
Preliminary Study of Body Temperature Emissivity in Rabbits Selected for Litter Size Residual Variability




Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals
2.3. Traits
2.4. Statistical Analysis
3. Results
3.1. Correlated Response to Selection in Temperature Emissivity
3.2. Effect of Season and Lactating Status of Does in Eyeball Temperature
4. Discussion
4.1. Correlated Response to Selection in Temperature Emissivity
4.2. Effect of Season and Lactating Status of Does in Eyeball Temperature
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Widowski, T.M.; Curtis, S.E.; Graves, C.N. THE NEUTROPHIL:LYMPHOCYTE RATIO IN PIGS FED CORTISOL. Can. J. Anim. Sci. 1989, 69, 501–504. [Google Scholar] [CrossRef]
- Urbanová, M.; Kramářová, E.; Chloupek, J.; Najmanová, M. Evaluation of stress in laboratory rabbits used for teaching purposes. Acta Vet. Brno 2019, 88, 249–255. [Google Scholar] [CrossRef]
- Stewart, M.; Webster, J.R.; Schaefer, A.L.; Cook, N.J.; Scott, S.L. Infrared thermography as a non-invasive tool to study animal welfare. Anim. Welf. 2005, 14, 319–325. [Google Scholar]
- Ricci, G.D.; Da Silva-Miranda, K.O.; Titto, C.G. Infrared thermography as a non-invasive method for the evaluation of heat stress in pigs kept in pens free of cages in the maternity. Comput. Electron. Agric. 2019, 157, 403–409. [Google Scholar] [CrossRef]
- de Lima, V.; Piles, M.; Rafel, O.; López-Béjar, M.; Ramón, J.; Velarde, A.; Dalmau, A. Use of infrared thermography to assess the influence of high environmental temperature on rabbits. Res. Vet. Sci. 2013, 95, 802–810. [Google Scholar] [CrossRef]
- Marai, I.; Ayyat, M.; El-Monem, U.A. Growth Performance and Reproductive Traits at First Parity of New Zealand White Female Rabbits as Affected by Heat Stress and Its Alleviation under Egyptian Conditions. Trop. Anim. Health Prod. 2001, 33, 451–462. [Google Scholar] [CrossRef] [PubMed]
- McManus, C.; Tanure, C.B.; Peripolli, V.; Seixas, L.; Fischer, V.; Gabbi, A.M.; Menegassi, S.R.O.; Stumpf, M.T.; Kolling, G.J.; Dias, E.; et al. Infrared thermography in animal production: An overview. Comput. Electron. Agric. 2016, 123, 10–16. [Google Scholar] [CrossRef]
- Blasco, A. Bayesian Data Analysis for Animal Scientists; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Beloumi, D.; Blasco, A.; Muelas, R.; Santacreu, M.A.; García, M.D.L.L.; Argente, M.-J. Inflammatory Correlated Response in Two Lines of Rabbit Selected Divergently for Litter Size Environmental Variability. Animals 2020, 10, 1540. [Google Scholar] [CrossRef] [PubMed]
- Argente, M.J.; García, M.L.; Zbyňovská, K.; Petruška, P.; Capcarova, M.; Blasco, A. Correlated response to selection for litter size environmental variability in rabbits’ resilience. Animal 2019, 13, 2348–2355. [Google Scholar] [CrossRef]
- García, M.L.; Blasco, A.; Argente, M.J. Correlated response in body condition and energy mobilisation in rabbits selected for litter size variability. Animal 2019, 13, 784–789. [Google Scholar] [CrossRef]
- Rushen, J.; A Taylor, A.; de Passillé, A.M. Domestic animals’ fear of humans and its effect on their welfare. Appl. Anim. Behav. Sci. 1999, 65, 285–303. [Google Scholar] [CrossRef]
- Gerencsér, Z.; Matics, Z.; Szabó, R.T.; Kustos, K.; Mikó, A.; Nagy, I.; Odermatt, M.; Atkári, T.; Szendrő, Z. Aggressiveness, Mating Behaviour and Lifespan of Group Housed Rabbit Does. Animals 2019, 9, 708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, J.; Baum, M.J. Neuroendocrine Regulation of GnRH Release in Induced Ovulators. Front. Neuroendocr. 2000, 21, 220–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valera, M.; Bartolomé, E.; Sánchez, M.J.; Molina, A.; Cook, N.; Schaefer, A. Changes in Eye Temperature and Stress Assessment in Horses During Show Jumping Competitions. J. Equine Vet. Sci. 2012, 32, 827–830. [Google Scholar] [CrossRef]
- Bartolomé, E.; Sánchez, M.J.; Molina, A.; Schaefer, A.L.; Cervantes, I.; Valera, M. Using eye temperature and heart rate for stress assessment in young horses competing in jumping competitions and its possible influence on sport performance. Animal 2013, 7, 2044–2053. [Google Scholar] [CrossRef]
- Vianna, D.M.L.; Carrive, P. Changes in cutaneous and body temperature during and after conditioned fear to context in the rat. Eur. J. Neurosci. 2005, 21, 2505–2512. [Google Scholar] [CrossRef] [Green Version]
- Marai, I.; Habeeb, A.; Gad, A. Rabbits’ productive, reproductive and physiological performance traits as affected by heat stress: A review. Livest. Prod. Sci. 2002, 78, 71–90. [Google Scholar] [CrossRef]
- Jaén-Téllez, J.A.; Sánchez-Guerrero, M.J.; López-Campos, J.I.; Valera, M.; González-Redondo, P. Acute stress assessment using infrared thermography in fattening rabbits reacting to handling under winter and summer conditions. Span. J. Agric. Res. 2020, 18, e0502. [Google Scholar] [CrossRef]
- Argente, M.-J.; García, M.D.L.L.; Birlanga, V.; Muelas, R. Relationship between cortisol and acute phase protein concentrations in female rabbits. Vet. J. 2014, 202, 172–175. [Google Scholar] [CrossRef]
- Bosco, A.D.; Rebollar, P.; Boiti, C.; Zerani, M.; Castellini, C. Ovulation induction in rabbit does: Current knowledge and perspectives. Anim. Reprod. Sci. 2011, 129, 106–117. [Google Scholar] [CrossRef] [PubMed]
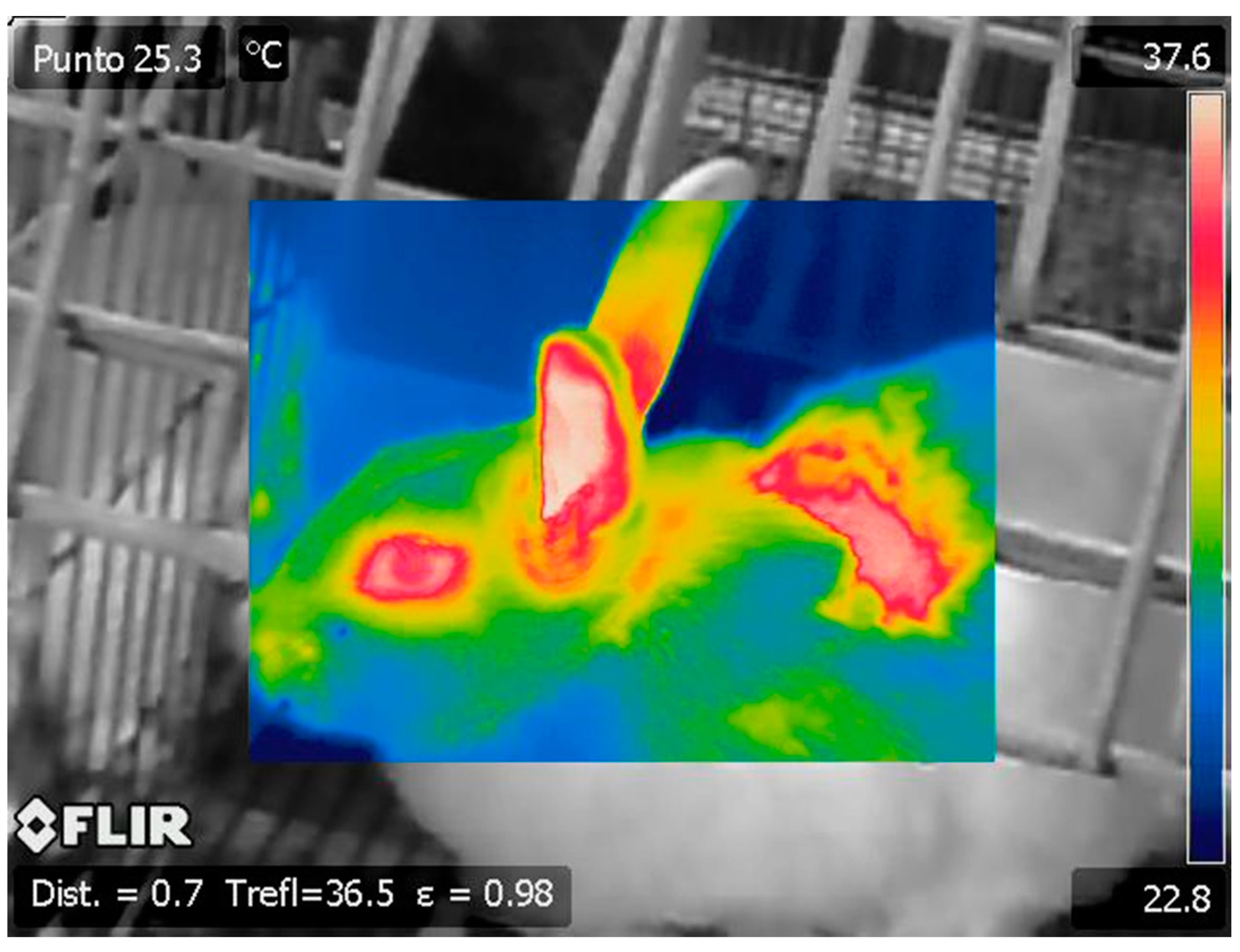

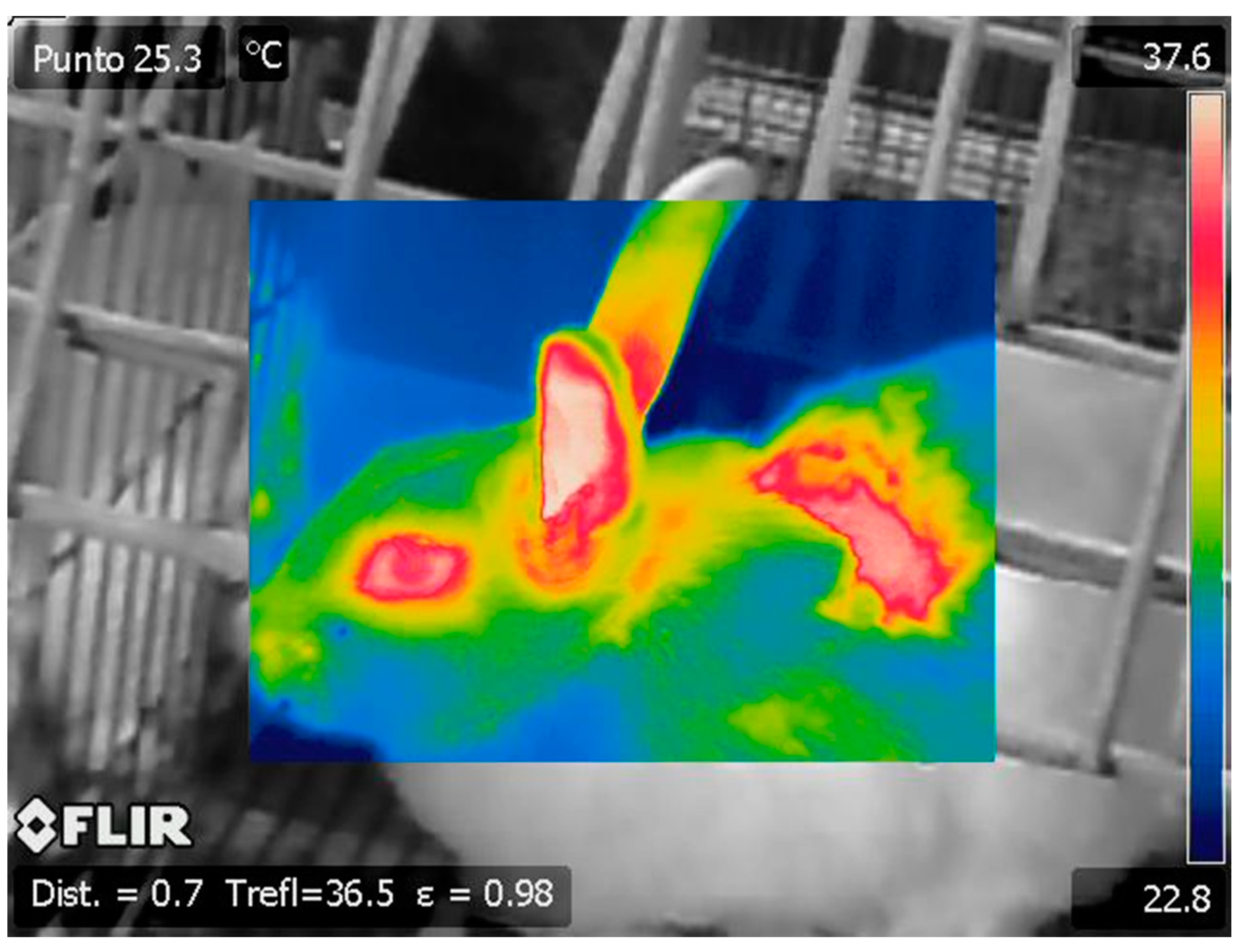{kind=link}
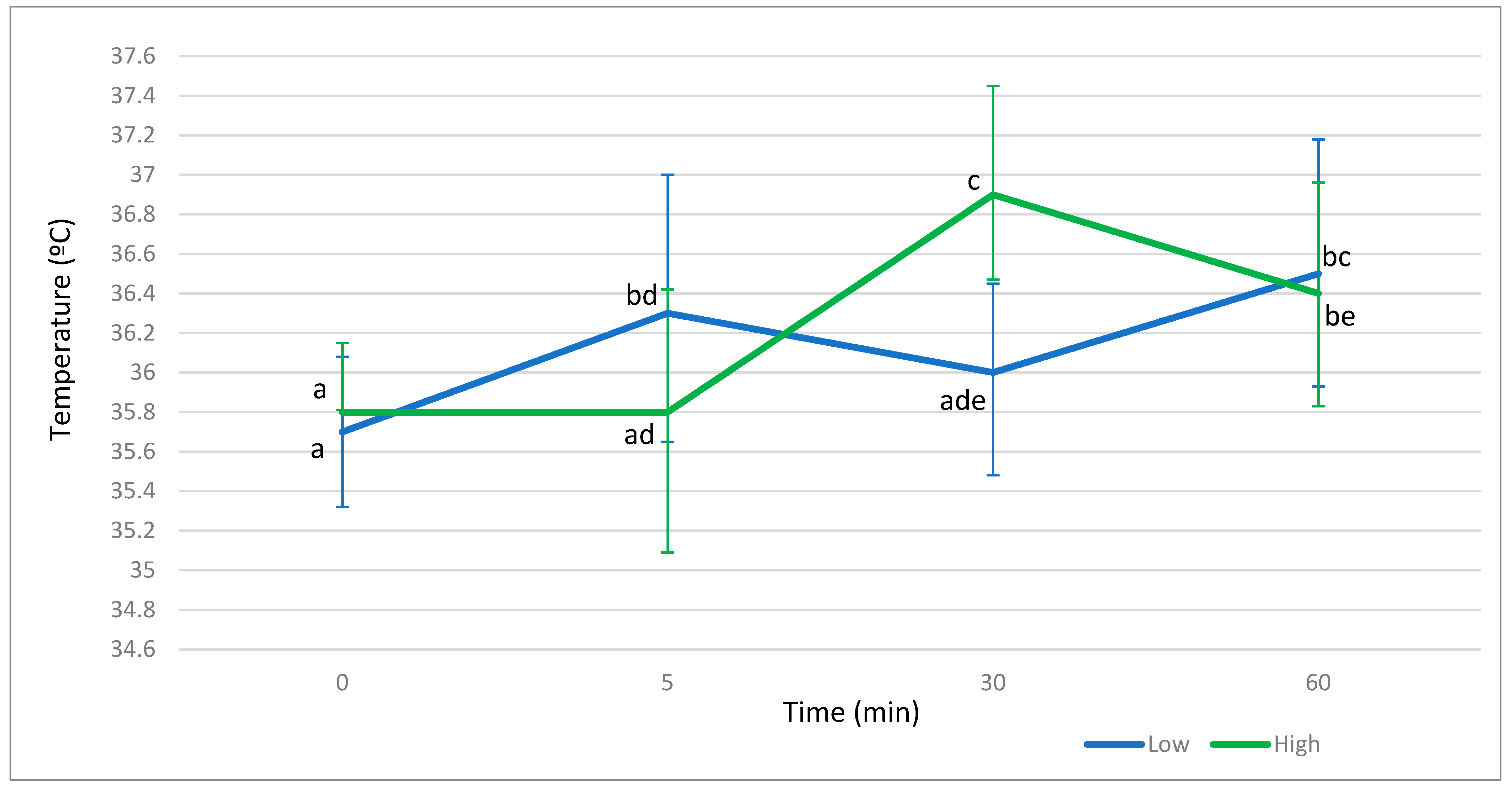
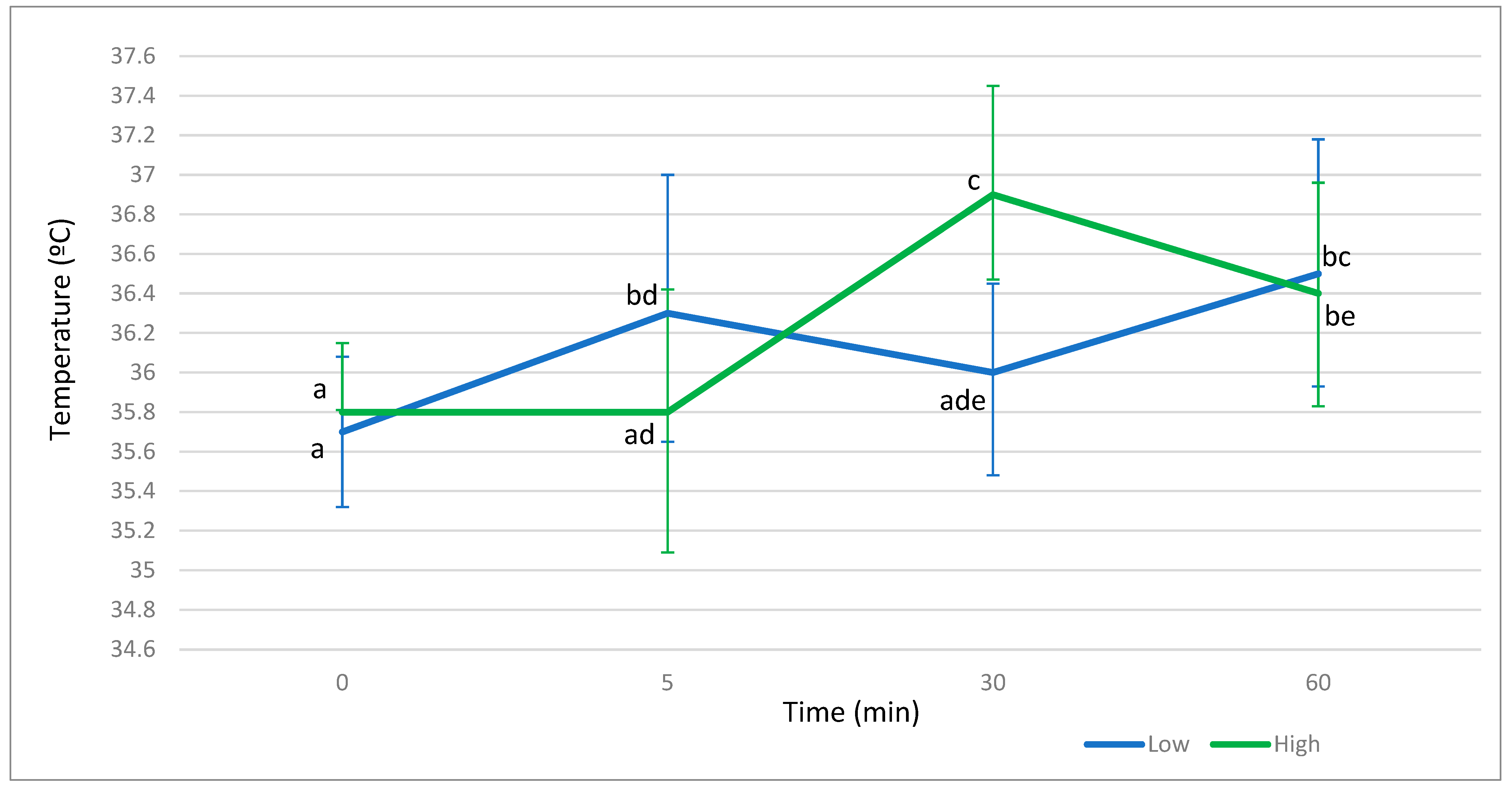{kind=link}
| Environment 1 | Environment 2 | |
|---|---|---|
| Number of sessions | 4 | 7 |
| Number of does | 46 | 47 |
| Mean temperature (°C) | 12.5 | 18.1 |
| Standard deviation (°C) | 0.95 | 1.51 |
| Time (n = 93) | Environment 1 | Environment 2 | DEnv1-Env2 | HPD95% | p |
|---|---|---|---|---|---|
| Basal | 32.19 | 33.28 | −1.09 | −1.78, −0.39 | 1 |
| Basal-60 min | 2.01 | 2.32 | −0.31 | −1.07, 0.44 | 0.79 |
| Time (n = 93) | Non-Lactating (°C) | Lactating (°C) | Dnl-l | HPD95% | p |
|---|---|---|---|---|---|
| Basal | 33.21 | 32.61 | 0.60 | −0.01, 1.27 | 0.97 |
| Basal-60 min | 1.82 | 2.37 | −0.55 | −1.30, 0.25 | 0.92 |
| Temperature (n = 93) | Fertile (°C) | Non-Fertile (°C) | Df-nf | HPD95% | p |
|---|---|---|---|---|---|
| Basal | 33.99 | 32.85 | 0.12 | −0.49, 0.78 | 0.64 |
| Basal-60 min | 1.91 | 2.28 | −0.37 | −1.12, 0.39 | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agea, I.; García, M.d.l.L.; Argente, M.-J. Preliminary Study of Body Temperature Emissivity in Rabbits Selected for Litter Size Residual Variability. Agriculture 2021, 11, 604. https://doi.org/10.3390/agriculture11070604
Agea I, García MdlL, Argente M-J. Preliminary Study of Body Temperature Emissivity in Rabbits Selected for Litter Size Residual Variability. Agriculture. 2021; 11(7):604. https://doi.org/10.3390/agriculture11070604
Chicago/Turabian StyleAgea, Iván, María de la Luz García, and María-José Argente. 2021. "Preliminary Study of Body Temperature Emissivity in Rabbits Selected for Litter Size Residual Variability" Agriculture 11, no. 7: 604. https://doi.org/10.3390/agriculture11070604
APA StyleAgea, I., García, M. d. l. L., & Argente, M.-J. (2021). Preliminary Study of Body Temperature Emissivity in Rabbits Selected for Litter Size Residual Variability. Agriculture, 11(7), 604. https://doi.org/10.3390/agriculture11070604