
Probiotic Delivery through Non-Dairy Plant-Based Food Matrices

 ,
, 

Abstract

1. Introduction
2. Plant-Based Matrices and Their Features

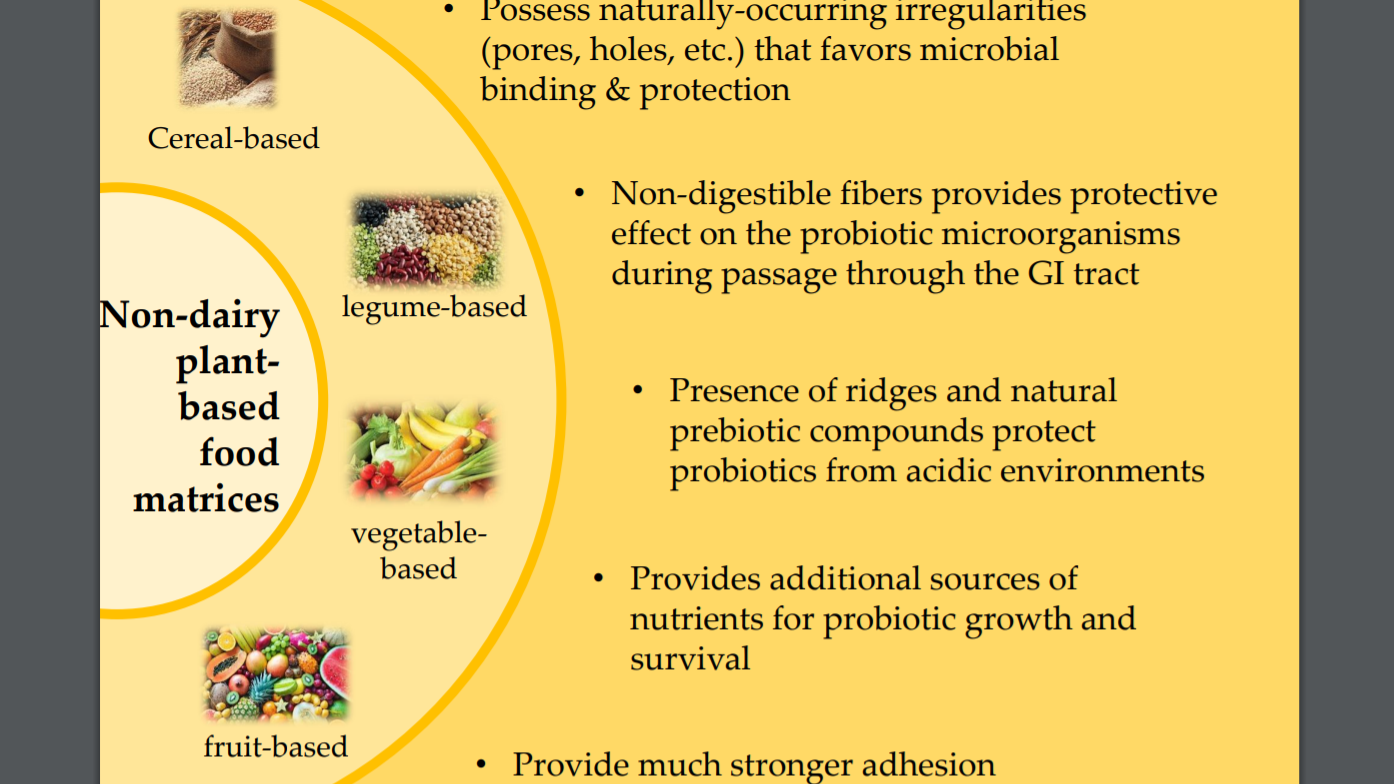{kind=link}
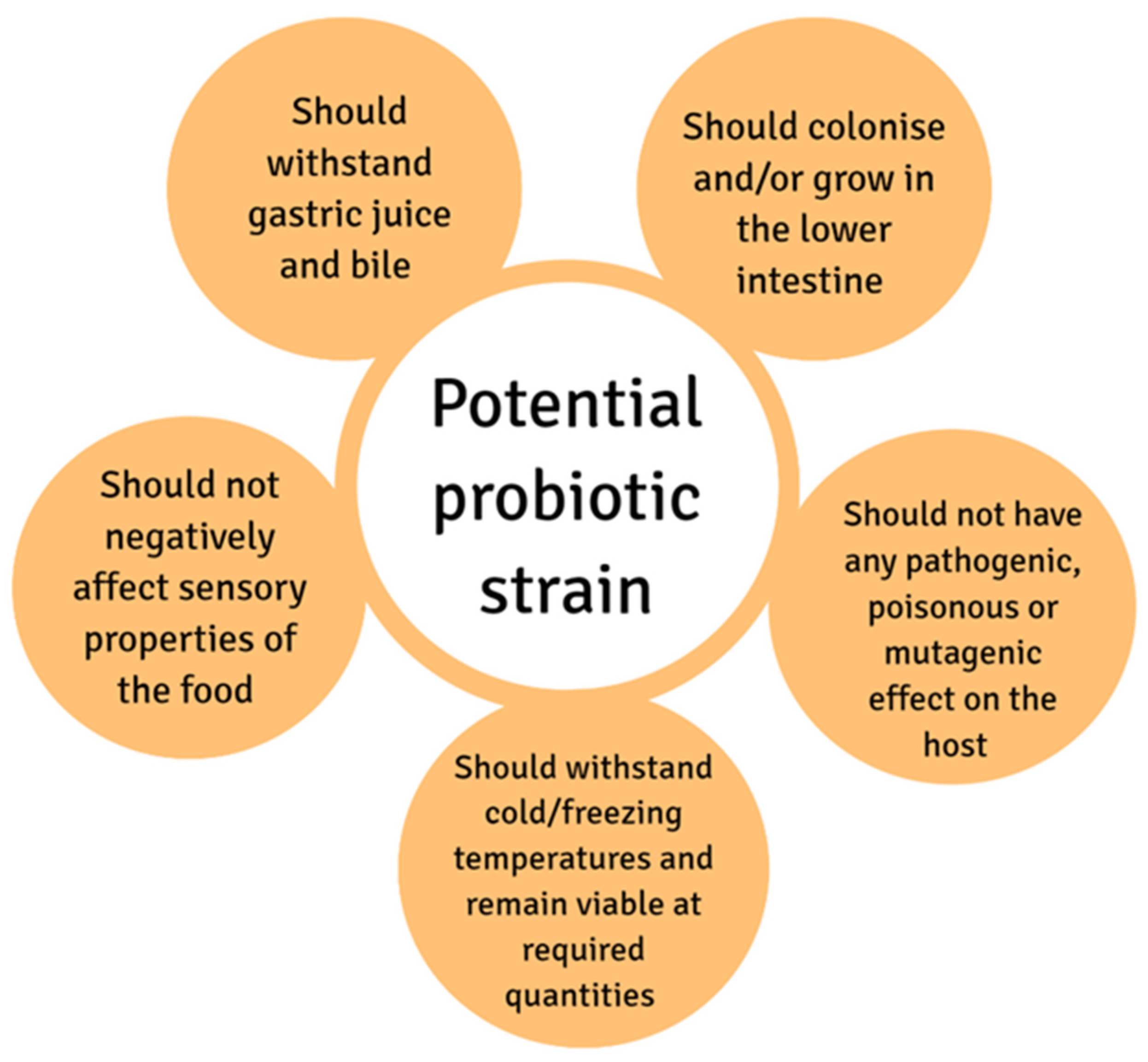{kind=link}
| Type | Name | Principal Raw Material | Geographical Regions in Which the Beverage Is Popular | Probiotic Microorganisms Isolated | References |
|---|---|---|---|---|---|
| Cereal and legume-based | Boza | Wheat, rye, millet mixed with sugar/saccharine | Bulgaria, Albania, Turkey, Romania | LABs: Lctobacillus acidophilus, Lb. fermentum, Lb. coprophilus, Lb. brevis, Leuconostoc reffinolactis, Leuconostoc mesenteroides Yeast: Sachharomyces cerevisiae, Candida tropicalis, Candida glabata, Geotrichum penicilliatum, Geotrichum candidum | [19,20] |
| Bushera | Sorghum or millet flour from germinated sorghum or millet grains | Western highlands of Uganda | Lactobacillus sp., Lactococcus sp., Leuconostoc sp., Enterococcus sp., Streptococcus sp. | [21] | |
| Mahewa/amahewa | Corn meal ferment with sorghum, millet malt or wheat flour | Africa and some Arabian Gulf countries | Lc. lactis ssp. lactis | [19,22] | |
| Pozol | Cooking maize in a lime solution (1% w/v) | South-eastern Mexico | Lactococcus lactis | [23] | |
| Togwa | Maize flour and finger millet | Japan & China | Lactobacillus spp., Streptococcus spp., and Lb. plantarum | [2,24] | |
| Ogi | Maize | Nigeria and west Africa | Lb. acidophilus, Lb. plantarum, Lb. brevis, Lb. fermentum, S. cerevisiae, Rhodotorula graminis, Candida krusei, C. tropicalis, Geotrichum candidum, Geotrichum fermentum | [25,26] | |
| Kefir Soy | Soya beans | Greece | Lb. brevis, Lb. kefir, Lb. mesenteroides, Lb. helveticus, Kluyveromyces maxianus and Kluyveromyces lactis | [9] | |
| Ricera | Rice | Lb. acidophilus, Lb. bulgaricus, B. bifidum, S. thermophilus | |||
| Uji | Maize or sorghum | Kenya | Lb. plantarum, Lb. fermentum, Lb. cellobiaosus, Lb. buchneri, Pediococcus nacidilactice, Pediococcus penosaceus, Lb. rhamnosus, S. thermophilus | [27] | |
| Velli | Oat brans and fruits | Russia | B. bifidum, Lb. acidophilus | [9] | |
| Tempeh | Soymilk | Lb. rhamnosus, Bifidobacterium spp. | [9] | ||
| Vegetable-based | Shalgam | black carrot, bulgur flour, sourdough, salt, turnip, and drinkable water | Southern Turkey | Lb. plantarum, Lb. paracasei subsp. paracasei, Lb. brevis, Lb. fermentum | [9,28,29] |
| Kanji | Black carrot and beetroot | Northern India | Lb. plantarum | [30] | |
| Sayurasin | Mustard and cabbage | Indonesia | Lb. farciminis, Lb. fermentum, Lb. namurensis, Lb. plantarum, Lb. helveticus, Lb. brevis, Lb. versmoldensis, Lb. casei Lb. rhamnosus, Lb. fabifermentans, Lb. satsumensis | [31] | |
| Suan-tsai | Mustard | Taiwan | Pediococcus pentosaceus Tetragenococcus halophilus | [32] | |
| Yan-dong-gua | Wax gourd | Taiwan | Weissela paramesenteroides W. cibaria | [33] | |
| Soido | Bamboo shoots (Melocanna bambusoides, Bambusatulda and Dendrocalamus giganteus) | North-eastern states of India | Lc. lactis, Lb. brevis, Leu. fallax | [34] | |
| Jiang-sun | Bamboo shoots | Taiwan | Lb. plantarum | [35] | |
| Dochi | Black beans | Taiwan | Enterococcus faecium | [36] | |
| Jiang-gua | Cucumber | Taiwan | Wissella cibaria, Leuconostoc lactis | [37] | |
| Dua muoi | Mustard and beet | Vietnam | Lb. fermentum, Lb. pentosus, Lb. plantarum | [38] | |
| Ca muoi | Egg plant | Vietnam | Lb. fermentum, Lb. pentosus, Lb. plantarum | [38] | |
| Nozawana-Zuke | Japan | Lb. curvatus | [39] | ||
| Yan-tsai-shin | Broccoli stems | Taiwan | Lb. plantarum, Enterococcus sulphurous, Weissella paramesenteroides, W. minor, Leuconostoc mesenteroides and W. cibaria | [40] | |
| Chinese sauerkraut | Cabbage | China | Lb. mesenteroides, Lb. plantarum, Lb. brevis, Lb. rhamnosus and Lb. plantarum | [41,42] | |
| Kimchi | Napa cabbage and Korean radish | Korea | Leuconostoc mesenteriodes, Leu. carnosum, Lactobacillus curvatus, Lb. pentosus, Weissela kimchi, W. cibaria and Pediococcus pentosaceus | [43] | |
| Dhamuoi | Cabbage | Nepal and India | Leuconostoc mesenteroides and Lb. plantarum, | [44] | |
| Gundruk | Cabbage | Nepal and India | Pediococcus and Lactobacillus spp. | [44] | |
| Paocai | Cabbage, celery, cucumber and radish | China | L. pentosus, L. plantarum, Leuconostoc mesenteroides, L. brevis, L. lactis, and L. fermentum | [45,46] | |
| Kenkey/kormi/kokoe/dorkunu | Maize, millet, sorghum | Africa | Lb. plantarum, Lb. brevis, Lb. fermentum, Lb. reuteri | [47] | |
| Koozh | Rice and millet flour | South India | W. paramesenteroides, Lb. plantarum, Lb. fermentum | [48] | |
| Fruit-based | Pobuzihi | Cummingcordia/glue berry | Taiwan | Lb. plantarum Enterococcus casseliflavus Wissella cibaria | [49] |
| Xi-gua-mian | Water melon | Taiwan | Lb. plantarum Pediococcus pentosaceus | [50] | |
| Hardaliye | Red grape/red grape enriched with crushed mustard seeds and benzoic acid | Turkey | Lb. paracasei ssp. paracasei Lb. casei ssp. pseudoplantarum Lb. brevis, Lb. pontis, Lb. acetotolerans, Lb. sanfransisco, Lb. vaccinostercus | [51] | |
| Tempoyak | Durian fruit | Malaysia | Lb. mali, Lb. brevis, Lb. mesenteroides, Lb. fermentum | [9] |
3. Non-Dairy Plant-Based Probiotic Products
3.1. Cereal-Based Products
3.1.1. Oat-Based Products
3.1.2. Malt-Based Products
3.1.3. Wheat-Based Products
3.1.4. Rice-Based Products
3.1.5. Maize-Based Products
3.1.6. Millet-Based Products
3.2. Legume-Based Products
3.2.1. Soya-Based Products
3.2.2. Chick-Pea-Based Products
3.2.3. Miscellaneous Legume-Based Products
3.3. Vegetable-Based Products
3.3.1. Carrot-Based Products
3.3.2. Beetroot-Based Products
3.3.3. Cabbage/Broccoli-Based Products
3.3.4. Tomato-Based Products
| Product Type | Product | Probiotic Strains | Viability at the End of Storage | Storage Time | References |
|---|---|---|---|---|---|
| Cereal-based | Fermented oat flour | Lb. plantarum | 108 cfu/g | 21 d at 4 °C | [10] |
| Oat-based drink | Lb. plantarum B28 | 106–107 cfu/mL | 24 d at 4 °C | [56] | |
| Oat bars | B. lactis Bb-12 | 109 cfu/25 g | 7–14 d at 4 °C | [118] | |
| Probiotic fermented oat beverage incorporated with guava, orange & passion fruit by-products | Lb. casei Lc-1 | >108 cfu/mL | 28 d at 4 °C | [119] | |
| Emma beverage | Lb. plantarum Lb. acidophilus | 108 cfu/mL | 30 d at 4 °C | [67] | |
| Yoghurt-like beverages from rice, rice and soy, rice and barley, rice and emmer, rice and oat | Lb. plantarum 6E and 6M | 108 cfu/mL | 30 d at 4 °C | [57] | |
| Milk-based maize/rice pudding | Lb. acidophilus La5 and 1748 B. animalis Bb12, and Lb. rhamnosus GG grown separately | 108–109 cfu/g | 21 d at 4–6°C | [120] | |
| Maize-based fermented beverages | Saccharomyces cerevisiae CCMA 0732, S. cerevisiae CCMA 0731, and Pichia kluyveri CCMA 0615 in combination with Lb. paracasei LBC-81 | >106 cfu/mL | 28 d at 4 °C | [75] | |
| Fermented beverage produced from maize and rice supplemented with fructooligosaccharides | Lactobacillus plantarum CCMA 0743, Torulaspora delbrueckii CCMA 0235, and Lb. acidophilus LACA 4 in mixed culture | ≥107 cfu/mL | 28 d at 4 °C | [76] | |
| Legume-based | Soya frozen dessert | Lb. acidophilus Lb. paracasei B. lactis Lb. rhamnosus S. boulardii | 107 cfu/g ~105 cfu/g | 28 d at −18 °C | [121] |
| Probiotic soymilk | B. breve strain Yakult | 109 cfu/mL | 20 d at 10 °C | [122] | |
| Beverage containing chick-pea & coconut extract in 9:1 ratio | Lb. paracasei LBC 81 | >108 cfu/mL | 10 d at 4 °C | [99] | |
| Probiotic roasted chick-peas | Lb. plantarum 299v | >109 cfu/mL | 3 months at 4 °C | [100] | |
| Vegetable-based | Probiotic beetroot juice | Lb. casei 431 | 106–108 cfu/mL | 28 d at 4 °C | [106] |
| Probiotic cabbage juice | Lb. plantarum Lb. rhamnosus Lb. brevis | >109 cfu/mL >1010 cfu/mL >109 cfu/mL | 30 d at 4 °C | [107] | |
| Probiotic blanched cabbage | Lb. paracasei LMG P22043 | >108 cfu/g | 30 d at 4 °C | [109] | |
| Probiotic cabbage juice | Lb. plantarum C3 Lb. delbrueckii D7 | >107 cfu/mL >105 cfu/mL | 28 d at 4 °C | [108] | |
| Probiotic tomato juice | Lb. acidophilus Lb. casei Lb. plantarum Lb. delbrueckii | 109 cfu/mL 108 cfu/mL 106 cfu/mL 108 cfu/mL | 28 d at 4 °C | [116] | |
| Fruit-based | Probiotic apple juice supplemented with oligofructose | Lb. paracasei ssp. paracasei | >106 cfu/mL | 28 d at 4 °C (in a glass container) | [123] |
| Osmotically dehydrated probiotic cut apple | Lb.plantarum 299v | >107 cfu/mL | 6 d at 4 °C | [124] | |
| Probiotic enriched apple snacks | Lb. plantarum SICC | >106 cfu/g | 120 d at 25 °C | [125] | |
| Probiotic pineapple juice | Meyerozyma caribbica 9 D | 107 cfu/g | 21 d at 4 °C | [126] | |
| Probiotic pineapple juice | Lb. casei NRRL B-442 | >106 cfu/mL | 21 d at 4 °C | [127] | |
| Probiotic mixed pineapple and jussara juice | Lb. rhamnosus GG | >107 cfu/mL | 28 d at 8 °C | [128] | |
| Spray-dried or freeze-dried probiotic orange juice powder | Lb. plantarum 299v P.acidilactici HA-6111-2 | 108 cfu/mL | 180 d at 4 °C | [129] | |
| Synbiotic orange juice | Lb. paracasei ssp. paracasei | >106 cfu/mL | 28 d at 4 °C | [130] | |
| Probiotic orange juice fortified with nettle | Lb. rhamnosus ATCC 53103 | ~107 cfu/mL | 28 d at 4 °C | [131] | |
| Probiotic orange juice | P. acidilactici CE51 | >107 cfu/mL | 35 d at 4 °C | [132] | |
| Probiotic cashew apple juice | Lb. casei NRRL B-442 | >108 cfu/mL | 42 d at 4 °C | [133] | |
| Non-fermented probiotic beverage from Jucara fruit blended with banana and strawberry | Lb. plantarum CNPC003 Lb. casei BGP93 | ~106 cfu/mL >107 cfu/mL | 90 d at 4 °C | [134] | |
| Probiotic Sohiong fruit juice powder | Lb. plantarum | >106 cfu/mL | 36 d at 25 °C | [135] | |
| Probiotic cataloupe juice | Lb. casei NRRL B-442 | >108 cfu/mL | 42 d at 4 °C | [136] | |
| Honeydew melon | Lb. casei NCIMB 4114 | >108 cfu/mL | [137] | ||
| Probiotic pomegranate juice | Lb. plantarum Lb. delbrueckii | Not available | 28 d at 4 °C | [138] | |
| Table olives | Lb. rhamnosus Lb. paracasei B. bifidum B. longum | 106–108 cfu/g | 90 d at 4 °C | [139] |
3.4. Fruit-Based Probiotic Products
3.4.1. Apple-Based Products
3.4.2. Pineapple-Based Products
3.4.3. Orange-Based Products
3.5. Miscellaneous Products
3.5.1. Products from Fruit by-Products
3.5.2. Plant-Based Dairy Alternatives
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ranadheera, C.S.; Naumovski, N.; Ajlouni, S. Non-bovine milk products as emerging probiotic carriers: Recent developments and innovations. Curr. Opin. Food Sci. 2018, 22, 109–114. [Google Scholar] [CrossRef]
- Molin, G. Probiotics in foods not containing milk or milk constituents, with special reference to Lactobacillus plantarum 299v. Am. J. Clin. Nutr. 2001, 73. [Google Scholar] [CrossRef] [PubMed]
- Ranadheera, C.S.; Vidanarachchi, J.K.; Rocha, R.S.; Cruz, A.G.; Ajlouni, S. Probiotic delivery through fermentation: Dairy vs. non-dairy beverages. Fermentation 2017, 3, 67. [Google Scholar] [CrossRef]
- Joint FAO/WHO Working Group. Report on Drafting Guidelines for the Evaluation of Probiotics in Food; WHO: London, OT, Canada, 2002. [Google Scholar]
- Peres, C.M.; Peres, C.; Hernández-Mendoza, A.; Malcata, F.X. Review on fermented plant materials as carriers and sources of potentially probiotic lactic acid bacteria—With an emphasis on table olives. Trends Food Sci. Technol. 2012, 26, 31–42. [Google Scholar] [CrossRef]
- Marques, T.M.; Cryan, J.F.; Shanahan, F.; Fitzgerald, G.F.; Ross, R.P.; Dinan, T.G.; Stanton, C. Gut microbiota modulation and implications for host health: Dietary strategies to influence the gut-brain axis. Innov. Food Sci. Emerg. Technol. 2014, 22, 239–247. [Google Scholar] [CrossRef]
- Ranadheera, C.S.; Evans, C.A.; Baines, S.K.; Balthazar, C.F.; Cruz, A.G.; Esmerino, E.A.; Freitas, M.Q.; Pimentel, T.C.; Wittwer, A.E.; Naumovski, N.; et al. Probiotics in Goat Milk Products: Delivery Capacity and Ability to Improve Sensory Attributes. Compr. Rev. Food Sci. Food Saf. 2019. [Google Scholar] [CrossRef]
- Mattia, A.; Merker, R. Regulation of probiotic substances as ingredients in foods: Premarket approval or “generally recognized as safe” notification. Clin. Infect. Dis. 2008, 46. [Google Scholar] [CrossRef]
- Panghal, A.; Janghu, S.; Virkar, K.; Gat, Y.; Kumar, V.; Chhikara, N. Potential non-dairy probiotic products—A healthy approach. Food Biosci. 2018, 21, 80–89. [Google Scholar] [CrossRef]
- Russo, P.; de Chiara, M.L.V.; Capozzi, V.; Arena, M.P.; Amodio, M.L.; Rascón, A.; Dueñas, M.T.; López, P.; Spano, G. Lactobacillus plantarum strains for multifunctional oat-based foods. LWT Food Sci. Technol. 2016, 68, 288–294. [Google Scholar] [CrossRef]
- Haas, R.; Schnepps, A.; Pichler, A.; Meixner, O. Cow milk versus plant-based milk substitutes: A comparison of product image and motivational structure of consumption. Sustainability 2019, 11, 5046. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Domínguez, R.; Budaraju, S.; Roselló-Soto, E.; Barba, F.J.; Mallikarjunan, K.; Roohinejad, S.; Lorenzo, J.M. Effect of innovative food processing technologies on the physicochemical and nutritional properties and quality of non-dairy plant-based beverages. Foods 2020, 9, 288. [Google Scholar] [CrossRef]
- Martins, E.M.F.; Ramos, A.M.; Vanzela, E.S.L.; Stringheta, P.C.; de Oliveira Pinto, C.L.; Martins, J.M. Products of vegetable origin: A new alternative for the consumption of probiotic bacteria. Food Res. Int. 2013, 51, 764–770. [Google Scholar] [CrossRef]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Çetin, B. Production of probiotic mixed pickles (turşu) and microbiological properties. Afr. J. Biotechnol. 2011, 10, 14926–14931. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends Food Sci. Technol. 2005, 16, 57–69. [Google Scholar] [CrossRef]
- Sapers, G.M. Efficacy of Washing and Sanitizing Methods for Disinfection of Fresh Fruit and Vegetable Products. Food Technol. Biotechnol. 2001, 39, 305–311. [Google Scholar]
- Ranadheera, R.D.C.S.; Baines, S.K.; Adams, M.C. Importance of food in probiotic efficacy. Food Res. Int. 2010, 43, 1–7. [Google Scholar] [CrossRef]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Gotcheva, V.; Pandiella, S.S.; Angelov, A.; Roshkova, Z.G.; Webb, C. Microflora identification of the Bulgarian cereal-based fermented beverage boza. Process Biochem. 2000, 36, 127–130. [Google Scholar] [CrossRef]
- Muyanja, C.M.B.K.; Narvhus, J.A.; Treimo, J.; Langsrud, T. Isolation, characterisation and identification of lactic acid bacteria from bushera: A Ugandan traditional fermented beverage. Int. J. Food Microbiol. 2003, 80, 201–210. [Google Scholar] [CrossRef]
- Gadaga, T.H.; Mutukumira, A.N.; Narvhus, J.A.; Feresu, S.B. A review of traditional fermented foods and beverages of Zimbabwe. Int. J. Food Microbiol. 1999, 53, 1–11. [Google Scholar] [CrossRef]
- Wacher, C.; Cañas, A.; Bárzana, E.; Lappe, P.; Ulloa, M.; Owens, J.D. Microbiology of Indian and Mestizo pozol fermentations. Food Microbiol. 2000, 17, 251–256. [Google Scholar] [CrossRef]
- Oi, Y.; Kitabatake, N. Chemical Composition of an East African Traditional Beverage, Togwa. J. Agric. Food Chem. 2003, 51, 7024–7028. [Google Scholar] [CrossRef]
- David, O.M.; Famurewa, O. Prophylactic and bio-therapeutic benefits of ‘ogi’: A lactic acid fermented food. Bull. Biol. Sci. 2010, 2, 72–77. [Google Scholar]
- Omemu, A.M. Fermentation dynamics during production of ogi, a Nigerian fermented cereal porridge. Rep. Opin. 2011, 3, 8–17. [Google Scholar]
- Kort, R.; Westerik, N.; Mariela Serrano, L.; Douillard, F.P.; Gottstein, W.; Mukisa, I.M.; Tuijn, C.J.; Basten, L.; Hafkamp, B.; Meijer, W.C.; et al. A novel consortium of Lactobacillus rhamnosus and Streptococcus thermophilus for increased access to functional fermented foods. Microb. Cell Fact. 2015, 14. [Google Scholar] [CrossRef]
- Erten, H.; Tanguler, H.; Canbaş, A. A traditional Turkish lactic acid fermented beverage: Shalgam (Salgam). Food Rev. Int. 2008, 24, 352–359. [Google Scholar] [CrossRef]
- Tanguler, H.; Erten, H. Occurrence and growth of lactic acid bacteria species during the fermentation of shalgam (salgam), a traditional Turkish fermented beverage. LWT Food Sci. Technol. 2012, 46, 36–41. [Google Scholar] [CrossRef]
- Lamba, J.; Goomer, S.; Saxena, S.K. Study the lactic acid bacteria content in traditional fermented Indian drink: Kanji. Int. J. Gastron. Food Sci. 2019, 16. [Google Scholar] [CrossRef]
- Sukara, E.; Salamah, A.; Dinoto, A.; Mangunwardoyo, W. Identification of lactic acid bacteria in sayur asin from Central Java (Indonesia) based on 16S rDNA sequence. Int. Food Res. J. 2014, 21, 527–532. [Google Scholar]
- Chen, Y.S.; Yanagida, F.; Hsu, J.S. Isolation and characterization of lactic acid bacteria from suan-tsai (fermented mustard), a traditional fermented food in Taiwan. J. Appl. Microbiol. 2006, 101, 125–130. [Google Scholar] [CrossRef]
- Lan, W.T.; Chen, Y.S.; Yanagida, F. Isolation and characterization of lactic acid bacteria from Yan-dong-gua (fermented wax gourd), a traditional fermented food in Taiwan. J. Biosci. Bioeng. 2009, 108, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P. Himalayan Fermented Foods: Microbiology, Nutrition, and Ethnic Values; CRC Press: Boca Raton, FL, USA, 2009; ISBN 9781420093254. [Google Scholar]
- Chen, Y.S.; Wu, H.C.; Liu, C.H.; Chen, H.C.; Yanagida, F. Isolation and characterization of lactic acid bacteria from jiang-sun (fermented bamboo shoots), a traditional fermented food in Taiwan. J. Sci. Food Agric. 2010, 90, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Yanagida, F.; Hsu, J.S. Isolation and characterization of lactic acid bacteria from dochi (fermented black beans), a traditional fermented food in Taiwan. Lett. Appl. Microbiol. 2006, 43, 229–235. [Google Scholar] [CrossRef]
- Chen, Y.S.; Wu, H.C.; Lo, H.Y.; Lin, W.C.; Hsu, W.H.; Lin, C.W.; Lin, P.Y.; Yanagida, F. Isolation and characterisation of lactic acid bacteria from Jiang-gua (fermented cucumbers), a traditional fermented food in Taiwan. J. Sci. Food Agric. 2012, 92, 2069–2075. [Google Scholar] [CrossRef]
- Nguyen, D.T.L.; Van Hoorde, K.; Cnockaert, M.; De Brandt, E.; Aerts, M.; Binh Thanh, L.; Vandamme, P. A description of the lactic acid bacteria microbiota associated with the production of traditional fermented vegetables in Vietnam. Int. J. Food Microbiol. 2013, 163, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Iida, A.; Toyama, Y.; Fukuda, K. Characterization of the bacteriocinogenic lactic acid bacteria lactobacillus curvatus strain YI08 isolated from nozawana-zuke pickles. Food Sci. Technol. Res. 2010, 16, 253–262. [Google Scholar] [CrossRef][Green Version]
- Chen, Y.S.; Liou, M.S.; Ji, S.H.; Yu, C.R.; Pan, S.F.; Yanagida, F. Isolation and characterization of lactic acid bacteria from Yan-tsai-shin (fermented broccoli stems), a traditional fermented food in Taiwan. J. Appl. Microbiol. 2013, 115, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, X.; Li, D.; Niu, C.; Yang, Z. Study on the identification and resistant properties of Lactobacillus plantarum isolated from sauerkraut. Food Sci. Technol. 2010, 31, 141–144. [Google Scholar]
- Yang, Z.; Li, S.; Zhang, X.; Zeng, X.; Li, D.; Zhao, Y.; Zhang, J. Capsular and slime-polysaccharide production by Lactobacillus rhamnosus JAAS8 isolated from Chinese sauerkraut: Potential application in fermented milk products. J. Biosci. Bioeng. 2010, 110, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.K.; Han, T.W.; Shin, H.Y.; Jin, I.N.; Ghim, S.Y. Diversity and antibacterial activity of lactic acid bacteria isolated from Kimchi. Korean J. Microbiol. Biotechnol. 2003, 31, 191–196. [Google Scholar]
- Dahal, N.R.; Karki, T.B.; Swamylingappa, B.; Li, Q.; Gu, G. Traditional foods and beverages of Nepal-a review. Food Rev. Int. 2005, 21, 1–25. [Google Scholar] [CrossRef]
- Feng, M.; Chen, X.; Li, C.; Nurgul, R.; Dong, M. Isolation and Identification of an Exopolysaccharide-Producing Lactic Acid Bacterium Strain from Chinese Paocai and Biosorption of Pb(II) by Its Exopolysaccharide. J. Food Sci. 2012, 77. [Google Scholar] [CrossRef]
- Yan, P.M.; Xue, W.T.; Tan, S.S.; Zhang, H.; Chang, X.H. Effect of inoculating lactic acid bacteria starter cultures on the nitrite concentration of fermenting Chinese paocai. Food Control 2008, 19, 50–55. [Google Scholar] [CrossRef]
- Nout, M.J.R. Rich nutrition from the poorest—Cereal fermentations in Africa and Asia. Food Microbiol. 2009, 26, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.S.; Varman, D.R.; Kanmani, P.; Yuvaraj, N.; Paari, K.A.; Pattukumar, V.; Arul, V. Isolation, Characterization and Identification of a Potential Probiont from South Indian Fermented Foods (Kallappam, Koozh and Mor Kuzhambu) and Its Use as Biopreservative. Probiotics Antimicrob. Proteins 2010, 2, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-S.; Wu, H.-C.; Wang, C.-M.; Lin, C.-C.; Jhong, Y.-J.; Yanagida, F. Isolation and characterization of lactic acid bacteria from pobuzihi (fermented cummingcordia), a traditional fermented food in Taiwan. Folia Microbiol. 2012, 58, 103–109. [Google Scholar] [CrossRef]
- Chen, Y.S.; Wu, H.C.; Yu, C.R.; Chen, Z.Y.; Lu, Y.C.; Yanagida, F. Isolation and characterization of lactic acid bacteria from xi-gua-mian (fermented watermelon), a traditional fermented food in Taiwan. Ital. J. Food Sci. 2016, 28, 9–14. [Google Scholar] [CrossRef]
- Arici, M.; Coskun, F. Hardaliye: Fermented grape juice as a traditional Turkish beverage. Food Microbiol. 2001, 18, 417–421. [Google Scholar] [CrossRef]
- Shori, A.B. Influence of food matrix on the viability of probiotic bacteria: A review based on dairy and non-dairy beverages. Food Biosci. 2016, 13, 1–8. [Google Scholar] [CrossRef]
- Vasudha, S.; Mishra, H.N. Non-dairy probiotic beverages. Int. Food Res. J. 2013, 20, 7–15. [Google Scholar]
- Charalampopoulos, D.; Wang, R.; Pandiella, S.S.; Webb, C. Application of cereals and cereal components in functional foods: A review. Int. J. Food Microbiol. 2002, 79, 131–141. [Google Scholar] [CrossRef]
- Lamsal, B.P.; Faubion, J.M. The beneficial use of cereal and cereal components in probiotic foods. Food Rev. Int. 2009, 25, 103–114. [Google Scholar] [CrossRef]
- Angelov, A.; Gotcheva, V.; Kuncheva, R.; Hristozova, T. Development of a new oat-based probiotic drink. Int. J. Food Microbiol. 2006, 112, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Lanera, A.; Trani, A.; Gobbetti, M.; Di Cagno, R. Yogurt-like beverages made of a mixture of cereals, soy and grape must: Microbiology, texture, nutritional and sensory properties. Int. J. Food Microbiol. 2012, 155, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Rathore, S.; Salmerón, I.; Pandiella, S.S. Production of potentially probiotic beverages using single and mixed cereal substrates fermented with lactic acid bacteria cultures. Food Microbiol. 2012, 30, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; López, P.; Capozzi, V.; de Palencia, P.F.; Dueñas, M.T.; Spano, G.; Fiocco, D. Beta-glucans improve growth, viability and colonization of probiotic microorganisms. Int. J. Mol. Sci. 2012, 13, 6026–6039. [Google Scholar] [CrossRef]
- Capozzi, V.; Menga, V.; Digesù, A.M.; De Vita, P.; Van Sinderen, D.; Cattivelli, L.; Fares, C.; Spano, G. Biotechnological production of vitamin B2-enriched bread and pasta. J. Agric. Food Chem. 2011, 59, 8013–8020. [Google Scholar] [CrossRef]
- Kedia, G.; Vázquez, J.A.; Charalampopoulos, D.; Pandiella, S.S. In vitro fermentation of oat bran obtained by debranning with a mixed culture of human fecal bacteria. Curr. Microbiol. 2009, 58, 338–342. [Google Scholar] [CrossRef]
- Bazzano, L.A.; He, J.; Ogden, L.G.; Loria, C.M.; Whelton, P.K. Dietary fiber intake and reduced risk of coronary heart disease in US men and women: The National Health and Nutrition Examination Survey I Epidemiologic Follow-up Study. Arch. Intern. Med. 2003, 163, 1897–1904. [Google Scholar] [CrossRef]
- Martensson, O.; Staaf, J.; Duenas-Chasco, M.; Irastorza, A.; Holst, O. A fermented, ropy, non-dairy oat product based on the exopolysaccharide-producing strain Pediococcus damnosus. Adv. Food Sci. 2002, 24, 4–11. [Google Scholar]
- Mårtensson, O.; Biörklund, M.; Lambo, A.M.; Dueñas-Chasco, M.; Irastorza, A.; Holst, O.; Norin, E.; Welling, G.; Öste, R.; Önning, G. Fermented, ropy, oat-based products reduce cholesterol levels and stimulate the bifidobacteria flora in humans. Nutr. Res. 2005, 25, 429–442. [Google Scholar] [CrossRef]
- Salazar, N.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Exopolysaccharides Produced by Lactic Acid Bacteria and Bifidobacteria as Fermentable Substrates by the Intestinal Microbiota. Crit. Rev. Food Sci. Nutr. 2016, 56, 1440–1453. [Google Scholar] [CrossRef]
- Rozada-Sánchez, R.; Sattur, A.P.; Thomas, K.; Pandiella, S.S. Evaluation of Bifidobacterium spp. for the production of a potentially probiotic malt-based beverage. Process Biochem. 2008, 43, 848–854. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Trani, A.; Gobbetti, M. Manufacture and characterization of functional emmer beverages fermented by selected lactic acid bacteria. Food Microbiol. 2011, 28, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Battistini, C.; Gullón, B.; Ichimura, E.S.; Gomes, A.M.P.; Ribeiro, E.P.; Kunigk, L.; Moreira, J.U.V.; Jurkiewicz, C. Development and characterization of an innovative synbiotic fermented beverage based on vegetable soybean. Braz. J. Microbiol. 2018, 49, 303–309. [Google Scholar] [CrossRef]
- Charalampopoulos, D.; Pandiella, S.S.; Webb, C. Evaluation of the effect of malt, wheat and barley extracts on the viability of potentially probiotic lactic acid bacteria under acidic conditions. Int. J. Food Microbiol. 2003, 82, 133–141. [Google Scholar] [CrossRef]
- Tian, W.; Hu, R.; Chen, G.; Zhang, Y.; Wang, W.; Li, Y. Potential bioaccessibility of phenolic acids in whole wheat products during in vitro gastrointestinal digestion and probiotic fermentation. Food Chem. 2021, 362, 130135. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; Ray, M.; Adak, A.; Halder, S.K.; Das, A.; Jana, A.; Parua (Mondal), S.; Vágvölgyi, C.; Das Mohapatra, P.K.; Pati, B.R.; et al. Role of probiotic Lactobacillus fermentum KKL1 in the preparation of a rice based fermented beverage. Bioresour. Technol. 2015, 188, 161–168. [Google Scholar] [CrossRef]
- Borah, T.; Gogoi, B.; Khataniar, A.; Gogoi, M.; Das, A.; Borah, D. Probiotic characterization of indigenous Bacillus velezensis strain DU14 isolated from Apong, a traditionally fermented rice beer of Assam. Biocatal. Agric. Biotechnol. 2019, 18, 101008. [Google Scholar] [CrossRef]
- Nath, S.; Roy, M.; Sikidar, J.; Deb, B.; Sharma, I.; Guha, A. Characterization and in-vitro screening of probiotic potential of novel Weissella confusa strain GCC_19R1 isolated from fermented sour rice. Curr. Res. Biotechnol. 2021, 3, 99–108. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, H.; Yan, X.; Zhao, S. Preparation of a probiotic rice tablet: Sensory evaluation and antioxidant activity during gastrointestinal digestion. LWT 2020, 124, 108911. [Google Scholar] [CrossRef]
- Menezes, A.G.T.; Ramos, C.L.; Dias, D.R.; Schwan, R.F. Combination of probiotic yeast and lactic acid bacteria as starter culture to produce maize-based beverages. Food Res. Int. 2018, 111, 187–197. [Google Scholar] [CrossRef]
- Freire, A.L.; Ramos, C.L.; Schwan, R.F. Effect of symbiotic interaction between a fructooligosaccharide and probiotic on the kinetic fermentation and chemical profile of maize blended rice beverages. Food Res. Int. 2017, 100, 698–707. [Google Scholar] [CrossRef]
- Helland, M.H.; Wicklund, T.; Narvhus, J.A. Growth and metabolism of selected strains of probiotic bacteria, in maize porridge with added malted barley. Int. J. Food Microbiol. 2004, 91, 305–313. [Google Scholar] [CrossRef]
- Palaniswamy, S.K.; Govindaswamy, V. In-vitro probiotic characteristics assessment of feruloyl esterase and glutamate decarboxylase producing Lactobacillus spp. isolated from traditional fermented millet porridge (kambu koozh). LWT Food Sci. Technol. 2016, 68, 208–216. [Google Scholar] [CrossRef]
- Oh, Y.J.; Jung, D.S. Evaluation of probiotic properties of Lactobacillus and Pediococcus strains isolated from Omegisool, a traditionally fermented milletalcoholic beverage in Korea. LWT Food Sci. Technol. 2015, 63, 437–444. [Google Scholar] [CrossRef]
- Oh, Y.J.; Lee, Y.N.; Hong, C.R.; Moon, G.E.; Jung, D.S. Effect of Aging Temperatures on the Quality of Omegisool, a Traditionally Fermented Alcohol Beverage in Jeju. Natl. Sci. Inst. Seoul Women’s Univ. 2012, 24, 151–158. [Google Scholar]
- Swieca, M.; Kordowska-Wiater, M.; Pytka, M.; Gawlik-Dziki, U.; Seczyk, L.; Złotek, U.; Kapusta, I. Nutritional and pro-health quality of lentil and adzuki bean sprouts enriched with probiotic yeast Saccharomyces cerevisiae var. boulardii. LWT 2019, 100, 220–226. [Google Scholar] [CrossRef]
- Tsai, T.Y.; Chen, L.Y.; Pan, T.M. Effect of probiotic-fermented, genetically modified soy milk on hypercholesterolemia in hamsters. J. Microbiol. Immunol. Infect. 2014, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bedani, R.; Rossi, E.A.; Saad, S.M.I. Impact of inulin and okara on Lactobacillus acidophilus La-5 and Bifidobacterium animalis Bb-12 viability in a fermented soy product and probiotic survival under in vitro simulated gastrointestinal conditions. Food Microbiol. 2013, 34, 382–389. [Google Scholar] [CrossRef]
- Champagne, C.P.; Green-Johnson, J.; Raymond, Y.; Barrette, J.; Buckley, N. Selection of probiotic bacteria for the fermentation of a soy beverage in combination with Streptococcus thermophilus. Food Res. Int. 2009, 42, 612–621. [Google Scholar] [CrossRef]
- Donkor, O.N.; Henriksson, A.; Vasiljevic, T.; Shah, N.P. α-Galactosidase and proteolytic activities of selected probiotic and dairy cultures in fermented soymilk. Food Chem. 2007, 104, 10–20. [Google Scholar] [CrossRef]
- Farnworth, E.R.; Mainville, I.; Desjardins, M.P.; Gardner, N.; Fliss, I.; Champagne, C. Growth of probiotic bacteria and bifidobacteria in a soy yogurt formulation. Int. J. Food Microbiol. 2007, 116, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, Z.; Zhang, Q.; Yan, L.; Chen, W.; Liu, X.M.; Zhang, H.P. Fermentation characteristics and transit tolerance of probiotic Lactobacillus casei Zhang in soymilk and bovine milk during storage. J. Dairy Sci. 2009, 92, 2468–2476. [Google Scholar] [CrossRef]
- WaiYee, F.; MinTze, L. Evaluation of proteolytic and ACE-inhibitory activity of Lactobacillus acidophilus in soy whey growth medium via response surface methodology. LWT Food Sci. Technol. 2010, 43, 563–567. [Google Scholar]
- Costa, K.K.F.D.; Soares Júnior, M.S.; Rosa, S.I.R.; Caliari, M.; Pimentel, T.C. Changes of probiotic fermented drink obtained from soy and rice byproducts during cold storage. LWT Food Sci. Technol. 2017, 78, 23–30. [Google Scholar] [CrossRef]
- Chen, M.; Mustapha, A. Survival of freeze-dried microcapsules of α-galactosidase producing probiotics in a soy bar matrix. Food Microbiol. 2012, 30, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Marazza, J.A.; Nazareno, M.A.; de Giori, G.S.; Garro, M.S. Enhancement of the antioxidant capacity of soymilk by fermentation with Lactobacillus rhamnosus. J. Funct. Foods 2012, 4, 594–601. [Google Scholar] [CrossRef]
- Singh, B.P.; Vij, S. Growth and bioactive peptides production potential of Lactobacillus plantarum strain C2 in soy milk: A LC-MS/MS based revelation for peptides biofunctionality. LWT Food Sci. Technol. 2017, 86, 293–301. [Google Scholar] [CrossRef]
- Miraghajani, M.; Zaghian, N.; Mirlohi, M.; Feizi, A.; Ghiasvand, R. The Impact of Probiotic Soy Milk Consumption on Oxidative Stress Among Type 2 Diabetic Kidney Disease Patients: A Randomized Controlled Clinical Trial. J. Ren. Nutr. 2017, 27, 317–324. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Marchesin, J.; Celiberto, L.S.; Orlando, A.B.; de Medeiros, A.I.; Pinto, R.A.; Zuanon, J.A.S.; Spolidorio, L.C.; dos Santos, A.; Taranto, M.P.; Cavallini, D.C.U. A soy-based probiotic drink modulates the microbiota and reduces body weight gain in diet-induced obese mice. J. Funct. Foods 2018, 48, 302–313. [Google Scholar] [CrossRef]
- Mishra Pandey, S.; Mishra, H.N. Optimization of the prebiotic & probiotic concentration and incubation temperature for the preparation of synbiotic soy yoghurt using response surface methodology. LWT Food Sci. Technol. 2015, 62, 458–467. [Google Scholar] [CrossRef]
- Norouzi, S.; Pourjafar, H.; Ansari, F.; Homayouni, A. A Survey on the survival of Lactobacillus paracasei in fermented and non-fermented frozen soy dessert. Biocatal. Agric. Biotechnol. 2019, 21. [Google Scholar] [CrossRef]
- Albuquerque, M.A.C.; Yamacita, D.S.; Bedani, R.; LeBlanc, J.G.; Saad, S.M.I. Influence of passion fruit by-product and fructooligosaccharides on the viability of Streptococcus thermophilus TH-4 and Lactobacillus rhamnosus LGG in folate bio-enriched fermented soy products and their effect on probiotic survival and folate bio-accessibility under in vitro simulated gastrointestinal conditions. Int. J. Food Microbiol. 2019, 292, 126–136. [Google Scholar]
- Hussein, H.; Awad, S.; El-Sayed, I.; Ibrahim, A. Impact of chickpea as prebiotic, antioxidant and thickener agent of stirred bio-yoghurt. Ann. Agric. Sci. 2020, 65, 49–58. [Google Scholar] [CrossRef]
- Mesquita, M.C.; Leandro, E.D.S.; de Alencar, E.R.; Botelho, R.B.A. Fermentation of chickpea (Cicer arietinum L.) and coconut (Coccus nucifera L.) beverages by Lactobacillus paracasei subsp paracasei LBC 81: The influence of sugar content on growth and stability during storage. LWT 2020, 132, 109834. [Google Scholar] [CrossRef]
- Paredes-Toledo, J. Roasted chickpeas as a probiotic carrier to improve L. plantarum 299v survival during storage. LWT 2021, 146. [Google Scholar] [CrossRef]
- Mridula, D.; Sharma, M. Development of non-dairy probiotic drink utilizing sprouted cereals, legume and soymilk. LWT Food Sci. Technol. 2015, 62, 482–487. [Google Scholar] [CrossRef]
- Swieca, M.; Gawlik-Dziki, U.; Jakubczyk, A.; Bochnak, J.; Sikora, M.; Suliburska, J. Nutritional quality of fresh and stored legumes sprouts—Effect of Lactobacillus plantarum 299v enrichment. Food Chem. 2019, 288, 325–332. [Google Scholar] [CrossRef]
- Wu, H.; Rui, X.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Mung bean (Vigna radiata) as probiotic food through fermentation with Lactobacillus plantarum B1-6. LWT Food Sci. Technol. 2015, 63, 445–451. [Google Scholar] [CrossRef]
- Romero-Espinoza, A.M.; Serna-Saldivar, S.O.; Vintimilla-Alvarez, M.C.; Briones-García, M.; Lazo-Vélez, M.A. Effects of fermentation with probiotics on anti-nutritional factors and proximate composition of lupin (Lupinus mutabilis sweet). LWT 2020, 130, 109658. [Google Scholar] [CrossRef]
- Panghal, A.; Virkar, K.; Kumar, V.; Dhull, S.B.; Gat, Y.; Chhikara, N. Development of probiotic beetroot drink. Curr. Res. Nutr. Food Sci. 2017, 5, 257–262. [Google Scholar] [CrossRef]
- Gamage, S.M.; Mihirani, M.K.S.; Perera, O.D.A.N.; Weerahewa, H.L.D. Development of synbiotic beverage from beetroot juice using beneficial probiotic Lactobacillus Casei 431. Ruhuna J. Sci. 2016, 7, 64. [Google Scholar] [CrossRef][Green Version]
- Jaiswal, A.K.; Abu-Ghannam, N. Kinetic studies for the preparation of probiotic cabbage juice: Impact on phytochemicals and bioactivity. Ind. Crop. Prod. 2013, 50, 212–218. [Google Scholar] [CrossRef]
- Yoon, K.Y.; Woodams, E.E.; Hang, Y.D. Production of probiotic cabbage juice by lactic acid bacteria. Bioresour. Technol. 2006, 97, 1427–1430. [Google Scholar] [CrossRef]
- Sarvan, I.; Valerio, F.; Lonigro, S.L.; de Candia, S.; Verkerk, R.; Dekker, M.; Lavermicocca, P. Glucosinolate content of blanched cabbage (Brassica oleracea var. capitata) fermented by the probiotic strain Lactobacillus paracasei LMG-P22043. Food Res. Int. 2013, 54, 706–710. [Google Scholar] [CrossRef]
- Cho, Y.H.; Hong, S.M.; Kim, C.H. Isolation and characterization of lactic acid bacteria from Kimchi, Korean traditional fermented food to apply into fermented dairy products. Korean J. Food Sci. Anim. Resour. 2013, 33, 75–82. [Google Scholar] [CrossRef]
- Kawahara, T.; Otani, H. Stimulatory effect of lactic acid bacteria from commercially available Nozawana-zuke pickle on cytokine expression by mouse spleen cells. Biosci. Biotechnol. Biochem. 2006, 70, 411–417. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tomita, S.; Watanabe, J.; Kuribayashi, T.; Tanaka, S.; Kawahara, T. Metabolomic evaluation of different starter culture effects on water-soluble and volatile compound profiles in nozawana pickle fermentation. Food Chem. Mol. Sci. 2021, 2, 100019. [Google Scholar] [CrossRef]
- Demirci, T.; Sert, D.; Aktaş, K.; Atik, D.S.; Öztürk Negiş, H.İ.; Akın, N. Influence of hot and cold break tomato powders on survival of probiotic L. paracasei subsp. paracasei F19, texture profile and antioxidative activity in set-type yoghurts. LWT 2020, 118. [Google Scholar] [CrossRef]
- García-Hernández, J.; Hernández-Pérez, M.; Peinado, I.; Andrés, A.; Heredia, A. Tomato-antioxidants enhance viability of L. reuteri under gastrointestinal conditions while the probiotic negatively affects bioaccessibility of lycopene and phenols. J. Funct. Foods 2018, 43, 1–7. [Google Scholar] [CrossRef]
- George-Okafor, U.; Ozoani, U.; Tasie, F.; Mba-Omeje, K. The efficacy of cell-free supernatants from Lactobacillus plantarum Cs and Lactobacillus acidophilus ATCC 314 for the preservation of home-processed tomato-paste. Sci. Afr. 2020, 8. [Google Scholar] [CrossRef]
- Yoon, K.Y.; Woodams, E.E.; Hang, Y.D. Probiotication of tomato juice by lactic acid bacteria. J. Microbiol. 2004, 42, 315–318. [Google Scholar]
- King, V.A.E.; Huang, H.Y.; Tsen, J.H. Fermentation of tomato juice by cell immobilized Lactobacillus acidophilus. Mid-Taiwan J. Med. 2007, 12, 1–7. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Kurvinen, T.; Rissanen, P. Use of a probiotic Bifidobacterium in a dry food matrix, an in vivo study. Int. J. Food Microbiol. 2004, 95, 103–106. [Google Scholar] [CrossRef]
- Casarotti, S.N.; Borgonovi, T.F.; Batista, C.L.F.M.; Penna, A.L.B. Guava, orange and passion fruit by-products: Characterization and its impacts on kinetics of acidification and properties of probiotic fermented products. LWT 2018, 98, 69–76. [Google Scholar] [CrossRef]
- Helland, M.H.; Wicklund, T.; Narvhus, J.A. Growth and metabolism of selected strains of probiotic bacteria in milk- and water-based cereal puddings. Int. Dairy J. 2004, 14, 957–965. [Google Scholar] [CrossRef]
- Heenan, C.N.; Adams, M.C.; Hosken, R.W.; Fleet, G.H. Survival and sensory acceptability of probiotic microorganisms in a nonfermented frozen vegetarian dessert. LWT Food Sci. Technol. 2004, 37, 461–466. [Google Scholar] [CrossRef]
- Shimakawa, Y.; Matsubara, S.; Yuki, N.; Ikeda, M.; Ishikawa, F. Evaluation of Bifidobacterium breve strain Yakult-fermented soymilk as a probiotic food. Int. J. Food Microbiol. 2002, 81, 131–136. [Google Scholar] [CrossRef]
- Pimentel, T.C.; Madrona, G.S.; Garcia, S.; Prudencio, S.H. Probiotic viability, physicochemical characteristics and acceptability during refrigerated storage of clarified apple juice supplemented with Lactobacillus paracasei ssp. paracasei and oligofructose in different package type. LWT Food Sci. Technol. 2015, 63, 415–422. [Google Scholar] [CrossRef]
- Emser, K.; Barbosa, J.; Teixeira, P.; Bernardo de Morais, A.M.M. Lactobacillus plantarum survival during the osmotic dehydration and storage of probiotic cut apple. J. Funct. Foods 2017, 38, 519–528. [Google Scholar] [CrossRef]
- Cui, L.; Niu, L.-Y.; Li, D.-J.; Liu, C.-Q.; Liu, Y.-P.; Song, J.-F. Effects of different drying methods on quality, bacterial viability and storage stability of probiotic enriched apple snacks. J. Integr. Agric. 2018, 17, 247–255. [Google Scholar] [CrossRef]
- Amorim, J.C.; Piccoli, R.H.; Duarte, W.F. Probiotic potential of yeasts isolated from pineapple and their use in the elaboration of potentially functional fermented beverages. Food Res. Int. 2018, 107, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.G.M.; Fonteles, T.V.; De Jesus, A.L.T.; Rodrigues, S. Sonicated pineapple juice as substrate for L. casei cultivation for probiotic beverage development: Process optimisation and product stability. Food Chem. 2013, 139, 261–266. [Google Scholar] [CrossRef]
- de Almeida Bianchini Campos, R.C.; Martins, E.M.F.; de Andrade Pires, B.; do Carmo Gouveia Peluzio, M.; da Rocha Campos, A.N.; Ramos, A.M.; de Castro Leite Júnior, B.R.; de Oliveira Martins, A.D.; da Silva, R.R.; Martins, M.L. In vitro and in vivo resistance of Lactobacillus rhamnosus GG carried by a mixed pineapple (Ananas comosus L. Merril) and jussara (Euterpe edulis Martius) juice to the gastrointestinal tract. Food Res. Int. 2019, 116, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.; Borges, S.; Amorim, M.; Pereira, M.J.; Oliveira, A.; Pintado, M.E.; Teixeira, P. Comparison of spray drying, freeze drying and convective hot air drying for the production of a probiotic orange powder. J. Funct. Foods 2015, 17, 340–351. [Google Scholar] [CrossRef]
- da Costa, G.M.; de Carvalho Silva, J.V.; Mingotti, J.D.; Barão, C.E.; Klososki, S.J.; Pimentel, T.C. Effect of ascorbic acid or oligofructose supplementation on L. paracasei viability, physicochemical characteristics and acceptance of probiotic orange juice. LWT Food Sci. Technol. 2017, 75, 195–201. [Google Scholar] [CrossRef]
- Sengun, I.Y.; Kirmizigul, A.; Atlama, K.; Yilmaz, B. The viability of Lactobacillus rhamnosus in orange juice fortified with nettle (Urtica dioica L.) and bioactive properties of the juice during storage. LWT 2020, 118. [Google Scholar] [CrossRef]
- Cristiny de Oliveira Vieira, K.; Da Silva Ferreira, C.; Toso Bueno, E.B.; De Moraes, Y.A.; Campagnolo Gonçalves Toledo, A.C.; Nakagaki, W.R.; Pereira, V.C.; Winkelstroter, L.K. Development and viability of probiotic orange juice supplemented by Pediococcus acidilactici CE51. LWT 2020, 130. [Google Scholar] [CrossRef]
- Pereira, A.L.F.; Maciel, T.C.; Rodrigues, S. Probiotic beverage from cashew apple juice fermented with Lactobacillus casei. Food Res. Int. 2011, 44, 1276–1283. [Google Scholar] [CrossRef]
- Ribeiro, A.P.D.O.; Gomes, F.D.S.; dos Santos, K.M.O.; da Matta, V.M.; Sá, D.D.G.C.F.D.; Santiago, M.C.P.D.A.; Conte, C.; Costa, S.D.D.O.; Ribeiro, L.D.O.; Godoy, R.L.D.O.; et al. Development of a probiotic non-fermented blend beverage with juçara fruit: Effect of the matrix on probiotic viability and survival to the gastrointestinal tract. LWT 2020, 118, 108756. [Google Scholar] [CrossRef]
- Vivek, K.; Mishra, S.; Pradhan, R.C. Characterization of spray dried probiotic Sohiong fruit powder with Lactobacillus plantarum. LWT 2020, 117. [Google Scholar] [CrossRef]
- Fonteles, T.V.; Costa, M.G.M.; de Jesus, A.L.T.; Rodrigues, S. Optimization of the Fermentation of Cantaloupe Juice by Lactobacillus casei NRRL B-442. Food Bioprocess Technol. 2012, 5, 2819–2826. [Google Scholar] [CrossRef]
- Saw, L.K.; Chen, S.; Wong, S.H.; Tan, S.A.; Goh, L.K. Fermentation of tropical fruit juices by lactic acid bacteria. In Proceedings of the 12th Asean Food Conference, Bangkok, Thailand, 16–18 June 2011. [Google Scholar]
- Mousavi, Z.E.; Mousavi, S.M.; Razavi, S.H.; Emam-Djomeh, Z.; Kiani, H. Fermentation of pomegranate juice by probiotic lactic acid bacteria. World J. Microbiol. Biotechnol. 2011, 27, 123–128. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Valerio, F.; Lonigro, S.L.; De Angelis, M.; Morelli, L.; Callegari, M.L.; Rizzello, C.G.; Visconti, A. Study of adhesion and survival of lactobacilli and bifidobacteria on table olives with the aim of formulating a new probiotic food. Appl. Environ. Microbiol. 2005, 71, 4233–4240. [Google Scholar] [CrossRef]
- Han, J.; Hang, F.; Guo, B.; Liu, Z.; You, C.; Wu, Z. Dextran synthesized by Leuconostoc mesenteroides BD1710 in tomato juice supplemented with sucrose. Carbohydr. Polym. 2014, 112, 556–562. [Google Scholar] [CrossRef]
- Rodrigues, S.; Silva, L.C.A.; Mulet, A.; Cárcel, J.A.; Fernandes, F.A.N. Development of dried probiotic apple cubes incorporated with Lactobacillus casei NRRL B-442. J. Funct. Foods 2018, 41, 48–54. [Google Scholar] [CrossRef]
- Akman, P.K.; Uysal, E.; Ozkaya, G.U.; Tornuk, F.; Durak, M.Z. Development of probiotic carrier dried apples for consumption as snack food with the impregnation of Lactobacillus paracasei. LWT 2019, 103, 60–68. [Google Scholar] [CrossRef]
- Haffner, F.B.; Pasc, A. Freeze-dried alginate-silica microparticles as carriers of probiotic bacteria in apple juice and beer. LWT 2018, 91, 175–179. [Google Scholar] [CrossRef]
- Xu, X.; Bao, Y.; Wu, B.; Lao, F.; Hu, X.; Wu, J. Chemical analysis and flavor properties of blended orange, carrot, apple and Chinese jujube juice fermented by selenium-enriched probiotics. Food Chem. 2019, 289, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Fernández-Quirós, B.; Ludwig, I.A.; Pradas, I.; Crozier, A.; Moreno-Rojas, J.M. Catabolism of citrus flavanones by the probiotics Bifidobacterium longum and Lactobacillus rhamnosus. Eur. J. Nutr. 2018, 57, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Rasika, D.; Vidanarachchi, J.; Rocha, R.; Balthazar, C.; Cruz, A.; Sant’Ana, A.S.; Ranadheera, C. Plant-based milk substitutes as emerging probiotic carriers. Curr. Opin. Food Sci. 2021. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasika, D.M.D.; Vidanarachchi, J.K.; Luiz, S.F.; Azeredo, D.R.P.; Cruz, A.G.; Ranadheera, C.S. Probiotic Delivery through Non-Dairy Plant-Based Food Matrices. Agriculture 2021, 11, 599. https://doi.org/10.3390/agriculture11070599
Rasika DMD, Vidanarachchi JK, Luiz SF, Azeredo DRP, Cruz AG, Ranadheera CS. Probiotic Delivery through Non-Dairy Plant-Based Food Matrices. Agriculture. 2021; 11(7):599. https://doi.org/10.3390/agriculture11070599
Chicago/Turabian StyleRasika, D. M. D., Janak K. Vidanarachchi, Selma F. Luiz, Denise Rosane Perdomo Azeredo, Adriano G. Cruz, and Chaminda Senaka Ranadheera. 2021. "Probiotic Delivery through Non-Dairy Plant-Based Food Matrices" Agriculture 11, no. 7: 599. https://doi.org/10.3390/agriculture11070599
APA StyleRasika, D. M. D., Vidanarachchi, J. K., Luiz, S. F., Azeredo, D. R. P., Cruz, A. G., & Ranadheera, C. S. (2021). Probiotic Delivery through Non-Dairy Plant-Based Food Matrices. Agriculture, 11(7), 599. https://doi.org/10.3390/agriculture11070599