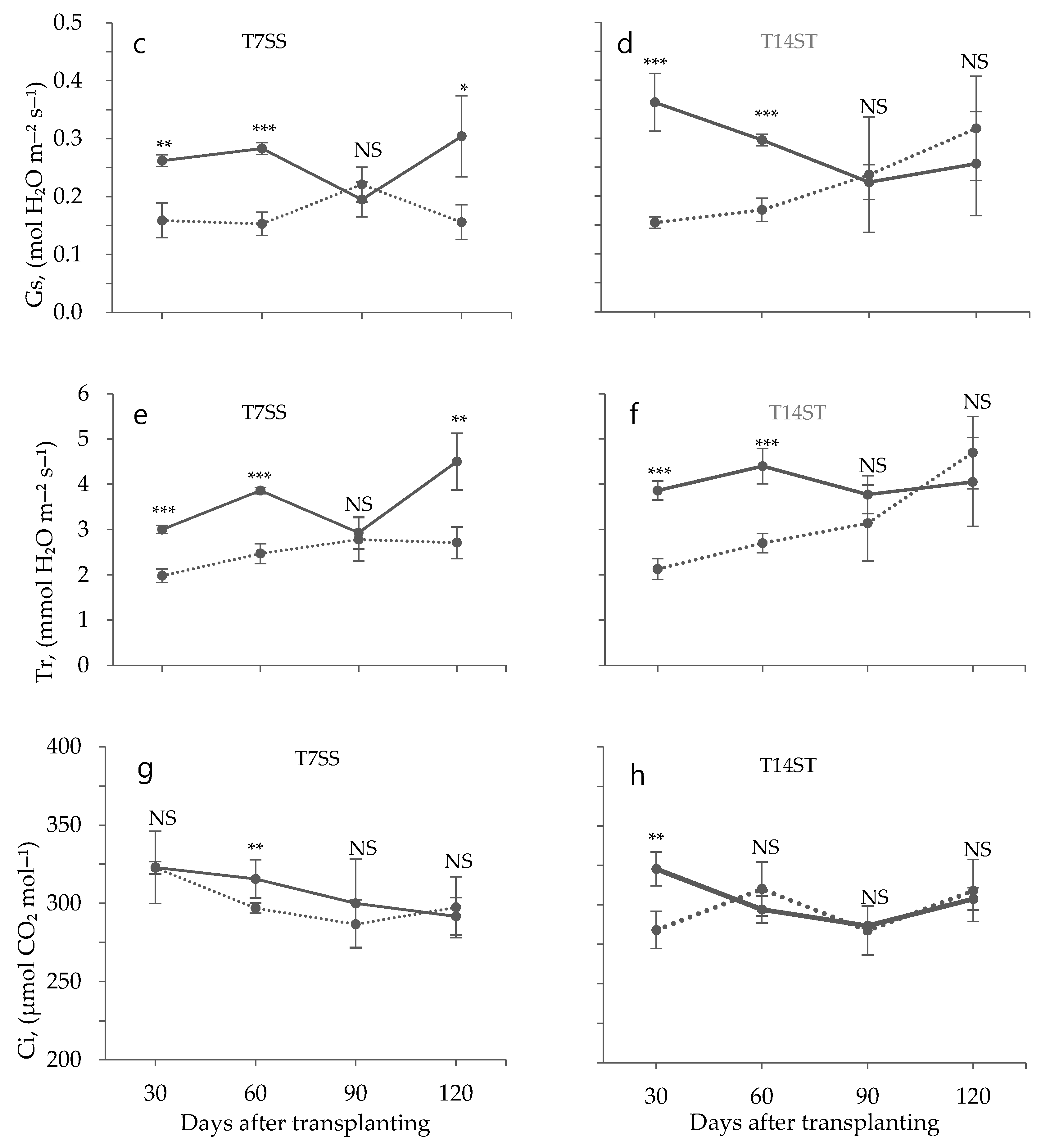
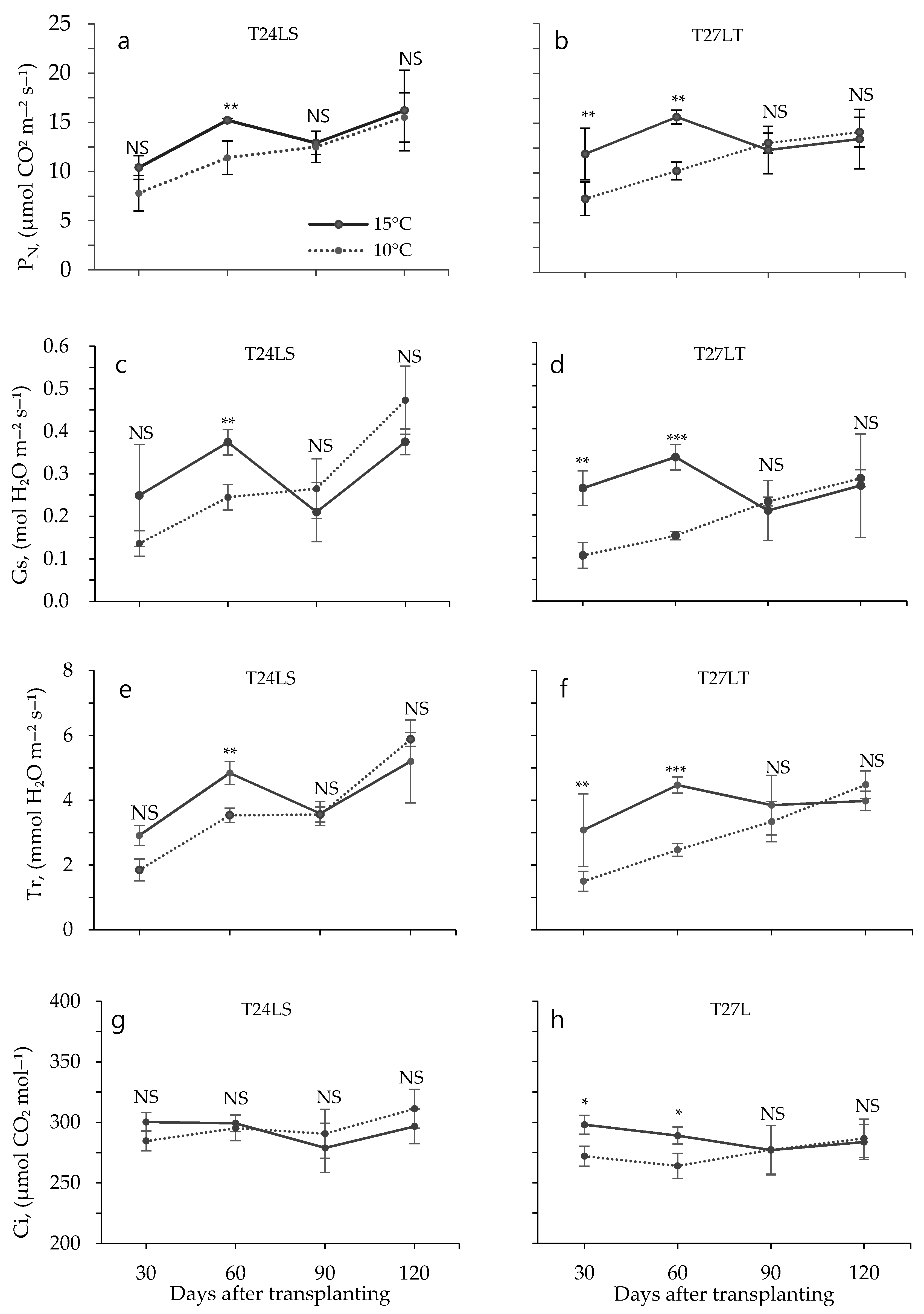
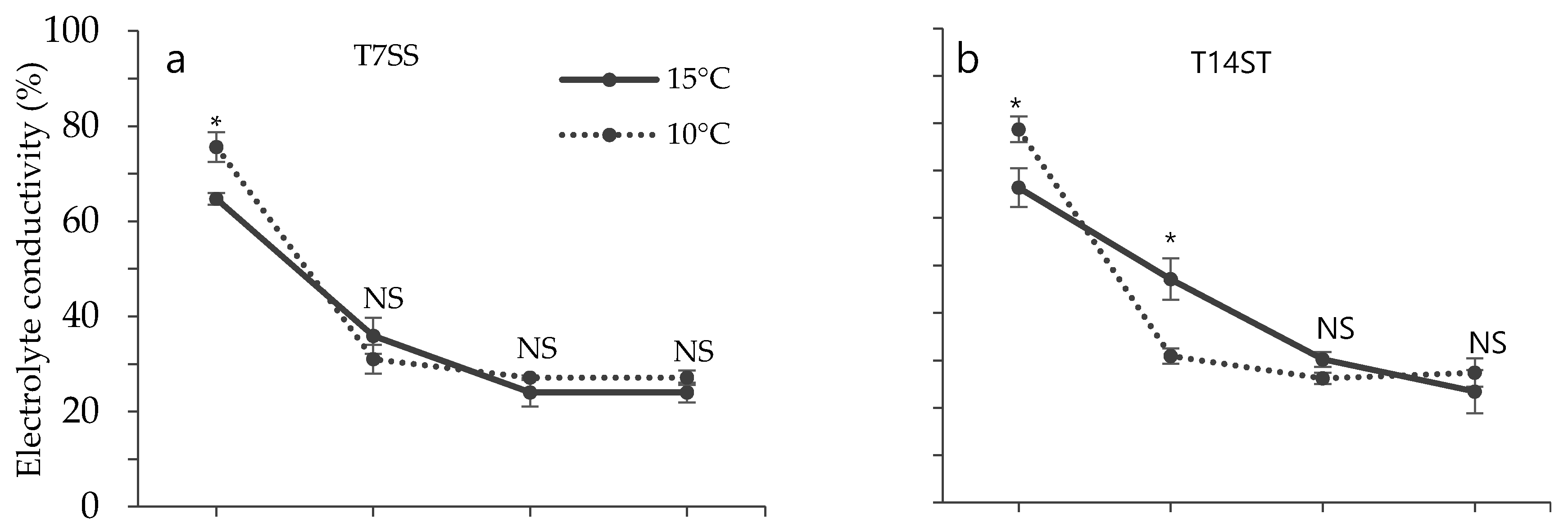
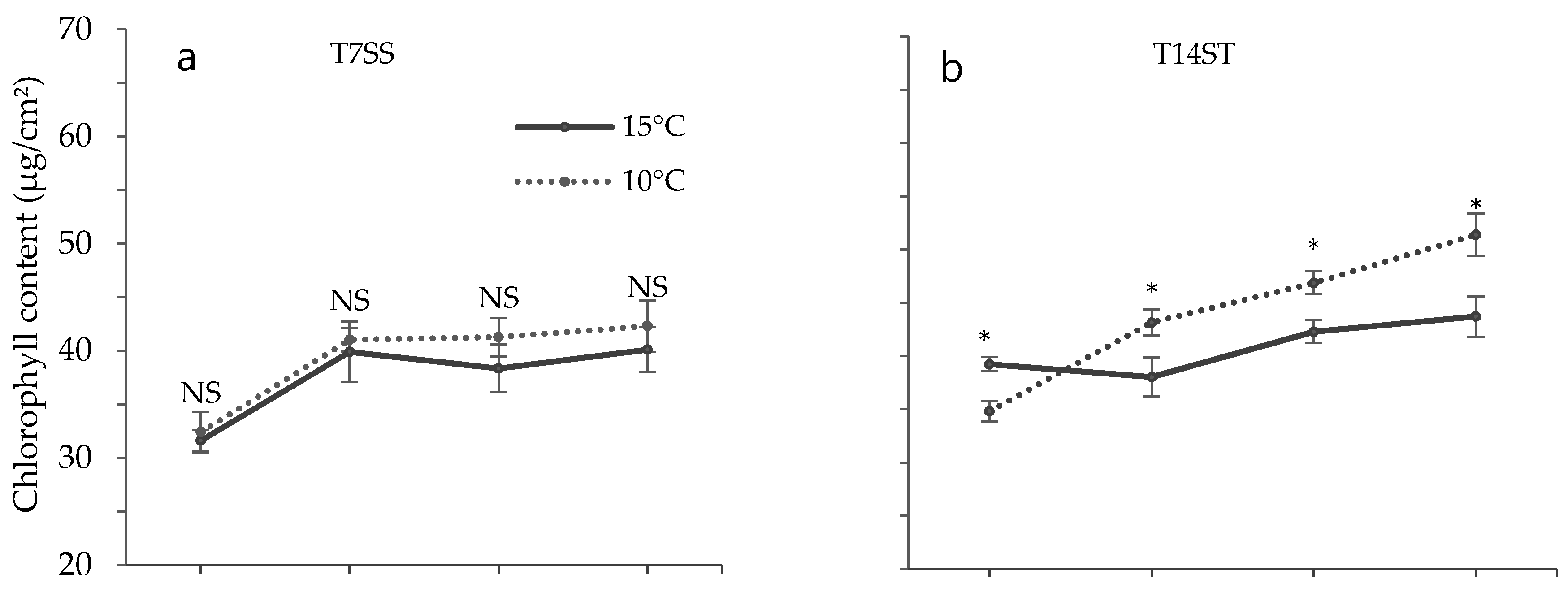
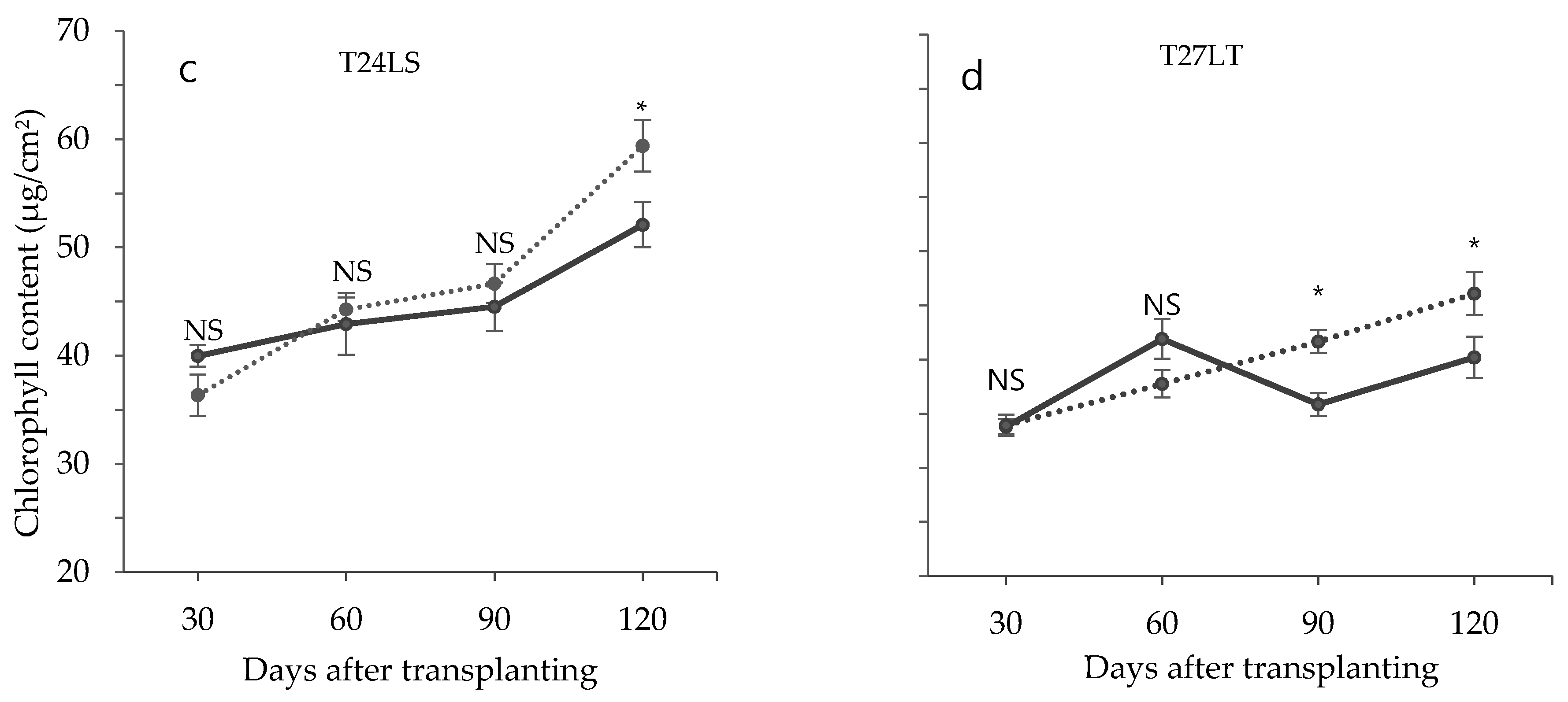
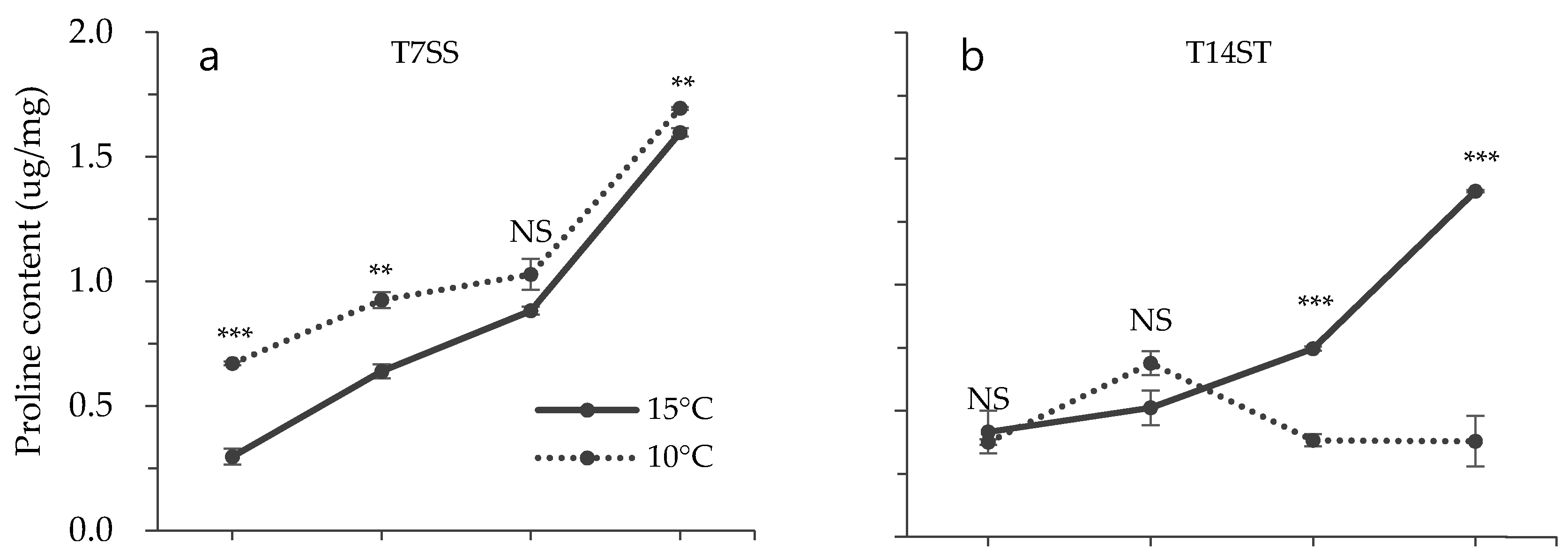
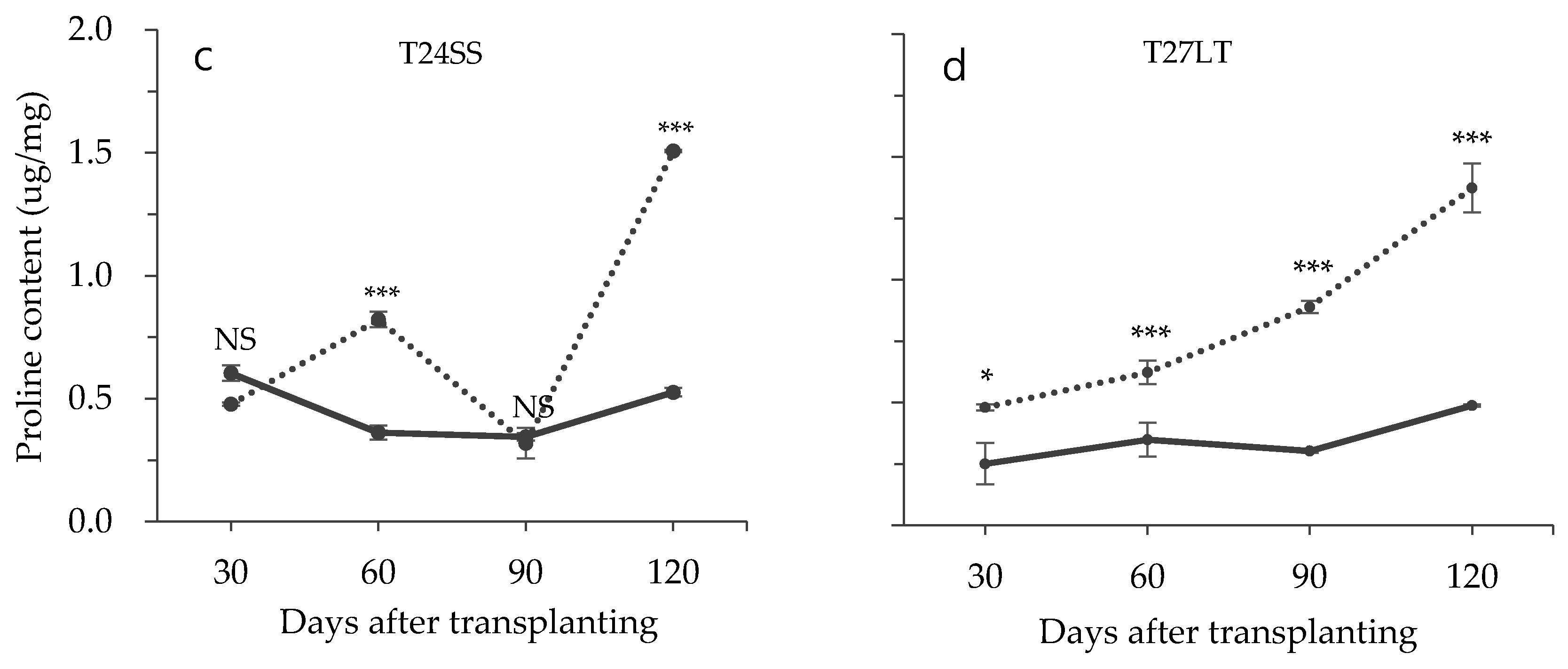
3.2. Effect of Low Temperature on Biochemical Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Hoek, I.; Ten Cate, C.H.H.; Keijzer, C.J.; Schel, J.H.; Dons, H.J. Development of the Fifth Leaf is Indicative for Whole Plant Performance at Low Temperature in Tomato. Ann. Bot. 1993, 72, 367–374. [Google Scholar] [CrossRef]
- Venema, J.H.; Posthumus, F.; de Vries, M.; van Hasselt, P.R. Differential response of domestic and wild Lycopersicon species to chilling under low light: Growth, carbohydrate content, photosynthesis and the xanthophyll cycle. J. Physiol. Plant. 1999, 105, 81–88. [Google Scholar] [CrossRef]
- Liu, H.; Ouyang, B.; Zhang, J.; Wang, T.; Li, H.; Zhang, Y.; Yu, C.; Ye, Z. Differential Modulation of Photosynthesis, Signaling, and Transcriptional Regulation between Tolerant and Sensitive Tomato Genotypes under Cold Stress. PLoS ONE 2012, 7, e50785. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, X.; Chen, D.; Li, J.; Zhang, Y.; Wang, A. A comparison of the low temperature transcriptomes of two tomato genotypes that differ in freezing tolerance: Solanum lycopersicum and Solanum habrochaites. BMC Plant Biol. 2015, 15, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittwer, S.H.; Tuebner, F.G. Cold exposure of tomato seedlings and flower formation. J. Am. Soc. Hortic. Sci. 1957, 67, 369–376. [Google Scholar]
- Hurd, R.G.; Graves, C.J. Some effects of air and root temperatures on the yield and quality of glasshouse tomatoes. J. Hortic. Sci. 1985, 60, 359–371. [Google Scholar] [CrossRef]
- Sherzod, R.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Kim, J.H.; Nam, C.W.; Chae, W.B. Traits Affecting Low Temperature Tolerance in Tomato and Its Application to Breeding Program. Plant Breed. Biotechnol. 2019, 7, 350–359. [Google Scholar] [CrossRef]
- Calvert, A. Effect of the Early Environment on Development of Flowering in the Tomato: I. Temperature. J. Hortic. Sci. 1957, 32, 9–17. [Google Scholar] [CrossRef]
- Rylski, I. Fruit set and development of seeded and seedless tomato fruits under diverse regimes of temperature and pollination. J. Am. Soc. Hortic. Sci. 1979, 104, 835–838. [Google Scholar]
- Picken, A.J.F. A review of pollination and fruit set in the tomato (Lycopersicon esculentum Mill.). J. Hortic. Sci. 1984, 59, 1–13. [Google Scholar] [CrossRef]
- Ercan, N.; Vural, H. the effects of low temperatures on fruit set of tomatoes. Acta Hortic. 1994, 36, 65–72. [Google Scholar] [CrossRef]
- Graham, D.; Patterson, B.D. Responses of Plants to Low, Nonfreezing Temperatures: Proteins, Metabolism, and Acclimation. Annu. Rev. Plant Physiol. 1982, 33, 347–372. [Google Scholar] [CrossRef]
- Foolad, M.; Lin, G. Relationship between Cold Tolerance during Seed Germination and Vegetative Growth in Tomato: Germplasm Evaluation. J. Am. Soc. Hortic. Sci. 2000, 125, 679–683. [Google Scholar] [CrossRef]
- Zhang, X.; Fowler, S.G.; Cheng, H.; Lou, Y.; Rhee, S.Y.; Stockinger, E.J.; Thomashow, M.F. Freezing-sensitive tomato has a functional CBF cold response pathway, but a CBF regulon that differs from that of freezing-tolerantArabidopsis. Plant J. 2004, 39, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Wang, H.-S.; Yang, S.; Tang, X.-F.; Duan, M.; Meng, Q.-W. Overexpression of endoplasmic reticulum omega-3 fatty acid desaturase gene improves chilling tolerance in tomato. Plant Physiol. Biochem. 2009, 47, 1102–1112. [Google Scholar] [CrossRef]
- Miura, K.; Shiba, H.; Ohta, M.; Kang, S.W.; Sato, A.; Yuasa, T.; Iwaya-Inoue, M.; Kamada, H.; Ezura, H. SlICE1 encoding a MYC-type transcription factor controls cold tolerance in tomato, Solanum lycopersicum. Plant Biotechnol. 2012, 29, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Campos, P.S.; Quartin, V.N.; Ramalho, J.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Munir, S.; Liu, H.; Xing, Y.; Hussain, S.; Ouyang, B.; Zhang, Y.; Li, H.; Ye, Z. Overexpression of calmodulin-like (ShCML44) stress-responsive gene from Solanum habrochaites enhances tolerance to multiple abiotic stresses. Sci. Rep. 2016, 6, 31772. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Wang, Y.; Wang, Q.; Dang, N.; Wang, L.; Liu, C.; Zhu, J.; Zhan, X. The tomato 2-oxoglutarate-dependent dioxygenase gene SlF3HL is critical for chilling stress tolerance. Hortic. Res. 2019, 6, 45. [Google Scholar] [CrossRef] [Green Version]
- Claussen, W. Proline as a measure of stress in tomato plants. Plant Sci. 2005, 168, 241–248. [Google Scholar] [CrossRef]
- Rosales, M.A.; Rios, J.J.; Castellano, R.; López-Carrión, A.I.; Romero, L.; Ruíz, J.M. Proline metabolism in cherry tomato exocarp in relation to temperature and solar radiation. J. Hortic. Sci. Biotechnol. 2007, 82, 739–744. [Google Scholar] [CrossRef]
- Ding, F.; Liu, B.; Zhang, S. Exogenous melatonin ameliorates cold-induced damage in tomato plants. Sci. Hortic. 2017, 219, 264–271. [Google Scholar] [CrossRef]
- Janssen, L.H.J.; Van Oeveren, J.C.; Van Hasselt, P.R.; Kuiper, P.J.C. Genotypic variation in chlorophyll fluorescence parameters, photosynthesis and growth of tomato grown at low temperature and low irradiance. Photosynthetica 1995, 31, 301–314. [Google Scholar]
- Xiaoa, F.; Yang, Z.; Zhua, L. Low temperature and weak light affect greenhouse tomato growth and fruit quality. J. Plant Sci. 2018, 6, 16–24. [Google Scholar] [CrossRef]
- Hu, W.H.; Zhou, Y.H.; Du, Y.S.; Xia, X.J.; Yu, J.Q. Differential response of photosynthesis in greenhouse—And field-ecotypes of tomato to long-term chilling under low light. J. Plant Physiol. 2006, 163, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Goodstal, F.J.; Kohler, G.R.; Randall, L.B.; Bloom, A.J.; St Clair, D.A. A major QTL introgressed from wild Lycopersicon hirsutum confers chilling tolerance to cultivated tomato (Lycopersicon esculentum). J. Theor. Appl. Genet. 2005, 111, 898–905. [Google Scholar] [CrossRef]
- De Koning, A.N.M. The effect of different day/night temperature regimes on growth, development and yield of glasshouse tomatoes. J. Hortic. Sci. 1988, 63, 465–471. [Google Scholar] [CrossRef]
- Elings, A.; Kempkes, F.L.K.; Kaarsemaker, R.C.; Ruijs, M.N.A.; Van De Braak, N.J.; Dueck, T.A. The energy balance and en-ergy-saving measures in greenhouse tomato cultivation. Acta Hortic. 2005, 691, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Sherzod, R.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Chae, W.B. Physiological traits associated with high temperature tolerance differ by fruit types and sizes in tomato (Solanum lycopersicum L.). Hortic. Environ. Biotechnol. 2020, 61, 837–847. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Angeles Morales, M.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. Plant Physiol. 2005, 162, 281–2896. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Ghorbanpour, A.; Salimi, A.; Ghanbary, M.A.T.; Pirdashti, H.; Dehestani, A. The effect of Trichoderma harzianum in mitigating low temperature stress in tomato (Solanum lycopersicum L.) plants. Sci. Hortic. 2018, 230, 134–141. [Google Scholar] [CrossRef]
- Fernandez-Munoz, R.; Cuartero, J. Effects of temperature and irradiance on stigma exertion, ovule viability and embryo development in tomato. J. Hortic. Sci. 1991, 66, 395–401. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.J.; Salvucci, M.E.; Schultz, C.J.; Rumsewicz, M.P.; Johnson, K.L.; Jones, B.J.; Gaspar, Y.M.; Bacic, A. Sensitivity of Photosynthesis in a C4 Plant, Maize, to Heat Stress. Plant Physiol. 2002, 129, 1773–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, E.-Y.; Rajametov, S.N.; Cho, M.-C.; Jeong, H.-B.; Chae, W.-B. Factors Affecting Tolerance to Low Night Temperature Differ by Fruit Types in Tomato. Agriculture 2021, 11, 681. https://doi.org/10.3390/agriculture11070681
Yang E-Y, Rajametov SN, Cho M-C, Jeong H-B, Chae W-B. Factors Affecting Tolerance to Low Night Temperature Differ by Fruit Types in Tomato. Agriculture. 2021; 11(7):681. https://doi.org/10.3390/agriculture11070681
Chicago/Turabian StyleYang, Eun-Young, Sherzod Nigmatullayevich Rajametov, Myeong-Cheoul Cho, Hyo-Bong Jeong, and Won-Byoung Chae. 2021. "Factors Affecting Tolerance to Low Night Temperature Differ by Fruit Types in Tomato" Agriculture 11, no. 7: 681. https://doi.org/10.3390/agriculture11070681
APA StyleYang, E.-Y., Rajametov, S. N., Cho, M.-C., Jeong, H.-B., & Chae, W.-B. (2021). Factors Affecting Tolerance to Low Night Temperature Differ by Fruit Types in Tomato. Agriculture, 11(7), 681. https://doi.org/10.3390/agriculture11070681